

Abstract
Background
Infection by dengue virus (DENV) is a major public health concern in hundreds of tropical and subtropical countries. French Polynesia (FP) regularly experiences epidemics that initiate, or are consecutive to, DENV circulation in other South Pacific Island Countries (SPICs). In January 2009, after a decade of serotype 1 (DENV-1) circulation, the first cases of DENV-4 infection were reported in FP. Two months later a new epidemic emerged, occurring about 20 years after the previous circulation of DENV-4 in FP. In this study, we investigated the epidemiological and molecular characteristics of the introduction, spread and genetic microevolution of DENV-4 in FP.
Methodology/Principal Findings
Epidemiological data suggested that recent transmission of DENV-4 in FP started in the Leeward Islands and this serotype quickly displaced DENV-1 throughout FP. Phylogenetic analyses of the nucleotide sequences of the envelope (E) gene of 64 DENV-4 strains collected in FP in the 1980s and in 2009–2010, and some additional strains from other SPICs showed that DENV-4 strains from the SPICs were distributed into genotypes IIa and IIb. Recent FP strains were distributed into two clusters, each comprising viruses from other but distinct SPICs, suggesting that emergence of DENV-4 in FP in 2009 resulted from multiple introductions. Otherwise, we observed that almost all strains collected in the SPICs in the 1980s exhibit an amino acid (aa) substitution V287I within domain I of the E protein, and all recent South Pacific strains exhibit a T365I substitution within domain III.
Conclusions/Significance
This study confirmed the cyclic re-emergence and displacement of DENV serotypes in FP. Otherwise, our results showed that specific aa substitutions on the E protein were present on all DENV-4 strains circulating in SPICs. These substitutions probably acquired and subsequently conserved could reflect a founder effect to be associated with epidemiological, geographical, eco-biological and social specificities in SPICs.
Introduction
Almost all tropical and subtropical regions of the world are concerned by the risk of dengue. Every year, dengue virus (DENV) causes more than 50 million infections, 500 000 hospitalizations and 12 500 deaths, mostly children [1]. Currently there is no vaccine or effective anti-viral therapy and vector control measures regularly fail to prevent the emergence of dengue epidemics. DENV is a single-stranded, positive-sense RNA virus belonging to the genus Flavivirus, family Flaviviridae, transmitted by Aedes mosquitoes, principally Aedes (Stegomyia) aegypti but also Aedes albopictus and some endemic vectors, like Aedes polynesiensis in the Polynesian triangle [2]. Infection with dengue virus may result in a wide spectrum of clinical manifestations accounting for the classification of the disease as dengue (fever, rash, headache, myalgia, arthralgia, nausea, vomiting) with or without warning signs (abdominal pain, persistent vomiting, clinical fluid accumulation, mucosal bleed) or severe dengue (characterized by plasma leakage) with or without hemorrhage [3]. There are four distinct serotypes of dengue virus (DENV-1 to -4) and infection with one serotype provides no long-term cross-protective immunity against the three others. Based on the sequence of the envelope gene (E), each serotype may be divided into distinct phylogenetic clusters or genotypes [4]. Both epidemiological observations and in vitro studies suggest that some genotypes may have different epidemic potential [5]–[7].
In the South Pacific, the earliest dengue epidemics may have occurred during the nineteenth century [8]. However, dengue only became a significant health problem during World War II, because of the spread of the main dengue vector, Ae. aegypti. The first documented outbreak of DENV in the South Pacific occurred in 1943–1944 and was due to serotype 1. When dengue reappeared 20 years later in French Polynesia (FP) it was due to DENV-3 [9]–[10]. The first cases of severe dengue in the Pacific were reported in the 1970s [11]–[13]. Beginning in 2000, outbreaks of DENV in South Pacific Island Countries (SPICs) have been due to DENV-1 [14]–[19] but in 2008, DENV-4 circulation was detected [20]–[22] for the first time since the 1980's [23], [24]. In January 2009, about 20 years after the last reported case, DENV-4 was detected in two members from the same family returning to FP from New Caledonia (NC) where this serotype had recently been detected. Two months later FP experienced a large dengue outbreak.
Here, we report the epidemiological and molecular characteristics of the introduction, spread and genetic microevolution of DENV-4 in FP.
Materials and Methods
Epidemiological and biological data
Laboratory diagnosis of dengue was performed on sera from dengue suspected patients using ELISA tests for the detection of either DENV NS1 (Platelia™ Dengue NS1 Ag, Bio-Rad Laboratories, Marnes-la-Coquette, France) or anti-DENV IgM (Dengue IgM Capture ELISA, Panbio Diagnostics, Brisbane, Australia). A DENV serotype-specific multiplex real-time RT-PCR using primers and probes designed by Johnson et al. (2005) [25] was performed on sera containing DENV NS1 or from patients who have travelled recently to areas where dengue is known to occur. Data from January 2009 to December 2010 were recorded by the Health Department (Bureau de Veille Sanitaire, Tahiti, French Polynesia) based on reports transmitted by Institut Louis Malardé (ILM, Tahiti, French Polynesia) and the principal hospital in Tahiti (Centre Hospitalier de la Polynésie Française).
Viruses
The details of the viruses employed are shown in Table 1. Briefly, DENV-4 strains collected in FP in the 1980s were obtained from the DENV collection of ILM. At the time they were collected, these strains were amplified by one passage on C6/36 Ae. albopictus cells and cell supernatants were then preserved at −80°C. DENV-4 strains recently collected in FP were obtained from sera positive by DENV-4 real-time RT-PCR performed by the clinical laboratory from ILM. One of these sera was recovered from a patient recently returned from NC [PF-NC09/160109-136]. Two additional DENV-4 strains collected in NC and Wallis & Futuna [NC09/060209-1528, WF09/010409-0001] were kindly provided by Dr M Dupont-Rouzeyrol, Institut Pasteur de Nouvelle Calédonie, and Dr JF Yvon, clinical laboratory from Sia Hospital, Wallis. Five additional strains collected in the SPICs in 2008–2009 [Kiribati KI08/266, Tonga TO08/14, Fiji FJ08/3953, Vanuatu VU09/4214 and Western Samoa WS08/73] were sequenced as previously described [22]. Additional E gene sequences of 73 strains were retrieved from GenBank.
