

Abstract
The powdery mildew diseases, caused by fungal species of the Erysiphales, have an important economic impact on a variety of plant species and have driven basic and applied research efforts in the field of phytopathology for many years. Although the first taxonomic reports on the Erysiphales date back to the 1850's, advances into the molecular biology of these fungal species have been hampered by their obligate biotrophic nature and difficulties associated with their cultivation and genetic manipulation in the laboratory. The discovery in the 1990's of a few species of powdery mildew fungi that cause disease on Arabidopsis has opened a new chapter in this research field. The great advantages of working with a model plant species have translated into remarkable progress in our understanding of these complex pathogens and their interaction with the plant host. Herein we summarize advances in the study of Arabidopsis-powdery mildew interactions and discuss their implications for the general field of plant pathology. We provide an overview of the life cycle of the pathogens on Arabidopsis and describe the structural and functional changes that occur during infection in the host and fungus in compatible and incompatible interactions, with special emphasis on defense signaling, resistance pathways, and compatibility factors. Finally, we discuss the future of powdery mildew research in anticipation of the sequencing of multiple powdery mildew genomes. The cumulative body of knowledge on powdery mildews of Arabidopsis provides a valuable tool for the study and understanding of disease associated with many other obligate biotrophic pathogen species.
INTRODUCTION
From the 1943 first proposal of Arabidopsis as a genetic model organism to the publication of its entire genome sequence in 2000 by the Arabidopsis Genome Initiative, this small mustard plant has been the source of a large body of biological information on many topics, some highlighted within this book, including the realm of plant pathology (see Meyerowitz, 2001 for a retrospective of Arabidopsis biology).
Heralded by Friedrich Laibach as a great genetic tool (see Leonelli, 2007 for a brief history of Arabidopsis research), due to its small genome, its relatively short generation time, high fecundity and ease of mutagenesis, Arabidopsis has been used extensively for the past 20 years to unravel basic plant defense response mechanisms that reach beyond the plant biology boundaries (e.g. Ratcliff et al., 1997; Felix et al., 1999; Robatzek et al., 2006; Navarro et al., 2006; Shen et al., 2007). The recent identification of common resistance genes and basic defense-related pathways among cereals and Arabidopsis (Collins et al., 2003; Consonni et al., 2006; Jensen et al., 2007, Shen et al., 2007) further reinforce the value of Arabidopsis as a scientific tool for plant-microbe interaction studies.
The scientific progress made on Arabidopsis pathology research has gained momentum in the past few years. In particular, the study of obligate biotrophic and hemibiotrophic pathogens such as powdery mildews (ascomycetes), downy mildews (oomycetes) and bacteria has significantly contributed to our current knowledge about the molecular basis of basal and isolate-specific defense mechanisms. The discovery and functional characterization of bacterial and oomycete pathogen effectors also brought us closer to a general understanding of their disease-causing mechanisms. In this review, we summarize the scientific progress that has been made in the study of powdery mildew diseases in Arabidopsis and discuss the features of this pathosystem(s) that make it the prototype for the study and understanding of biotrophic fungi-plant interactions in general.
The Magic Roundabout: Life cycle of powdery mildews on Arabidopsis
The causal agents of powdery mildew diseases encompass a diverse range of pathogenic species under the broad umbrella of the order Erysiphales, phylum of Ascomycota (see Braun et al., 2002 for more details). Worldwide, approximately 500 powdery mildew species are able to colonize about 10,000 distinct plant species (Takamatsu, 2004). The disease is characterized by the appearance of white powder-like fungal growth, which consists of the mycelium and spore carriers (conidiophores), on the surface of infected plant organs (predominantly leaves, Figure 1A). Powdery mildew is particularly prevalent in temperate and humid climates where it frequently causes significant yield losses and reductions in product quality in agricultural settings. Among the economically important plant species susceptible to powdery mildew are cereals (e.g. wheat and barley), members of the Solanaceae (e.g. tomato), Cucurbitaceae (e.g. squash), grapevine, and ornamental plants (e.g. roses), (Adam and Somerville, 1996). Although natural and induced resistance to the powdery mildew disease has been known and extensively studied in cereals such as barley (Jørgensen, 1994), it is with the relatively recent identification of fungal species pathogenic on Arabidopsis thaliana that conceptual models of plant resistance and host compatibility are beginning to emerge.
Figure 1.

Macroscopic infection phenotypes of susceptible and resistant Arabidopsis lines.
Rosette leaves of 5–6 week old A. thaliana ecotypes Col-0 (A), Do-0 (B), and Sorbo (C) as well as the powdery mildew resistant mutant pmr6-3 (D) at 13 days post-inoculation with G. orontii. Completion of the asexual powdery mildew life cycle is evidenced by the occurrence of abundant sporulation (white powder) on inoculated rosette leaves of the susceptible accession, Col-0. Younger leaves without disease symptoms emerged after inoculation with fungal conidiospores. Note the difference in appearance of infected leaves of resistant accessions Do-0 and Sorbo. Resistance in both accessions is assumed to be governed by RPW8 (Göllner et al., 2008).
Erysiphe cruciferarum (Koch and Slusarenko, 1990), Golovinomyces (syn. Erysiphe) cichoracearum (Adam and Somerville, 1996) and Golovinomyces (syn. Erysiphe) orontii (Plotnikova et al., 1998) are powdery mildews of cucurbits and crucifers that are able to infect Arabidopsis (Table 1). In addition, the less-extensively characterized tomato powdery mildew pathogen Oidium neolycopersici can successfully reproduce on Arabidopsis (Bai et al., 2008; Ellis and Turner, 2001; Jones et al., 2001; Göllner et al., 2008; Whipps et al., 1998; Xiao et al., 2001; Table 1). Although all aforementioned species can colonize Arabidopsis under controlled laboratory conditions, the fungal species range, frequency and intensity of infections in natural Arabidopsis habitats has, to our knowledge, not been studied. Preliminary observations suggest that powdery mildew does not appear to represent a particularly prevalent disease of Arabidopsis in natural settings (K. Schmid, personal communication). Initial surveys of resistance and susceptibility to various powdery mildew isolates were performed on a broad range of Arabidopsis accessions collected throughout the world (Adam et al., 1999). However, more in-depth pathology and genetic studies involved a defined number of Arabidopsis ecotypes and mutants and are summarized below.
Table 1.
Distinguishing anatomical and developmental features of four powdery mildew species pathogenic on Arabidopsisa.
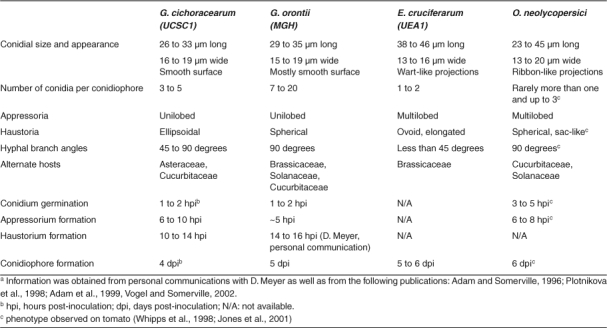
In laboratory conditions, fungal infections are performed by brushing, touching or shaking infected material onto Arabidopsis leaves. Within 1 or 2 hours, a conidium that has landed on a leaf will imbibe and germinate, producing a primary germ tube that differentiates at its terminus a specialized infection structure, the appressorium (Figure 2, 3 and 4A). The appressorium defines the site at which the fungal pathogen attempts to breach the underlying cell wall. A mixture of enzymatic activities and pressure from the penetration peg (2 to 4 MPa in the grass powdery mildew Blumeria graminis; reviewed in Tucker and Talbot, 2001) acting on the epidermal cell pave the way for plant cell invasion. Penetration attempts are typically accompanied on the host side by the production of cell wall appositions (CWA, also termed papillae), which are thought to represent a physical and chemical blockade deployed to arrest infection (see discussion below). Successful penetration results in host plasma membrane invagination to accommodate the primary feeding structure of the fungus, the haustorium (Figure 5, reviewed in O′Connell and Panstruga, 2006). In biotrophic pathogens haustoria are believed to be responsible for the uptake of sugars and amino acids from the plant to the fungal mycelium (Hahn et al., 1997; Voegele et al., 2001) and are thought to actively deliver proteins (effectors) to suppress plant immune responses as is the case for rust fungi (Dodds et al., 2004; O′Connell and Panstruga, 2006). Interestingly, powdery mildew fungi do not invade deeper plant tissues, such as mesophyll cells, but rather limit their growth to the epidermal layer. It has been suggested that the lack of or lower photosynthetic activity of this plant cell layer makes it a powerful nutrient sink that is efficiently exploited by the powdery mildew pathogens (Zeyen et al., 2002). Subsequent to haustorial establishment, secondary hyphal proliferation occurs exclusively on the surface of the leaf or stem (Figure 4B and C). Along the hyphae, new appressoria can form and penetrate epidermal cells and further develop secondary haustoria (Figure 3). Conidiation (the formation and release of asexual spores) completes the vegetative life cycle of the pathogen (Figure 4D).
Figure 2.
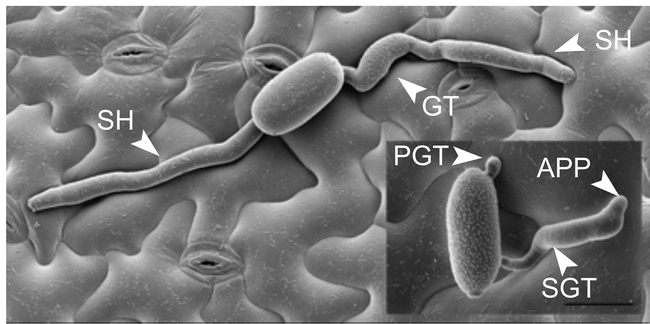
Host and non-host powdery mildew interactions in Arabidopsis.
Scanning electron micrographs of Arabidopsis leaf surface carrying germinated spores of adapted G. orontii (large image) and non-adapted B. graminis f. sp. hordei at 48 hours post-inoculation. Note that Bgh forms a primary (PGT) and a secondary germ tube (SGT), the latter of which differentiates into an appressorium (APP), while G. orontii produces only one germ tube (GT). In the case of Bgh, infection is arrested at this stage in approximately 95% of the cases. In contrast, G. orontii has already formed secondary hyphae (SH) indicating successful host cell penetration and haustorium formation (Photo courtesy of D. Meyer). Scale bar: 20 μm.
Figure 3.
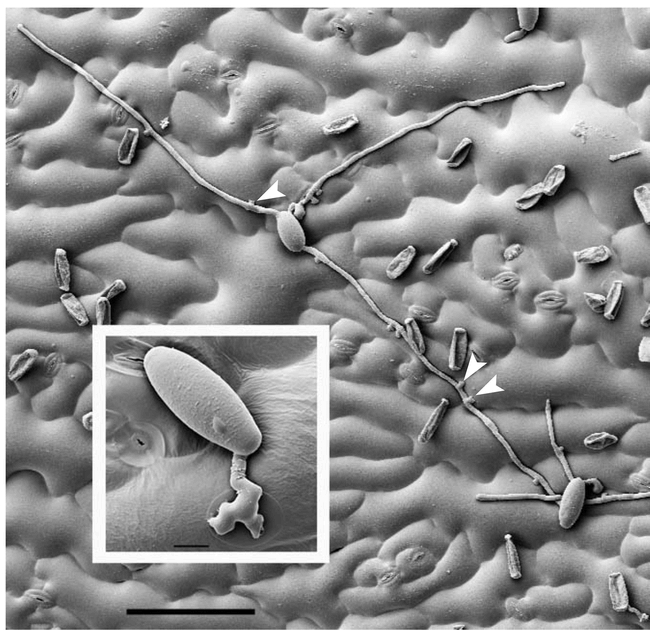
O. neolycopersici growing on Arabidopsis Col-0 at four days after inoculation.
Notice the lobate appressoria (arrowheads) that form at regular intervals along the secondary hyphae. The disease index on Col-0 is usually 2.6 (approximately 30% leaf coverage at 15 days post-inoculation; Bai et al., 2008). Scale bar: 10 μm (inset), 100 μm (large picture).
Figure 4.
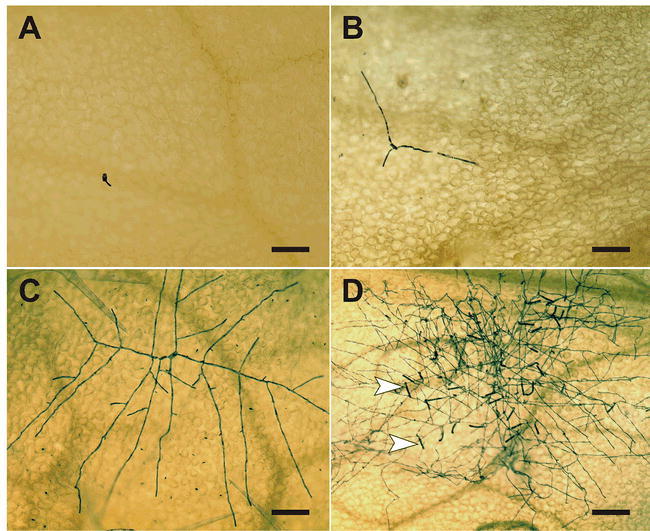
Microscopic analysis of the development of a powdery mildew colony.
The micrographs show the expansion of a G. orontii colony on the surface of a Col-0 rosette leaf. The series of events starts with a germinated spore at 24 hours post-inoculation (A) (see also Figure 2 for further details on spore germination) and continues with initial hyphal elongation (following successful establishment of the first haustorium inside a host cell) at 48 hours post-inoculation (B). Subsequently, a multi-branched mycelium develops (C; photo taken at 63 hours post-inoculation) and the appearance of numerous conidiophores (arrowheads) from a fully expanded fungal colony from 5 days post-inoculation onwards completes the asexual life cycle (D). Fungal structures were highlighted by Coomassie Blue staining of cleared leaf samples. Scale bar: 100 μm (A–C), 200 μm (D).
Figure 5.
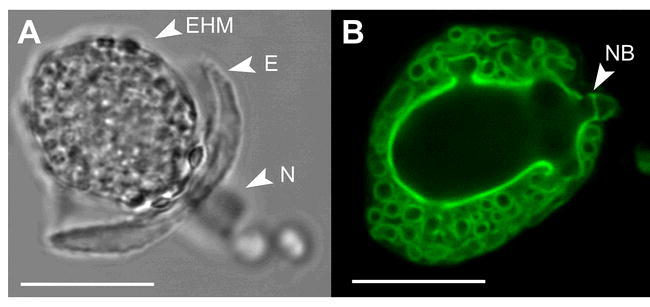
Haustorial complexes of G. orontii.
