

Abstract
Tight junctions (TJs) are structures between cells where cells appear in the closest possible contact. They are responsible for sealing compartments when epithelial sheets are generated. They regulate the permeability of ions, (macro) molecules and cells via the paracellular pathway. Their structure at the electron microscopic level has been well known since the 1970s; however, only recently has their macromolecular composition been revealed. This review first examines the major macromolecular components of the TJs (occludin, claudins, junctional adhesion molecule and tricellulin) and then the associated macromolecules at the intracellular plaque [zonula occludens (ZO)-1, ZO-2, ZO-3, AF-6, cingulin, 7H6]. Emphasis is given to their interactions in order to begin to understand the mode of assembly of TJs. The functional significance of TJs is detailed and several mechanisms and factors involved are discussed briefly. Emphasis is given to the role of intestinal TJs and the alterations observed or speculated in diverse disease states. Specifically, intestinal TJs may exert a pathogenetic role in intestinal (inflammatory bowel disease, celiac disease) and extraintestinal diseases (diabetes type 1, food allergies, autoimmune diseases). Additionally, intestinal TJs may be secondarily disrupted during the course of diverse diseases, subsequently allowing the bacterial translocation phenomenon and promoting the systemic inflammatory response, which is often associated with clinical deterioration. The major questions in the field are highlighted.
Keywords: Tight junctions, Occludin, Claudins, Junctional adhesion molecule, Tricellulin, Intestinal permeability
INTRODUCTION
The direct observation of how close cells can be came with the use of the transmission electron microscope. Cell biologists of the 1950s and 1960s observed several structures that seemed to bring the cells into close contact and connect with each other, and gave to these junctions several names, descriptive of either the structure or the function served by the structure. Thus, the names of occluding junctions [or tight junctions (TJs)], anchoring junctions (or adherens junctions) and communicating junctions (or gap junctions) were coined.
TJs are the key elements for creating two different barriers: the first barrier is between the apical and basolateral cell membrane (lipid bilayer) compartment, thus keeping the protein and lipid composition of these two membrane domains qualitatively different by restricting their exchange. The second barrier is between the apical and basal compartment defined by the epithelium in which the TJ is present, thus restricting the passage of water, solutes and cells from the “outer” to the “inner” compartment and vice versa, also known as paracellular pathway.
TJs were seen in the most apical part of the lateral cell membrane of polarized cells, forming continuous circumferential contacts. What morphologists observed with the use of transmission electron microscopy was an obliteration of the extracellular space between cells at certain points; the morphological data led them to suggest that there are points of the closest possible apposition between the cell membranes of cells involved. The use of freeze-fracture techniques added important new information by showing these “contact points’ as fibrillar rows of intramembrane particles, forming a branching network around the cell. The chemical composition of these particles became a subject of intense investigation. It was originally proposed that these particles are composed solely of lipids[1]. Starting from the early 1990s, proteins localized at and involved in the formation of TJs have been identified. The first member of this family, occludin, is described in the report by Furuse et al[2] in 1993 and since then the field has witnessed a wealth of new information, which will be briefly reviewed in this report.
MACROMOLECULAR COMPOSITION OF TJs
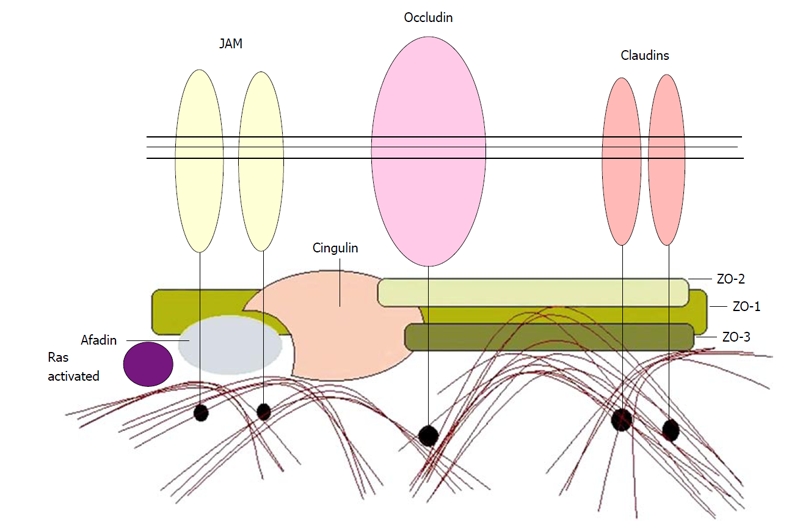
To date, four groups of macromolecules are considered as bona fide integral components of the TJ: occludin(s), claudins, junctional adhesion molecule (JAM) and tricellulin. It is remarkable that the members of the first two groups cross the cell membrane four times, thus creating two extracellular loops and three intracellular domains, including the amino terminal domain, the carboxy terminal domain and the very short loop-connecting segment. In contrast, JAM crosses the cell membrane only once (Figures 1-3). Recently, tricellulin was identified as another integral TJ protein, quite homologous to occludin, with 4 transmembrane domains, preferentially localized at the tight junctional strands of tricellular contacts of epithelial cells.
Figure 1.

Occludin. WW sequences are highly preserved 35-45 aminoacid segments, containing Trp and Pro. They recognize proline rich domains. Ser 379 and Thr 375 are casein kinase 2 (CK2) phosphorylation sites. PKC: Protein kinase C; PI3K: Phosphatidylinositol 3 kinase; PP2A: Protein phosphatase 2A; ZO: Zonula occludens.
Figure 3.

Junctional adhesion molecule.
Occludin
The name comes from the Latin verb “occludere”, which means to restrict passage. It was first isolated by Furuse et al[2] in 1993 in cultures of chicken liver cells, based on the results of intensive screening of monoclonal antibodies. It is a component of TJs, highly conserved among species and without tissue specific isoforms (recently only one occludin isoforms resulting from alternative splicing has been reported)[3]. Occludin is only expressed under normal conditions in cells that form TJs. When cells that do not form TJs are led via transfection to express occludin, it is concentrated at cell contact sites, forming only a small number of short strands identified in freeze-fracture replicas[4]. However, when occludin was cotransfected with claudin-1, tight junctional fibrils were formed[4]. In addition, immunoreplica electron microscopy has documented the presence of occludin in tight junctional strands[5].
Occludin (Figure 1) consists of 504 amino acids. The amino terminal portion is intracellular and contains 57 amino acids. This portion contains a WW domain (PPYP), which is motif participating in the interaction with other signaling and regulatory proteins[6]. The four transmembrane domains consist of 21-24 amino acid residues, mostly hydrophobic. The two extracellular loops are 43 amino acids long each and the intravellular loop is 10 amino acids long. Both extracellular loops are rich in tyrosine and glycine (in the range of 20%-30%); however, the functional significance of this is not known yet[2]. The very long intracellular carboxyl terminal part (50% of the sequence) has several sites of potential interaction with other macromolecules and phosphorylation sites[2,7]. It is a highly hydrophilic sequence, containing stretches of charged amino acids, like EEEEE (aa 347-351) and RRGRRRRR (aa 363-370).
The electrophoretic mobility of occludin in polyacrylamide gels has revealed a wide range of bands from 62kDa to 82 kDa. This difference in mobility has been assigned to post-translational modifications and more precisely to differential phosphorylation on both tyrosine and serine/threonine residues. It has been proposed that the site and the degree of phosphorylation of occludin are important parameters in defining the localization and the function of the molecule. For example, highly phosphorylated occludin, especially in serine/threonine residues, is detected mainly in the junctional area and correlates with restricted transcellular permeability. On the contrary, non-phosphorylated occludin is detected intracellularly, in the vesicular compartment, and is considered to contribute less to transcellular permeability, being a pool of macromolecules easily subjected to regulatory stimuli. As mentioned above, the site of phosphorylation is also important. It has been suggested that high degree of phosphorylation at tyrosine residues correlates with loss of function and higher transcellular permeability.
Recently, a novel alternatively spliced form of occludin has been described[3], in which 56 amino acids are present close to the amino terminal of the molecule. This new form termed occludin 1B has been detected in TJs and widely co-expressed with the known occludin.
Claudins
The name claudin comes from the Latin verb “claudere”, which means “to close”. Claudins, identified after occludin, are transmembrane proteins extremely crucial for TJ formation. The existence of another TJ component, in addition to occludin, was suspected when Saitou et al[8] created an occludin deficient cell line that was surprisingly able to form TJ strands. These TJs appeared structurally normal, but this finding should not underestimate the role of occludin since a later studied model of occludin deficient mice[9] presented abnormalities from several tissues e.g., chronic inflammation and hyperplasia of the gastric epithelium, calcification in the brain, testicular atrophy, thinning of the compact bone and loss of cytoplasmic granules in striated duct cells of the salivary glands.
Furuse et al[10] identified the first two members of the claudin superfamily, claudins 1 and 2. Comparing their sequence to already known proteins, they found similarity to at least three membrane-associated proteins: rat ventral prostate-1, clostridium perfringens enterotoxin receptor and oligodendrocyte specific protein[11]. Up until now, the claudin family consists of 24 members in humans[12,13] (Table 1).
Table 1.
Distinctive characteristics of claudins
| Claudin | Distinctive characteristics |
| 1 | Present in high resistance epithelia (collecting segment), absent in leaky epithelia (proximal tubule) |
| Crucial for the mammalian epidermal barrier | |
| Mutations cause neonatal sclerosing cholangitis | |
| Prognostic value in colon and thyroid cancer | |
| 2 | Present in leaky epithelia (proximal tubule) and absent in tight epithelia |
| Present in the choroids plexus epithelium | |
| 3 | Present in the tighter segments of the nephron |
| Up-regulated in ovarian, breast, prostate and pancreatic tumors | |
| 4 | Induces selective decrease in sodium permeability |
| Present in the tighter segments of the nephron | |
| Alternative name: CPE-R | |
| Up-regulated in ovarian, breast, prostate and pancreatic tumors | |
| 5 | Frequently deleted in velo cardio facial syndrome |
| Constitutes TJ strands in endothelial cells and it is transiently expressed during the development of retinal pigment epithelium | |
| 6 | Present in embryonic epithelia |
| Its overexpression in transgenic mice generates a defective epidermal permeability barrier | |
| 7 | Down regulated in head and neck squamous cell carcinomas |
| Upregulated in stomach cancer | |
| 8 | Present in the tighter segments of the nephron |
| 10 | Prognostic value in hepatocellular carcinoma (recurrence) |
| 11 | Present in oligodendrocytes and sertoli cells; also named OSP |
| 14 | Expressed in the sensory epithelium of the organ of Corti Mutations cause autosomal recessive deafness |
| 15 | Present in endothelial cells |
| 16 | Critical for Mg2+ and Ca2+ resorption in the human thick ascending limb of Henle |
| Mutations cause familial hypomagnesaemia | |
| 18 | Expressed in the lung and stomach |
Claudins 9, 12, 13, 17, 19-24: Insufficient data. CPE-R: Clostridium perfringens enterotoxin receptor; OSP: Oligodendrocyte sertoli protein.
