

Abstract
Cryptosporidiosis is a leading cause of persistent diarrhea in children in impoverished and developing countries and has both a short- and long-term impact on the growth and development of affected children. An animal model of cryptosporidial infection that mirrors closely the complex interaction between nutritional status and infection in children, particularly in vulnerable settings such as post-weaning and malnourishment, is needed to permit exploration of the pathogenic mechanisms involved. Weaned C57BL/6 mice received a protein-deficient (2%) diet for 3–12 days, then were infected with 5 × 107 excysted C. parvum oocyts, and followed for rate of growth, parasite stool shedding, and intestinal invasion/morphometry. Mice had about 20% reduction in weight gain over 12 days of malnutrition and an additional 20% weight loss after C. parvum challenge. Further, a significantly higher fecal C. parvum shedding was detected in malnourished infected mice compared to the nourished infected mice. Also, higher oocyst counts were found in ileum and colon tissue samples from malnourished infected mice, as well as a significant reduction in the villous height–crypt depth ratio in the ileum. Tissue Th1 cytokine concentrations in the ileum were significantly diminished by malnutrition and infection. mRNA for toll-like receptors 2 and 4 were diminished in malnourished infected mice. Treatment with nitazoxanide did not prevent weight loss or parasite stool shedding. These findings indicate that, in the weaned animal, malnutrition intensifies cryptosporidial infection, while cryptosporidial infection further impairs normal growth. Depressed TLR2 and 4 signaling and Th1 cytokine response may be important in the mechanisms underlying the vicious cycle of malnutrition and enteric infection.
Cryptosporidium parvum is an obligate intracellular protozoan parasite that invades and resides in the epithelial cells of the small intestine (Tzipori and Ward, 2002; Sasahara et al., 2003). Discovered by Tyzzer in 1907, it is now well recognized as a major cause of diarrheal illness and gastroenteritis in children and adults worldwide (Dillingham et al., 2002; Huang et al., 2004; Tzipori and Widmer, 2008). Cryptosporidiosis is associated with severe life-threatening illness among immunocompromised individuals, most notably those with AIDS (Pozio et al., 1997). Different from immunocompetent adults, in whom the disease is usually a self-limited diarrhea, people with AIDS are susceptible to a devastating form manifested by chronic, voluminous diarrhea and increased mortality (Pantenburg et al., 2008; Borad and Ward, 2010). Furthermore, in patients with AIDS, infections may result in malabsorption of antiretroviral drugs and, possibly, in increased drug resistance (Brantley et al., 2003). In resource-limited countries, the disease probably exerts most of its impact on pediatric health, particularly as an important cause of morbidity and mortality in young children (Agnew et al., 1998; Newman et al., 1999). In these areas, successive or persistent infections of Cryptosporidium spp. intensified by malnutrition have been considered responsible for substantial long-term consequences for the later physical and cognitive development of these children and even leading to death (Guerrant et al., 1999; Gendrel et al., 2003). Malnutrition itself can increase susceptibility to infection, which, in turn, contributes to further malnutrition, resulting in a vicious cycle, with potentially serious health consequences for the host (Guerrant et al., 2000; Katona and Katona-Apte, 2008).
Attempts to treat cryptosporidial infections in humans have included antimicrobial drugs, as well as other pharmacologic approaches, but have yielded limited or only partial success (Armson et al., 2003; Gargala, 2008; Rossignol, 2010). Nitazoxanide (NTZ) was effective in treating diarrhea in immunocompetent children and adults but is not effective in the absence of an appropriate immune response; in AIDS patients, restoration of the immune system, along with antimicrobial treatment, is necessary (Maggi et al., 2000). The development of effective intervention is hampered by the still incomplete knowledge about the human immune response toward C. parvum, how the parasite causes disease, and the contribution of immunocompromising conditions, e.g., AIDS or malnutrition.
Our goal is to study the interaction of malnutrition and C. parvum infection in weaned mice, a model that is more easily executed than previous neonatal models and correlates more closely to the complex interaction between the immune response and infectious agents in children transitioning from breast feeding to solid foods. We observed the weight loss caused by malnutrition, as well as infection, quantified fecal parasite shedding, and intestinal tissue parasite burden, and analyzed morphometric and histopathologic characteristics of crypts and villi from the ileal mucosa. In addition, we examined the effectiveness of the treatment with NTZ in this model and assessed the innate and adaptive immune responses by measuring toll-like receptors (TLRs) and Th1/Th2 cytokines production in the intestinal tissue.