Phase contrast (A) and epifluorescence (B) micrographs of G. orontii haustoria isolated from Arabidopsis leaves. Notice the highly convoluted and complex folding of the haustorial cell surface providing a large area for nutrient uptake from and effector delivery into the host. Haustoria were labeled with wheat germ agglutinin-FITC. EHM: extrahaustorial membrane, E: encasement, N: haustorial neck, NB: neck band. Scale bars: 20 μm.
The better-studied powdery mildew species on Arabidopsis, namely G. cichoracearum, G. orontii and E. cruciferarum, exhibit different infection intensities that are subject to environmental conditions such as temperature, humidity and light intensity (see Table 1). G. cichoracearum displays abundant conidiation visible to the naked eye, starting at 7 days post-inoculation (dpi), whereas G. orontii requires 10 days for full conidiation and E. cruciferarum sporulates little on Arabidopsis (Vogel and Somerville, 2002). The variability in infection phenotypes correlates with the phylogenetic relationship of the three species (the more virulent species G. cichoracearum and G. orontii being closely related to each other and each being more distantly related to E. cruciferarum; Adam et al., 1999) and is likely indicative of the level of adaptation of each fungus to Arabidopsis. In this sense, the study of the three different powdery mildew species provides a large body of information on the molecular pathways that control this interaction.
The Battleground: Structural and functional aspects of the fungal-plant interface
The intimate interaction established between the plant host and obligate biotrophic fungi such as powdery mildews testifies to the complex cellular processes deployed by both host and pathogen to establish the interface between the two living organisms. The plant cell modifies its architecture and metabolism in response to the invading fungal haustorium to support growth of the fungal colony. Conversely, the fungus adjusts its development and morphology to occupy this niche. Studies into the cellular and sub-cellular structures of the plant-pathogen interface provide valuable information regarding the establishment and maintenance of this fragile interaction (reviewed in O′Connell and Panstruga, 2006).
Among the first critical stages in the infection process of a variety of fungal species is the formation, after germination, of the appressorium and penetration peg. In Arabidopsis and other plant species, attacked cells react to penetration attempts through the formation of CWAs (Figure 6), which – despite sparse experimental evidence supporting this view – are commonly thought to provide both physical cell wall reinforcements as well as a chemical anti-microbial blockade against the invading pathogen (reviewed in Hardham et al., 2007 and Hückelhoven, 2007). Papillae are formed within a few hours after fungal penetration attempts in response to adapted and non-adapted pathogens. Remarkably, physical damage (by needle puncture, for example) can also induce the formation of papilla-like structures, which suggests that it is the pathogen-triggered wounding of plant cells that prompts the formation of CWAs (Aist, 1976). However, detailed analysis of the structures formed in response to non-biotic, mechanical wounds revealed that they are distinct in their composition and likely in their function from fungus-induced CWAs (Russo and Bushnell, 1989). In addition, no papilla formation was detected in response to mechanical (needle) stimulation of parsley suspension culture cells, despite detection of other responses associated with pathogen recognition such as reactive oxygen intermediates (ROI) production and induction of several elicitor-response genes (Gus-Meyer et al., 1998). Although there is no absolute association between papilla formation and resistance to fungal penetration, delayed formation of CWAs is correlated with enhanced fungal infection (Assaad et al., 2004). CWAs are composed of an apparently amorphous mixture of cellulose, pectin, callose, lignins, phenolics, silicon, H2O2 and derivatives as well as dedicated de novo-synthesized antimicrobial metabolites (phytoalexins) and fungal enzyme inhibitors (Figure 6). Some of these compounds are delivered within secretory vesicles while others are synthesized on the spot by cell wall-resident enzymes (reviewed in Hückelhoven, 2007). Each component of the CWA likely plays a role in defense. Structural components such as pectin, callose and cellulose are thought to provide physical reinforcement to the plant cell wall at the site of attack (Zeyen et al., 2002). In addition, released moieties of these carbohydrate polymers as well as enzymes involved in their synthesis have potent signaling capabilities and are thought to be involved in the modulation of downstream defense responses (see discussion below). For example, callose synthase GSL5/PMR4 protein (and/or callose itself) appears to be a negative regulator of SA-dependent defense responses and mutations in this gene render Arabidopsis resistant to G. orontii and G. cichoracearum (Jacobs et al., 2003; Nishimura et al., 2003).
Figure 6.
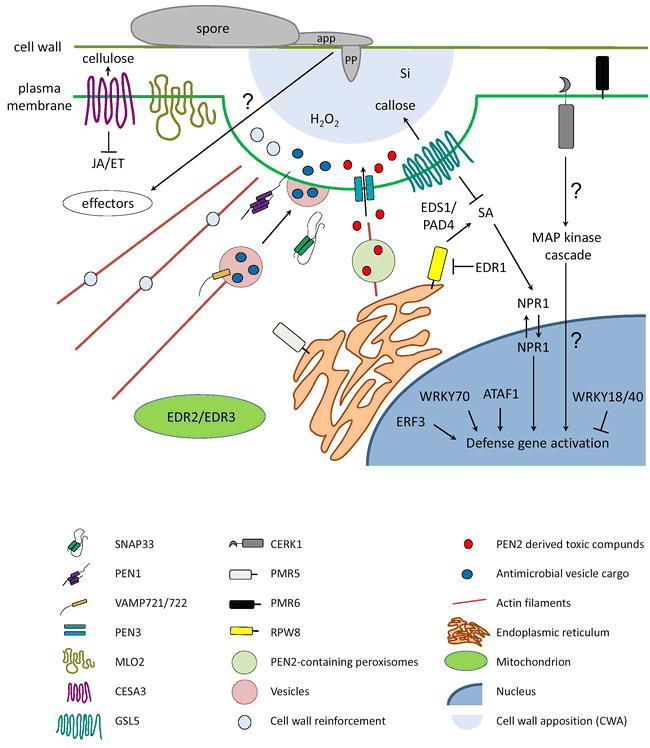
Schematic diagram illustrating genetically-anchored components in Arabidopsis powdery mildew susceptibility/resistance.
The figure depicts a section of a host cell attacked by a powdery mildew germling. Components coded by shape and color are explained in the legend below the scheme. app: appressorium, pp: penetration peg, Si: silicon. Question marks indicate presumed links/activities. For further details, see main text
Silicon (Si) is another component found in papillae (Figure 6) that was long thought to provide structural reinforcement to the cell wall (Zeyen et al., 2002). More recently, a comparative transcriptome analysis of Arabidopsis plants either supplied with or lacking Si, revealed that silicon modulates gene expression in a pathogen-dependent manner (Fauteux et al., 2006). Specifically, it appears to dampen general stress responses associated with pathogen infection and partially restores expression of genes encoding components of primary metabolism to normal conditions without affecting the expression of defense-related genes. Reduction of biotic stress responses results in reduced powdery mildew density and conidiation on leaves, effects that had been historically reported for a variety of other plant species treated with Si (Ghanmi et al., 2004).
To complete defense at the cell wall, pathogen-induced antimicrobial compounds (phytoalexins) are actively delivered to the site of fungal contact. Components of the basal defense machinery such as the PEN2 glycosyl hydrolase (associated with peroxisomes; Figure 6) and the plasma membrane-resident PEN3 ABC transporter (Figure 6) are thought to cooperate and generate toxic anti-microbial compounds that are pumped at the site of attack (Lipka et al., 2005; Stein et al., 2006; see below).
The delivery of cell wall components and anti-microbial compounds at CWAs is thought to occur at least in part in membrane-bound vesicles that move along tracks of actin microfilaments (Figure 6). This view is supported by the observation of massive cytoskeletal re-organization of plant cells upon pathogen attack. Reminiscent of findings in other plant-powdery mildew interactions (Kobayashi et al., 1997a; Opalski et al., 2005) fluorescence microscopy of GFP-tagged Arabidopsis cytoskeletal proteins revealed a rapid concentration of actin arrays towards the site of attack by the non-adapted fungus B. graminis f. sp. hordei. This rearrangement was accompanied by a mobilization of the nucleus and endoplasmic reticulum to the site of the emergent penetration peg and later towards the developing haustorium (Takemoto et al., 2006). Golgi-derived bodies circulate and make frequent stops below the incipient penetration peg, consistent with the hypothesis that the plant protein synthesis and secretory machinery is actively recruited to the emerging infection structures (Hardham et al., 2007). Mutations in PEN1, originally identified as a component of powdery mildew non-host resistance, did not affect actin re-organization upon challenge with the non-adapted barley powdery mildew fungus (Takemoto et al., 2006), suggesting that this protein acts downstream of actin re-organization. However, in barley cells, chemical and genetic interference with actin polymerization significantly affects basal defense and blocks at least partially non-host resistance against inappropriate powdery mildews (Kobayashi et al., 1997a and b; Miklis et al., 2007). The scenario is similar in Arabidopsis (Yun et al., 2003) and suggests an important functional as well as structural role for cytoskeletal components in plant defense responses (reviewed in Schmidt and Panstruga, 2008).
It is interesting to note that plant cytoskeleton and organelle responses documented for powdery mildew infections are strikingly similar to those observed during infection of several plant species with beneficial arbuscular mycorrhizal (AM) fungi (Strack et al., 2003). In this case, the plant cell guides the formation of intracellular arbuscules (analogous in structure and function to haustoria) through the formation of microtubule tracks and by re-organization of the actin network. In tomato, inhibition of microfilament formation interferes with the proper development of the fungus on root tissue (Timonen et al., 2006).
In pathogenic powdery mildew-plant interactions, components of both fungal and plant origin are implicated in the formation of the haustorium. Post-penetration, the plant plasma membrane invaginates and the haustorium starts to develop, initially as a bud, then as a relatively smooth globular body and finally matures into an intricate, highly convoluted and branched structure (Koh et al., 2005; Figure 5). The haustorium is a unicellular structure shielded from the plant environment by its own plasma membrane, cell wall, extrahaustorial matrix (an amorphous environment separating plant and fungal structures) and extrahaustorial membrane (EHM, a unique interface between the host and the pathogen, thought to be composed of a modified plant plasma membrane; Bracker, 1968). In studies of G. cichoracearum haustoria on Arabidopsis, fluorescently labeled host plasma membrane markers spread as far as the two haustorial neckbands that separate fungal from plant structures. Below the neckbands, the EHM has a distinct architecture and function from the plant and fungal membranes (Koh et al., 2005). ATPase and other enzymatic activities normally found at the plant plasma membrane are lacking in the EHM (Spencer-Phillips and Gay, 1981). The EHM enveloping haustoria of the pea powdery mildew, Erysiphe pisi, is twice as thick as normal host plasma membrane, is highly convoluted in healthy haustorial complexes and is resistant to detergent treatments (Gil and Gay, 1977). The biogenesis and molecular composition of EHMs covering the haustoria of biotrophic pathogens represents indeed one of the major mysteries of plant-powdery mildew interactions. Under optimal conditions, the life-span and transport capacity of a single haustorium appears to be sufficient to allow the fungus to complete its life cycle (Shirasu et al., 1999). However, defense responses deployed by the host can interfere with optimal haustorium development and function. In Arabidopsis cells infected with G. orontii, for example, mature haustoria are frequently encased (partially or fully) in what appears to be an extension of the papillary structure, characterized by presence of callose (D. Meyer, P. Schulze-Lefert, R. O′Connell and C. Micali, unpublished data). These encasements, when complete, appear to crush haustorial bodies and may restrict passage of nutrients to the fungus and pathogen effectors to the host. As such they may represent one of the several layers of resistance to powdery mildew infection that are discussed in more detail below.
Déjà vu: RPW8-mediated defense as a recurrent theme in natural powdery mildew resistance
Efforts in dissecting resistance to powdery mildews in Arabidopsis have exploited the natural variation in resistance phenotypes among several hundred accessions collected worldwide (Adam and Somerville, 1996; Adam et al., 1999; Göllner et al., 2008). In most accessions analyzed to date resistance is either polygenic, based on the atypical resistance (R) gene RESISTANCE TO POWDERY MILDEW 8 (RPW8), or on combinations thereof, with RPW8 representing the major Quantitative Trait Locus (Adam and Somerville, 1996; Xiao et al., 1997; Adam et al., 1999; Schiff et al., 2001; Wilson et al., 2001; Orgil et al., 2007; Göllner et al., 2008; Figure 1A–C, Figure 7). In general, more accessions are resistant to E. cruciferarum (76%) than to G. cichoracearum (63%) (Adam et al., 1999), while out of 64 accessions tested in an independent study, only 26% were resistant to G. orontii (Göllner et al, 2008). These numbers may reflect the degree of adaptation that each of these powdery mildew species possesses to cope with the variations of the antifungal defense repertoire that is common in natural Arabidopsis populations. Notably, the reference ecotype Col-0 was found to be susceptible to all four powdery mildew fungi known to successfully colonize A. thaliana (Xiao et al., 1997; Plotnikova et al., 1998; Adam et al., 1999).
Figure 7.
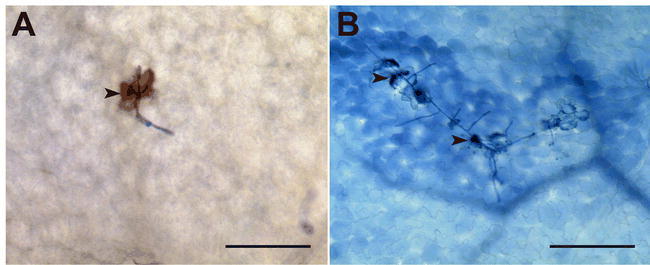
Typical cytological features of RPW8-mediated powdery mildew resistance in ecotype Ms-0.