Claudins are transmembrane proteins with MW ranging from 20 kDa to 27 kDa. They possess an intracellular amino-terminus, four transmembrane segments that form two extracellular and an intracellular loop, and a short carboxyterminal intramembrane tail (Figure 2)[14]. The first and fourth transmembrane part, as well as the first and second extracellular loops are highly conserved, whereas the COOH tail has the greatest variability among members of the claudin family[11]. The first extracellular loop is longer and more hydrophobic than the second one[11,14]; therefore it is possibly responsible for cell-to-cell contact. The second extracellular loop contains many charged residues, which regulate the affinity to ions[14] and possibly forms ion-selective paracellular channels. The carboxyterminal tail contains many phosphorylation sites, related to protein kinase C (PKC), casein kinase 2 (CK2) and a cAMP dependent kinase. Almost every claudin has a carboxyterminal Tyr-Val sequence, which acts as a PDZ-domain interacting motif. Exceptions are claudins 16, 11, 12 and 13. When claudins 1 and 2 were transfected in TJs, negative cells[4] induced cell-to-cell adhesion by forming TJ strands with no obvious structural and functional difference from their occludin positive homologues. At present, claudins are supposed to be the “backbone” of the TJ barrier[14-16].
Figure 2.

Claudin(s). PKC: Protein kinase C; CK2: Casein kinase 2; ZO: Zonula occludens.
TJs in vivo usually express 2 claudins[15] with two notable exceptions, the oligodendrocytes and the Sertoli cells, both expressing a single claudin, claudin-11[11]. Claudins are also characterized by a differential tissue expression pattern[15,17] and a developmental selectivity. Interestingly, nephron expresses different claudins in different segments and this variability may be the basis for differential epithelial permeability in these segments[18-20]: claudins 5 and 15 are expressed at endothelial cells, 2, 10 and 11 at the proximal tubule segment, 1, 3 and 8 at the distal tubule and 1, 3, 4 and 8 at the collecting duct segment. As proved by Turksen et al[21,22] and Reyes et al[18], the expression of claudins in murine nephron changes during development. Hellani et al[23], who studied the expression of claudin 11 on Sertoli cells, also verified the above observations.
The complexity at the expression of different claudins in different tissues, in different stages of their development and to more than one combination, suggests that these molecules participate in both homophilic and heterophilic interactions within the same cell or between opposing cell membranes[24]. The possible combination models that claudins follow to their polymerization are presented in Figure 2. At the level of electron microscopy, claudins form different strand patterns, related to the P face or the E face of the cytoplasmic membrane. Claudins 1, 3 and 11 form P face strands, whereas claudins 2 and 5 are related to the E face strand pattern[15,24-26].
Claudins form a transmembrane and transcellular net that serves paracellular permeability, epithelial polarization and conservation of the transepithelial resistance (TER)[11,14,16], as well as selective permeation of charged molecules and ions[27,28]. These parameters vary among different tissues according to their claudin expression, as long as claudins are the only known variable elements in TJs[11].
Finally, some members of the claudin family are receptors for extracellular ligands, such as Clostridium perfringens enterotoxin, that bind directly to claudins-4 (high affinity) and -3 (low affinity)[29-31].
JAM
JAM was first described in 1998 by Martìn-Padura et al[32]. It is a type I transmembrane protein of MW 43 kDa, belongs to the immunoglobulin superfamily (Figure 3) and is localized in close proximity to the TJs of epithelial and endothelial sheets.
When JAM is expressed in cells that do not form TJs under physiological conditions, tight junctional strands are not observed, as is the case with occludin and claudins[33]. In this case, JAM molecules are accumulated only when cell-cell contact is established and both cells express JAM[32], suggesting a specific polymerization pattern by homophilic interactions. It appears that JAM plays a role in the process of formation of TJs, since antibodies against JAM do not disrupt already formed TJs[34].
Structurally, JAM consists of an extracellular amino terminal segment, a transmembrane domain and an intracellular carboxy terminal segment[32]. The extracellular part consists of 215 amino acids and contains two variable (V) type IgG domains. The V-V arrangement is novel between Ig domains and differentiates JAM-subfamily from other subfamilies of the Ig superfamily[32]. The intracellular part consists of 45 amino acids, where motifs appropriate for interactions with occludin[32] as well as other TJ associated macromolecules exist. The recently revealed X-ray structure of JAM suggests that first a U-shaped homodimer is formed and several homodimers interact and form an extensive network[35].
JAMs are subdivided based on the expression of type I or II PDZ-binding motifs in the intracellular C-terminus, which suggests that the 2 types interact with unique scaffolding and cytoplasmic proteins. JAM-A, JAM-B, and JAM-C (or JAM-1, JAM-2, and JAM-3) have type II binding motifs, whereas the atypical JAMs, including JAM-4, Coxsackie and adenovirus receptor and endothelial selective adhesion molecule, contain type I PDZ-binding domains[36].
It has been proposed that JAM may contribute to free diffusion of proteins within the lipid bilayer but more importantly to the restriction of cellular passage (leukocytes, monocytes, lymphocytes, etc.) through TJs between endothelial cells[14,32,35,37].
Tricellulin
A recent addition to the list of TJ proteins is tricellulin. Structurally it is quite homologous to occludin, with 555 amino acids forming 4 transmembrane domains; both the amino and the carboxy termini are intracellular, forming two extracellular loops. It is localized at the tight junctional strands of tricellular contacts of epithelial cells[38]. It participates in epithelial barrier organization. It was recently found that recessive mutations in the tricellulin gene cause nonsyndromic deafness and that tricellulin participates in the junctions in cochlear and vestibular epithelial cells[39]. Recent studies indicate that tricellulin is also localized in special TJs of myelinating Schwann cells[40]. Tricellulin has also been proposed to play an inhibitory role in the process of epithelial to mesenchymal transition of gastric carcinoma cells[41].
MACROMOLECULAR COMPONENTS ASSOCIATED WITH TJs
Several macromolecules are concentrated underneath the TJs and constitute the “tight junctional plaque”. In general, their function is to promote interactions between them and the bona fide TJ macromolecules described above. These interactions can be mediated through: (1) PDZ motifs, that are present in several copies in most of the proteins of the tight junctional plaque; these motifs are usually 80-90 amino acids long and associate with certain protein motifs (e.g., S/TXV). Their presence in high numbers allows multiple and complicated interactions, leading to clustering and anchoring of transmembrane proteins[42]; (2) SH3 motifs, usually 50-70 amino acids long and frequently used as docking sites for several intracellular proteins; and (3) guanylate kinase (GK) motifs that are considered to be enzymatically inactive but could activate signaling pathways[43]. The proteins that contain them are collectively termed MAGUK proteins (Membrane-associated GK proteins). These proteins are beyond the scope of the present review so they will only be briefly mentioned, with emphasis on their interactions.
Zonula occludens-1
Zonula occludens-1 (ZO-1) is a 210-225 kDa phosphoprotein that interacts with occludin[44], claudins[45] and JAM[46] and also ZO-2[47], ZO-3[47], AF-6[46], cingulin[48] and the actin cytoskeleton[44]. Therefore, it plays a key role in bringing several components together and in connecting tight junctional proteins to the cytoskeleton. Cells that do not form TJs contain ZO-1 either dispersed in the cytoplasm or concentrated in cadherin-enriched adherens junctions[49] (Table 2).
Table 2.
Interaction between zonula occludens-1 and other macromolecules
| ZO-1 protein domain | Interacting molecules |
| 1st PDZ | C terminus of claudin |
| 2nd/3rd PDZ | JAM |
| GUK | Occludin |
| CAR | |
| 2nd PDZ | ZO-2s and ZO-3s 2nd PDZ |
| Actin cytoskeleton | |
| Actin binding protein 4.1 | |
| AF-6 | |
| Cingulin | |
| A catenin | |
| Connexins 43 and 45 |
ZO: Zonula occludens; CAR: Coxsackievirus and adenovirus receptor; JAM: Junction adhesion molecule.
ZO-2
ZO-2 is a 160 kDa protein present in two different isoforms. Table 3 shows the relationships of ZO-2 to other TJ molecules and cytosceleton fibers[14]. ZO-2 reacts with the splicing factor SC35[50] and with the transcription factors Fos, Jun and C/EBP[51].
Table 3.
Interaction between zonula occludens-2 and other macromolecules
| ZO-2 binding area | Interacting molecule |
| 1st PDZ | Claudin |
| 2nd PDZ | ZO-1 |
| GUK | Occludin |
| Cingulin | |
| C terminal proline rich domain | Actin |
| Actin binding protein 4.1 |
ZO: Zonula occludens.
ZO-3
ZO-3 is a 130 kDa that differs from the other two members of ZO group in that it lacks the long carboxy-tail (Table 4). According to the model of Wittchen et al[47], ZO-1 binds to ZO-2 and ZO-3; however, there is no direct interaction between ZO-2 and ZO-3 (Figure 4).
Table 4.
Interaction between zonula occludens-3 and other macromolecules
| ZO-3 binding sites | Interacting molecules | |
| 1st PDZ domain | Claudins | |
| 2nd PDZ domain | ZO-1 | |
| N terminus | Actin | Occludin |
| C terminus (class I PDZ binding motif TDL) | 6th PDZ domain PATJ | Cingulin |
| Connexin 45 |
ZO: Zonula occludens.
Figure 4.

Zonula occludens. JAM: Junctional adhesion molecule; ZO: Zonula occludens.
AF-6/Afadin
AF-6/Afadin is a 205 kDa protein that seems to be more important for TJ formation and development than for TJ stabilization[14]. It is expressed in both TJ and AJ, and it seems to correlate with: (1) ZO-1 and activated Ras protein have antagonistic effect for the same binding site[52]; (2) JAM; and (3) F actin. AF-6 also interacts with cingulin[48]. Afadin expresses a 190 kDa splicing variant[53], found at the postsynaptic densities of neuronal tissues.
Cingulin
Cingulin is named after the Latin word “cingere”, to encircle. It is a 140-160 kDa protein. It is characterized by a globular N terminal head, a central α-helical coiled-coil domain and a C terminal tail. Through its central rods, dimmers are formed that interact with ZO-1, ZO-2, ZO-3, JAM, F actin and myosin. The majority of these interactions are supported by its globular head[14]. Cingulin is a serine phosphorylated protein, independent from PKC[54].
7H6 antigen
One hundred and fifty five kilodaltons protein that belongs to a family of proteins with alpha helical coiled coil domain and an ATPase domain. It sustains phosphorylation and its phosphorylated form is detected at TJ areas, whereas the non-phosphorylated form dissociates form TJs[14].
ASSEMBLY OF TJs
During embryogenesis and formation of epithelial sheets, it appears that primordial “spot-junctions” are formed. These are found in protrusions of neighboring cells and consist of cadherin, nectin, AF-6 and ZO-1[55,56]. At a later stage, cadherin and nectin are removed from those spots destined to become TJs, while occludin and claudins are recruited[57]. AF-6 remains in TJs of endothelial sheets but is removed from mature epithelial sheets and JAM is detected only in its presence in the tight junctional area[46], strongly suggesting a co-operative role of JAM and AF-6 related to specific cellular transmigration.