Table 1. Characteristics of the 64 DENV-4 strains used for E-gene sequence analysis.
| ID | Country | Archipelago | Island | District | Year | GenBank |
| NC09/060209-1528 | NC | 2009 | JN832498 | |||
| WF09/010409-0001 | WF | 2009 | JN832499 | |||
| PF-NC09/160109-136 | PF (imp. from NC) | Tahiti | Mahina | 2009 | JN832507 | |
| PF82/5918 | PF | Windward Islands | Tahiti | 1982 | JN832500 | |
| PF83/6074 | Tahiti | 1983 | JN832501 | |||
| PF84/131-188 | Tahiti | 1984 | JN832502 | |||
| PF85/323-103 | Tahiti | 1985 | JN832503 | |||
| PF86/50-70 | Tahiti | 1986 | JN832504 | |||
| PF87/19-7 | Tahiti | 1987 | JN832505 | |||
| PF09/060309-96 | Tahiti | Papeari | 2009 | JN832514 | ||
| PF09/130309-109 | Tahiti | Punaauia | 2009 | JN832519 | ||
| PF09/160309-24 | Tahiti | Mahina | 2009 | JN832520 | ||
| PF09/230309-126 | Tahiti | Punaauia | 2009 | JN832521 | ||
| PF09/310309-162 | Tahiti | Pirae | 2009 | JN832524 | ||
| PF09/270409-182 | Tahiti | Faaa | 2009 | JN832530 | ||
| PF09/150609-91* | Tahiti | Punaauia | 2009 | JN832536 | ||
| PF09/220709-54* | Tahiti | Punaauia | 2009 | JN832541 | ||
| PF09/220909-50 | Tahiti | Papeete | 2009 | JN832545 | ||
| PF09/141009-111 | Tahiti | Papeete | 2009 | JN832546 | ||
| PF09/041109-131 | Tahiti | Papara | 2009 | JN832547 | ||
| PF09/041209-32 | Tahiti | Arue | 2009 | JN832548 | ||
| PF10/260110-67 | Tahiti | Punaauia | 2010 | JN832551 | ||
| PF10/050210-117 | Tahiti | Punaauia | 2010 | JN832552 | ||
| PF10/120310-31 | Tahiti | Papeete | 2010 | JN832554 | ||
| PF10/010410-215 | Tahiti | Mahina | 2010 | JN832555 | ||
| PF10/060410-11 | Tahiti | Papeete | 2010 | JN832556 | ||
| PF10/300410-48 | Tahiti | Punaauia | 2010 | JN832557 | ||
| PF10/170510-92 | Tahiti | 2010 | JN832558 | |||
| PF10/170510-191 | Tahiti | Punaauia | 2010 | JN832559 | ||
| PF10/170510-192 | Tahiti | Punaauia | 2010 | JN832560 | ||
| PF10/150610-28 | Tahiti | Papara | 2010 | JN832561 | ||
| PF09/020309-54 | Moorea | 2009 | JN832512 | |||
| PF09/190809-58 | Moorea | 2009 | JN832543 | |||
| PF10/190110-98 | Moorea | 2010 | JN832550 | |||
| PF10/010310-32 | Moorea | 2010 | JN832553 | |||
| PF09/090309-122 | Leeward Islands | Raiatea | 2009 | JN832516 | ||
| PF09/080409-93 | Raiatea | 2009 | JN832526 | |||
| PF09/040509-242 | Raiatea | 2009 | JN832531 | |||
| PF09/120609-125 | Raiatea | 2009 | JN832535 | |||
| PF09/270709-74 | Raiatea | 2009 | JN832542 | |||
| PF09/230209-116 | Bora Bora | 2009 | JN832509 | |||
| PF09/100309-172 | Bora Bora | 2009 | JN832517 | |||
| PF09/100309-208 | Bora Bora | 2009 | JN832518 | |||
| PF09/190509-238 | Bora Bora | 2009 | JN832533 | |||
| PF09/200809-131 | Bora Bora | 2009 | JN832544 | |||
| PF09/290109-69 | Taha'a | 2009 | JN832508 | |||
| PF09/270209-213 | Taha'a | 2009 | JN832511 | |||
| PF09/030309-39 | Taha'a | 2009 | JN832513 | |||
| PF09/060309-198 | Taha'a | 2009 | JN832515 | |||
| PF09/210409-134 | Maupiti | 2009 | JN832527 | |||
| PF09/190509-37 | Maupiti | 2009 | JN832532 | |||
| PF09/160609-119 | Maupiti | 2009 | JN832537 | |||
| PF88/130-74 | Marquesas archipelago | 1988 | JN832506 | |||
| PF09/240209-38 | Ua Pou | 2009 | JN832510 | |||
| PF09/270309-27 | Ua Pou | 2009 | JN832522 | |||
| PF09/270309-31 | Ua Pou | 2009 | JN832523 | |||
| PF09/081209-283 | UaPou | 2009 | JN832549 | |||
| PF09/290509-25 | Nuku Hiva | 2009 | JN832534 | |||
| PF09/070409-227 | Australes archipelago | Rimatara | 2009 | JN832525 | ||
| PF09/230409-06 | Rurutu | 2009 | JN832528 | |||
| PF09/250609-14 | Rurutu | 2009 | JN832538 | |||
| PF09/240409-72 | Tuamotu archipelago | Hao | 2009 | JN832529 | ||
| PF09/210709-11 | Hao | 2009 | JN832540 | |||
| PF09/080709-23 | Manihi | 2009 | JN832539 |
*patient with severe dengue.
Ethics statement
All dengue virus strains were obtained from sera initially sampled for dengue diagnostic and surveillance purposes (under medical prescription by a physician), and archived at ILM. The only information provided to the research laboratory was the sample laboratory ID number, the date of collection and the district/island of collection (no personal information on the patient was provided). The use of biological samples and the collection of information were performed in accordance with the French regulations.