Whole-cell hydrogen peroxide (H2O2) accumulation in a G. orontii-attacked leaf epidermal cell of the Ms-0 ecotype (A), which is known to express RPW8-based powdery mildew resistance (Xiao et al., 2001). H2O2 accumulation is highlighted as a brownish 3,3-diaminobenzidine (DAB) precipitate (Thordal-Christensen et al., 1997) inside the challenged cell (arrowhead). Trailing necrosis of G. orontii-challenged leaf epidermal cells highlighted by Trypan-blue staining (B). Exemplary dead cells are highlighted by arrowheads. Scale bars: 100 μm
RPW1, RPW2, RPW4 and RPW5 were described as semi-dominant resistance loci and localized to chromosomes II, III, IV and V, respectively (Adam and Somerville, 1996). Additional RPW genes, including RPW6 through RPW13 were later identified (Xiao et al., 1997; Adam et al., 1999; Wilson et al., 2001). In depth analysis of the genetic basis underlying natural resistance repeatedly pointed to locus RPW8 as a key player (found in accessions Co-3, Do-0, Ei-4, Ei-5, Kas-1, Ms-0, Nok-3, Shahdara and Wa-1; Wilson et al., 2001; Schiff et al., 2001; Xiao et al., 2001; Göllner et al. 2008). In Ms-0, the best-studied accession exhibiting RPW8-mediated resistance, the RPW8 locus comprises two naturally polymorphic and dominant genes, RPW8.1 and RPW8.2, which control resistance to a broad range of powdery mildew pathogens (Xiao et al., 2001). This scenario is in contrast to the situation with the majority of other R genes, which generally confer isolate-specific resistance (reviewed in Martin et al., 2003). In addition to the functional RPW8 copies, the Ms-0 locus contains the sequence-related paralogs HR1, HR2 and HR3, which have been shown not to contribute to resistance (Xiao et al., 2001). Two basic Arabidopsis haplotypes have been identified at the RPW8 locus based on the presence/absence of RPW8.1 and RPW8.2: one comprises both RPW8.1 and RPW8.2 and the other, found in susceptible ecotypes, contains HR4 in replacement of RPW8.1 and RPW8.2 (Xiao et al., 2001; Xiao et al., 2004).
The RPW8.1 and RPW8.2 proteins share 45% identity at the amino acid level and lack the typical nucleotide-binding site (NBS) and leucine-rich repeat (LRR) motifs found in most R proteins (Martin et al., 2003). Instead, they carry a predicted coiled-coil (CC) domain and an N-terminal transmembrane (TM) helix that is assumed to localize RPW8 to endomembranes (Wang et al., 2007; Figure 6). However, as in the case of most R genes, RPW8-mediated resistance is associated with the expression of pathogenesis-related (PR) genes and triggers a hypersensitive response (HR; evidenced by whole cell callose deposition, H2O2 accumulation and cell death) in response to pathogen attack (Xiao et al., 2001; Xiao et al., 2003; Xiao et al., 2005; Göllner et al., 2008; Figure 7). Signaling in RPW8-mediated resistance has been proposed to occur through a feed back amplification loop in the salicylic acid (SA) pathway and requires the classical components PHYTOALEXIN DEFICIENT 4 (PAD4), ENHANCED DISEASE SUSCEPTIBILITY 1 (EDS1), EDS5 and NONEXPRESSOR OF PATHOGENESIS-RELATED PROTEIN 1 (NPR1) (Figure 6). Resistance conferred by RPW8 is however independent of the ethylene (ET) and jasmonic acid (JA)-dependent signaling pathways, since mutations in CORONATINE INSENSITIVE 1 (COI1) and ETHYLENE INSENSITIVE 2 (EIN2) do not compromise RPW8-mediated resistance (Xiao et al., 2005). In addition to protection from powdery mildew fungi, RPW8 conveys resistance to oomycetes (Hyaloperonospora parasitica, the causal agent of the Arabidopsis downy mildew disease) and the cauliflower mosaic virus, but not to bacteria (Pseudomonas syringae), suggesting that this protein is a component of a larger signaling conduit leading to broad-spectrum resistance to biotrophs, rather than a bona fide R gene (Wang et al., 2007). Since the pathways controlling resistance against biotrophic and necrotrophic pathogens appear often to be antagonistic (Feys and Parker, 2000; Glazebrook, 2005; Gupta et al., 2000; Ton et al., 2002), it is not surprising that RPW8 expression also leads to enhanced susceptibility to necrotrophic fungi such as Alternaria and Botrytis species (Wang et al., 2007). In this respect, Arabidopsis is confronted with the same dilemma as other plant species: enhanced protection against one type of pathogen might open the door for another. Consequently, occurrence of RPW8 is associated with both fitness benefits and costs, depending on the presence or absence of the concerned pathogens (Orgil et al., 2007).
Despite in-depth searches into Arabidopsis natural isolates from diverse geographical locations, no true race-specific resistance genes against powdery mildew fungi have been identified yet (Göllner et al., 2008). This is in stark contrast to other plant species such as barley (Halterman et al., 2001), grapevine (Barker et al., 2005) and tomato (Bai et al., 2005; Li et al., 2007), where in each case multiple R genes recognizing specific fungal avirulence determinants exist. This striking observation suggests that the Arabidopsis-powdery mildew pathosystem might have developed relatively recently and has not had time to mature the classical Avr/R gene pairs observed in other pathosystems. Alternatively, it is possible that Arabidopsis is not the primary host of these powdery mildew species in nature (Schulze-Lefert and Vogel, 2000). Finally, the emergence of RPW8-based broad spectrum resistance in Arabidopsis may have eliminated the evolutionary driving force for the acquisition of prototypical R genes conferring race-specific resistance. This possibility underscores the importance of the RPW8 locus as a main component of defense against powdery mildews in Arabidopsis (Xiao et al., 2004; Orgil et al., 2007; Göllner et al., 2008).
Non-host Defenses: Trespassers will be shot! Survivors will be shot again!
Non-host resistance has been defined as the capacity of a particular plant species to resist infection by all genetic variants of a pathogen that normally infect other host species (Thordal-Christensen, 2003). Recent in-depth analysis of genes initially identified as “non-host” defense components suggests that plants deploy a similar arsenal in response to both adapted and nonadapted invaders. The intensity and rapidity of the defense response appear to be essential in determining whether a particular microbe will successfully colonize the plant or be blocked in its attempts.
In Arabidopsis, resistance to non-adapted powdery mildews such as Blumeria graminis f. sp. hordei (Bgh; pathogenic on barley) and Erysiphe pisi (pathogenic on pea) is readily detectable at the penetration stage, by the arrest of approximately 80 to 90% of host cell entry attempts. Rare cases of haustorium formation are usually accompanied by callose encasement and often hypersensitive death of the attacked epidermal cell, which prevents any further fungal development (Collins et al., 2003; Assaad et al., 2004; Lipka et al., 2005; Stein et al., 2006). In forward genetic screens, four penetration (pen) mutants impaired in pre-invasion defenses have been identified, indicating that the respective wild type genes (PEN1 through PEN4) are essential for non-host resistance to Bgh and E. pisi (Collins et al., 2003; Lipka et al., 2005; Stein et al., 2006; M. Lim and S. Somerville, unpublished data). All four mutants show enhanced penetration and haustorium formation by the two non-host powdery mildew species, yet fail to support further fungal development and sporulation, suggesting that additional defense responses downstream of entry control are at play.
PEN1 encodes a NSF (N-ethylmaleimide–sensitive factor) attachment protein receptor (SNARE) domain-containing and plasma-membrane resident syntaxin (Collins et al., 2003). Fluorophore-tagged versions of PEN1 have been shown to focally accumulate beneath papillae at sites of attempted fungal penetration (Assaad et al., 2004; Bhat et al., 2005). The PEN1 protein associates via the adaptor protein SNAP33 to Vesicle Associated Membrane Proteins (VAMP), preferentially VAMP721 and VAMP722, to form ternary SNARE complexes (Kwon et al., 2008a; Figure 6). In eukaryotes, ternary SNARE complexes mediate membrane fusion events between vesicles and target membranes (reviewed in Lipka et al., 2007). In the context of antifungal defense, the association of PEN1, SNAP33 and VAMP721/722 is believed to tether secretory vesicles to the plasma membrane and eventually force the release of vesicle cargo to the outside of the infected cell (Kwon et al., 2008a). Although the nature of the vesicles and the contents of the cargo are currently unknown, one can not help but compare the action of PEN1 to that of other syntaxins involved in cell-mediated resistance and inflammatory responses in mammalian systems (Skula et al., 2000). In this well-studied system, immune cells such as cytotoxic T cells and Natural Killer cells can deliver suicide orders such as the release of GranzymeB-packaged vesicles, to virus-infected cells. By analogy to these mammalian defense systems, PEN1 is postulated to mediate the release of anti-microbial and possibly cell-wall reinforcing compounds at the site of fungal attack that spare the plant cell from cell death (Kwon et al., 2008b). Delivery of anti-microbial and cell-wall components within specialized vesicles supposedly depend on intact cytoskeleton tracks (see discussion above and Figure 6).
Similarly, the PEN2 and PEN3 proteins also contribute to penetration resistance at the cell periphery. PEN2 shows sequence similarity to glycosyl hydrolases and PEN3 is a predicted ATP binding cassette (ABC) transporter (Lipka et al., 2005; Stein et al., 2006). PEN2 was recently shown to possess unexpected myrosinase activity in vitro, processing indole glucosinolates into apparently novel toxic products that may act to block further fungal progression (P. Bednarek, personal communication; Figure 6). PEN3 is thought to cooperate with PEN2 in pumping out these toxic compounds at the site of fungal penetration (Lipka et al., 2005; Stein et al., 2006). In accordance with this hypothesis, functional PEN3-GFP localizes to the plasma membrane and accumulates at the site of Bgh penetration (Figure 6). Beyond fungal penetration, the activities of PEN2 and PEN3 are thought to further play a role in resistance subsequent to invasion, by poisoning the fungal haustorium with toxic products delivered to the extrahaustorial matrix (Stein et al., 2006). In contrast to pen1 mutants, which show no dramatic alterations in response to appropriate powdery mildews, pen2 is hyper-susceptible to the adapted powdery mildews G. orontii and G. cichoracearum (Lipka et al., 2005). In addition strong chlorosis symptoms were observed on pen3 after inoculation with the adapted powdery mildew species, G. cichoracearum (Stein et al., 2006). This chlorosis phenotype is proposed to be the result of high levels of toxic PEN2-derived products, which accumulate in response to repeated epidermal penetration by G. cichoracearum in the absence of PEN3 (Stein et al., 2006). It is notable that the requirements for non-host resistance appear to strongly depend on the nature of the attacking pathogen, since the PEN1 syntaxin is functionally dispensable for effective pre-invasion (penetration) resistance in non-host interactions with different Colletotrichum species (Shimada et al., 2006). Similarly, pen2 and pen3 but not pen1 mutants showed enhanced susceptibility to the inappropriate hemibiotrophic oomycete pathogen Phytophthora infestans and the necrotrophic fungus Plectosphaerella cucumerina, suggesting that the two former components of non-host defense play a wider role in preventing pathogen attack than the latter (Lipka et al., 2005; Stein et al., 2006).
In addition to pre-invasion resistance controlled by PEN genes, post-invasive defenses restrict pathogen growth after the formation of the primary haustorium. The effects of post-penetration defenses were revealed by the removal, through mutation, of penetration resistance associated with PEN genes (Lipka et al., 2005). Genes encoding the defense components PAD4, EDS1 and SAG101 (SENESCENCE ASSOCIATED GENE 101; Lipka et al., 2005; Stein et al., 2006) are involved in the establishment of basal defense during compatible interactions and the execution of isolate-specific resistance conferred by a subset of R genes (Parker et al., 1996; Feys et al., 2005). In the double mutants pen2 pad4 and pen2 eds1, the non-adapted fungus Bgh was able to develop secondary hyphae while in the triple mutant pen2 pad4 sag101, microcolonies were formed and the pathogen was able to complete its life cycle, effectively abolishing non-host resistance in this interaction (Lipka et al., 2005). In addition, triple mutants exhibited full susceptibility to another non-adapted pathogen, E. pisi, allowing sporulation comparable to the interaction between wild-type plants and an adapted powdery mildew fungus such as G. orontii (Lipka et al., 2005).
Taken together, cumulative mutations in all layers of the non-host defense machinery can transform a non-adapted powdery mildew fungus into a full-fledged pathogen, suggesting that it is largely the ability or inability to overcome species-specific defense barriers that determines the success or failure of a microbe in a given plant-pathogen interaction. Conversely, as outlined below, mutations in so-called compatibility genes can partially or completely close the door to adapted fungi.
Passing on the Message: Defense signaling in Arabidopsis-powdery mildew interactions
It is currently widely accepted (although exceptions exist, see below) that defense signaling in response to biotrophic pathogens typically converges on a pathway controlled by salicylic acid (Glazebrook, 2005). In contrast, defense pathways controlled by JA and ET are overall antagonistic to the SA pathway and are in general known to govern resistance to necrotrophs. Early studies revealed a collection of SA pathway genes involved in basal defense, including EDS1, PAD4, and NPR1. Individuals carrying mutations in these genes allow enhanced growth of the compatible powdery mildew pathogens G. cichoracearum and G. orontii (Reuber et al., 1998; Glazebrook, 2005), demonstrating the importance of these SA signaling components in limiting growth of powdery mildews on Arabidopsis.
NPR1 and its homologues are major convergence points for SA and JA signaling. NPR1 is a positive regulator of SA signaling and controls basal defense to a variety of pathogens including powdery mildews. Under non-challenged conditions NPR1 exists as cytoplasmic oligomers. Upon increase in SA levels triggered by pathogen attack, it monomerises and shuttles to the nucleus where it interacts directly with TGA transcription factors and activates PR genes and genes encoding WRKY transcription factors responsible for plant defense and cell death (reviewed in Loake and Grant, 2007; Figure 6). The NPR1 homologue NPR4 appears to have a similar role in basal defense. T-DNA insertion mutants in NPR4 are compromised in their resistance to G. cichoracearum (Liu et al., 2005). NPR4 shares 36% amino acid identity to NPR1, its transcript is induced upon SA treatment and it mediates both SA- and JA-dependent gene induction, in a parallel but distinct pathway from NPR1.
In addition to SA signaling, the JA and ET signaling pathways can also contribute to powdery mildew resistance. Although these pathways do not appear to be induced in compatible powdery mildew infections (Reuber et al., 1998; Zimmerli et al., 2004; Glazebrook, 2005), mutants in CESA3 (cev1), encoding a plasma membrane-resident cellulose synthase (Figure 6), produce JA and ET constitutively, have elevated stress response gene expression and are resistant to G. cichoracearum (Ellis et al., 2002a and b). Induction of JA responsive genes can also be reproduced by treatment of wild type plants with cellulose synthesis inhibitors, suggesting a role for plant cell wall signaling in general defense responses. However, the artificial means used to induce JA signaling in this mutant and the fact that JA-dependent defense responses are not typically triggered by compatible powdery mildew species (or may actively be suppressed) argues against an authentic role of JA and ET in resistance against these fungal pathogens under natural conditions.