The interactions between the macromolecules described above obviously play an important role in the assembly process. Although several facts of the assembly process are not known yet, one can highlight observations that are the first important clues we have for understanding this process. Phosphorylation and dephosphorylation of occludin is a crucial post-translational modification since it has been connected with assignment of occludin to different subcellular compartments. Kinases and phosphatases involved are expected to be major players in the assembly-disassembly process. The intracellular domain of occludin is key to these events, since it contains several motifs that allow binding to the other macromolecules (see above). The last 150 amino acids close to the carboxyl terminal are very highly conserved among species and it is believed that they contain the self-association domain, critical for fibril formation.
Freeze-fracture immunostaining data suggest that usually two different types of claudins are present in tight junctional strands. This finding suggests that specific areas belonging to the short intracellular domains of claudins are responsible for crucial interactions between claudins and between one claudin and one occludin molecule. Although the molecular basis for these interactions is not known yet, it is certain that they will prove of great importance in understanding how the differential distribution of claudins in tissues leads to the great variety of electrical resistance and permeability observed among epithelial and endothelial sheets.
Another important aspect for our understanding of TJ assembly and functionality is the precise nature of the associations between extracellular loops of occludin and claudins. The length of each loop, similar in occludin and very different in claudins and the specific amino acid composition, mentioned above, are likely to be major factors in the formation of the barrier (Figure 5).
Figure 5.
Assembly of tight junction proteins. JAM: Junctional adhesion molecule; ZO: Zonula occludens.
FUNCTIONAL SIGNIFICANCE OF TJs-REGULATORY MECHANISMS
TJs are crucial structures for the establishment and the stability of epithelial barriers. They mediate cell to cell adhesion, thus creating a mechanical and charged fence for selective permeability of macromolecules and ions. Ionic permeability is expressed through the parameter of TER. Macromolecular and ionic permeability seems to be quite independent, as they sustain controversial remodeling under the same conditions[58].
More than a simple paracellular fence, the transmembrane component of TJs blocks the circulation of proteins and lipids between the apical and basolateral membrane, mediating membrane polarization. Finally, the intracellular component triggers a variety of signaling pathways and communicates with the nucleus, acting as a “sensor” for extracellular events.
TJ transmembrane and intracellular proteins interact to cytoskeletal elements and undergo various types of phosphorylation. They also regulate protein expression, participating in vesicle trafficking. Pit and Rab proteins are the intracellular proteins that mediate such a function[14]. Finally, TJs interact with the actin cytoskeleton, and directly or indirectly are related to other membrane structures and macromolecules, like adherens junctions[14,16], gap junctions and proteoglycans[14].
The extracellular part of the TJs, consisting of the extracellular part of transmembrane proteins, acts as a multidynamic ligand for both homophilic and heterophilic interactions. It interacts with other cells participating with homodimers (occludin) and homopolymers (JAM) of heteropolymers (claudins). Additionally, free antigens from the extracellular space are ligands for extracellular TJ domains. Between these antigens are bacterial toxins, whose role in the pathogenetic mechanisms is discussed below.
TJs are in a dynamic balance, regulated both from intracellular and extracellular events. Intracellular events that may influence TJ stability are related to energy depletion and cAMP level changes. ATP depletion downregulates TJs[59], whereas cAMP induces increase of TER and reduced paracellular permeability[58]. Cell cycle also affects TJs morphology, according to development of differentiation of the cell. Cell cycle regulation on TJs can be both quantitative (enhanced or reduced expression of the same molecular substates) and qualitative (changes of molecular substrates, e.g., differential expression of claudins during development). Extracellular events with the potential for TJ regulation include the following five aspects. Direct interaction of TJ proteins to other cell proteins; a typical example is endothelial interaction to leukocyte membrane antigens that induces site-specific TJ dissociation[60]; direct interactions to external antigens: claudins 3 and 4 and occludin are both receptors for Clostridium perfringens enterotoxin[29,61,62]; indirect paracellular (cytokine) effects and hormonic stimulus: (1) proteases[63] - leukocytes are proposed to secrete proteases that destroy TJs by macromolecular cleavage; (2) interleukins[60,64,65] - interleukins can have enhancing or dissociative effect on TJs, depending on the type of interleukin acting and the tissue studied; (3) interferons[60,64,66] - like interleukins, interferons can have both up or downregulating effect on different epithelia; (4) Igs[67] - IgM and IgG interact with coxsackievirus and adenovirus receptor (CAR) at inflammation sites, which may play a role at neutrophil transmigration; (5) GFs[58,68,69] - growth factors generally act on epithelia by lysis of their barrier, thereby dissociation of TJs. Both fibroblast growth factor, hepatocyte growth factor/scattering factor and vascular endothelial growth factor decreased TER and increased macromolecular permeability when adjusted to epithelia. However, other growth factors may have opposite results; (6) LTs[70] - Leukotriene D4 was found to induce reorganization of actin network and consequently affect TJs. There is evidence that phosphatidylinositol 3 kinase (PI3K) is involved in the signaling pathway used by LTD4; and (7) Glucocorticoids[71] - although still not completely understood, one study shows that they may destroy epithelial barrier by a mechanism of serine/threonine phosphorylation; oxidative stress: increased oxidative stress is associated with paracellular barrier dysfunction[72,73]; calcium level imbalance: low calcium is related to TJ disturbance[72,74].
Each stimulus affects TJs using a regulatory mechanism. So far the molecular basis of TJ regulation is partially known and continuous investigation reveals its complexity. Phosphorylation is probably the most common, but not the only regulatory pathway, and affects serine/threonine or tyrosine residues, usually with opposite results. Thus, serine/threonine high phosphorylation level is detected when TJ integrity and morphological stability is maximal, whereas low phosphorylation level on the same residues causes TJ dissociation and cytoplasmic localization of occludin. In contrast, tyrosine phosphorylation is related to TJ dissociation without cytoplasmic occludin localization[72,75-78], and block of tyrosine phosphorylation after TJ disruption leads to inability to reassemble[59]. An increasing number of protein kinases are detected to associate directly or indirectly with TJ intracellular components, including protein phosphatase 2A, PKC and atypical PCKs, CK2, PI3K, cAMP dependent kinase, Tyrosine kinases, Mitogen-activated protein kinases and protein tyrosine phosphatases. Another posttranslational mechanism that may participate in TJ regulation is N Glycozylation, occurring on CAR[79]. Proteolytic cleavage of occludin is mediated by PMNs during their transmigration and leads to a 22 kDa molecule without barrier-forming potential[63]. Finally, there is evidence that TJ composition can be regulated by transcription and translation modulation. For instance, symplekin and GATA-4 have been found to regulate TJ components such as claudin-2 and ZO-2[80-82].
TJs AT THE INTESTINAL BARRIER - CLINICAL IMPLICATIONS
General considerations and interactions
TJs are expressed by the intestinal columnar epithelium. As the TJs of every epithelial tissue, there are occludin and special claudins expressed at intestinal TJs that mainly regulate the permeability of the epithelial layer under normal conditions. TJ strands are copolymers of heterogeneous claudin species and occludin, and heterogeneous claudin species constitute the backbone of TJ strands in situ[24].
Occludin associated with ZO-1 in a linear strand is detected among the intestinal epithelia and is strictly related to the differentiation status[83]. In highly differentiated adenocarcinomas, occludin and ZO-1 are normally expressed and form TJ structures. On the contrary, low differentiation carcinomas are characterized by low or absent occludin expression, whereas ZO-1, although normally expressed, appears to concentrate at the inner membrane area in a dotted pattern.
The intestinal epithelium is in continuous contact with the microbial ecosystem of the gut, with which it establishes a dynamic relationship. Based mainly on culturing techniques it, had initially been estimated that more than 500 bacterial species inhabit the human gut[84]. With the advancement of metagenomic technology, the magnitude of bacterial species diversity was raised to 15 000 to 36 000 species based on rRNA sequence analysis[85]. A recent release of the data from the Metagenomics of the Human Intestinal Tract project revealed a total of 3.3 million non-redundant microbial genes in human fecal specimens[86]. Intestinal epithelial cells and bacteria interact in a continuous so-called “cross-talk”, for mutual benefit. TJs participate in this interaction in a primary or a secondary way. They possess proteins that act as pathogen receptors and directly bind to the bacterial wall. Moreover, TJs can be secondarily affected by cytokine expression and by rearrangement of the actin cytoskeleton induced by other bacterial mechanisms, such as intimin-Tir mechanism of enteropathogenic Escherichia coli[87]. Some known interactions between bacterial strains and tight junctional macromolecules are described in (Table 5)[29,31,61,62,87-93]. Probiotics have a positive effect on TJ barrier, thus enhancing the epithelial resistance to pathogens, also reducing the paracellular permeability of antigens that may cause inflammation[94].
Table 5.
Interaction between bacterial strains and tight junctional molecules
| Vibrio cholerae[87,88] | It expresses zonula occludens toxin that reversibly increases paracellular permeability, triggering phospholipase C and protein kinase Ca dependent actin polymerization |
| This process is primary or secondary related to TJ disruption | |
| Shigella flexneri[89] | Secretes heat stable proteins that affect intestinal cells and lead to TJ disruption, even in the absence of living bacteria |
| Clostridium perfringens[29,31,61,62] | Its enterotoxin interacts with high affinity to claudin-4, therefore also known as CPE-R |
| Lower affinity receptors are claudin-3 and occludin. CPE is proposed to be a multifunctional toxin that first induces cell damage at the level of the cell membrane, and thereby relates to TJ proteins, causing structural and functional alterations[61] | |
| Michl et al[31] have studied the effect of CPE on pancreatic cell cancers that expressed claudin 4[31], and they suggest that targeting of claudin-4 expressing tumors with CPE can represent a promising treatment method | |
| Clostridium difficile[90] | This pathogenic microorganism, known etiologic factor of pseudomembranous colitis, secretes two toxins TcdA and TcdB that act through the Rho GTPase pathway to produce cell damage |
| Study for their effect on epithelial TJ structure assumed that they lead to actin rearrangement, actin-ZO1 dissociation and dissociation of TJ components with changes of their cytoplasmic localization[90] | |
| EPEC[91-93] | EPEC secretes through the type III secretion mechanism[87] the EspF protein, that is dose-dependently related to TER and epithelial barrier disruption and cytoplasmic localization of occludin[91] |
| These effects seem to relate primary with phosphorylation of 20 kDa myosin light chain and cytoskeletal contraction. Occludin appears dephosphorylated on serine/threonine residues[92] | |
| The pathogenic action of EPEC on the intestinal epithelium is reversed by Saccharomyces boulardii[93] |
CPE: Clostridium perfringens enterotoxin; CPE-R: Clostridium perfringens enterotoxin receptor; EPEC: Enteropathogenic Escherichia coli; TJ: Tight junction; ZO: Zonula occludens; GTPase: Guanosine triphosphatase.
Dendritic cells, antigen presenting cells that belong to the subepithelial immune system, mediate a novel mechanism of bacterial uptake[95]. DCs send dendrites that express TJ proteins (occludin, claudin 1, ZO1) through the gaps between epithelial cells. These dendrites disassociate transcellular interactions, creating new ones between their surface proteins and previous cells. In this way, they send their dendrites to the epithelial surface and sample bacteria, without disrupting the integrity of the epithelial barrier.