Nucleotide sequencing
Sera or whole blood on filter paper cards (LDA22, France) were used as a source of DENV. RNA was extracted from sera using the QIAamp® Viral RNA Mini Kit (Qiagen, Germany) or the Easymag extraction system (Biomérieux, France) according to manufacturers' instructions. For whole blood, five filter paper spots per patient were incubated for 20 min at room temperature in lysis buffer from the QIAamp® Viral RNA Mini Kit. After centrifugation for 10 min at 8000 rpm, the extraction process was pursued using the supernatant, according to the manufacturer's instructions. Amplification of the E gene was then performed using the OneStep RT-PCR® Kit (Qiagen, Germany) and two manually designed oligonucleotides primer pairs [D4E/777F (5′- GCT TGG AAA CAT GCT CAG AG-3′) and D4E/1766R (5′- ACA TGT GGT TTC CAT CAC CG-3′); D4E/1639F (5′-TGG TGA CAT TCA AGG TTC CTC-3′) and D4E/2509R (5′-ACT GTT CTG TCC AAG TGT GC-3′)] in order to produce two overlapping fragments covering the complete E gene. For two sequences [PF09/270309-27, PF09/270309-31], a second round of PCR was required to obtain sufficient DNA. The PCR products were purified using the QIAquick PCR Purification Kit (Qiagen, Germany), the subsequent sequencing reactions performed with the ABI PRISM® BigDye™ Terminator Cycle Sequencing Ready Reaction Kit (Applied Biosystems, USA). and the product analyzed on the ABI Prism 310 automated sequencer (Perkin-Elmer, Applied Biosystems). Overlapping nucleotide sequences were assembled with AutoAssembler software (Perkin Elmer). Nucleotide sequences were aligned with the multiple sequence alignment software ClustalW integrated in MEGA version 5 [26]. Phylogenetic analysis was performed with MEGA5 using the Maximum likelihood based on the Kimura 2-parameter method [27]. The robustness of the original tree was tested with 1000 bootstrap replications [28]. Sequences available on GenBank and corresponding to strains belonging to all major branches of previously published DENV-4 phylogenies were included in the analysis and selected to maximize the representation of genotypes, countries and years of isolation. Those that were very closely related to one another in the data set were removed from the analysis. Mean distances between and within groups were calculated for all sequences (MEGA5).
Results
Epidemiology
Two patients with DENV-4 infections were detected in January 2009 in Tahiti, Society Islands. The patients were members from the same family recently returned from NC where a DENV-4 outbreak had recently commenced concomitantly with DENV-1 transmission. These were the first cases of infection with DENV-4 detected in FP for more than 20 years. The French Polynesia Public Health and Hygiene Department immediately applied vector control measures by spraying insecticide against adult mosquitoes and destroying breeding sites around the home of these patients. The insecticide treatment was repeated 10 days later. No new DENV-4 cases were detected in Tahiti in the following weeks. However, at the end of February, DENV-4 was detected in three patients living in Taha'a, five in Bora-Bora, one in Tahiti (Society Islands) and one in Ua Pou (Marquesas Islands). None of these patients had traveled out from FP during the two previous weeks. An additional patient from Taha'a with an onset of symptoms at the end of January was found subsequently, by RT-PCR, to also have had a DENV-4 infection. From January to March 2009, the number of patients with DENV-4 infections increased dramatically and DENV-4 had displaced DENV-1 3 months later (Table 2). From January 2009 to December 2010, ILM reported 8375 presumptive cases of dengue to the Health Department, of which 2629 were confirmed subsequently by serological or virological testing. One hundred of these were due to DENV-1 and 763 to DENV-4 infections (Figure 1). During this period, 87 hospitalizations were reported of which three were dengue severe cases.
Table 2. Spatial and temporal distribution of confirmed cases of DENV-1 and DENV-4 infections between January and December 2009.
| Society archipelago | Marquesas archipelago | Austral archipelago | Tuamotu archipelago | Gambier archipelago | |||
| Total | Windward Islands | Leeward Islands | |||||
| January | |||||||
| DENV-1 | 11 | Tahiti 11 | |||||
| DENV-4 | 3 | Tahiti 2 | Tahaa 1 | ||||
| February | |||||||
| DENV-1 | 29 | Tahiti 29 | |||||
| DENV-4 | 10 | Tahiti 1 | Tahaa 3 | Ua Pou 1 | |||
| Bora Bora 5 | |||||||
| March | |||||||
| DENV-1 | 39 | Tahiti 37 | |||||
| Moorea 2 | |||||||
| DENV-4 | 132 | Tahiti 41 | Tahaa 42 | Ua Pou 3 | |||
| Moorea 3 | Bora Bora 33 | ||||||
| Raiatea 10 | |||||||
| April | |||||||
| DENV-1 | 20 | Tahiti 17 | Raiatea 1 | ||||
| Moorea 2 | |||||||
| DENV-4 | 262 | Tahiti 130 | Tahaa 14 | Ua Pou 8 | Rimatara 1 | Hao 2 | |
| Moorea 25 | Bora Bora 55 | Nuku-Hiva 1 | Rurutu 2 | ||||
| Raiatea 21 | Fatu-Hiva 1 | ||||||
| Maupiti 2 | |||||||
| May | |||||||
| DENV-1 | 2 | Tahiti 2 | |||||
| DENV-4 | 126 | Tahiti 80 | Bora Bora 9 | Ua Pou 4 | Tubuai 1 | ||
| Moorea 10 | Raiatea 9 | Nuku-Hiva 6 | |||||
| Maupiti 4 | Ua Huka 1 | ||||||
| Huahine 2 | |||||||
| June | |||||||
| DENV-1 | 1 | Tahiti 1 | |||||
| DENV-4 | 165 | Tahiti 112 | Bora Bora 4 | Nuku-Hiva 1 | Rurutu 6 | Hao 4 | |
| Moorea 13 | Raiatea 15 | Raivavae 2 | Fakarava 3 | ||||
| Maupiti 2 | Manihi 1 | ||||||
| Huahine 1 | Rangiroa 1 | ||||||
| July | |||||||
| DENV-4 | 35 | Tahiti 23 | Raiatea 1 | Fatu-Hiva 1 | Hao 1 | ||
| Moorea 4 | Huahine 1 | Manihi 4 | |||||
| August | |||||||
| DENV-4 | 7 | Tahiti 3 | Bora Bora 1 | Hao 1 | |||
| Moorea 2 | |||||||
| September | |||||||
| DENV-4 | 9 | Tahiti 5 | Nuku-Hiva 2 | Mangareva 1 | |||
| Hiva-Oa 1 | |||||||
| October | |||||||
| DENV-4 | 5 | Tahiti 4 | Rangiroa 1 | ||||
| November | |||||||
| DENV-4 | 2 | Tahiti 2 | |||||
| December | |||||||
| DENV-4 | 7 | Tahiti 6 | Ua Pou 1 | ||||
Figure 1. Monthly recorded dengue suspected and confirmed cases from January 2009 to December 2010.