The Right Stuff: Arabidopsis-powdery mildew compatibility factors
The biotrophic lifestyle of powdery mildew fungi dictates that they infect plants and establish colonies in a “quiet” manner, so as not to arouse the defense mechanisms of the host (Panstruga, 2003). They likely also usurp host infrastructure for transport of nutrients and building blocks and coerce the host cell into accommodating their impressive haustorial feeding structures (Figure 5, Mendgen and Hahn, 2002; Schulze-Lefert and Panstruga, 2003). Specific host genes and/or proteins termed compatibility or susceptibility factors are believed to be essential for successful pathogenesis by a given microbe, and a lack of these factors is predicted to result in resistance to an otherwise virulent pathogen (Vogel and Somerville, 2000; Panstruga, 2003). To isolate compatibility factors, two independent genetic screens were performed in the late 1990s and identified many new components that paint an ever more complex picture of Arabidopsis cellular mechanisms of defense.
A hallmark of constitutive defense is the transcriptional activation of PR genes such as PR1 (reviewed in Loake and Grant, 2007). In an attempt to identify novel elements of resistance, three powdery mildew (G. cichoracearum) resistant mutants that do not constitutively express PR1 were isolated in the lab of R. Innes and named enhanced disease resistance (edr) 1, edr2 and edr3 (Frye and Innes, 1998; Frye et al., 2001; Tang and Innes, 2002; Tang et al., 2005a and b; Tang et al., 2006). All three mutants display late-onset of resistance (5 to 8 days post-inoculation) that is characterized by accelerated cell death in mesophyll cells leading to large necrotic lesions and limited or absent conidiation of the pathogen. Resistance in all cases is SA-dependent and JA-independent.
EDR1 encodes a CTR1-like protein kinase and negative regulator of disease resistance (Frye et al., 2001). Presence of EDR1 limits the transcriptional amplification of RPW8.1 and reduces RPW8-conditioned host cell death in response to pathogen attack (Xiao et al., 2005; Figure 6). EDR2 encodes a mitochondrial protein (Figure 6) with a pleckstrin homology domain and a steroidogenic acute regulatory protein-related lipid-transfer (START) motif that are each indicative of a role in lipid binding and signaling. Both EDR1 and EDR2 function in a common pathway since double mutants display resistance phenotypes identical to the respective single mutant plants (Tang et al., 2005a). In addition, both mutants display enhanced senescence in response to ethylene. In contrast, EDR3 appears to function in an independent pathway from EDR1 and EDR2 and the corresponding mutant does not display an early senescence phenotype. Like EDR2, EDR3 localizes at least in part to mitochondria and encodes a dynamin-like protein with an N-terminal GTPase domain and a C-terminal GTPase effector region (Tang et al., 2006; Figure 6). In mammals such proteins are responsible for membrane tubulation and vesicle pinching and regulate mitochondrial dynamics associated with apoptosis (programmed cell death; Tang et al., 2006). The characterization of edr mutants suggests a link between SA-mediated resistance, mitochondrial function and programmed cell death and further stresses the broad parallels that exist between animal and plant immune responses (Ausubel, 2005). Indeed, a central feature of defense responses in mammalian systems is the ability to undergo apoptosis in response to pathogen (or non-self) detection (Hiscott et al., 2006). Often the apoptotic program is either initiated or amplified in mitochondria and components of the mitochondrial apoptosome are absolutely required for completion of programmed cell death (Keeble and Gilmore, 2007).
In an independent screen performed by Vogel and Somerville to recover loss-of-susceptibility to G. cichoracearum mutants in Arabidopsis, the powdery mildew resistant (pmr) mutants pmr1 to pmr6 were isolated and to date four of them have been cloned (Vogel and Somerville, 2000; Vogel et al., 2002; Jacobs et al., 2003; Nishimura et al., 2003; Vogel et al., 2004; Consonni et al., 2006; Figure 1D). PMR2 is allelic to MLO2 (see also below), PMR4 (synonyms CALS12 and GSL5) encodes a wound- and pathogen-associated callose synthase, PMR5 belongs to a large family of plant-specific genes of unknown function and PMR6 encodes a glycosyl-phosphatidyl-inositol (GPI)-anchored pectatelyase-like protein (Vogel et al., 2002; Jacobs et al., 2003; Nishimura et al., 2003; Vogel et al., 2004; Consonni et al., 2006). In pmr4, pmr5 and pmr6 mutants resistance is entirely established post penetration (M. Humphry, unpublished data) and results in reduced colony size and conidiophore formation in interactions with both G. cichoracearum and G. orontii. The PMR genes cloned to date are involved in different cellular activities, underscoring the diversity of plant biological processes that are required for successful powdery mildew pathogenesis.
PMR4 controls callose synthesis at cell wall appositions (papillae) that form beneath infection and wound sites and that are believed to provide a physical barrier to fungal penetration (Aist and Bushnell, 1991; Figure 6). It therefore came as a surprise that a loss-of-function in a callose synthase renders a plant more resistant to powdery mildew infection (Jacobs et al., 2003; Nishimura et al., 2003). A comparative analysis of genes that are differentially expressed in wild-type and pmr4 plants upon infection with G. cichoracearum revealed that resistance to powdery mildews is likely the result of constitutive activation of the SA signaling pathway that is further enhanced upon pathogen attack (Nishimura et al., 2003). Mutations in genes encoding components of the salicylic acid signaling pathway indeed abolished pmr4-based resistance without restoring callose deposition at papillae. Callose and/or callose-synthase-based signaling was shown to activate basal defense responses that are effective against a variety of pathogens including G. orontii and H. parasitica (Jacobs et al., 2003; Nishimura et al., 2003). In this sense, PMR4 is likely less a fungal–specific compatibility molecule and more a general basal defense switch located at the cell wall that is effective against a broad range of pathogens.
Two additional pmr mutants stress the potential importance of cell wall integrity surveillance in resistance mechanisms. PMR6 encodes a putative pectate lyase with proposed pectin-degrading activity localized at the cell wall. The pmr6 mutant displays an increased pectin and uronic acid content in the cell wall and exhibits a stunted phenotype (Vogel et al., 2002). In pmr5, similar changes in cell wall components have been reported. PMR5 codes for a member of a large family of plant-specific proteins of unknown function that is likely targeted to the endoplasmic reticulum/secretory pathway (Figure 6). Although the exact mechanism leading to powdery mildew resistance is unknown, it is independent of SA, ET and JA signaling, since mutations in any of the known defense pathways have no effect on pmr5- or pmr6-based resistance. However, pmr5/6 double mutants show further increased resistance compared to the respective single mutants, indicating that the two genes likely control parallel and independent defense responses. Furthermore, the fact that pmr5 and pmr6 are resistant to G. cichoracearum and G. orontii, but fully susceptible to unrelated pathogens such as virulent strains of either P. syringae or H. parasitica, suggests that these two proteins, in contrast to the PMR4 callose synthase gene, may in fact be true powdery mildew-specific compatibility factors (Vogel and Somerville, 2000; Vogel et al., 2002 and 2004).
MLO proteins: A league of their own
Isolated in the same genetic screen as genes PMR4, PMR5 and PMR6, PMR2 encoding MLO2 (MILDEW LOCUS O) has an essential role in the establishment of compatibility with the powdery mildew species G. cichoracearum and G. orontii. Together with the phylogenetically closely related paralogs MLO6 and MLO12, which act in concert with MLO2 in partial functional redundancy (Consonni et al., 2006), MLO2 controls entry of powdery mildew fungi in Arabidopsis epidermal cells (Figure 8). In this respect, MLO2, MLO6 and MLO12 and the barley homologue - and first member of this protein family to be identified - Mlo (Panstruga, 2005), represent the prototype for molecules that mediate compatibility between powdery mildews and their plant hosts. Loss of particular MLO protein(s) in barley, tomato and Arabidopsis render these plant hosts immune to powdery mildew attack (Bai et al., 2008). The conserved function of MLO proteins in plant defense predates the divergence of dicots and monocots and underscores the importance of this protein family in the establishment of compatible plant-powdery mildew interactions (Consonni et al., 2006). MLO proteins are integral membrane proteins with seven transmembrane domains (Figure 6) that have so far only been identified in plants (Devoto et al., 2003). In Arabidopsis they are encoded by a family of 15 genes and appear to have diverse functions in addition to plant defense. However, only the three closely-related members MLO2, MLO6 and MLO12 seem to play a role in powdery mildew compatibility.
Figure 8.

Resistance in Arabidopsis mlo2 mlo6 mlo12 triple mutants is characterized by complete failure of successful host cell invasion.
The micrograph shows attempted penetration of numerous G. orontii sporelings at 48 hours post-inoculation on a highly resistant mlo2 mlo6 mlo12 triple mutant in the genetic background of otherwise susceptible Col-0 (Consonni et al., 2006). Note the aborted fungal entry evidenced by a lack of secondary hyphae compared to the situation in a Col-0 wild type plant (Figure 4B). Scale bar: 100 μm.
Consistent with previous findings in barley (Jarosch et al., 1999, Kumar et al., 2001), mlo2 mlo6 double and mlo2 mlo6 mlo12 triple mutants show enhanced susceptibility to the necrotrophic pathogens Alternaria alternata and A. brassicicola and the hemibiotrophic oomycete Phytophthora infestans, indicating that MLO proteins modulate the infection process of pathogens with diverse lifestyles (Consonni et al., 2006). Barley MLO and MLO2 from Arabidopsis each interact via a conserved peptide domain in their cytoplasmic carboxyl terminus with the Ca2+ sensor calmodulin (Kim et al., 2002; Bhat et al., 2005) and are thought to regulate defense responses against powdery mildews via PEN1 (ROR2 in barley)–dependent mechanisms at the cell periphery (Collins et al., 2003; Schulze-Lefert, 2004, Panstruga, 2005). Similar to non-host resistance, mlo-mediated resistance in A. thaliana does not depend on the JA/ET or SA signaling pathways (Collins et al., 2003; Lipka et al., 2005; Stein et al., 2006; Consonni et al., 2006). The physical proximity of MLO to the PEN1-dependent non-host defense machinery at the sites of fungal attack (Bhat et al., 2005), the recently observed transcriptional co-expression of these genes in Arabidopsis (M. Humphry, unpublished data) and the genetic and cytological similarities between mlo-based and non-host resistance (Humphry et al., 2006) point to a tight mechanistic link between MLO and the basal defense apparatus. The fact that presence of MLO is absolutely required by powdery mildew fungi to successfully infect host plants suggests that these pathogens possibly exploit MLO function(s) to suppress basal defense responses (Panstruga, 2005).
Gene Switches: Transcription factors and gene regulation involved in defense against powdery mildews
In plants, recognition of microbial attack and/or pathogen entry and initiation of defense responses results in a dramatic re-programming of gene expression (Schenk et al., 2000, Zimmerli et al., 2004). Whether mediated by direct interaction between a pathogen receptor and transcription factors (Shen et al., 2007) or, as more often observed in animal systems, by a complex signal transduction cascade (mediated by MAP kinases; Asai et al., 2002), the complete activation of defense responses relies in part on the activity of so-called WRKY transcriptional regulators (reviewed in Eulgem and Somssich, 2007).
In Arabidopsis, the WRKY family is composed of 72 expressed members displaying various degrees of functional redundancy with respect to the diverse biological roles they are involved in, including plant defense. WRKY70 for example, is transcriptionally induced by SA signaling and mediates basal resistance to a variety of pathogens including G. cichoracearum (Li et al., 2006; Wang et al., 2006; Ülker et al., 2007; Figure 6). In contrast, WRKY18 and WRKY40 act as negative regulators of basal defense against G. orontii (Figure 6) and double mutants in these genes are fully resistant to fungal attack (Shen et al., 2007). In the analogous interaction of the monocot barley with its cognate powdery mildew pathogen, Bgh, WRKY18 and 40 counterparts WRKY1 and WRKY2 were shown to interact with the CC domain of MLA immune receptors (Shen et al., 2007). Under normal (pathogen-free) conditions, WRKY1 and WRKY2 are proposed to suppress the transcription of genes involved in basal defense. Upon pathogen challenge and activation by a matching fungal avirulence protein, the MLA receptor is re-localized to the nucleus where it recruits WRKY1 and WRKY2 away from the promoters of defense genes. As expected, silencing of these WRKY transcription factors results in enhanced basal defense against Bgh (Shen et al., 2007, Eckey et al., 2004).
In addition to WRKY transcription factors, the NAC transcription factors, barley NAC6 and its Arabidopsis homolog ATAF1, were recently identified as positive regulators of transcription involved in early defense against powdery mildews. Silencing (barley) or knock out (Arabidopsis) of the respective gene resulted in decreased penetration resistance upon inoculation with Bgh while overexpression (in barley) resulted in increased resistance (Figure 6). These findings suggest a conserved contribution of NAC transcription factors in pre-haustorial defense (Jensen et al., 2007).
A member of another group of transcription factors, the wheat Ethylene Response Factor ERF3, has been also suggested to act as a positive regulator of early defense against powdery mildews (Zhang et al., 2007; Figure 6). ERF3 expression is induced by pathogen challenge (Bgh) and the gene product operates as a GCC box-binding transcriptional activator of SA signaling genes (Zhang et al., 2007). Similarly, heterologous overexpression of other members of this family from tomato in Arabidopsis resulted in enhanced resistance to the adapted powdery mildew, G. orontii, further corroborating their role as positive regulators of transcription in pathogen defense (Gu et al., 2002).