Clinical implications
Several lines of evidence suggest that TJs are involved in the pathogenesis of inflammatory and non-inflammatory intestinal diseases. TJ disruption leads to an inadequate epithelial barrier and, subsequently, to an incontrollable water and electrolyte loss. Secondly, intestinal TJ disruption is implicated in the pathogenesis of diverse extraintestinal autoimmune and inflammatory diseases. Thirdly, intestinal TJs may be secondarily disrupted in the course of several intestinal or extraintestinal diseases, leading to their further aggravation through promotion of systemic responses to endotoxin escape from the gut lumen.
Pathogenetic role in intestinal diseases: Changes at the level of TJs are related to the inflammatory bowel diseases (IBD), Crohn’s disease and ulcerative colitis. These are diseases that display a course of recurrent exacerbation and subsidence periods. At the peak of the IBDs, polymorphonuclear neutrophils transmigrate through the epithelial layer and induce inflammation at the intestinal surface. Transmigration is accompanied by an increase of the epithelial permeability to ions and macromolecules, as a result of downregulation of TJ. More specifically, occludin seems to be downregulated in a mechanism different from that of other TJ proteins[96]. The observation that, in clinically asymptomatic Crohn’s disease patients, increased intestinal epithelial permeability precedes clinical relapse by as much as 1 year, raised the assumption that permeability defect may be an early event in disease reactivation. Further evidence supporting that abnormal intestinal permeability occurs early in the pathogenesis of Crohn’s disease is provided by a study demonstrating intestinal barrier disruption, even in the non-inflamed parts of ileum from patients with Crohn’s disease[97]. The primary and potentially genetically determined association of intestinal TJs disruption with the evolution of IBD came from studies demonstrating increased gut permeability in otherwise healthy relatives of people with IBD[98]. However, the genetic factors implicated in these phenomena have not been revealed yet and the opposite opinion, that TJ disruption is a secondary event to the inflammatory process in IBD, is still under consideration[98,99]. Notably, disease exacerbations and the risk of developing IBD have been associated with emotional stress[100]. Experimental and clinical data provided evidence that psychological stress exerts injurious effects on intestinal TJs and increases gut permeability in IBD and irritable bowel syndrome[101,102]. Therefore, the pathogenetic role of intestinal TJs disruption in IBD beyond its potential genetic basis has an environmental stress-related component as well.
Celiac disease is an immune-mediated enteropathy triggered by an inappropriate T cell-mediated response to ingested gluten and its component gliadin. Clinical and experimental studies suggest that altered intestinal barrier function might play an inciting role in the development of celiac disease by allowing gliadin to cross the intestinal barrier and activate the immune system. Zonulin is the 47 kDa intestinal epithelial protein analogue of zonula occludens toxin (Zot) of Vibrio cholerae. Zonulin is normally expressed and secreted to the surface of intestinal and other epithelia (heart, brain). Zonulin and Zot have the same receptor on the cell surface and both trigger actin polymerization by PLC and PKCa pathways. Zonulin has been proposed to be the initial factor for the pathogenesis of celiac disease[87]. The pathogenetic role of intestinal TJs disruption in celiac disease is supported by studies demonstrating that increased intestinal permeability exists prior to disease onset, persists in asymptomatic patients who were on a gluten-free diet and is also present in a significant proportion of healthy first-degree relatives of patients with celiac disease who also have increased intestinal permeability[103-106].
Pathogenetic role in extraintestinal diseases: A combination of predisposing genetics, dysregulated intestinal barrier function and aberrant immune responses play an inciting role in type 1 diabetes. Increased gut permeability is believed to facilitate increased exposure to antigens that can trigger autoimmune destruction of the insulin-producing pancreatic beta cells[107]. Several lines of experimental and clinical data suggest that intestinal TJs disruption and increased gut permeability is an early event with a pathogenetic association with disease onset. Specifically, experimental studies with the Biobreeding diabetes-prone rat (BBDP), an inbred line in which autoimmune diabetes spontaneously develops when weaned onto a normal diet, showed that increased intestinal permeability, associated with decreased expression of the TJ protein claudin-1, precede the onset of insulitis and clinical diabetes[108]. Also, increased intestinal permeability was found in diabetic patients at various stages of disease progression and their relatives; however, prediabetic subjects had the greatest increase[109,110]. Zonulin has been proposed to be the initial factor for the pathogenesis of diabetes mellitus type 1[110].
Food allergies are expressed as adverse multisystemic immune-mediated reactions to ingested food proteins/antigens. Current pathogenetic aspects highlight the pivotal role of increased intestinal permeability, which permits increased dietary antigen transport across the intestinal barrier and exposure of dietary antigens to the mucosal immune system, leading to the development of the dietary antigen-specific response. In support of this hypothesis, intestinal permeability in infants with food allergies was significantly increased compared with that seen in healthy children[111]. In subjects with food allergies, intestinal permeability remained increased even after 6 mo of an allergen-free diet, indicating that the increased permeability probably preexists and is independent of food antigen stimulation[112]. Additional data supporting a role for increased intestinal permeability in the development of food antigen sensitization and food allergies are provided by recent clinical studies that demonstrate an association between increased intestinal permeability and the development of new-onset food allergies in patients after liver and heart transplantation under immunosuppressant therapy. These allergies were not related to the passive transfer of food antigen-specific IgE or lymphocytes from donors to previously nonallergic recipients, as were developed even in cases of non-allergic donors[113,114]. It is considered that immunosuppressive agents disrupt intestinal TJs, thus increasing permeability and facilitating presentation of food allergens to the immune system[115].
Beyond diabetes type 1, a similar mechanism has been proposed to contribute to the pathogenesis of diverse autoimmune diseases like atopic dermatitis, ankylosing spondylitis, Hashimoto’s thyroiditis and autoimmune hepatitis. The classical paradigm of autoimmune pathogenesis involving specific gene makeup and exposure to environmental triggers has been recently challenged by the addition of a third element, the loss of intestinal barrier function[116]. Disruption of intestinal TJs in genetically predisposed subjects might trigger a pathological immune response. According to this theory, once the autoimmune process is activated, it is not auto-perpetuating, but rather can be modulated or even reversed by preventing the continuous interplay between genes and environment, or in other words, by preventing loss of gut barrier function[116]. A zonulin - induced intestinal TJs disruption has been proposed as the responsible mechanism of barrier dysfunction allowing antigens to invade subepithelial and cause autoimmune reactions[87].
Secondarily affected in intestinal and extraintestinal diseases: Diverse intestinal and extraintestinal diseases during their course exert injurious effects on the integrity of intestinal TJs. Disruption of intestinal barrier function further subsequently aggravates through promotion of a systemic inflammatory response and the structural and functional integrity of the diseased and other organs, leading to clinical deterioration. Thus, although secondarily affected, dysfunction of the gut barrier exerts a pivotal role for the clinical outcome of diverse diseases. There are two main theories of how this may occur. The first one supports the view that after an initial insult, the intestinal barrier is compromised, thus allowing the passage of intestinal bacteria and endotoxins in mesenteric lymph nodes, portal circulation and normally sterile extraintestinal tissues (bacterial translocation)[117]. This process causes systemic infections and promotes a systemic inflammatory response both associated with distant organ failure and development of a septic state[118]. The second theory supports that, after the initial injurious insult and disruption of gut barrier integrity, bacteria and endotoxins crossing the mucosal barrier activate an intestinal inflammatory response, even when these translocating factors are trapped within the gut wall or intestinal lymph nodes and do not reach the systemic circulation[119]. Thus, the gut becomes a proinflammatory organ and gut-derived inflammatory factors, carried mainly in mesenteric lymph, induce a systemic inflammatory response and multiple organ failure[119-121]. In both theories, the intestinal TJs are further disrupted under the influence of systemic cytokinemia, further aggravating intestinal barrier function and leading to a vicious cycle.
The above described mechanisms may occur in diverse clinical states such as in intestinal ischemia[122], hemorrhagic shock[123,124], in critically ill patients[125], total parenteral nutrition[126], radiation enteritis[127], burns[128], ileus[129], acute pancreatitis[130], sepsis[131], cardiac bypass surgery[132], chemotherapy[133], obstructive jaundice[134,135], alcoholic liver disease[136], liver resections[137,138] and liver cirrhosis[139-141].
FUTURE DIRECTIONS
Protection against external stimuli and challenging factors is mainly offered by the skin with its several layers, containing cells at different levels of differentiation, in order to better serve this function. In an analogous way, the epithelium covering the gastro-intestinal tract protects against several “foreign factors” whether they may be chemicals, microorganisms or of a different nature. At the same time, the intestine has to serve an equally important opposite function; the selective permeability of needed nutrients from the intestinal lumen into the circulation and into the internal milieu in general. These essential life-sustaining opposite functions are performed in the intestine almost exclusively by a monolayer epithelium, the intestinal epithelium, and one of the key elements, if not the most important for this “compartmentalization” function, is the presence and the versatility of existing TJs.
The study of the structure of TJs has led to exciting new information and now over 40 macromolecular components are in the picture. However, we still lack detailed information about the mode of extracellular associations between occludin and claudins and the mechanism(s) by which they form a tight seal between cell membranes. The fact that several molecular combinations exist in different tissues certainly suggests that there is extremely fine tuning in determination of paracellular permeability, in a highly tissue-specific manner. Information is also missing regarding the details of assembly and disassembly of TJs and the exact role of each factor affecting the phenomena. Further understanding of these processes will allow us to design pharmacological interventions with impressive tissue specificity and will endow us with a better understanding of several pathogenetic mechanisms at the molecular level. Pharmacological modulation of intestinal permeability in a specific manner will enable us to treat and even prevent several intestinal and extraintestinal diseases, possibly overcoming the issue of severe side effects observed with current treatment modalities. Furthermore, understanding the specific differences in nucleotide/protein sequences of tight junctional macromolecules among individuals will allow the establishment of “personalized medicine” therapeutic modalities in the future.