Molecular epidemiology of DENV-4 in FP
The 64 E gene sequences generated in the present study were combined with 5 E gene sequences from DENV-4 strains collected in the South Pacific between 2008 and 2009 and a data set of 73 E gene sequences available on GenBank representing the global genetic variability of DENV-4 (Figure 2). The phylogenetic tree shows four distinct clusters named genotype I, II, III and IV. These four clusters have been previously described as genotype I or Southeast Asia, II or Indonesia, III, and IV or Sylvatic or Malaysia [6], [29]–[31]. All sequences of DENV-4 collected in the South Pacific belonged to genotype II. The average distances between genotype II and the three others varied from 0.073 to 0.156, with standard errors (SE) of 0.005 and 0.011 respectively. Within genotype II, viruses clustered into two distinct clades previously defined as IIa and IIb [32]. The average distance between these two clades is 0.063 with a SE of 0.007. The Clade IIb consisted of strains isolated from 1979 to 2008 in the South Pacific, the Caribbean, South and Central America, along with two strains isolated in Indonesia in 1976 and 1977. Within clade IIb, viruses collected in FP from 1983 to 1988 formed a distinct group supported by a bootstrap value of 99% which we have called the PF80s group. Clade IIa was composed, almost exclusively, of viruses collected in the South Pacific and Southeast Asia since 2000. Within clade IIa, all DENV-4 strains collected in the South Pacific from 2007 to 2010 formed a distinct group supported by a bootstrap value of 100% and which we have named the Oceania group. Within the Oceania group, the strains collected in FP were distributed into two clusters (Figure 3). The first one, PF/NC/VU, comprised most (83%) of the FP strains and was close to the strains from NC and Vanuatu. The second cluster, PF/WF, included nine FP strains and was closer to the strain from Wallis & Futuna. Within each cluster, several FP strains shared identical E gene consensus sequences.
Figure 2. Evolutionary relationships of DENV-4 E gene sequences.
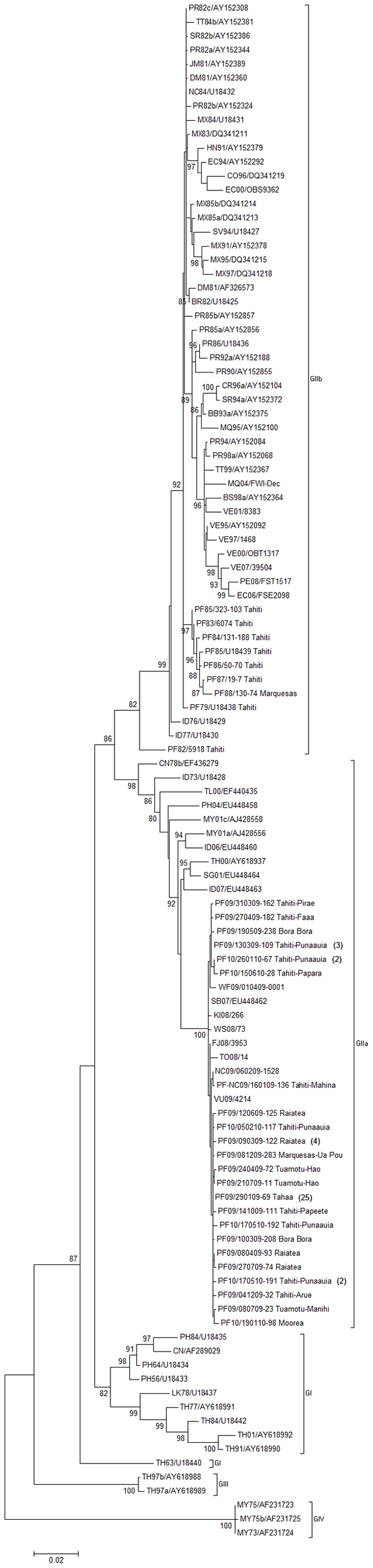
ML original tree derived from 110 DENV-4 E gene sequences. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) is shown for values over 80. The number of strains with identical E gene sequence is indicated in parenthesis (these additional strains could have been collected in a different district or island and at a different date than the strain that appears in the tree).
Figure 3. Evolutionary relationships of DENV-4 Oceania group within clade IIa.

ML original tree derived from 63 DENV-4 E gene sequences. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) is shown next to the branches.
Based on the Solomon Island [SB07/EU448462] E gene sequence, 53 sites exhibited a nucleotide (nt) substitution of which 18 were present on more than one strain but didn't lead to an amino acid (aa) substitution (Table 3). The analysis of the 142 E gene sequences included in this study revealed 79 aa mutated sites, of which one aa substitution (T365I) was specifically encountered within the Oceania group and another (V287I) was only found in the PF80s group (Table 4).
Table 3. Significant nucleotide substitutions on the E gene (1485 bp) within the DENV-4 clade IIa Oceania group.