The Ups and Downs of Gene Expression: Transcriptional changes in response to powdery mildew attack
The broad range of transcriptional changes that occur in Arabidopsis during powdery mildew infection (Zimmerli et al., 2004, Fabro et al., 2008) reflect both defense responses mounted by the plant as well as possible manipulation of the host by the fungal pathogen. With the sequencing of the Arabidopsis genome and availability of novel technologies for transcript profiling (oligonucleotide-based microarray chips) it has been possible to peek into the gene expression changes that occur during the infection process. Several dedicated websites such as Genevestigator (https://www.genevestigator.ethz.ch/) and ATGenExpress (http://www.arabidopsis.org/info/expression/ATGenExpress.jsp) provide publicly available collections of microarray data on Arabidopsis challenged with a variety of pathogens including powdery mildews, which represent a largely unexplored treasure of information. However, the spatial resolution of microarray data is limiting, since most microarray experiments are performed on whole leaf samples and as such only provide a gross overview about changes in transcript abundance in pathogen-challenged leaf tissue. Since powdery mildews exclusively colonize the epidermal layer of the leaf, many tissue-specific transcriptional changes are likely to be masked by proportionately larger amounts of mesophyll transcripts. Even within the epidermis, attacked cells and successfully colonized cells each differ in their transcript profile from non-attacked cells (Gjetting et al., 2004 and 2007). Owing to these limitations, expression levels of numerous genes in infected host cells might be considerably under-estimated. Employment of techniques that allow transcript profiling of powdery mildew-challenged Arabidopsis leaves at cellular resolution, e.g. based on RNA extraction via microcapillaries or by laser microdissection (Kehr et al., 2003), is thus urgently required to refine the currently existing data sets.
A previous study analyzing transcriptional changes in Arabidopsis compared the response observed in host (G. cichoracearum) and non-host interactions (Bgh; Zimmerli et al., 2004) at 8, 18 and 24 hours post-inoculation. Surprisingly, many of the differentially regulated genes displayed a similar response in both host and non-host combinations. However, in the non-host interaction, the response was more rapid and of greater amplitude than in the compatible interaction, supporting the hypothesis that a common defense machinery is activated upon attack by adapted and non-adapted pathogens and reinforcing the notion that adapted pathogens are able to partially suppress basal defense in their host species.
Among the genes that exhibit lower transcript abundance after pathogen challenge, a majority encode proteins that are involved in photosynthesis, metabolism, transport and transcription/translation. Consistent with these functions, half of the gene products are either known or predicted to be chloroplast residents. This observation corroborates further studies indicating that Arabidopsis suffers up to 30% reduction in the gain of dry weight following Bgh infections (Zimmerli et al., 2004). This decrease is more modest in compatible infections with G. cichoracearum. An inverse relationship between defense responses and plant growth has been also observed in interactions between cereals and powdery mildews and underscores the metabolic costs associated with activation of defense pathways (Swarbrick et al., 2006; Wright et al., 1995a and b). Intriguingly, a recent microarray analysis of the compatible interaction between Arabidopsis and G. cichoracearum, revealed that host photosynthesis is likely stimulated by the fungus, through the induction of genes involved in chlorophyll binding and chlorophyll a oxygenase (Fabro et al., 2008). Though it is conceivable that an increase in photosynthetic rate is required, as the metabolic demands of the infected epidermal tissue increase, these findings are somewhat contradictory to those obtained in the study of Zimmerli et al. (2004). Manipulation of the host metabolism by the fungal parasite thus likely involves upas well as downregulation of photosynthetic genes.
However, many gene transcripts related to defense responses accumulate upon both host and non-host powdery mildew attack (e.g. transcripts encoding glycosyl hydrolases and β-1,3-glucanase). Consistently, many of the respective genes contain in their up-stream promoter region the cis-regulatory element OCS, which is known to be responsive to auxin, SA and hydrogen peroxide. Transcripts specifically accumulating during non-host interactions encode defensins and derive from genes that are regulated by JA and ethylene. SA signaling therefore does not appear to play an important role in non-host defense against Bgh. Alternatively, SA- controlled genes may only become activated once the penetration defenses have failed.
In a separate experiment, analysis of the transcriptional response of Arabidopsis leaves to chitin (the main fungal cell wall polysaccharide) and chitin derivatives identified several genes involved in defense responses (Ramonell et al., 2002). Genes encoding plant-derived chitinases exhibit higher transcript accumulation upon pathogen challenge compared to control conditions and chitin oligomers obtained as fungal cell wall degradation products constitute potent signaling molecules responsible for induced resistance to G. cichoracearum. This resistance response appears to be activated through signaling routes that are distinct from the SA, JA and ET pathways (Ramonell et al., 2002). Genes encoding PR5, two WRKY transcription factors and several disease resistance proteins belonging to the TIR-NB-LRR family were induced upon chitin treatment (Ramonell et al., 2005). Genes with reduced transcript abundance included ETR2, encoding a trans-membrane protein kinase with a LRR domain, the defensin PDF2.5, RAR1 (encoding a protein with two zinc-finger-like domains that is required for the accumulation of many R proteins) and several other genes coding for zinc-finger proteins. To functionally validate the microarray experiments, T-DNA insertion mutants within seven of the differentially expressed genes were tested for susceptibility to G. cichoracearum. Three of the seven mutants, encoding a putative RING zinc-finger and two putative disease resistance proteins, displayed enhanced susceptibility to the pathogen, further reinforcing the role of chitin sensing and signaling in plant defense responses (Ramonell et al., 2005).
The critical role played by chitin perception in plant defense responses was recently strengthened by the cloning of the presumptive Arabidopsis chitin (co-)receptor LysM Receptor-Like Kinase1 gene (LysM RLK1 or CERK1) by two independent groups (Miya et al., 2007, Wan et al., 2008). The gene codes for a plasma membrane-resident protein with a single transmembrane domain, three extracellular LysM domains and a cytoplasmic serine/threo-nine kinase domain (Figure 6). Mutations in this gene abolish virtually all changes in gene transcription normally observed upon chitin perception by the plant, without affecting other elicitor perception such as flagellin sensing, suggesting that the protein is part of the chitin receptor complex (Wan et al., 2008). It is conceivable that in analogy to the FLS2 receptor kinase, which is responsible for flagellin-triggered signaling (Asai et al., 2002), LysM RLK1/CERK feeds into MAP kinase signaling and defense gene activation (Figure 6). Interestingly, although loss of LysM RLK1 caused enhanced susceptibility to the host fungus G. cichoracearum at the post-penetration stage (Wan et al., 2008), it did not affect the plant response to the adapted hemibiotrophic fungus Colletotrichum higginsianum and resulted in only a marginal increase in susceptibility to the non-adapted necrotrophic fungus Alternaria brassicicola. These observations suggest that pathogen perception by the plant is microbe–specific and implies the existence of more elicitor-specific receptors yet to be discovered.
In addition to genes involved in defense responses, genes that are induced because they are likely required for pathogen growth have also been identified (Fotopoulos et al., 2003). The observation that Arabidopsis leaves infected with G. cichoracearum have an enhanced sugar uptake compared to non-infected leaves prompted an investigation of sugar transporter expression upon powdery mildew challenge. G. cichoracearum induces expression of STP4, encoding a glucose transporter as well as βfruct1, coding for a cell wall invertase, in several cell types of the leaf (most notably the epidermis and vascular bundle) irrespective of whether they are penetrated by the fungus or not. This transcriptional induction is sustained for up to 6 days post-inoculation and is not reproducible in a non-host interaction with Bgh. A similar role for sugar transporters in fungal development was also found in the interaction between the rust fungus Uromyces fabae and broad bean (Vicia faba). However, in this case, the hexose transporter is of fungal origin and resides in the haustorial membrane (Voegele et al., 2001), suggesting that exchange of nutrients at the plant/biotroph interface requires transport facilitators on both sides. It is not surprising that sugars, and therefore sugar biosynthesis and transport, would be important for fungal growth and development and enhanced by successful fungal pathogens. The mechanisms leading to this genetic and physiological manipulation remain an enigma. However, new efforts into powdery mildew studies promise to lift the veil on the mystery.
Through the Looking Glass: Powdery mildew biology
A large body of knowledge on plant compatibility and resistance to powdery mildews has considerably advanced our understanding of these diseases. However, our lack of information about many functional aspects of the biology of powdery mildew fungi has created a plant-centric view of this pathosystem. At the present time, we obtain biological information about the fungus predominantly by indirectly looking in the reflection provided by Arabidopsis research. Although the image of this “mirror” is valuable it may sometimes be skewed and it certainly is incomplete. The difficulties hampering direct powdery mildew research stem mainly from their obligate biotrophic lifestyle. The fungi can not be maintained on artificial media and their vegetative spores, at least in the case of the four species pathogenic on Arabidopsis, can not be stored frozen or lyophilized (see long-term storage methods for some powdery mildew species reported in Bardin et al., 2007). Although spores germinate on artificial hydrophobic substrates, growth does not progress beyond the formation of an appressorium. In addition to these hurdles that prevent any forward genetic screens for pathogenicity mutants, reproducible genetic manipulation of these pathogens is not feasible at present.
Despite these difficulties, genomic and transcript analysis methods have recently been used to crack the code of powdery mildew genes and provide a glimpse into their biology. Several expressed sequence tags (ESTs) and microarray analyses have been performed on Bgh (Both et al., 2005a and b) and the genome of this cereal pathogen has been sequenced and is currently being annotated by a scientific consortium in the U.K. (P. Spanu, S. Gurr, N. Talbot, J. Brown, C. Ridout and co-workers, personal communication; see also the BluGen database at http://www.blugen.org/). Several thousand ESTs and data from one development-specific microarray analysis of Bgh transcripts are publicly accessible (http://cogeme.ex.ac.uk/index.html; Soanes and Talbot, 2006; Soanes et al., 2002; Both et al., 2005a). In addition, the genomes of G. orontii and E. pisi are currently being sequenced by next generation high-throughput DNA sequencing techniques (pyrosequencing) at the Max-Planck Institute for Plant Breeding Research (C. Micali, S. Noir, M. Benjdia, K. Stüber, P. Schulze-Lefert and R. Panstruga, unpublished data). This effort should provide a starting point to the exploration of powdery mildew biology. Among the first cloned genes from a powdery mildew species that is able to colonize A. thaliana, a putative ATPase transporter gene (PMA1) from G. cichoracearum was shown to be induced starting at 3 days and until 6 days post-inoculation on Arabidopsis leaves (Fotopoulos et al., 2006). The complete lack of expression of the gene at 1 day post-inoculation may suggest its selective expression in haustoria, which are not abundant at that early infection stage. A similar induction of a plasma membrane H+ ATPase reported for the barley pathogen Bgh further supports a role for ATPase pumps in the establishment of a proton potential that can drive the transport of sugars and other nutrients to the fungal haustorium (Both et al., 2005a).
In addition to genes involved in basic fungal housekeeping functions such as nutrition and development, genes that play a direct role in pathogenicity begin to emerge, as more and more fungal pathogen genomes and transcriptomes are being analyzed. The isolation and characterization of bacterial, fungal and oomycete effector and avirulence protein coding genes has opened a new chapter in plant-microbe interactions and has reshaped the way we think about disease and disease resistance (see Bent and Mackie, 2007 for a review of the subject). Although no genes directly involved in virulence have so far been identified in any of the powdery mildew fungi colonizing Arabidopsis, more than 25 avirulence genes and their cognate host R genes are known for Bgh.
Recently, the isolation and characterization of the first avirulence genes, AVRk1 and AVRa10, from Bgh has provided an exciting starting point for further research in the field (Ridout et al., 2006). AVRk1 and its paralogue AVRa10 are tightly-linked avirulence genes and encode proteins recognized by the barley resistance genes Mlk1 and Mla10, respectively. Using conventional AFLP marker segregation in two crosses between virulent and avirulent Bgh strains, the two genes were mapped within a 7.5 kb. interval interspersed with several repetitive sequences. AVRk1 is a 422 amino acid protein with no homology to any known proteins in the databases. A premature stop codon at amino acid 14 in this protein renders it undetectable by the MLk1 R protein and is responsible for virulence of certain Bgh strains on Mlk1 plants. Overexpression of either AVRk1 or AVRa10 in barley epidermal cells causes cell death that is dependent on the presence of the cognate R protein, MLk1 or MLa10, respectively. In addition, overexpression of the same genes enhances penetration by Bgh in the absence of the cognate R protein, indicating a role for AVRk1 and AVRa10 in disease establishment. AVRk1 and AVRa10 appear to be part of a large gene family, with at least five additional expressed paralogs (based on a survey of the current Bgh EST database) and more than 30 genomic copies. The close proximity of the genes to retrotransposon sequences suggests a possible mechanism of gene amplification and genome-wide dissemination. In addition, the multicopy nature of AVRk1 and AVRa10 provides the possibility for mutation and escape from host detection, without the evolutionary cost associated with the loss of a virulence gene.
Do AVRk1 and AVRa10 represent typical effector/avirulence protein encoding genes in powdery mildew species? Preliminary analysis suggests the existence of AVRk1-like sequences in the genomes of E. pisi and G. orontii, also nested within or in close proximity to retrotransposable elements (C. Micali and R. Panstruga, unpublished results). Whether these sequences are transcribed and encode bona fide genes, and whether these genes play a role in pathogenesis are among the many questions that remain to be answered. Based on our current knowledge of effector-encoding genes from other fungal and oomycete pathogens, there appears to be no prototypical protein effector structure. For example, a canonical N-terminal signal peptide for passage through the secretory pathway, a hallmark of nearly all characterized fungal effectors (Bent and Mackie, 2007), is lacking in AVRk1, AVRa10 and its Bgh, E. pisi and G. orontii homologues.
The genetic and molecular study of powdery mildew fungi is still in its infancy. The contribution from genome sequencing efforts will greatly enhance our understanding of these fungi. In the case of powdery mildew fungi, a comparative analysis of the genomes of Bgh, E. pisi and G. orontii, will provide a precious glimpse into their virulence arsenal. In addition, ultrastructural and biochemical studies of the haustoria will likely provide valuable information about the nutrient uptake and host manipulation strategies of these pathogens. Further studies on the lifestyle, sexual reproduction and population structure of powdery mildew species in nature will help to bridge the gap in knowledge we currently have about these fungi and their interaction and disease-causing mechanisms in plants.