Footnotes
Peer reviewers: Yan-Fang Guan, Dr., Easton Hospital, PA, 250 South 21st Street, Easton, MA 18042, United States; Jean-Francois Beaulieu, Professor, Department of Anatomy and Cell Biology, Faculty of Medicine and Health Sciences, Université de Sherbrooke, Sherbrooke, Qué J1H 5N4, Canada
S- Editor Wu X L- Editor Roemmele A E- Editor Zheng XM
References
- 1.Kachar B, Reese TS. Evidence for the lipidic nature of tight junction strands. Nature. 1982;296:464–466. doi: 10.1038/296464a0. [DOI] [PubMed] [Google Scholar]
- 2.Furuse M, Hirase T, Itoh M, Nagafuchi A, Yonemura S, Tsukita S, Tsukita S. Occludin: a novel integral membrane protein localizing at tight junctions. J Cell Biol. 1993;123:1777–1788. doi: 10.1083/jcb.123.6.1777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Muresan Z, Paul DL, Goodenough DA. Occludin 1B, a variant of the tight junction protein occludin. Mol Biol Cell. 2000;11:627–634. doi: 10.1091/mbc.11.2.627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Furuse M, Sasaki H, Fujimoto K, Tsukita S. A single gene product, claudin-1 or -2, reconstitutes tight junction strands and recruits occludin in fibroblasts. J Cell Biol. 1998;143:391–401. doi: 10.1083/jcb.143.2.391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Fujimoto K. Freeze-fracture replica electron microscopy combined with SDS digestion for cytochemical labeling of integral membrane proteins. Application to the immunogold labeling of intercellular junctional complexes. J Cell Sci. 1995;108(Pt 11):3443–3449. doi: 10.1242/jcs.108.11.3443. [DOI] [PubMed] [Google Scholar]
- 6.Traweger A, Fang D, Liu YC, Stelzhammer W, Krizbai IA, Fresser F, Bauer HC, Bauer H. The tight junction-specific protein occludin is a functional target of the E3 ubiquitin-protein ligase itch. J Biol Chem. 2002;277:10201–10208. doi: 10.1074/jbc.M111384200. [DOI] [PubMed] [Google Scholar]
- 7.Cordenonsi M, Turco F, D'atri F, Hammar E, Martinucci G, Meggio F, Citi S. Xenopus laevis occludin. Identification of in vitro phosphorylation sites by protein kinase CK2 and association with cingulin. Eur J Biochem. 1999;264:374–384. doi: 10.1046/j.1432-1327.1999.00616.x. [DOI] [PubMed] [Google Scholar]
- 8.Saitou M, Fujimoto K, Doi Y, Itoh M, Fujimoto T, Furuse M, Takano H, Noda T, Tsukita S. Occludin-deficient embryonic stem cells can differentiate into polarized epithelial cells bearing tight junctions. J Cell Biol. 1998;141:397–408. doi: 10.1083/jcb.141.2.397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Saitou M, Furuse M, Sasaki H, Schulzke JD, Fromm M, Takano H, Noda T, Tsukita S. Complex phenotype of mice lacking occludin, a component of tight junction strands. Mol Biol Cell. 2000;11:4131–4142. doi: 10.1091/mbc.11.12.4131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Furuse M, Fujita K, Hiiragi T, Fujimoto K, Tsukita S. Claudin-1 and -2: novel integral membrane proteins localizing at tight junctions with no sequence similarity to occludin. J Cell Biol. 1998;141:1539–1550. doi: 10.1083/jcb.141.7.1539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Heiskala M, Peterson PA, Yang Y. The roles of claudin superfamily proteins in paracellular transport. Traffic. 2001;2:93–98. doi: 10.1034/j.1600-0854.2001.020203.x. [DOI] [PubMed] [Google Scholar]
- 12.Van Itallie CM, Anderson JM. Claudins and epithelial paracellular transport. Annu Rev Physiol. 2006;68:403–429. doi: 10.1146/annurev.physiol.68.040104.131404. [DOI] [PubMed] [Google Scholar]
- 13.Tsukita S, Furuse M, Itoh M. Multifunctional strands in tight junctions. Nat Rev Mol Cell Biol. 2001;2:285–293. doi: 10.1038/35067088. [DOI] [PubMed] [Google Scholar]
- 14.González-Mariscal L, Betanzos A, Nava P, Jaramillo BE. Tight junction proteins. Prog Biophys Mol Biol. 2003;81:1–44. doi: 10.1016/s0079-6107(02)00037-8. [DOI] [PubMed] [Google Scholar]
- 15.Tsukita S, Furuse M. Occludin and claudins in tight-junction strands: leading or supporting players? Trends Cell Biol. 1999;9:268–273. doi: 10.1016/s0962-8924(99)01578-0. [DOI] [PubMed] [Google Scholar]
- 16.Fanning AS, Mitic LL, Anderson JM. Transmembrane proteins in the tight junction barrier. J Am Soc Nephrol. 1999;10:1337–1345. doi: 10.1681/ASN.V1061337. [DOI] [PubMed] [Google Scholar]
- 17.Rahner C, Mitic LL, Anderson JM. Heterogeneity in expression and subcellular localization of claudins 2, 3, 4, and 5 in the rat liver, pancreas, and gut. Gastroenterology. 2001;120:411–422. doi: 10.1053/gast.2001.21736. [DOI] [PubMed] [Google Scholar]
- 18.Reyes JL, Lamas M, Martin D, del Carmen Namorado M, Islas S, Luna J, Tauc M, González-Mariscal L. The renal segmental distribution of claudins changes with development. Kidney Int. 2002;62:476–487. doi: 10.1046/j.1523-1755.2002.00479.x. [DOI] [PubMed] [Google Scholar]
- 19.Enck AH, Berger UV, Yu AS. Claudin-2 is selectively expressed in proximal nephron in mouse kidney. Am J Physiol Renal Physiol. 2001;281:F966–F974. doi: 10.1152/ajprenal.2001.281.5.F966. [DOI] [PubMed] [Google Scholar]
- 20.Kiuchi-Saishin Y, Gotoh S, Furuse M, Takasuga A, Tano Y, Tsukita S. Differential expression patterns of claudins, tight junction membrane proteins, in mouse nephron segments. J Am Soc Nephrol. 2002;13:875–886. doi: 10.1681/ASN.V134875. [DOI] [PubMed] [Google Scholar]
- 21.Turksen K, Troy TC. Claudin-6: a novel tight junction molecule is developmentally regulated in mouse embryonic epithelium. Dev Dyn. 2001;222:292–300. doi: 10.1002/dvdy.1174. [DOI] [PubMed] [Google Scholar]
- 22.Turksen K, Troy TC. Permeability barrier dysfunction in transgenic mice overexpressing claudin 6. Development. 2002;129:1775–1784. doi: 10.1242/dev.129.7.1775. [DOI] [PubMed] [Google Scholar]
- 23.Hellani A, Ji J, Mauduit C, Deschildre C, Tabone E, Benahmed M. Developmental and hormonal regulation of the expression of oligodendrocyte-specific protein/claudin 11 in mouse testis. Endocrinology. 2000;141:3012–3019. doi: 10.1210/endo.141.8.7625. [DOI] [PubMed] [Google Scholar]
- 24.Furuse M, Sasaki H, Tsukita S. Manner of interaction of heterogeneous claudin species within and between tight junction strands. J Cell Biol. 1999;147:891–903. doi: 10.1083/jcb.147.4.891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Morita K, Sasaki H, Furuse M, Tsukita S. Endothelial claudin: claudin-5/TMVCF constitutes tight junction strands in endothelial cells. J Cell Biol. 1999;147:185–194. doi: 10.1083/jcb.147.1.185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Morita K, Sasaki H, Fujimoto K, Furuse M, Tsukita S. Claudin-11/OSP-based tight junctions of myelin sheaths in brain and Sertoli cells in testis. J Cell Biol. 1999;145:579–588. doi: 10.1083/jcb.145.3.579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Colegio OR, Van Itallie CM, McCrea HJ, Rahner C, Anderson JM. Claudins create charge-selective channels in the paracellular pathway between epithelial cells. Am J Physiol Cell Physiol. 2002;283:C142–C147. doi: 10.1152/ajpcell.00038.2002. [DOI] [PubMed] [Google Scholar]
- 28.Van Itallie C, Rahner C, Anderson JM. Regulated expression of claudin-4 decreases paracellular conductance through a selective decrease in sodium permeability. J Clin Invest. 2001;107:1319–1327. doi: 10.1172/JCI12464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Katahira J, Inoue N, Horiguchi Y, Matsuda M, Sugimoto N. Molecular cloning and functional characterization of the receptor for Clostridium perfringens enterotoxin. J Cell Biol. 1997;136:1239–1247. doi: 10.1083/jcb.136.6.1239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.McClane BA. Clostridium perfringens enterotoxin and intestinal tight junctions. Trends Microbiol. 2000;8:145–146. doi: 10.1016/s0966-842x(00)01724-8. [DOI] [PubMed] [Google Scholar]
- 31.Michl P, Buchholz M, Rolke M, Kunsch S, Löhr M, McClane B, Tsukita S, Leder G, Adler G, Gress TM. Claudin-4: a new target for pancreatic cancer treatment using Clostridium perfringens enterotoxin. Gastroenterology. 2001;121:678–684. doi: 10.1053/gast.2001.27124. [DOI] [PubMed] [Google Scholar]
- 32.Martìn-Padura I, Lostaglio S, Schneemann M, Williams L, Romano M, Fruscella P, Panzeri C, Stoppacciaro A, Ruco L, Villa A, et al. Junctional adhesion molecule, a novel member of the immunoglobulin superfamily that distributes at intercellular junctions and modulates monocyte transmigration. J Cell Biol. 1998;142:117–127. doi: 10.1083/jcb.142.1.117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Itoh M, Sasaki H, Furuse M, Ozaki H, Kita T, Tsukita S. Junctional adhesion molecule (JAM) binds to PAR-3: a possible mechanism for the recruitment of PAR-3 to tight junctions. J Cell Biol. 2001;154:491–497. doi: 10.1083/jcb.200103047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Liu Y, Nusrat A, Schnell FJ, Reaves TA, Walsh S, Pochet M, Parkos CA. Human junction adhesion molecule regulates tight junction resealing in epithelia. J Cell Sci. 2000;113(Pt 13):2363–2374. doi: 10.1242/jcs.113.13.2363. [DOI] [PubMed] [Google Scholar]
- 35.Kostrewa D, Brockhaus M, D'Arcy A, Dale GE, Nelboeck P, Schmid G, Mueller F, Bazzoni G, Dejana E, Bartfai T, et al. X-ray structure of junctional adhesion molecule: structural basis for homophilic adhesion via a novel dimerization motif. EMBO J. 2001;20:4391–4398. doi: 10.1093/emboj/20.16.4391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ebnet K, Suzuki A, Ohno S, Vestweber D. Junctional adhesion molecules (JAMs): more molecules with dual functions? J Cell Sci. 2004;117:19–29. doi: 10.1242/jcs.00930. [DOI] [PubMed] [Google Scholar]
- 37.Cunningham SA, Arrate MP, Rodriguez JM, Bjercke RJ, Vanderslice P, Morris AP, Brock TA. A novel protein with homology to the junctional adhesion molecule. Characterization of leukocyte interactions. J Biol Chem. 2000;275:34750–34756. doi: 10.1074/jbc.M002718200. [DOI] [PubMed] [Google Scholar]