| nt positions | |||||||||||||||||||
| Cluster | Strains | 15 | 63 | 213 | 273 | 375 | 498 | 549 | 606 | 657 | 738 | 885 | 957 | 978 | 1146 | 1365 | 1386 | 1446 | 1465 |
| none | SB07/EU448462 | G | C | A | T | A | C | C | A | T | G | G | A | T | T | T | A | T | C |
| KI08/266 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | |
| WS08/73 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | |
| PF/WF | WF09/010409-0001 | - | - | - | - | T | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Punaauia 09 Group (3 strains) | - | - | - | - | T | - | - | - | - | - | - | - | - | - | - | - | - | T | |
| PF09/310309-162 Tahiti-Pirae | - | - | - | - | T | - | - | - | - | - | - | - | - | - | - | - | - | T | |
| PF09/270409-182 Tahiti-Faaa | - | - | - | - | T | - | - | - | - | - | - | - | - | - | - | - | - | T | |
| PF09/190509-238 Bora Bora | - | - | - | - | T | - | - | - | - | - | - | - | - | - | - | - | - | T | |
| Punaauia 10a Group (2 strains) | - | - | - | - | T | - | - | - | C | - | - | - | - | - | C | - | - | T | |
| PF10/150610-28 Tahiti-Papara | - | - | - | - | T | - | - | - | C | - | - | - | - | - | C | - | - | T | |
| none | FJ08/3953 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | G | - | - |
| TO08/14 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | G | - | - | |
| PF/NC/VU | VU09/4214 | - | - | - | - | - | - | - | - | - | A | - | - | - | - | - | G | - | - |
| PF-NC09/160109-136 Tahiti-Mahina | - | - | - | - | - | - | T | - | - | A | - | - | - | - | - | G | - | - | |
| NC09/060209-1528 | - | - | - | - | - | - | T | - | - | A | - | - | - | A | - | G | - | - | |
| PF09/10 Group (25 strains) | - | - | - | - | - | - | - | - | - | A | - | - | - | A | - | G | - | - | |
| PF09/100309-208 Bora Bora | - | - | - | - | - | - | - | - | - | A | - | - | - | A | - | G | - | - | |
| PF09/120609-125 Raiatea | - | - | - | - | - | - | - | - | - | A | - | - | - | A | - | G | - | - | |
| PF09/141009-111 Tahiti-Papeete | - | - | - | - | - | - | - | - | - | A | - | - | - | A | - | G | - | - | |
| PF09/041209-32 Tahiti-Arue | - | - | - | - | - | - | - | - | - | A | - | - | - | A | - | G | - | - | |
| PF09/081209-283 Marquesas-Ua Pou | - | - | - | - | - | - | - | - | - | A | - | - | - | A | - | G | - | - | |
| PF10/050210-117 Tahiti-Punaauia | - | - | - | - | - | - | - | - | - | A | - | - | - | A | - | G | - | - | |
| PF10/170510-192 Tahiti-Punaauia | - | - | - | - | - | - | - | - | - | A | - | - | - | A | - | G | - | - | |
| Maupiti 09 Group (4 strains) | A | - | - | C | - | - | - | - | - | A | - | - | - | A | - | G | - | - | |
| PF09/080409-93 Raiatea | - | - | - | - | - | - | - | - | - | A | - | G | - | A | - | G | - | - | |
| PF09/270709-74 Raiatea | - | - | - | - | - | - | - | - | - | A | - | G | - | A | - | G | - | - | |
| PF09/240409-72 Tuamotu-Hao | - | - | - | - | - | T | - | - | - | A | - | - | - | A | - | G | - | - | |
| PF09/210709-11 Tuamotu-Hao | - | - | - | - | - | T | - | - | - | A | - | - | - | A | - | G | - | - | |
| PF09/080709-23 Tuamotu-Manihi | - | T | - | - | - | - | - | - | - | A | - | - | - | A | - | G | - | - | |
| Moorea 10 Group (2 strains) | - | T | G | - | - | - | - | - | - | A | A | - | - | A | - | G | C | - | |
| Punaauia 10b Group (2 strains) | - | - | - | - | - | - | - | G | - | A | - | - | C | A | - | G | - | - | |
- Identical to Solomon.
Table 4. Relevant amino acid substitutions on the E protein based on the consensus sequence of 142 DENV-4 strains.
| Clade | Group | Substitutions | Comments |
| IIa | Oceania | M/I34T | TO08/14 |
| I/A46T* | Clade IIa (except WF09/010409-1); Clade IIb (except PF82/5918) | ||
| I/T46A | WF09/010409-1 | ||
| L82P | TO08/14 | ||
| S120L | Clade IIa (except MY01a) | ||
| S156P | TO08/14 | ||
| V160M | PF10/170510-192 | ||
| V173I | PF09/310309-162 | ||
| N276S | PF09/141009-111 | ||
| K323Q | PF09/190509-238 | ||
| V335I | Oceania group, MY01a (Clade IIa); TH63 (Genotype I); Genotype IV | ||
| T365I* | Oceania group | ||
| I380V | KI08/266 | ||
| D/E384N* | PF10/050210-117, MY01a (Clade IIa); LK78 (Genotype I); Clade IIb | ||
| IIb | PF80s | I/A46T* | Clade IIb (except PF82/5918); Clade IIa (except WF09/010409-1) |
| V/A141I | PF85/U18439 | ||
| D/S154G | PF88/130-74 | ||
| T155I | PF82/5918; ID73, CN78b (Clade IIa); PH56, TH63 (Genotype I) | ||
| K/E202R | PF79; TH63, LK78 (Genotype I) | ||
| T221A | PF82/5918; CN, PH56, PH64, PH84 (Genotype I) | ||
| A/T222V | PF87/19-7 | ||
| S/L/A227T | PF80s group (except PF79/U18438, PF82/5918, PF85/323-103) | ||
| V287I | PF80s group (except PF79/U18438) | ||
| I351V | PF80s group (except PF79/U18438); most strains from Genotype IIb; CN, PH84 (Genotype I) | ||
| P356L | PF88/130-74 | ||
| F357L | Clade IIb; ID73 (Genotype IIa); PH56, PH64, PH84, TH63, TH84 (Genotype I) | ||
| D/E384N* | Clade IIb; PF10/050210-117, MY01(Clade IIa); LK78 (Genotype I) |
*non conservative aa substitution; strains of interest are in bold characters.