Footnotes
Citation: Micali C., Göllner K., Humphry M., Consonni C., and Panstruga R. (2008) The Powdery Mildew Disease of Arabidopsis: A Paradigm for the Interaction between Plants and Biotrophic Fungi. The Arabidopsis Book 6:e0115. doi:10.1199/tab.0115
elocation-id: e0115
Published on: October 2, 2008
Present address: University of Aachen, Department for Biology III, Worringer Weg 1, 52056 Aachen, Germany
References
- Adam L., Ellwood S., Wilson I., Saenz G., Xiao S., Oliver R. P., Turner J. G., Somerville S. Comparison of Erysiphe cichoracearum and E. cruciferarum and a survey of 360 Arabidopsis thaliana accessions for resistance to these two powdery mildew pathogens. Mol. Plant-Microbe Interact. 1999;126(1):1031–1043. doi: 10.1094/MPMI.1999.12.12.1031. [DOI] [PubMed] [Google Scholar]
- Adam L., Somerville S. C. Genetic characterization of five powdery mildew disease resistance loci in Arabidopsis thaliana. Plant J. 1996;96(1):341–356. doi: 10.1046/j.1365-313x.1996.09030341.x. [DOI] [PubMed] [Google Scholar]
- Aist J. R. Papillae and related wound plugs of plant cells. Ann. Rev. Phytopathol. 1976;146(1):145–163. [Google Scholar]
- Aist J. R., Bushnell W. R. Invasion of plants by powdery mildew fungi, and cellular mechanisms of resistance. In: Cole G. T., Hoch H. C., editors. The fungal spore and disease initiation in plants and animals. 1. Vol. 6. Plenum press; New York: 1991. pp. 321–345. [Google Scholar]
- Asai T., Tena G., Plotnikova J., Willmann M. R., Chiu W. L., Gomez-Gomez L., Boller T., Ausubel F. M., Sheen J. MAP kinase signalling cascade in Arabidopsis innate immunity. Nature. 2002;4156(1):977–983. doi: 10.1038/415977a. [DOI] [PubMed] [Google Scholar]
- Assaad F. F., Qiu J. L., Youngs H., Ehrhardt D., Zimmerli L., Kalde M., Wanner G., Peck S. C., Edwards H., Ramonell K., Somerville C. R., Thordal-Christensen H. The PEN1 syntaxin defines a novel cellular compartment upon fungal attack and is required for the timely assembly of papillae. Mol. Biol. Cell. 2004;156(1):5118–5129. doi: 10.1091/mbc.E04-02-0140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ausubel F. M. Are innate immune signaling pathways in plants and animals conserved. Nature Immunol. 2005;66(1):973–979. doi: 10.1038/ni1253. [DOI] [PubMed] [Google Scholar]
- Bai Y. L., Pavan S., Zheng Z., Zappel N. F., Reinstädler A., Lotti C., De Giovanni C., Ricciardi L., Lindhout P., Visser R., Theres K., Panstruga R. Naturally occurring broad-spectrum powdery mildew resistance in a Central American tomato accession is caused by loss of Mlo function. Mol. Plant-Microbe Interact. 2008;216(1):30–39. doi: 10.1094/MPMI-21-1-0030. [DOI] [PubMed] [Google Scholar]
- Bai Y. L., van der Hulst R., Bonnema G., Marcel B. C., Meijer-Dekens F., Niks R. E., and Lindhout P. Tomato defense to Oidium neolycopersici: Dominant Ol genes confer isolate-dependent resistance via a different mechanism than recessive ol-2. Mol. Plant–Microbe Interact. 2005;186(1):354–362. doi: 10.1094/MPMI-18-0354. [DOI] [PubMed] [Google Scholar]
- Bardin M., Suliman E. M., Sage-Palloix A., Mohamed Y. F., Nicot P. C. Inoculum production and long-term conservation methods for cucurbits and tomato powdery mildews. 2007. Mycol. Res. 2007;1116(1):740–747. doi: 10.1016/j.mycres.2007.03.015. [DOI] [PubMed] [Google Scholar]
- Barker C. L., Donald T., Pauquet J., Ratnaparkhe M. B., Bouquet A., Adam-Blondon A. F., Thomas M. B., Dry I. Genetic and physical mapping of the grapevine powdery mildew resistance gene, Run1, using a bacterial artificial chromosome library. Theoret. Appl. Genet. 2005;1116(1):370–377. doi: 10.1007/s00122-005-2030-8. [DOI] [PubMed] [Google Scholar]
- Bent A. F., Mackey D. Elicitors, Effectors and R Genes: The new paradigm and a lifetime supply of questions. Annu. Rev. Phytopathol. 2007;456(1):399–436. doi: 10.1146/annurev.phyto.45.062806.094427. [DOI] [PubMed] [Google Scholar]
- Bhat R. A., Miklis M., Schmelzer E., Schulze-Lefert P., Panstruga R. Recruitment and interaction dynamics of plant penetration resistance components in a plasma membrane microdomain. Proc. Natl. Acad. USA. 2005;1026(1):3135–3140. doi: 10.1073/pnas.0500012102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Both M., Csukai M., Stumpf M. P. H., Spanu P. D. Gene expression profiles of Blumeria graminis indicate dynamic changes to primary metabolism during development of an obligate biotrophic pathogen. Plant Cell. 2005a;176(1):2107–2122. doi: 10.1105/tpc.105.032631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Both M., Eckert S. E., Csukai M., Müller E., Dimopoulos G., Spanu P. D. Transcript profiles of Blumeria graminis development reveal a cluster of genes that are potential virulence determinants. Mol. Plant-Microbe Interact. 2005b;186(1):125–133. doi: 10.1094/MPMI-18-0125. [DOI] [PubMed] [Google Scholar]
- Bracker C. E. Ultrastructure of the haustorial apparatus of Erysiphe graminis and its relationship to the epidermal cell of barley. Phytopathology. 1968;586(1):12–30. [Google Scholar]
- Braun U., Cook R. T. A., Inman A. J., and Shin H. D. The taxonomy of the powdery mildews. In: Belanger R. R., Bushnell W. R., Dik A. J., Carver T. L. W., editors. The Powdery Mildews, a comprehensive treatise. 1. Vol. 6. The American Phytopathological Society Press; St. Paul: 2002. pp. 56–65. [Google Scholar]
- Collins N. C., Thordal-Christensen H., Lipka V., Bau S., Kombrink E., Qiu J. L., Hückelhoven R., Stein M., Freialdenhoven A., Somerville S. C., Schulze-Lefert P. SNARE-protein-mediated disease resistance at the plant cell wall. Nature. 2003;4256(1):973–977. doi: 10.1038/nature02076. [DOI] [PubMed] [Google Scholar]
- Consonni C., Humphry M. E., Hartmann H. A., Livaja M., Durner J., Westphal L., Vogel J., Lipka V., Kemmerling B., Schulze-Lefert P., Somerville S. C., Panstruga R. Conserved requirement for a plant host cell protein in powdery mildew pathogenesis. Nat. Genet. 2006;386(1):716–720. doi: 10.1038/ng1806. [DOI] [PubMed] [Google Scholar]
- Devoto A., Piffanelli P., Elliott C., Simmons C., Taramino G., Goh C. S., Cohen F. E., Emerson B. C., Schulze-Lefert P., Panstruga R. Molecular phylogeny and evolution of the plant-specific seven-transmembrane MLO family. J. Mol. Evol. 2003;566(1):77–88. doi: 10.1007/s00239-002-2382-5. [DOI] [PubMed] [Google Scholar]
- Dodds P. N., Lawrence G. J., Catanzariti A. M., Ayliffe M. A., Ellis J. G. The Melampsora lini AvrL567 avirulence genes are expressed in haustoria and their products are recognized inside plant cells. Plant Cell. 2004;166(1):755–768. doi: 10.1105/tpc.020040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eckey C., Korell M., Leib K., Biedenkopf D., Jansen C., Langen G., Kogel K. H. Identification of powdery mildew-induced barley genes by cDNA-AFLP: functional assessment of an early expressed MAP kinase. Plant Mol. Biol. 2004;556(1):1–15. doi: 10.1007/s11103-004-0275-2. [DOI] [PubMed] [Google Scholar]
- Ellis C., Karafyllidis I., Turner J. G. Constitutive activation of jasmonate signaling in an Arabidopsis mutant correlates with enhanced resistance to Erysiphe cichoracearum, Pseudomonas syringae, and Myzus persicae. Mol Plant-Microbe Interact. 2002a;156(1):1025–1030. doi: 10.1094/MPMI.2002.15.10.1025. [DOI] [PubMed] [Google Scholar]
- Ellis C., Karafyllidis I., Wasternack C., Turner J. G. The Ara-bidopsis mutant cev1 links cell wall signaling to jasmonate and ethylene responses. Plant Cell. 2002b;146(1):1557–1566. doi: 10.1105/tpc.002022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellis C., Turner J. G. The Arabidopsis mutant cev1 has constitutively active jasmonate and ethylene signal pathways and enhanced resistance to pathogens. Plant Cell. 2001;136(1):1025–1033. doi: 10.1105/tpc.13.5.1025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eulgem T., Somssich I. E. Networks of WRKY transcription factors in defense signaling. Curr. Opin. Plant Biol. 2007;106(1):366–371. doi: 10.1016/j.pbi.2007.04.020. [DOI] [PubMed] [Google Scholar]
- Fabro G., Di Rienzo J. A., Voigt C. A., Sevchenko T., Dehesh K., Somerville S., Alvarez M. E. Genome-wide expression profiling Arabidopsis at the stage of Golovinomyces cichoracearum haustorium formation. Plant Physiol. 2008;1466(1):1421–1439. doi: 10.1104/pp.107.111286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fauteux F., Chain F., Belzile F., Menzies J. G., Belanger R. R. The protective role of silicon in the Arabidopsis-powdery mildew pathosystem. Proc. Natl. Acad. USA. 2006;1036(1):17554–17559. doi: 10.1073/pnas.0606330103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Felix G., Duran J. D., Volko S., Boller T. Plants have a sensitive perception system for the most conserved domain of bacterial flagellin. Plant J. 1999;186(1):265–276. doi: 10.1046/j.1365-313x.1999.00265.x. [DOI] [PubMed] [Google Scholar]
- Feys B. J., Parker J. E. Interplay of signaling pathways in plant disease resistance. Trends Genet. 2000;166(1):449–455. doi: 10.1016/s0168-9525(00)02107-7. [DOI] [PubMed] [Google Scholar]
- Feys B. J., Wiermer M., Bhat R. A., Moisan L. J., Medina-Escobar N., Neu C., Cabral A., Parker J. E. Arabidopsis SENESCENCE-ASSOCIATED GENE101 stabilizes and signals within an ENHANCED DISEASE SUSCEPTIBILITY1 complex in plant innate immunity. Plant Cell. 2005;176(1):2601–2613. doi: 10.1105/tpc.105.033910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fotopoulos V., Gilbert M. J., Pittman J. K., Marvier A. C., Buchanan A. J., Sauer N., Hall J. L., Williams L. E. The monosaccharide transporter gene, AtSTP4, and the cell-wall invertase, Atβfruct1, are induced in Arabidopsis during infection with the fungal biotroph Erysiphe cichoracearum. Plant Physiol. 2003;1326(1):821–829. doi: 10.1104/pp.103.021428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fotopoulos V., Holmes R., Hall J. L., Williams L. E. Isolation, cloning and expression analysis of EcPMA1, a putative plasma membrane H+ -ATPase transporter gene from the biotrophic pathogenic fungus Erysiphe cichoracearum. Mycol. Res. 2006;1106(1):28–37. doi: 10.1016/j.mycres.2005.08.011. [DOI] [PubMed] [Google Scholar]
- Frye C. A., Innes R. W. An Arabidopsis mutant with enhanced resistance to powdery mildew. Plant Cell. 1998;106(1):947–956. doi: 10.1105/tpc.10.6.947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frye C. A., Tang D. Z., Innes R. W. Negative regulation of defense responses in plants by a conserved MAPKK kinase. Proc. Natl. Acad. Sci. USA. 2001;986(1):373–378. doi: 10.1073/pnas.98.1.373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghanmi D., McNally D. J., Benhamou N., Menzies J. G., Belanger R. R. Powdery mildew of Arabidopsis thaliana: a pathosystem for exploring the role of silicon in plant-microbe interactions. Physiol. Mol. Plant Pathol. 2004;646(1):189–199. [Google Scholar]
- Gil F., Gay J. L. Ultrastructural and physiological properties of host interfacial components of haustoria of Erysiphe pisi in vivo and in vitro. Physiol. Plant Pathol. 1977;106(1):1–12. [Google Scholar]
- Gjetting T., Carver T. L., Skøt L., Lyngkjaer M. F. Differential gene expression in individual papilla-resistant and powdery mildew-infected barley epidermal cells. Mol. Plant Microbe Interact. 2004;176(1):729–738. doi: 10.1094/MPMI.2004.17.7.729. [DOI] [PubMed] [Google Scholar]
- Gjetting T., Hagedorn P. H., Schweizer P., Thordal-Christensen H., Carver T. L., Lyngkjaer M. F. Single-cell transcript profiling of barley attacked by the powdery mildew fungus. Mol. Plant Microbe Interact. 2007;206(1):235–246. doi: 10.1094/MPMI-20-3-0235. [DOI] [PubMed] [Google Scholar]
- Glazebrook J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 2005;436(1):205–227. doi: 10.1146/annurev.phyto.43.040204.135923. [DOI] [PubMed] [Google Scholar]
- Göllner K., Schweizer P., Bai Y., Panstruga R. Natural genetic resources of Arabidopsis thaliana reveal a high prevalence and unexpected phenotypic plasticity of RPW8-mediated powdery mildew resistance. New Phytol. 2008;1776(1):725–742. doi: 10.1111/j.1469-8137.2007.02339.x. [DOI] [PubMed] [Google Scholar]
- Gu Y. Q., Wildermuth M. C., Chakravarthy S., Loh Y. T., Yang C., He X., Han Y., Martin G. B. Tomato transcription factors Pti4, Pti5, and Pti6 activate defense responses when expressed in Arabidopsis. Plant Cell. 2002;146(1):817–831. doi: 10.1105/tpc.000794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gupta V., Willits M. G., Glazebrook J. Arabidopsis thaliana EDS4 contributes to salicylic acid (SA)-dependent expression of defense responses: evidence for inhibition of jasmonic acid signaling by SA. Mol. Plant-Microbe Interact. 2000;136(1):503–511. doi: 10.1094/MPMI.2000.13.5.503. [DOI] [PubMed] [Google Scholar]