- 38.Ikenouchi J, Furuse M, Furuse K, Sasaki H, Tsukita S, Tsukita S. Tricellulin constitutes a novel barrier at tricellular contacts of epithelial cells. J Cell Biol. 2005;171:939–945. doi: 10.1083/jcb.200510043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Riazuddin S, Ahmed ZM, Fanning AS, Lagziel A, Kitajiri S, Ramzan K, Khan SN, Chattaraj P, Friedman PL, Anderson JM, et al. Tricellulin is a tight-junction protein necessary for hearing. Am J Hum Genet. 2006;79:1040–1051. doi: 10.1086/510022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kikuchi S, Ninomiya T, Tatsumi H, Sawada N, Kojima T. Tricellulin is expressed in autotypic tight junctions of peripheral myelinating Schwann cells. J Histochem Cytochem. 2010;58:1067–1073. doi: 10.1369/jhc.2010.956326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Masuda R, Semba S, Mizuuchi E, Yanagihara K, Yokozaki H. Negative regulation of the tight junction protein tricellulin by snail-induced epithelial-mesenchymal transition in gastric carcinoma cells. Pathobiology. 2010;77:106–113. doi: 10.1159/000278293. [DOI] [PubMed] [Google Scholar]
- 42.Kim E, Niethammer M, Rothschild A, Jan YN, Sheng M. Clustering of Shaker-type K+ channels by interaction with a family of membrane-associated guanylate kinases. Nature. 1995;378:85–88. doi: 10.1038/378085a0. [DOI] [PubMed] [Google Scholar]
- 43.Kim E, Naisbitt S, Hsueh YP, Rao A, Rothschild A, Craig AM, Sheng M. GKAP, a novel synaptic protein that interacts with the guanylate kinase-like domain of the PSD-95/SAP90 family of channel clustering molecules. J Cell Biol. 1997;136:669–678. doi: 10.1083/jcb.136.3.669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Fanning AS, Jameson BJ, Jesaitis LA, Anderson JM. The tight junction protein ZO-1 establishes a link between the transmembrane protein occludin and the actin cytoskeleton. J Biol Chem. 1998;273:29745–29753. doi: 10.1074/jbc.273.45.29745. [DOI] [PubMed] [Google Scholar]
- 45.Itoh M, Furuse M, Morita K, Kubota K, Saitou M, Tsukita S. Direct binding of three tight junction-associated MAGUKs, ZO-1, ZO-2, and ZO-3, with the COOH termini of claudins. J Cell Biol. 1999;147:1351–1363. doi: 10.1083/jcb.147.6.1351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Ebnet K, Schulz CU, Meyer Zu Brickwedde MK, Pendl GG, Vestweber D. Junctional adhesion molecule interacts with the PDZ domain-containing proteins AF-6 and ZO-1. J Biol Chem. 2000;275:27979–27988. doi: 10.1074/jbc.M002363200. [DOI] [PubMed] [Google Scholar]
- 47.Wittchen ES, Haskins J, Stevenson BR. Protein interactions at the tight junction. Actin has multiple binding partners, and ZO-1 forms independent complexes with ZO-2 and ZO-3. J Biol Chem. 1999;274:35179–35185. doi: 10.1074/jbc.274.49.35179. [DOI] [PubMed] [Google Scholar]
- 48.Cordenonsi M, D'Atri F, Hammar E, Parry DA, Kendrick-Jones J, Shore D, Citi S. Cingulin contains globular and coiled-coil domains and interacts with ZO-1, ZO-2, ZO-3, and myosin. J Cell Biol. 1999;147:1569–1582. doi: 10.1083/jcb.147.7.1569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Itoh M, Nagafuchi A, Yonemura S, Kitani-Yasuda T, Tsukita S, Tsukita S. The 220-kD protein colocalizing with cadherins in non-epithelial cells is identical to ZO-1, a tight junction-associated protein in epithelial cells: cDNA cloning and immunoelectron microscopy. J Cell Biol. 1993;121:491–502. doi: 10.1083/jcb.121.3.491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.González-Mariscal L, Islas S, Contreras RG, García-Villegas MR, Betanzos A, Vega J, Diaz-Quiñónez A, Martín-Orozco N, Ortiz-Navarrete V, Cereijido M, et al. Molecular characterization of the tight junction protein ZO-1 in MDCK cells. Exp Cell Res. 1999;248:97–109. doi: 10.1006/excr.1999.4392. [DOI] [PubMed] [Google Scholar]
- 51.Betanzos A, Huerta M, Lopez-Bayghen E, Azuara E, Amerena J, González-Mariscal L. The tight junction protein ZO-2 associates with Jun, Fos and C/EBP transcription factors in epithelial cells. Exp Cell Res. 2004;292:51–66. doi: 10.1016/j.yexcr.2003.08.007. [DOI] [PubMed] [Google Scholar]
- 52.Yamamoto T, Harada N, Kano K, Taya S, Canaani E, Matsuura Y, Mizoguchi A, Ide C, Kaibuchi K. The Ras target AF-6 interacts with ZO-1 and serves as a peripheral component of tight junctions in epithelial cells. J Cell Biol. 1997;139:785–795. doi: 10.1083/jcb.139.3.785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Mandai K, Nakanishi H, Satoh A, Obaishi H, Wada M, Nishioka H, Itoh M, Mizoguchi A, Aoki T, Fujimoto T, et al. Afadin: A novel actin filament-binding protein with one PDZ domain localized at cadherin-based cell-to-cell adherens junction. J Cell Biol. 1997;139:517–528. doi: 10.1083/jcb.139.2.517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Citi S, Denisenko N. Phosphorylation of the tight junction protein cingulin and the effects of protein kinase inhibitors and activators in MDCK epithelial cells. J Cell Sci. 1995;108(Pt 8):2917–2926. doi: 10.1242/jcs.108.8.2917. [DOI] [PubMed] [Google Scholar]
- 55.Yonemura S, Itoh M, Nagafuchi A, Tsukita S. Cell-to-cell adherens junction formation and actin filament organization: similarities and differences between non-polarized fibroblasts and polarized epithelial cells. J Cell Sci. 1995;108(Pt 1):127–142. doi: 10.1242/jcs.108.1.127. [DOI] [PubMed] [Google Scholar]
- 56.Asakura T, Nakanishi H, Sakisaka T, Takahashi K, Mandai K, Nishimura M, Sasaki T, Takai Y. Similar and differential behaviour between the nectin-afadin-ponsin and cadherin-catenin systems during the formation and disruption of the polarized junctional alignment in epithelial cells. Genes Cells. 1999;4:573–581. doi: 10.1046/j.1365-2443.1999.00283.x. [DOI] [PubMed] [Google Scholar]
- 57.Ando-Akatsuka Y, Yonemura S, Itoh M, Furuse M, Tsukita S. Differential behavior of E-cadherin and occludin in their colocalization with ZO-1 during the establishment of epithelial cell polarity. J Cell Physiol. 1999;179:115–125. doi: 10.1002/(SICI)1097-4652(199905)179:2<115::AID-JCP1>3.0.CO;2-T. [DOI] [PubMed] [Google Scholar]
- 58.Dye JF, Leach L, Clark P, Firth JA. Cyclic AMP and acidic fibroblast growth factor have opposing effects on tight and adherens junctions in microvascular endothelial cells in vitro. Microvasc Res. 2001;62:94–113. doi: 10.1006/mvre.2001.2333. [DOI] [PubMed] [Google Scholar]
- 59.Tsukamoto T, Nigam SK. Role of tyrosine phosphorylation in the reassembly of occludin and other tight junction proteins. Am J Physiol. 1999;276:F737–F750. doi: 10.1152/ajprenal.1999.276.5.F737. [DOI] [PubMed] [Google Scholar]
- 60.Oshima T, Laroux FS, Coe LL, Morise Z, Kawachi S, Bauer P, Grisham MB, Specian RD, Carter P, Jennings S, et al. Interferon-gamma and interleukin-10 reciprocally regulate endothelial junction integrity and barrier function. Microvasc Res. 2001;61:130–143. doi: 10.1006/mvre.2000.2288. [DOI] [PubMed] [Google Scholar]
- 61.Katahira J, Sugiyama H, Inoue N, Horiguchi Y, Matsuda M, Sugimoto N. Clostridium perfringens enterotoxin utilizes two structurally related membrane proteins as functional receptors in vivo. J Biol Chem. 1997;272:26652–26658. doi: 10.1074/jbc.272.42.26652. [DOI] [PubMed] [Google Scholar]
- 62.McClane BA. The complex interactions between Clostridium perfringens enterotoxin and epithelial tight junctions. Toxicon. 2001;39:1781–1791. doi: 10.1016/s0041-0101(01)00164-7. [DOI] [PubMed] [Google Scholar]
- 63.Burns AR, Bowden RA, MacDonell SD, Walker DC, Odebunmi TO, Donnachie EM, Simon SI, Entman ML, Smith CW. Analysis of tight junctions during neutrophil transendothelial migration. J Cell Sci. 2000;113(Pt 1):45–57. doi: 10.1242/jcs.113.1.45. [DOI] [PubMed] [Google Scholar]
- 64.Ahdieh M, Vandenbos T, Youakim A. Lung epithelial barrier function and wound healing are decreased by IL-4 and IL-13 and enhanced by IFN-gamma. Am J Physiol Cell Physiol. 2001;281:C2029–C2038. doi: 10.1152/ajpcell.2001.281.6.C2029. [DOI] [PubMed] [Google Scholar]
- 65.Nishiyama R, Sakaguchi T, Kinugasa T, Gu X, MacDermott RP, Podolsky DK, Reinecker HC. Interleukin-2 receptor beta subunit-dependent and -independent regulation of intestinal epithelial tight junctions. J Biol Chem. 2001;276:35571–35580. doi: 10.1074/jbc.M106013200. [DOI] [PubMed] [Google Scholar]
- 66.Lechner J, Krall M, Netzer A, Radmayr C, Ryan MP, Pfaller W. Effects of interferon alpha-2b on barrier function and junctional complexes of renal proximal tubular LLC-PK1 cells. Kidney Int. 1999;55:2178–2191. doi: 10.1046/j.1523-1755.1999.00487.x. [DOI] [PubMed] [Google Scholar]
- 67.Ito M, Kodama M, Masuko M, Yamaura M, Fuse K, Uesugi Y, Hirono S, Okura Y, Kato K, Hotta Y, et al. Expression of coxsackievirus and adenovirus receptor in hearts of rats with experimental autoimmune myocarditis. Circ Res. 2000;86:275–280. doi: 10.1161/01.res.86.3.275. [DOI] [PubMed] [Google Scholar]
- 68.Martin TA, Mansel RE, Jiang WG. Antagonistic effect of NK4 on HGF/SF induced changes in the transendothelial resistance (TER) and paracellular permeability of human vascular endothelial cells. J Cell Physiol. 2002;192:268–275. doi: 10.1002/jcp.10133. [DOI] [PubMed] [Google Scholar]
- 69.Pedram A, Razandi M, Levin ER. Deciphering vascular endothelial cell growth factor/vascular permeability factor signaling to vascular permeability. Inhibition by atrial natriuretic peptide. J Biol Chem. 2002;277:44385–44398. doi: 10.1074/jbc.M202391200. [DOI] [PubMed] [Google Scholar]
- 70.Massoumi R, Sjölander A. The inflammatory mediator leukotriene D4 triggers a rapid reorganisation of the actin cytoskeleton in human intestinal epithelial cells. Eur J Cell Biol. 1998;76:185–191. doi: 10.1016/S0171-9335(98)80033-2. [DOI] [PubMed] [Google Scholar]