Discussion
The recent emergence of DENV-4 in the SPICs occurred after a decade of active DENV-1 circulation and about 20 years after the previous circulation of DENV-4 in the region [23], [24]. This observation is consistent with previous epidemiological data suggesting that, in the region and particularly in FP, sustained transmission of a predominant serotype/genotype occurs following a 20 to 25 years periodic cycle [33], [34]. DENV-1 was still circulating in FP when DENV-4 started to be transmitted locally, but from July 2009, by performing DENV serotype-specific RT-PCR on all sera previously tested positive using NS1 test, we only found DENV-4 (Table 2). Up to November 2011, the only DENV-1 case reported was a traveler just coming back from San-Martin, Caribbean (data not shown). Previous studies already demonstrated that the sensitivity of the Platelia™ Dengue NS1 Ag test depends on the infecting DENV serotype, however the sensitivity of the test was found to be at least identical or higher for DENV-1 compared to DENV-4 [35], [36]. Our results suggest that DENV-4 displaced DENV-1. Consistently, the displacement of the DENV serotype previously circulating by the newly introduced serotype is a characteristic that has been constantly observed in FP over time [33], [34].Since the second semester of 2008, at the time the transmission of DENV-4 in the South Pacific region was dramatically increasing, dengue surveillance was reinforced in FP. However, despite an active tracking, health authorities did not reach to prevent and control the introduction of DENV-4 in FP. Beside the two DENV-4 cases imported from NC to Tahiti in mid-January, additional introduction events that remained undetected might have occurred, particularly in the Leeward islands (Society Islands) where the firsts cases due to local transmission were reported (Table 2). Additional DENV-4 cases were then progressively reported in other FP archipelagos, the spread of the virus being supported by the high frequency of inter-island travel, mostly by plane. Reminding that the DENV-1 epidemic in 2001 also started in the Leeward Islands [23], this place should be considered as a main entry point for new emergent DENV serotypes. Consistently, the Leeward Islands, notably Bora Bora, are major touristic stations in FP.
In order to complement the epidemiological data and get a better understanding of the events that might have led to the introduction and spread of DENV-4 in FP, we conducted a phylogenetic analysis on the complete E gene of DENV-4 strains collected recently and in the 1980s in FP. We also included in the study some strains collected in other SPICs in 2008–2009 and additional sequences available on GenBank (Figure 2). The resulting phylogenetic tree displays the four distinct genotypes already described [4], [29]–[31]. Furthermore, it shows that DENV-4 strains collected in FP, both in the 1980s and in 2009–2010, belong to genotype II and are distributed in clades IIa and IIb [4], [32]. These two clades are distant from more than 6% of genetic divergence and could be therefore reasonably classified in two distinct genotypes [4]. Clearly, these clades evolved separately from a common ancestor originating from South-East Asia. Interestingly, only clade IIa continued to circulate beyond the year 2000 in Southeast Asia and in the Pacific region. In contrast strains belonging to clade IIb were isolated in Central and South America and in the Caribbean at least until 2008 [37], [38]. Indeed, DENV-4 genotype IIb is still predominating in the Americas. The replacement of genotype IIb by the genotypes predominating in Southeast Asia (I, IIa, III, IV) did not occur; although introductions of genotype I were reported in Brazil in 2007 [39]. This contrasts with the situation observed for DENV-2, where the Southeast Asian genotype totally replaced the American genotype previously circulating [40], [41].
Our results corroborate previous studies showing that both in the 1980s and in the early year 2000, DENV-4 had been introduced into the region from Southeast Asia [22], [24], [30]. In addition, we found that all FP strains along with other DENV-4 strains recently collected in the SPICs form a distinct group rooted by the Solomon Island strain [SB07/EU448462], we called Oceania group (Figure 3). Within the Oceania group, the FP strains are distributed into two distinct clusters: PF/NC/VU that also comprises strains collected in NC and Vanuatu, and PF/WF that includes the strain from Wallis & Futuna. This observation suggests that the recent emergence of DENV-4 in FP results from multiple introductions of strains circulating in different SPICs. Both clusters circulated from March 2009 to June 2010. However, PF/NC/VU is widely distributed in the different FP archipelagos whereas PF/WF only comprises strains collected in the Society archipelago. Interestingly, the early dissemination of the virus started in the Leeward Islands within PF/NC/VU and in Tahiti within PF/WF. This observation suggests that in addition to be a main entry point for emergent DENV serotypes, the Leeward Islands should also be considered as an efficient starting point for active dissemination of the virus throughout all FP. The analysis of the nt substitutions within the two clusters compared to the Solomon Island strain [SB07/EU448462] reveals two nt substitutions specific to PF/NC/VU and another substitution specific to PF/WF (Table 3). Moreover, within each cluster, all FP strains share one additional nt substitution. Particularly, all FP strains in cluster PF/WF exhibit a nt substitution (C1465T) in the genomic E/NS1 junction region which is propitious to the study of evolutionary characteristics [4]. All of these nt substitutions are synonymous, showing that no major mutational event occurred on the E gene from the first emergence of DENV-4 genotype IIa in the Solomon Islands in 2007 to the current circulation in FP. Interestingly, some specific missense mutations resulting in aa substitutions occurred on the E gene of the strains collected in Kiribati and Tonga (Table 4).
A more global analysis of the aa changes on the E protein based on the consensus aa sequence of 142 DENV-4 strains reveals mutations that are specific to strains collected in the South Pacific. Within genotype IIb, all the strains composing the PF80s group, except [PF79/U18438], exhibited the same substitution V287I within the domain I of the E gene [42]. Within genotype IIa, all the strains belonging to the Oceania group show two substitutions (V335I and T365I) already described [22], [42]. These mutations occurred within the domain III that mediates DENV binding to cellular receptors and has been proved to be highly immunogenic [42], [43]. Our results show that in the 1980s like in the recent emergence of DENV-4 in the region, specific aa substitutions might have been acquired and subsequently conserved on all strains circulating in the SPICs during periods ranging from 3 (2007–2010) to 5 (1983–1988) years. In a previous study, investigating the microevolution of DENV-1 during 6 years of sustained circulation in FP, two specific aa substitutions on the E protein, that appeared during the endemic transmission period and were rapidly stabilized, were suggestive of an adaptation to the mosquito vector [18]. In the present study, the aa substitutions on the E protein found on all DENV-4 strains collected in the SPICs could reflect a founder effect, to be associated with either one or several factors characterizing the “South Pacific Islands context”: insularity, climate, presence of endemic vector species, small human population sizes and low population flows.
Acknowledgments
We are grateful to Myrielle Dupont-Rouzeyrol, Olivia O'Connor and Suzanne Chanteau from the Institut Pasteur de Nouvelle Calédonie (Nouméa, New Caledonia) and Jean-François Yvon from the Laboratoire d'analyses de l'hôpital de Sia (Wallis, Wallis & Futuna) for providing blood samples blotted on filter paper cards from DENV-4 infected patients.
Footnotes
Competing Interests: The authors have declared that no competing interests exist.