- Gus-Meyer S., Naton B., Hahlbrock K., Schmelzer E. Local mechanical stimulation induces components of the pathogen defense response in parsley. Proc. Natl. Acad. Sci. USA. 1998;146(1):8398–8403. doi: 10.1073/pnas.95.14.8398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hahn M., Neef U., Struck C., Göttfert M., Mendgen K. A putative amino acid transporter is specifically expressed in haustoria of the rust fungus Uromyces fabae. Mol. Plant-Microbe Interact. 1997;106(1):438–445. doi: 10.1094/MPMI.1997.10.4.438. [DOI] [PubMed] [Google Scholar]
- Halterman D., Zhou F. S., Wei F. S., Wise R. P., Schulze-Lefert P. The MLA6 coiled-coil, NBS-LRR protein confers AvrMla6-dependent resistance specificity to Blumeria graminis f. sp. hordei in barley and wheat. Plant J. 2001;256(1):335–348. doi: 10.1046/j.1365-313x.2001.00982.x. [DOI] [PubMed] [Google Scholar]
- Hardham A., Jones D. A., Takemoto D. Cytoskeleton and cell wall function in penetration resistance. Curr. Opin. Plant Biol. 2007;106(1):342–348. doi: 10.1016/j.pbi.2007.05.001. [DOI] [PubMed] [Google Scholar]
- Hiscott J., Rongtuan L., Peyman N., Paz S. MasterCARD: A priceless link to innate Immunity. Trends Mol. Med. 2006;126(1):53–56. doi: 10.1016/j.molmed.2005.12.003. [DOI] [PubMed] [Google Scholar]
- Hückelhoven R. Cell wall-associated mechanisms of disease resistance and susceptibility. Annu. Rev. Phytopathol. 2007;456(1):101–127. doi: 10.1146/annurev.phyto.45.062806.094325. [DOI] [PubMed] [Google Scholar]
- Humphry M., Consonni C., Panstruga R. mlo-based powdery-mildew immunity: Silver bullet or simply non-host resistance Mol. Plant Pathol. 2006;76(1):605–610. doi: 10.1111/j.1364-3703.2006.00362.x. [DOI] [PubMed] [Google Scholar]
- Jacobs A. K., Lipka V., Burton R. A., Panstruga R., Strizhov N., Schulze-Lefert P., Fincher G. B. An Arabidopsis callose synthase, GSL5, is required for wound and papillary callose formation. Plant Cell. 2003;156(1):2503–2513. doi: 10.1105/tpc.016097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jarosch B., Kogel K. H., Schaffrath U. The ambivalence of the barley Mlo locus: Mutations conferring resistance against powdery mildew (Blumeria graminis f. sp. hordei) enhance susceptibility to the rice blast fungus Magnaporthe grisea. Mol. Plant-Microbe Interact. 1999;126(1):508–514. [Google Scholar]
- Jensen M. K., Rung J. H., Gregersen P. L., Gjetting T., Fuglsang A. T., Hansen M., Joehnk N., Lyngkjaer M. F., Collinge D. B. The HvNAC6 transcription factor: a positive regulator of penetration resistance in barley and Arabidopsis. Plant Mol. Biol. 2007;656(1):137–150. doi: 10.1007/s11103-007-9204-5. [DOI] [PubMed] [Google Scholar]
- Jones H., Whipps J. M., Gurr S. J. The tomato powdery mildew fungus Oidium neolycopersici. Mol. Plant Pathol. 2001;26(1):303–309. doi: 10.1046/j.1464-6722.2001.00084.x. [DOI] [PubMed] [Google Scholar]
- Jørgensen J. H. Genetics of powdery mildew resistance in barley. Crit. Rev. Plant Sci. 1994;136(1):97–119. [Google Scholar]
- Keeble J. A., Gilmore A. P. Apoptosis commitment-translating survival signals into decisions on mitochondria. Cell Res. 2007;176(1):976–984. doi: 10.1038/cr.2007.101. [DOI] [PubMed] [Google Scholar]
- Kehr J. Single cell technology. Curr. Opin. Plant Biol. 2003;66(1):617–621. doi: 10.1016/j.pbi.2003.09.002. [DOI] [PubMed] [Google Scholar]
- Kim M. C., Panstruga R., Elliott C., Müller J., Devoto A., Yoon H. W., Park H. C., Cho M. J., Schulze-Lefert P. Calmodulin interacts with MLO protein to regulate defence against mildew in barley. Nature. 2002;4166(1):447–451. doi: 10.1038/416447a. [DOI] [PubMed] [Google Scholar]
- Kobayashi Y., Kobayashi I., Funaki Y., Fujimoto S., Takemoto T., Kunoh H. Dynamic reorganization of microfilaments and microtubules is necessary for the expression of non-host resistance in barley coleoptile cells. Plant J. 1997a;116(1):525–537. [Google Scholar]
- Kobayashi Y., Yamada M., Kobayashi I., Kunoh H. Actin microfilaments are required for the expression of nonhost resistance in higher plants. Plant Cell. Physiol. 1997b;386(1):725–733. [Google Scholar]
- Koch E., Slusarenko A. J. Fungal pathogens of Arabidopsis thaliana (L.) Heyhn. Bot. Helv. 1990;1006(1):257–268. [Google Scholar]
- Koh S., Andre A., Edwards H., Ehrhardt D., Somerville S. Arabidopsis thaliana subcellular responses to compatible Erysiphe cichoracearum infections. Plant J. 2005;446(1):516–529. doi: 10.1111/j.1365-313X.2005.02545.x. [DOI] [PubMed] [Google Scholar]
- Kumar J., Hückelhoven R., Beckhove U., Nagarajan S., Kogel K. H. A compromised Mlo pathway affects the response of barley to the necrotrophic fungus Bipolaris sorokiniana (teleomorph: Cochliobolus sativus) and its toxins. Phytopathol. 2001;916(1):127–133. doi: 10.1094/PHYTO.2001.91.2.127. [DOI] [PubMed] [Google Scholar]
- Kwon C., Neu C., Pajonk S., Yun H. S., Lipka U., Humphry M., Bau S., Straus M., Kwaaitaal M., Rampelt H., El Kasmi F., Jürgens G., Parker J., Panstruga R., Lipka V., Schulze-Lefert P. Co-option of a default secretory pathway for plant immune responses. Nature. 2008a;4516(1):835–840. doi: 10.1038/nature06545. [DOI] [PubMed] [Google Scholar]
- Kwon C., Panstruga R., Schulze-Lefert P. Les liaisons dangereuses: immunological synapse formation in animals and plants. Trends Immunol. 2008b;296(1):159–166. doi: 10.1016/j.it.2008.01.004. [DOI] [PubMed] [Google Scholar]
- Leonelli S. Arabidopsis, the botanical Drosophila: from mouse cress to model organism. Endeavour. 2007;316(1):34–38. doi: 10.1016/j.endeavour.2007.01.003. [DOI] [PubMed] [Google Scholar]
- Li C., Bonnema G., Che D., Dong L., Lindhout P., Visser R., Bai Y. Biochemical and molecular mechanisms involved in monogenic resistance responses to tomato powdery mildew. Mol. Plant-Microbe Interact. 2007;206(1):1161–72. doi: 10.1094/MPMI-20-9-1161. [DOI] [PubMed] [Google Scholar]
- Li J., Brader G., Kariola T., Palva E. T. WRKY70 modulates the selection of signaling pathways in plant defense. Plant J. 2006;466(1):477–491. doi: 10.1111/j.1365-313X.2006.02712.x. [DOI] [PubMed] [Google Scholar]
- Lipka V., Dittgen J., Bednarek P., Bhat R., Wiermer M., Stein M., Landtag J., Brandt W., Rosahl S., Scheel D., Llorente F., Molina A., Parker J., Somerville S., Schulze-Lefert P. Pre- and postinvasion defenses both contribute to nonhost resistance in Arabidopsis. Science. 2005;3106(1):1180–1183. doi: 10.1126/science.1119409. [DOI] [PubMed] [Google Scholar]
- Lipka V., Kwon C., Panstruga R. SNARE-ware: the role of SNARE-domain proteins in plant biology. Annu. Rev. Cell Dev. Biol. 2007;236(1):147–174. doi: 10.1146/annurev.cellbio.23.090506.123529. [DOI] [PubMed] [Google Scholar]
- Liu G., Holub E. B., Alonso J. M., Ecker J. R., Fobert P. R. An Arabidopsis NPR1-like gene, NPR4, is required for disease resistance. Plant J. 2005;416(1):304–318. doi: 10.1111/j.1365-313X.2004.02296.x. [DOI] [PubMed] [Google Scholar]
- Loake G., Grant M. Salicylic acid in plant defense: The players and protagonists. Curr. Opin. Plant Biol. 2007;106(1):466–472. doi: 10.1016/j.pbi.2007.08.008. [DOI] [PubMed] [Google Scholar]
- Martin G. B., Bogdanove A. J., Sessa G. Understanding the functions of plant disease resistance proteins. Annu. Rev. Plant Biol. 2003;546(1):23–61. doi: 10.1146/annurev.arplant.54.031902.135035. [DOI] [PubMed] [Google Scholar]
- Mendgen K., Hahn M. Plant infection and the establishment of fungal biotrophy. Trends Plant Sci. 2002;76(1):352–357. doi: 10.1016/s1360-1385(02)02297-5. [DOI] [PubMed] [Google Scholar]
- Meyerowitz E. M. Prehistory and history of Arabidopsis research. Plant Physiol. 2001;1256(1):15–19. doi: 10.1104/pp.125.1.15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miya A., Albert P., Shinya T., Desaki Y., Ichimura K., Shirasu K., Narusaka Y., Kawakami N., Kaku H., Shibuya N. CERK1, a LysM receptor kinase, is essential for chitin elicitor signaling in Arabidopsis. Proc. Natl. Acad. Sci. USA. 2007;1046(1):19613–19618. doi: 10.1073/pnas.0705147104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miklis M., Consonni C., Bhat R. A., Lipka V., Schulze-Lefert P., Panstruga R. Barley MLO modulates actin-dependent and actin-independent antifungal defense pathways at the cell periphery. Plant Physiol. 2007;1446(1):1132–1143. doi: 10.1104/pp.107.098897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Navarro L., Dunoyer P., Jay F., Arnold B., Dharmasiri N., Estelle M., Voinnet O., Jones J. D. A plant miRNA contributes to antibacterial resistance by repressing auxin signaling. Science. 2006;3126(1):436–439. doi: 10.1126/science.1126088. [DOI] [PubMed] [Google Scholar]
- Nishimura M. T., Stein M., Hou B. H., Vogel J. P., Edwards H., Somerville S. C. Loss of a callose synthase results in salicylic acid-dependent disease resistance. Science. 2003;3016(1):969–972. doi: 10.1126/science.1086716. [DOI] [PubMed] [Google Scholar]
- O'Connell R. J., Panstruga R. Tête à tête inside a plant cell: establishing compatibility between plants and biotrophic fungi and oomycetes. New Phytol. 2006;1716(1):699–718. doi: 10.1111/j.1469-8137.2006.01829.x. [DOI] [PubMed] [Google Scholar]
- Opalski K. S., Schultheiss H., Kogel K. H., Hückelhoven R. The receptor-like MLO protein and the RAC/ROP family G-protein RACB modulate actin reorganization in barley attacked by the biotrophic powdery mildew fungus Blumeria graminis f.sp. hordei. Plant J. 2005;416(1):291–303. doi: 10.1111/j.1365-313X.2004.02292.x. [DOI] [PubMed] [Google Scholar]
- Orgil U., Araki H., Tangchaiburana S., Berkey R., Xiao S. Intraspecific genetic variations, fitness cost and benefit of RPW8, a disease resistance locus in Arabidopsis thaliana. Genetics. 2007;1766(1):2317–2333. doi: 10.1534/genetics.107.070565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Panstruga R. Establishing compatibility between plants and obligate biotrophic pathogens. Curr. Opin. Plant Biol. 2003;66(1):320–326. doi: 10.1016/s1369-5266(03)00043-8. [DOI] [PubMed] [Google Scholar]
- Panstruga R. Serpentine plant MLO proteins as entry portals for powdery mildew fungi. Biochem. Soc. Trans. 2005;336(1):389–392. doi: 10.1042/BST0330389. [DOI] [PubMed] [Google Scholar]
- Parker J. E., Holub E. B., Frost L. N., Falk A., Gunn N. D., Daniels M. J. Characterization of eds1, a mutation in Arabidopsis suppressing resistance to Peronospora parasitica specified by several different RPP genes. Plant Cell. 1996;86(1):2033–2046. doi: 10.1105/tpc.8.11.2033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plotnikova J. M., Reuber T. L., Ausubel F. M. Powdery mildew pathogenesis on Arabidopsis thaliana. Mycologia. 1998;906(1):1009–1016. [Google Scholar]
- Ramonell K., Berrocal-Lobo M., Koh S., Wan J., Edwards H., Stacey G., Somerville S. Loss-of-Function mutations in chitin responsive genes show increased susceptibility to the powdery mildew pathogen Erysiphe cichoracearum. Plant Physiol. 2005;1386(1):1027–1036. doi: 10.1104/pp.105.060947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramonell K. M., Zhang B., Ewing R. M., Chen Y., Xu D., Stacey G., Somerville S. Microarray analysis of chitin elicitation in Arabidopsis thaliana. Mol. Plant Pathol. 2002;36(1):301–311. doi: 10.1046/j.1364-3703.2002.00123.x. [DOI] [PubMed] [Google Scholar]
- Ratcliff F., Harrison B. D., Baulcombe D. C. A similarity between viral defense and gene silencing in plants. Science. 1997;2766(1):1558–1560. doi: 10.1126/science.276.5318.1558. [DOI] [PubMed] [Google Scholar]
- Reuber T. L., Plotnikova J. M., Dewdney J., Rogers E. E., Wood W., Ausubel F. M. Correlation of defense gene induction defects with powdery mildew susceptibility in Arabidopsis enhanced disease susceptibility mutants. Plant J. 1998;166(1):473–485. doi: 10.1046/j.1365-313x.1998.00319.x. [DOI] [PubMed] [Google Scholar]
- Ridout C., Skamnioti P., Porritt O., Sacristan S., Jones J. D., Brown J. K. Multiple avirulence paralogues in cereal powdery mildew fungi may contribute to parasite fitness and defeat of plant resistance. Plant Cell. 2006;186(1):2402–2414. doi: 10.1105/tpc.106.043307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robatzek S., Chinchilla D., Boller T. Ligand-induced endocytosis of the pattern recognition receptor FLS2 in Arabidopsis. Genes Dev. 2006;206(1):537–542. doi: 10.1101/gad.366506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russo V. M., Bushnell W. R. Responses of barley cells to puncture by microneedles and to attempted penetration by Erysiphe graminis f. sp. hordei. Can. J. Bot. 1989;676(1):2912–2921. [Google Scholar]