- 71.Singer KL, Stevenson BR, Woo PL, Firestone GL. Relationship of serine/threonine phosphorylation/dephosphorylation signaling to glucocorticoid regulation of tight junction permeability and ZO-1 distribution in nontransformed mammary epithelial cells. J Biol Chem. 1994;269:16108–16115. [PubMed] [Google Scholar]
- 72.Rao RK, Baker RD, Baker SS, Gupta A, Holycross M. Oxidant-induced disruption of intestinal epithelial barrier function: role of protein tyrosine phosphorylation. Am J Physiol. 1997;273:G812–G823. doi: 10.1152/ajpgi.1997.273.4.G812. [DOI] [PubMed] [Google Scholar]
- 73.Rao RK, Basuroy S, Rao VU, Karnaky Jr KJ, Gupta A. Tyrosine phosphorylation and dissociation of occludin-ZO-1 and E-cadherin-beta-catenin complexes from the cytoskeleton by oxidative stress. Biochem J. 2002;368:471–481. doi: 10.1042/BJ20011804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Ma TY, Tran D, Hoa N, Nguyen D, Merryfield M, Tarnawski A. Mechanism of extracellular calcium regulation of intestinal epithelial tight junction permeability: role of cytoskeletal involvement. Microsc Res Tech. 2000;51:156–168. doi: 10.1002/1097-0029(20001015)51:2<156::AID-JEMT7>3.0.CO;2-J. [DOI] [PubMed] [Google Scholar]
- 75.Staddon JM, Herrenknecht K, Smales C, Rubin LL. Evidence that tyrosine phosphorylation may increase tight junction permeability. J Cell Sci. 1995;108(Pt 2):609–619. doi: 10.1242/jcs.108.2.609. [DOI] [PubMed] [Google Scholar]
- 76.Takeda H, Tsukita S. Effects of tyrosine phosphorylation on tight junctions in temperature-sensitive v-src-transfected MDCK cells. Cell Struct Funct. 1995;20:387–393. doi: 10.1247/csf.20.387. [DOI] [PubMed] [Google Scholar]
- 77.Morgado-Díaz JA, de Souza W. Evidence that increased tyrosine phosphorylation causes disassembly of adherens junctions but does not perturb paracellular permeability in Caco-2 cells. Tissue Cell. 2001;33:500–513. doi: 10.1054/tice.2001.0204. [DOI] [PubMed] [Google Scholar]
- 78.Atkinson KJ, Rao RK. Role of protein tyrosine phosphorylation in acetaldehyde-induced disruption of epithelial tight junctions. Am J Physiol Gastrointest Liver Physiol. 2001;280:G1280–G1288. doi: 10.1152/ajpgi.2001.280.6.G1280. [DOI] [PubMed] [Google Scholar]
- 79.Tomko RP, Xu R, Philipson L. HCAR and MCAR: the human and mouse cellular receptors for subgroup C adenoviruses and group B coxsackieviruses. Proc Natl Acad Sci U S A. 1997;94:3352–3356. doi: 10.1073/pnas.94.7.3352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Escaffit F, Boudreau F, Beaulieu JF. Differential expression of claudin-2 along the human intestine: Implication of GATA-4 in the maintenance of claudin-2 in differentiating cells. J Cell Physiol. 2005;203:15–26. doi: 10.1002/jcp.20189. [DOI] [PubMed] [Google Scholar]
- 81.Buchert M, Papin M, Bonnans C, Darido C, Raye WS, Garambois V, Pélegrin A, Bourgaux JF, Pannequin J, Joubert D, et al. Symplekin promotes tumorigenicity by up-regulating claudin-2 expression. Proc Natl Acad Sci U S A. 2010;107:2628–2633. doi: 10.1073/pnas.0903747107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Benoit YD, Paré F, Francoeur C, Jean D, Tremblay E, Boudreau F, Escaffit F, Beaulieu JF. Cooperation between HNF-1alpha, Cdx2, and GATA-4 in initiating an enterocytic differentiation program in a normal human intestinal epithelial progenitor cell line. Am J Physiol Gastrointest Liver Physiol. 2010;298:G504–G517. doi: 10.1152/ajpgi.00265.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Kimura Y, Shiozaki H, Hirao M, Maeno Y, Doki Y, Inoue M, Monden T, Ando-Akatsuka Y, Furuse M, Tsukita S, et al. Expression of occludin, tight-junction-associated protein, in human digestive tract. Am J Pathol. 1997;151:45–54. [PMC free article] [PubMed] [Google Scholar]
- 84.Manson JM, Rauch M, Gilmore MS. The commensal microbiology of the gastrointestinal tract. Adv Exp Med Biol. 2008;635:15–28. doi: 10.1007/978-0-387-09550-9_2. [DOI] [PubMed] [Google Scholar]
- 85.Frank DN, Pace NR. Gastrointestinal microbiology enters the metagenomics era. Curr Opin Gastroenterol. 2008;24:4–10. doi: 10.1097/MOG.0b013e3282f2b0e8. [DOI] [PubMed] [Google Scholar]
- 86.Qin J, Li R, Raes J, Arumugam M, Burgdorf KS, Manichanh C, Nielsen T, Pons N, Levenez F, Yamada T, et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature. 2010;464:59–65. doi: 10.1038/nature08821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Wang W, Uzzau S, Goldblum SE, Fasano A. Human zonulin, a potential modulator of intestinal tight junctions. J Cell Sci. 2000;113 Pt 24:4435–4440. doi: 10.1242/jcs.113.24.4435. [DOI] [PubMed] [Google Scholar]
- 88.Di Pierro M, Lu R, Uzzau S, Wang W, Margaretten K, Pazzani C, Maimone F, Fasano A. Zonula occludens toxin structure-function analysis. Identification of the fragment biologically active on tight junctions and of the zonulin receptor binding domain. J Biol Chem. 2001;276:19160–19165. doi: 10.1074/jbc.M009674200. [DOI] [PubMed] [Google Scholar]
- 89.Sakaguchi T, Köhler H, Gu X, McCormick BA, Reinecker HC. Shigella flexneri regulates tight junction-associated proteins in human intestinal epithelial cells. Cell Microbiol. 2002;4:367–381. doi: 10.1046/j.1462-5822.2002.00197.x. [DOI] [PubMed] [Google Scholar]
- 90.Nusrat A, von Eichel-Streiber C, Turner JR, Verkade P, Madara JL, Parkos CA. Clostridium difficile toxins disrupt epithelial barrier function by altering membrane microdomain localization of tight junction proteins. Infect Immun. 2001;69:1329–1336. doi: 10.1128/IAI.69.3.1329-1336.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.McNamara BP, Koutsouris A, O'Connell CB, Nougayréde JP, Donnenberg MS, Hecht G. Translocated EspF protein from enteropathogenic Escherichia coli disrupts host intestinal barrier function. J Clin Invest. 2001;107:621–629. doi: 10.1172/JCI11138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Simonovic I, Rosenberg J, Koutsouris A, Hecht G. Enteropathogenic Escherichia coli dephosphorylates and dissociates occludin from intestinal epithelial tight junctions. Cell Microbiol. 2000;2:305–315. doi: 10.1046/j.1462-5822.2000.00055.x. [DOI] [PubMed] [Google Scholar]
- 93.Czerucka D, Dahan S, Mograbi B, Rossi B, Rampal P. Saccharomyces boulardii preserves the barrier function and modulates the signal transduction pathway induced in enteropathogenic Escherichia coli-infected T84 cells. Infect Immun. 2000;68:5998–6004. doi: 10.1128/iai.68.10.5998-6004.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Lu L, Walker WA. Pathologic and physiologic interactions of bacteria with the gastrointestinal epithelium. Am J Clin Nutr. 2001;73:1124S–1130S. doi: 10.1093/ajcn/73.6.1124S. [DOI] [PubMed] [Google Scholar]
- 95.Rescigno M, Urbano M, Valzasina B, Francolini M, Rotta G, Bonasio R, Granucci F, Kraehenbuhl JP, Ricciardi-Castagnoli P. Dendritic cells express tight junction proteins and penetrate gut epithelial monolayers to sample bacteria. Nat Immunol. 2001;2:361–367. doi: 10.1038/86373. [DOI] [PubMed] [Google Scholar]
- 96.Kucharzik T, Walsh SV, Chen J, Parkos CA, Nusrat A. Neutrophil transmigration in inflammatory bowel disease is associated with differential expression of epithelial intercellular junction proteins. Am J Pathol. 2001;159:2001–2009. doi: 10.1016/S0002-9440(10)63051-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Söderholm JD, Olaison G, Peterson KH, Franzén LE, Lindmark T, Wirén M, Tagesson C, Sjödahl R. Augmented increase in tight junction permeability by luminal stimuli in the non-inflamed ileum of Crohn's disease. Gut. 2002;50:307–313. doi: 10.1136/gut.50.3.307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Fries W, Renda MC, Lo Presti MA, Raso A, Orlando A, Oliva L, Giofré MR, Maggio A, Mattaliano A, Macaluso A, et al. Intestinal permeability and genetic determinants in patients, first-degree relatives, and controls in a high-incidence area of Crohn's disease in Southern Italy. Am J Gastroenterol. 2005;100:2730–2736. doi: 10.1111/j.1572-0241.2005.00325.x. [DOI] [PubMed] [Google Scholar]
- 99.Gassler N, Rohr C, Schneider A, Kartenbeck J, Bach A, Obermüller N, Otto HF, Autschbach F. Inflammatory bowel disease is associated with changes of enterocytic junctions. Am J Physiol Gastrointest Liver Physiol. 2001;281:G216–G228. doi: 10.1152/ajpgi.2001.281.1.G216. [DOI] [PubMed] [Google Scholar]
- 100.Ringel Y, Drossman DA. Psychosocial aspects of Crohn's disease. Surg Clin North Am. 2001;81:231–52, x. doi: 10.1016/s0039-6109(05)70283-8. [DOI] [PubMed] [Google Scholar]
- 101.Piche T, Barbara G, Aubert P, Bruley des Varannes S, Dainese R, Nano JL, Cremon C, Stanghellini V, De Giorgio R, Galmiche JP, et al. Impaired intestinal barrier integrity in the colon of patients with irritable bowel syndrome: involvement of soluble mediators. Gut. 2009;58:196–201. doi: 10.1136/gut.2007.140806. [DOI] [PubMed] [Google Scholar]
- 102.Gareau MG, Silva MA, Perdue MH. Pathophysiological mechanisms of stress-induced intestinal damage. Curr Mol Med. 2008;8:274–281. doi: 10.2174/156652408784533760. [DOI] [PubMed] [Google Scholar]
- 103.Schulzke JD, Bentzel CJ, Schulzke I, Riecken EO, Fromm M. Epithelial tight junction structure in the jejunum of children with acute and treated celiac sprue. Pediatr Res. 1998;43:435–441. doi: 10.1203/00006450-199804000-00001. [DOI] [PubMed] [Google Scholar]
- 104.Madara JL, Trier JS. Structural abnormalities of jejunal epithelial cell membranes in celiac sprue. Lab Invest. 1980;43:254–261. [PubMed] [Google Scholar]
- 105.Vogelsang H, Schwarzenhofer M, Oberhuber G. Changes in gastrointestinal permeability in celiac disease. Dig Dis. 1998;16:333–336. doi: 10.1159/000016886. [DOI] [PubMed] [Google Scholar]