Funding: This work was supported by the “Fonds de Coopération Economique, Sociale et Culturelle pour le Pacifique” -Ministère des Affaires Etrangères et Européennes, France (Conventions N°138/1/2008, 75/1/2009), (URL: http://www.diplomatie.gouv.fr) and by the “Agence nationale pour la Recherche”, France (Décision attributive d'aide N°ANR-09-MIEN-028-01), URL: http://www.agence-nationale-recherche.fr. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.World Health Organization. Dengue and dengue haemorrhagic fever. Factsheet N°117 revised March 2009. Geneva: World Health Organization; 2008. Available: http://www.who.int/mediacentre/factsheets/fs117/en. Accessed 2011 Dec 1. [Google Scholar]
- 2.Rosen L, Rozeboom LE, Sweet BH, Sabin AB. The transmission of Dengue by Aedes polynesiensis Marks. Am J Trop Med Hyg. 1954;3:878–882. doi: 10.4269/ajtmh.1954.3.878. [DOI] [PubMed] [Google Scholar]
- 3.World Health Organization. Dengue: Guidelines for diagnosis, treatment, prevention and control. New edition. Geneva: TDR/WHO; 2009. Available: http://whqlibdoc.who.int/publications/2009/9789241547871_eng.pdf. Accessed 2011 Dec 1. [PubMed] [Google Scholar]
- 4.Rico-Hesse R. Molecular evolution and distribution of dengue viruses type 1 and 2 in nature. Virology. 1990;174:479–493. doi: 10.1016/0042-6822(90)90102-w. [DOI] [PubMed] [Google Scholar]
- 5.Pandey BD, Igarashi A. Severity-related molecular differences among nineteen strains of Dengue type 2 viruses. Microbiol Immunol. 2000;44:179–188. doi: 10.1111/j.1348-0421.2000.tb02480.x. [DOI] [PubMed] [Google Scholar]
- 6.Rico-Hesse R. Microevolution and virulence of Dengue viruses. Adv Virus Res. 2003;59:315–341. doi: 10.1016/s0065-3527(03)59009-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Anderson JR, Rico-Hesse R. Aedes aegypti vectorial capacity is determined by the infecting genotype of dengue virus. Am J Trop Med Hyg. 2006;75:886–882. [PMC free article] [PubMed] [Google Scholar]
- 8.Alliot FA. Etude sur une épidémie de dengue observée à Tahiti. Janvier à Mai 1885. Sa propagation à bord de l'aviso le Volage. Bordeaux Th Med 1886–1887 #78. 1887:35. [Google Scholar]
- 9.Laigret J, Rosen L, Scholammer G. Sur une épidémie de Dengue survenue à Tahiti en 1964. Relations avec les «Fièvres Hémorragiques» du Sud-Est asiatique. Bull Soc Pathol Exot. 1967;60:339–353. [PubMed] [Google Scholar]
- 10.Saugrain J, Rosen L, Outin-Fabre D, Moreau JP. Une récente épidémie d'arbovirose du type Dengue à Tahiti. Comparaison avec l'épidémie de 1964. Bull Soc Pathol Exot. 1970;63:631–641. [PubMed] [Google Scholar]
- 11.Singh N, Kierdzynski T, Lepers C, Benyon EK. Dengue in the Pacific – an update of the current situation. Pac Health Dialog. 2005;12:111–119. [PubMed] [Google Scholar]
- 12.Moreau JP, Rosen L, Saugrain J, Lagraulet J. An epidemic of Dengue on Tahiti associated with hemorrhagic manifestations. Am J Trop Med Hyg. 1973;22:237–241. doi: 10.4269/ajtmh.1973.22.237. [DOI] [PubMed] [Google Scholar]
- 13.Gubler DJ, Reed D, Rosen L, Hitchcock JR., Jr Epidemiologic, clinical, and virologic observations on Dengue in the Kingdom of Tonga. Am J Trop Med Hyg. 1978;27:581–589. doi: 10.4269/ajtmh.1978.27.581. [DOI] [PubMed] [Google Scholar]
- 14.A-Nuegoonpipat A, Berlioz-Arthaud A, Chow V, Endy T, Lowry K, et al. Sustained transmission of dengue virus type 1 in the Pacific due to repeated introductions of different Asian strains. Virology. 2004;329:505–512. doi: 10.1016/j.virol.2004.08.029. [DOI] [PubMed] [Google Scholar]
- 15.Imrie A, Zhao Z, Bennett SN, Kitsutani P, Laille M, et al. Molecular epidemiology of dengue in the Pacific: introduction of two distinct strains of dengue virus type-1 into Hawaii. Ann Trop Med Parasitol. 2006;100:327–336. doi: 10.1179/136485906X105589. [DOI] [PubMed] [Google Scholar]
- 16.Laille M, Roche C. Comparison of Dengue-1 virus envelope glycoprotein gene sequences from French Polynesia. Am J Trop Med Hyg. 2004;71:478–484. [PubMed] [Google Scholar]
- 17.Wiegant A, Renou L, Louette R, Melix G, Hirshauer C, et al. Dengue fever epidemiology and related control strategies in French Polynesia 2006–2007. Inform'ACTION #27. 2007. Available: http://www.spc.int/phs/ENGLISH/Publications/InformACTION/IA27-contents.htm. Accessed 2011 Dec 1.
- 18.Descloux E, Cao-Lormeau VM, Roche C, de Lamballerie X. Dengue 1 diversity and microevolution, French Polynesia 2001–2006: connection with epidemiology and clinics. PLoS Negl Trop Dis. 2009;3:e493. doi: 10.1371/journal.pntd.0000493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Daudens E, Lastère S, Hirschauer C, Cao-Lormeau VM, Louette R, et al. Epidemiology of dengue and control strategies in French Polynesia from 2006 to 2008. BEH thématique. 2009;48-49-50:499–503. [Google Scholar]
- 20.Lepers C. Dengue fever situation in the Pacific Island Countries and Territories, 30 September 2008. Inform'ACTION #29. 2008. Available: http://www.spc.int/phs/ENGLISH/Publications/InformACTION/IA29-contents.htm. Accessed 2011 Dec 1.