- Schenk P. M., Kazan K., Wilson I., Anderson J. P., Richmond T., Somerville S. C., Manners J. M. Coordinated plant defense responses in Arabidopsis revealed by microarray analysis. Proc. Natl. Acad. Sci. USA. 2000;976(1):11655–11660. doi: 10.1073/pnas.97.21.11655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schiff C. L., Wilson I. W., Somerville S. C. Polygenic powdery mildew disease resistance in Arabidopsis thaliana: Quantitative trait analysis of the accession Warschau-1. Plant Pathol. 2001;506(1):690–701. doi: 10.1093/genetics/158.3.1301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmidt S. M., Panstruga R. Cytoskeleton functions in plant-microbe interactions. Physiol. Mol. Plant Pathol. 2007;716(1):135–148. [Google Scholar]
- Schulze-Lefert P. Knocking on heaven's wall: pathogenesis of and resistance to biotrophic fungi at the cell wall. Curr Opin. Plant Biol. 2004;76(1):377–383. doi: 10.1016/j.pbi.2004.05.004. [DOI] [PubMed] [Google Scholar]
- Schulze-Lefert P., Panstruga R. Establishment of biotrophy by parasitic fungi and reprogramming of host cells for disease resistance. Annu. Rev. Phytopathol. 2003;416(1):641–467. doi: 10.1146/annurev.phyto.41.061002.083300. [DOI] [PubMed] [Google Scholar]
- Schulze-Lefert P., Vogel J. Closing the ranks to attack by powdery mildew. Trends Plant Sci. 2000;56(1):343–348. doi: 10.1016/s1360-1385(00)01683-6. [DOI] [PubMed] [Google Scholar]
- Shen Q. H., Saijo Y., Mauch S., Biskup C., Bieri S., Keller B., Seki H., Ülker B., Somssich I. E., Schulze-Lefert P. Nuclear activity of MLA immune receptors links isolate-specific and basal disease-resistance responses. Science. 2007;3156(1):1098–1103. doi: 10.1126/science.1136372. [DOI] [PubMed] [Google Scholar]
- Shimada C., Lipka V., O'Connell R., Okuno T., Schulze-Lefert P., Takano Y. Nonhost resistance in Arabidopsis-Colletotrichum interactions acts at the cell periphery and requires actin filament function. Mol. Plant-Microbe Interact. 2006;196(1):270–279. doi: 10.1094/MPMI-19-0270. [DOI] [PubMed] [Google Scholar]
- Shirasu K., Nielsen K., Piffanelli P., Oliver R., Schulze-Lefert P. Cell-autonomous complementation of mlo resistance using a biolistic transient expression system. Plant J. 1999;176(1):293–299. [Google Scholar]
- Skula A., Berglund L., Nielsen L. P., Nielsen S., Hoffmann H. J., Dahl R. Regulated exocytosis in immune function: are SNARE-proteins involved? Respir. Med. 2000;946(1):10–17. doi: 10.1053/rmed.1999.0700. [DOI] [PubMed] [Google Scholar]
- Soanes D. M., Skinner W., Keon J., Hargreaves J., Talbot N. J. Genomics of phytopathogenic fungi and the development of bioinformatic resources. Mol Plant-Microbe Interact. 2002;156(1):421–427. doi: 10.1094/MPMI.2002.15.5.421. [DOI] [PubMed] [Google Scholar]
- Soanes D. M., Talbot N. J. Comparative genomic analysis of phytopathogenic fungi using expressed sequence tag (EST) collections. Mol. Plant Pathol. 2006;76(1):61–70. doi: 10.1111/j.1364-3703.2005.00317.x. [DOI] [PubMed] [Google Scholar]
- Spencer-Phillips P. T. N., Gay J. L. Domains of ATPase in plasma membrane and transport through infected plant cells. New Phytol. 1981;896(1):393–400. [Google Scholar]
- Stein M., Dittgen J., Sánchez-Rodríguez C., Hou B. H., Molina A., Schulze-Lefert P., Lipka V., Somerville S. Arabidopsis PEN3/PDR8, an ATP binding cassette transporter, contributes to non-host resistance to inappropriate pathogens that enter by direct penetration. Plant Cell. 2006;186(1):731–746. doi: 10.1105/tpc.105.038372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strack D., Fester T., Hause B., Schliemann W., Walter M. H. Arbuscular mycorrhiza: Biological, chemical, and molecular aspects. J. Chem. Ecol. 2003;296(1):1955–1979. doi: 10.1023/a:1025695032113. [DOI] [PubMed] [Google Scholar]
- Swarbrick P. J., Schulze-Lefert P., Scholes J. D. Metabolic consequences of susceptibility and resistance (race-specific and broad-spectrum) in barley leaves challenged with powdery mildew. Plant Cell Environ. 2006;296(1):1061–1076. doi: 10.1111/j.1365-3040.2005.01472.x. [DOI] [PubMed] [Google Scholar]
- Takamatsu S. Phylogeny and evolution of the powdery mildew fungi (Erysiphales, Ascomycota) inferred from nuclear ribosomal DNA sequences. Mycoscience. 2004;456(1):147–157. [Google Scholar]
- Takemoto D., Jones D., Hardham A. Re-organisation of the cytoskeleton and endoplasmic reticulum in the Arabidopsis pen1-1 mutant inoculated with the non-adapted powdery mildew Blumeria graminis f. sp. hordei. Mol. Plant Pathol. 2006;76(1):553–563. doi: 10.1111/j.1364-3703.2006.00360.x. [DOI] [PubMed] [Google Scholar]
- Tang D., Ade J., Frye C. A., Innes R. W. A mutation in the GTP hydrolysis site of Arabidopsis dynamin-related protein 1E confers enhanced cell death in response to powdery mildew infection. Plant J. 2006;476(1):75–84. doi: 10.1111/j.1365-313X.2006.02769.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tang D., Ade J., Frye C. A., Innes R. W. Regulation of plant defense responses in Arabidopsis by EDR2, a PH and START domain-containing protein. Plant J. 2005a;446(1):245–257. doi: 10.1111/j.1365-313X.2005.02523.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tang D., Christiansen K. M., Innes R. W. Regulation of plant disease resistance, stress responses, cell death and ethylene signaling in Arabidopsis by the EDR1 protein kinase. Plant Physiol. 2005b;1386(1):1018–1026. doi: 10.1104/pp.105.060400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tang D., Innes R. W. Overexpression of a kinase-deficient form of the EDR1 gene enhances powdery mildew resistance and ethylene-induced senescence in Arabidopsis. Plant J. 2002;326(1):975–983. doi: 10.1046/j.1365-313x.2002.01482.x. [DOI] [PubMed] [Google Scholar]
- Thordal-Christensen H. Fresh insights into processes of nonhost resistance. Curr. Opin. Plant Biol. 2003;66(1):351–357. doi: 10.1016/s1369-5266(03)00063-3. [DOI] [PubMed] [Google Scholar]
- Thordal-Christensen H., Zhang Z. G., Wei Y. D., Collinge D. B. Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley- powdery mildew interaction. Plant J. 1997;116(1):1187–1194. [Google Scholar]
- Timonen S., Drew E., Smith S. E. Effect of cytoskeletal inhibitors on mycorrhizal colonisation of tomato roots. Symbiosis. 2006;416(1):81–86. [Google Scholar]
- Ton J., Van Pelt J. A., Van Loon L. C., Pieterse C. M. Differential effectiveness of salicylate-dependent and jasmonate/ethylene-dependent induced resistance in Arabidopsis. Mol. Plant-Microbe Interact. 2002;156(1):27–34. doi: 10.1094/MPMI.2002.15.1.27. [DOI] [PubMed] [Google Scholar]
- Tucker S. L., Talbot N. J. Surface attachment and pre-penetration stage development by plant pathogenic fungi. Annu. Rev. Phytopathol. 2001;396(1):385–417. doi: 10.1146/annurev.phyto.39.1.385. [DOI] [PubMed] [Google Scholar]
- Ülker B., Shahid Mukhtar M., Somssich I. E. The WRKY70 transcription factor of Arabidopsis influences both the plant senescence and defense signaling pathways. Planta. 2007;2266(1):125–137. doi: 10.1007/s00425-006-0474-y. [DOI] [PubMed] [Google Scholar]
- Voegele R. T., Struck C., Hahn M., Mendgen K. The role of haustoria in sugar supply during infection of broad bean by the rust fungus Uromyces fabae. Proc. Natl. Acad. Sci. USA. 2001;986(1):8133–8138. doi: 10.1073/pnas.131186798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vogel J. P., Raab T. K., Schiff C., Somerville S. C. PMR6, a pectate lyase-like gene required for powdery mildew susceptibility in Arabidopsis. Plant Cell. 2002;146(1):2095–2106. doi: 10.1105/tpc.003509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vogel J. P., Raab T. K., Somerville C. R., Somerville S. C. Mutations in PMR5 result in powdery mildew resistance and altered cell wall composition. Plant J. 2004;406(1):968–978. doi: 10.1111/j.1365-313X.2004.02264.x. [DOI] [PubMed] [Google Scholar]
- Vogel J., Somerville S. Powdery mildew of Arabidopsis: a model system for host-parasite interactions. In: Belanger R. R., Bushnell W. R., Dik A. J., Carver T. L. W., editors. The Powdery Mildews, a comprehensive treatise. 1. Vol. 6. The American Phytopathological Society Press; St. Paul: 2002. pp. 161–168. [Google Scholar]
- Vogel J., Somerville S. Isolation and characterisation of powdery mildew resistant Arabidopsis mutants. Proc. Natl. Acad. USA. 2000;976(1):1897–1902. doi: 10.1073/pnas.030531997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wan J. R., Zhang X. C., Neece D., Ramonell K. M., Clough S., Kim S. Y., Stacey M. G., Stacey G. A LysM receptor-like kinase plays a critical role in chitin signaling and fungal resistance in Arabidopsis. Plant Cell. 2008;206(1):471–481. doi: 10.1105/tpc.107.056754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang D., Amornsiripanitch N., Dong X. A genomic approach to identify regulatory nodes in the transcriptional network of systemic acquired resistance in plants. PLoS Pathogens. 2006;26(1):e123. doi: 10.1371/journal.ppat.0020123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang W., Devoto A., Turner J. G., Xiao S. Expression of the membrane-associated resistance protein RPW8 enhances basal defense against biotrophic pathogens. Mol. Plant-Microbe Interact. 2007;206(1):966–976. doi: 10.1094/MPMI-20-8-0966. [DOI] [PubMed] [Google Scholar]
- Whipps J. M., Budge S. P., Fenlon J. S. Characteristics and host range of tomato powdery mildew. Plant Pathol. 1998;476(1):36–48. [Google Scholar]
- Wilson I. W., Schiff C. L., Hughes D. E., Somerville S. C. Quantitative trait loci analysis of powdery mildew disease resistance in the Arabidopsis thaliana accession Kashmir-1. Genetics. 2001;1586(1):1301–1309. doi: 10.1093/genetics/158.3.1301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wright D. P., Baldwin B. C., Shephard M. C., Scholes J. D. Source-sink relationships in wheat leaves infected with powdery mildew. I. Alterations in carbohydrate metabolism. Physiol. Mol. Plant Pathol. 1995a;476(1):237–253. [Google Scholar]
- Wright D. P., Baldwin B. C., Shephard M. C., Scholes J. D. Source-sink relationships in wheat leaves infected with powdery mildew. II. Changes in the regulation of the Calvin cycle. Physiol. Mol. Plant Pathol. 1995b;476(1):255–267. [Google Scholar]
- Xiao S. Y., Calis O., Patrick E., Zhang G. G., Charoenwattana P., Muskett P., Parker J. E., Turner J. G. The atypical resistance gene, RPW8, recruits components of basal defence for powdery mildew resistance in Arabidopsis. Plant J. 2005;426(1):95–110. doi: 10.1111/j.1365-313X.2005.02356.x. [DOI] [PubMed] [Google Scholar]
- Xiao S. Y., Charoenwattana P., Holcombe L., Turner J. G. The Arabidopsis genes RPW8.1 and RPW8.2 confer induced resistance to powdery mildew diseases in tobacco. Mol. Plant-Microbe Interact. 2003;166(1):289–294. doi: 10.1094/MPMI.2003.16.4.289. [DOI] [PubMed] [Google Scholar]
- Xiao S. Y., Ellwood S., Calis O., Patrick E., Li T. X., Coleman M., Turner J. G. Broad-spectrum mildew resistance in Arabidopsis thaliana mediated by RPW8. Science. 2001;2916(1):118–120. doi: 10.1126/science.291.5501.118. [DOI] [PubMed] [Google Scholar]
- Xiao S. Y., Ellwood S., Findlay K., Oliver R. P., Turner J. G. Characterization of three loci controlling resistance of Arabidopsis thaliana accession Ms-0 to two powdery mildew diseases. Plant J. 1997;126(1):757–768. doi: 10.1046/j.1365-313x.1997.12040757.x. [DOI] [PubMed] [Google Scholar]
- Xiao S. Y., Emerson B., Ratanasut K., Patrick E., O'Neill C., Bancroft I., Turner J. G. Origin and maintenance of a broad-spectrum disease resistance locus in Arabidopsis. Mol. Biol. Evol. 2004;216(1):1661–1672. doi: 10.1093/molbev/msh165. [DOI] [PubMed] [Google Scholar]
- Yun B. W., Atkinson H. A., Gaborit C., Greenland A., Read N. D., Pallas J. A., Loake G. J. Loss of actin cytoskeletal function and EDS1 activity, in combination, severely compromises non-host resistance in Arabidopsis against wheat powdery mildew. Plant J. 2003;346(1):768–777. doi: 10.1046/j.1365-313x.2003.01773.x. [DOI] [PubMed] [Google Scholar]
- Zeyen R. J., Carver T. L. W., Lyngkjaer M. F. Epidermal cell papillae. The Powdery Mildews, a comprehensive treatise. In: Belanger R. R., Bushnell W. R., Dik A. J., Carver T. L. W., editors. 1. Vol. 6. The American Phytopathological Society Press; St. Paul: 2002. pp. 107–125. [Google Scholar]
- Zhang Z., Yao W., Dong N., Liang H., Liu H., Huang R. A novel ERF transcription activator in wheat and its induction kinetics after pathogen and hormone treatments. J. Exp. Bot. 2007;586(1):2993–3003. doi: 10.1093/jxb/erm151. [DOI] [PubMed] [Google Scholar]
- Zimmerli L., Stein M., Lipka V., Schulze-Lefert P., Somerville S. Host and non-host pathogens elicit different jasmonate/ethylene responses in Arabidopsis. Plant J. 2004;406(1):633–646. doi: 10.1111/j.1365-313X.2004.02236.x. [DOI] [PubMed] [Google Scholar]