- 106.van Elburg RM, Uil JJ, Mulder CJ, Heymans HS. Intestinal permeability in patients with coeliac disease and relatives of patients with coeliac disease. Gut. 1993;34:354–357. doi: 10.1136/gut.34.3.354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Mooradian AD, Morley JE, Levine AS, Prigge WF, Gebhard RL. Abnormal intestinal permeability to sugars in diabetes mellitus. Diabetologia. 1986;29:221–224. doi: 10.1007/BF00454879. [DOI] [PubMed] [Google Scholar]
- 108.Neu J, Reverte CM, Mackey AD, Liboni K, Tuhacek-Tenace LM, Hatch M, Li N, Caicedo RA, Schatz DA, Atkinson M. Changes in intestinal morphology and permeability in the biobreeding rat before the onset of type 1 diabetes. J Pediatr Gastroenterol Nutr. 2005;40:589–595. doi: 10.1097/01.mpg.0000159636.19346.c1. [DOI] [PubMed] [Google Scholar]
- 109.Vaarala O. Leaking gut in type 1 diabetes. Curr Opin Gastroenterol. 2008;24:701–706. doi: 10.1097/MOG.0b013e32830e6d98. [DOI] [PubMed] [Google Scholar]
- 110.Sapone A, de Magistris L, Pietzak M, Clemente MG, Tripathi A, Cucca F, Lampis R, Kryszak D, Cartenì M, Generoso M, et al. Zonulin upregulation is associated with increased gut permeability in subjects with type 1 diabetes and their relatives. Diabetes. 2006;55:1443–1449. doi: 10.2337/db05-1593. [DOI] [PubMed] [Google Scholar]
- 111.Laudat A, Arnaud P, Napoly A, Brion F. The intestinal permeability test applied to the diagnosis of food allergy in paediatrics. West Indian Med J. 1994;43:87–88. [PubMed] [Google Scholar]
- 112.Ventura MT, Polimeno L, Amoruso AC, Gatti F, Annoscia E, Marinaro M, Di Leo E, Matino MG, Buquicchio R, Bonini S, et al. Intestinal permeability in patients with adverse reactions to food. Dig Liver Dis. 2006;38:732–736. doi: 10.1016/j.dld.2006.06.012. [DOI] [PubMed] [Google Scholar]
- 113.Boyle RJ, Hardikar W, Tang ML. The development of food allergy after liver transplantation. Liver Transpl. 2005;11:326–330. doi: 10.1002/lt.20368. [DOI] [PubMed] [Google Scholar]
- 114.Legendre C, Caillat-Zucman S, Samuel D, Morelon S, Bismuth H, Bach JF, Kreis H. Transfer of symptomatic peanut allergy to the recipient of a combined liver-and-kidney transplant. N Engl J Med. 1997;337:822–824. doi: 10.1056/NEJM199709183371204. [DOI] [PubMed] [Google Scholar]
- 115.Groschwitz KR, Hogan SP. Intestinal barrier function: molecular regulation and disease pathogenesis. J Allergy Clin Immunol. 2009;124:3–20; quiz 21-2. doi: 10.1016/j.jaci.2009.05.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Visser J, Rozing J, Sapone A, Lammers K, Fasano A. Tight junctions, intestinal permeability, and autoimmunity: celiac disease and type 1 diabetes paradigms. Ann N Y Acad Sci. 2009;1165:195–205. doi: 10.1111/j.1749-6632.2009.04037.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Deitch EA. The role of intestinal barrier failure and bacterial translocation in the development of systemic infection and multiple organ failure. Arch Surg. 1990;125:403–404. doi: 10.1001/archsurg.1990.01410150125024. [DOI] [PubMed] [Google Scholar]
- 118.Deitch EA, Kemper AC, Specian RD, Berg RD. A study of the relationship among survival, gut-origin sepsis, and bacterial translocation in a model of systemic inflammation. J Trauma. 1992;32:141–147. [PubMed] [Google Scholar]
- 119.Senthil M, Brown M, Xu DZ, Lu Q, Feketeova E, Deitch EA. Gut-lymph hypothesis of systemic inflammatory response syndrome/multiple-organ dysfunction syndrome: validating studies in a porcine model. J Trauma. 2006;60:958–65; discussion 965-7. doi: 10.1097/01.ta.0000215500.00018.47. [DOI] [PubMed] [Google Scholar]
- 120.Deitch EA. Gut lymph and lymphatics: a source of factors leading to organ injury and dysfunction. Ann N Y Acad Sci. 2010;1207 Suppl 1:E103–E111. doi: 10.1111/j.1749-6632.2010.05713.x. [DOI] [PubMed] [Google Scholar]
- 121.Deitch EA, Xu D, Franko L, Ayala A, Chaudry IH. Evidence favoring the role of the gut as a cytokine-generating organ in rats subjected to hemorrhagic shock. Shock. 1994;1:141–145. doi: 10.1097/00024382-199402000-00010. [DOI] [PubMed] [Google Scholar]
- 122.Vollmar B, Menger MD. Intestinal ischemia/reperfusion: microcirculatory pathology and functional consequences. Langenbecks Arch Surg. 2011;396:13–29. doi: 10.1007/s00423-010-0727-x. [DOI] [PubMed] [Google Scholar]
- 123.Deitch EA, Bridges W, Baker J, Ma JW, Ma L, Grisham MB, Granger DN, Specian RD, Berg R. Hemorrhagic shock-induced bacterial translocation is reduced by xanthine oxidase inhibition or inactivation. Surgery. 1988;104:191–198. [PubMed] [Google Scholar]
- 124.Deitch EA, Senthil M, Brown M, Caputo F, Watkins A, Anjaria D, Badami C, Pisarenko V, Doucet D, Lu Q, et al. Trauma-shock-induced gut injury and the production of biologically active intestinal lymph is abrogated by castration in a large animal porcine model. Shock. 2008;30:135–141. doi: 10.1097/shk.0b013e318161724f. [DOI] [PubMed] [Google Scholar]
- 125.Gatt M, Reddy BS, MacFie J. Review article: bacterial translocation in the critically ill--evidence and methods of prevention. Aliment Pharmacol Ther. 2007;25:741–757. doi: 10.1111/j.1365-2036.2006.03174.x. [DOI] [PubMed] [Google Scholar]
- 126.Alverdy JC, Aoys E, Moss GS. Total parenteral nutrition promotes bacterial translocation from the gut. Surgery. 1988;104:185–190. [PubMed] [Google Scholar]
- 127.Vagianos C, Karatzas T, Scopa CD, Panagopoulos C, Tsoni I, Spiliopoulou I, Kalfarentzos F. Neurotensin reduces microbial translocation and improves intestinal mucosa integrity after abdominal radiation. Eur Surg Res. 1992;24:77–83. doi: 10.1159/000129191. [DOI] [PubMed] [Google Scholar]
- 128.Wang Z, Xiao G, Yao Y, Guo S, Lu K, Sheng Z. The role of bifidobacteria in gut barrier function after thermal injury in rats. J Trauma. 2006;61:650–657. doi: 10.1097/01.ta.0000196574.70614.27. [DOI] [PubMed] [Google Scholar]
- 129.Deitch EA, Bridges WM, Ma JW, Ma L, Berg RD, Specian RD. Obstructed intestine as a reservoir for systemic infection. Am J Surg. 1990;159:394–401. doi: 10.1016/s0002-9610(05)81280-2. [DOI] [PubMed] [Google Scholar]
- 130.Sharma B, Srivastava S, Singh N, Sachdev V, Kapur S, Saraya A. Role of probiotics on gut permeability and endotoxemia in patients with acute pancreatitis: a double-blind randomized controlled trial. J Clin Gastroenterol. 2011;45:442–448. doi: 10.1097/MCG.0b013e318201f9e2. [DOI] [PubMed] [Google Scholar]
- 131.Hunninghake GW, Doerschug KC, Nymon AB, Schmidt GA, Meyerholz DK, Ashare A. Insulin-like growth factor-1 levels contribute to the development of bacterial translocation in sepsis. Am J Respir Crit Care Med. 2010;182:517–525. doi: 10.1164/rccm.200911-1757OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Sun YJ, Chen WM, Zhang TZ, Cao HJ, Zhou J. Effects of cardiopulmonary bypass on tight junction protein expressions in intestinal mucosa of rats. World J Gastroenterol. 2008;14:5868–5875. doi: 10.3748/wjg.14.5868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.van Vliet MJ, Harmsen HJ, de Bont ES, Tissing WJ. The role of intestinal microbiota in the development and severity of chemotherapy-induced mucositis. PLoS Pathog. 2010;6:e1000879. doi: 10.1371/journal.ppat.1000879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Assimakopoulos SF, Tsamandas AC, Louvros E, Vagianos CE, Nikolopoulou VN, Thomopoulos KC, Charonis A, Scopa CD. Intestinal epithelial cell proliferation, apoptosis and expression of tight junction proteins in patients with obstructive jaundice. Eur J Clin Invest. 2011;41:117–125. doi: 10.1111/j.1365-2362.2010.02379.x. [DOI] [PubMed] [Google Scholar]
- 135.Assimakopoulos SF, Scopa CD, Vagianos CE. Pathophysiology of increased intestinal permeability in obstructive jaundice. World J Gastroenterol. 2007;13:6458–6464. doi: 10.3748/wjg.v13.i48.6458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Purohit V, Bode JC, Bode C, Brenner DA, Choudhry MA, Hamilton F, Kang YJ, Keshavarzian A, Rao R, Sartor RB, et al. Alcohol, intestinal bacterial growth, intestinal permeability to endotoxin, and medical consequences: summary of a symposium. Alcohol. 2008;42:349–361. doi: 10.1016/j.alcohol.2008.03.131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Kakkos SK, Kirkilesis J, Scopa CD, Arvaniti A, Alexandrides T, Vagianos CE. Nonabsorbable antibiotics reduce bacterial and endotoxin translocation in hepatectomised rats. HPB Surg. 1997;10:283–29; discussion 283-29;. doi: 10.1155/1997/49681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Assimakopoulos SF, Alexandris IH, Scopa CD, Mylonas PG, Thomopoulos KC, Georgiou CD, Nikolopoulou VN, Vagianos CE. Effect of bombesin and neurotensin on gut barrier function in partially hepatectomized rats. World J Gastroenterol. 2005;11:6757–6764. doi: 10.3748/wjg.v11.i43.6757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Ramos AR, Matte U, Goldani HA, Oliveira OL, Vieira SM, Silveira TR. Intestinal permeability assessed by 51Cr-EDTA in rats with CCl4 - induced cirrhosis. Arq Gastroenterol. 2010;47:188–192. doi: 10.1590/s0004-28032010000200014. [DOI] [PubMed] [Google Scholar]
- 140.Scarpellini E, Valenza V, Gabrielli M, Lauritano EC, Perotti G, Merra G, Dal Lago A, Ojetti V, Ainora ME, Santoro M, et al. Intestinal permeability in cirrhotic patients with and without spontaneous bacterial peritonitis: is the ring closed? Am J Gastroenterol. 2010;105:323–327. doi: 10.1038/ajg.2009.558. [DOI] [PubMed] [Google Scholar]
- 141.Lee S, Son SC, Han MJ, Kim WJ, Kim SH, Kim HR, Jeon WK, Park KH, Shin MG. Increased intestinal macromolecular permeability and urine nitrite excretion associated with liver cirrhosis with ascites. World J Gastroenterol. 2008;14:3884–3890. doi: 10.3748/wjg.14.3884. [DOI] [PMC free article] [PubMed] [Google Scholar]