- 21.Shu PY, Su CL, Liao TL, Yang CF, Chang SF, et al. Molecular characterization of dengue viruses imported into Taiwan during 2003–2007: geographic distribution and genotype shift. Am J Trop Med Hyg. 2009;80:1039–1046. [PubMed] [Google Scholar]
- 22.Li DS, Liu W, Guigon A, Mostyn C, Grant R, et al. Rapid displacement of dengue virus type 1 by type 4, pacific region, 2007–2009. Emerg Infect Dis. 2010;16:123–125. doi: 10.3201/eid1601.091275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Parc F, Pichon G, Tetaria C, Louis F, Laigret J. La Dengue due au virus de type 4 en Polynésie française. I - Epidémiologie générale, aspects cliniques particuliers. Med Trop. 1981;41:93–96. [PubMed] [Google Scholar]
- 24.Chungue E, Cassar O, Drouet MT, Guzman MG, Laille M, et al. Molecular epidemiology of Dengue-1 and Dengue-4 viruses. J Gen Virol. 1995;76:1877–1884. doi: 10.1099/0022-1317-76-7-1877. [DOI] [PubMed] [Google Scholar]
- 25.Johnson BW, Russell BJ, Lanciotti RS. Serotype-specific detection of dengue viruses in a fourplex real-time reverse transcriptase PCR assay. J Clin Microbiol. 2005;43:4977–4983. doi: 10.1128/JCM.43.10.4977-4983.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Tamura K, Peterson D, Peterson N, Stecheir G, Nei M, et al. MEGA5: Molecular Evolutionary Genetics Analysis using maximum likelihood evolutionary distance, and maximum parsimony methods. Mol Biol Evol. 2011;28:2731–2739. doi: 10.1093/molbev/msr121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kimura M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J Mol Evol. 1980;16:111–120. doi: 10.1007/BF01731581. [DOI] [PubMed] [Google Scholar]
- 28.Felsenstein J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution. 1985;39:783–791. doi: 10.1111/j.1558-5646.1985.tb00420.x. [DOI] [PubMed] [Google Scholar]
- 29.Lanciotti RS, Gubler DJ, Trent DW. Molecular evolution and phylogeny of dengue-4 viruses. J Gen Virol. 1997;78:2279–2284. doi: 10.1099/0022-1317-78-9-2279. [DOI] [PubMed] [Google Scholar]
- 30.Klungthong C, Zhang C, Mammen MP, Jr, Ubol S, Holmes EC. The molecular epidemiology of dengue virus serotype 4 in Bangkok, Thailand. Virology. 2004;329:168–179. doi: 10.1016/j.virol.2004.08.003. [DOI] [PubMed] [Google Scholar]
- 31.Klungthong C, Putnak R, Mammen M P, Li T, Zhang C. Molecular genotyping of dengue viruses by phylogenetic analysis of the sequences of individual genes. J Virol Methods. 2008;154:175–181. doi: 10.1016/j.jviromet.2008.07.021. [DOI] [PubMed] [Google Scholar]
- 32.AbuBakar S, Wong PF, Chan YF. Emergence of dengue virus type 4 genotype IIA in Malaysia. J Gen Virol. 2002;83:2437–2442. doi: 10.1099/0022-1317-83-10-2437. [DOI] [PubMed] [Google Scholar]
- 33.Chungue E, Deparis X, Murgue B. Dengue in French Polynesia: Major features, surveillance, molecular epidemiology and current situation. Pac Health Dialog. 1998;5:154–162. [Google Scholar]
- 34.Cao-Lormeau VM, Roche C, Descloux E, Viallon J, Lastère S, et al. Lost in French Polynesia: which strategies for a dengue virus to spread? Am J Trop Med Hyg. 2007;77(5 suppl):129. [Google Scholar]
- 35.Bessoff K, Delorey M, Sun W, Hunsperger E. Comparison of two commercially available dengue virus (DENV) NS1 capture enzyme-linked immunosorbent assays using a single clinical sample for diagnosis of acute DENV infection. Clin Vaccine Immunol. 2008;15(10):1513–1518. doi: 10.1128/CVI.00140-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Guzman MG, Jaenisch T, Gaczkowski R, Ty Hang VT, Sekaran SD, et al. Multi-country evaluation of the sensitivity and specificity of two commercially-available NS1 ELISA assays for dengue diagnosis. PLoS Negl Trop Dis. 2010;4(8):e811. doi: 10.1371/journal.pntd.0000811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Forshey BM, Morrison AC, Cruz C, Rocha C, Vilcarromero S, et al. Dengue virus serotype 4, Northeastern Peru, 2008. Emerg Infect Dis. 2009;15:1815–1818. doi: 10.3201/eid1511.090663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Tang Y, Quintana M, Zhang C, Li T, Sauls D, et al. Letter to the editor: molecular genotyping of Dengue Virus Types 2 and 4 from the Guatemalan and Honduran Epidemics of 2007 using the envelope glycoprotein gene. Virus Genes. 2011;42:200–203. doi: 10.1007/s11262-010-0567-3. [DOI] [PubMed] [Google Scholar]
- 39.De Melo FL, Romano CM, de Andrade Zanotto PM. Introduction of dengue virus 4 (DENV-4) genotype I into Brazil from Asia? PLoS Negl Trop Dis. 2009;3:e390. doi: 10.1371/journal.pntd.0000390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Rico-Hesse R, Harrison LM, Salas RA, Tovar D, Nisalak A, et al. Origins of dengue type 2 viruses associated with increased pathogenicity in the Americas. Virology. 1997;230:244–251. doi: 10.1006/viro.1997.8504. [DOI] [PubMed] [Google Scholar]
- 41.Bennett SN, Holmes EC, Chirivella M, Rodriguez DM, Beltran M, et al. Molecular evolution of dengue 2 virus in Puerto Rico: positive selection in the viral envelope accompanies clade reintroduction. J Gen Virol. 2006;87:885–893. doi: 10.1099/vir.0.81309-0. [DOI] [PubMed] [Google Scholar]
- 42.Modis Y, Ogata S, Clements D, Harrison SC. A ligand-binding pocket in the dengue virus envelope glycoprotein. Proc Natl Acad Sci USA. 2003;100:6986–6991. doi: 10.1073/pnas.0832193100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Guzman MG, Hermida L, Bernardo L, Ramirez R, Guillén G. Domain III of the envelope protein as a dengue vaccine target. Expert Rev Vaccines. 2010;9:137–147. doi: 10.1586/erv.09.139. [DOI] [PubMed] [Google Scholar]