MATERIALS AND METHODS
Animal studies
Animals and malnutrition protocol
Twenty-one-day-old female C57BL/6 mice were purchased from Charles River Laboratories, Inc. (Wilmington, Massachusetts). Mice were acclimated, fed a regular diet for 3 days, and then assigned to experimental groups matched for body weight. On day 24 of life, mice assigned to the nourished groups received chow containing 20% protein (Harland Labs, Madison, Wyoming), and mice assigned to the malnourished groups received isocaloric chow with 2% protein (Harland Labs). The animals remained on their diets for 3–12 days before oocyst challenge and continued their diet until the end of the experiment. In some experiments, mice were killed by cervical dislocation on the peak of infection (day 3 or 4 post-infection [PI]), at which time ileal and colonic specimens were collected and processed. The protocol described here was approved and is in accordance with the Institutional Animal Care and Use Committee policies of the University of Virginia.
Preparation and administration of C. parvum
The C. parvum oocysts were obtained from experimentally infected calves (Iowa isolate; Waterborne, Inc., New Orleans, Louisiana). Oocysts of C. parvum were stored in phosphate-buffered saline (PBS) at 4 C and used within 8 wk of their receipt from Waterborne. The concentration of oocysts in the stock solution was measured using a hemocytometer. Oocysts were excysted by adding 1 part sodium hypochlorite to 4 parts of the oocysts solution to make a final concentration of 20% sodium hypochlorite solution. The tube was briefly vortexed and incubated in room temperature for 10 min. The number of oocysts was then counted before and after hypochlorite to compute for the excystation rate (30–35% in these experiments). Each infected mouse received 75 μl of PBS containing freshly prepared excysted oocysts in a recently vortexed solution (5 × 107 oocysts per mouse) by oral gavage directly into the stomach.
Administration of NTZ
Nitazoxanide was purchased as a powder, formulated for oral administration to children (Alinia™; Romark Pharmaceuticals, Tampa, Florida). It was prepared as a suspension, containing 2 mg per 50 μl of water, a concentration allowing a 50-μl dose to be equivalent to 100–150 mg/kg depending on the weight of each animal. For mice treated with NTZ, each received 50 μl daily for 3 days via gavage, beginning 24 hr after challenge with excysted oocysts.
Stool collection and weight measurements
Mice were weighed daily, and stools were collected daily from the infected and non-infected groups in pre-weighed tubes and stored at −20 C until DNA or protein was extracted.
Analysis of samples
DNA extraction
The DNA was extracted from the frozen stool samples using Qiagen QIAamp DNA Stool Kit (Qiagen, Inc., Germantown, Maryland) with minor modifications as follows. First 400 μl Buffer ASL (warmed to 60 C) was added to each sample, and each sample was vortexed at 1,500 rpm overnight until homogenized. The samples were incubated at 82.5 C for 5 min and then vortexed for 1 min at full speed. The supernatant was pipetted into a new tube with 30 μl of proteinase K to which was added 400 μl of Buffer AL and incubated at 70 C for 10 min; 400 μl of ethanol (100%) was added and mixed by vortexing. The “Protocol for Isolation of DNA from Stool for Pathogen Detection” was then followed to completion, and the DNA was eluted in 200 μl Elution Buffer and stored at −20 C. DNA from tissue samples was extracted from frozen tissue samples using the QIAamp DNA Tissue Kit (Qiagen), following the protocol exactly.
Real-time PCR for C. parvum quantification
Extracted DNA (5 μl) was added to the master mix (20 μl) to give a total reaction volume of 25 μl per sample. Master mix was prepared by mixing 12.5 μl of Bio-Rad iQ SYBR Green Supermix (Bio-Rad Laboratories, Hercules, California), 5.5 μl of DEPC-treated nuclease free sterile water (Fisher Scientific, Pittsburgh, Pennsylvania), and 1.0 μl (6.2 μM) each of both forward and reverse primers (Invitrogen, Carlsbad, California). The primers target the 18s rRNA gene of the parasite (forward: 5′-CTGCGAATGGCTCATTATAACA-3′; reverse: 5′-AGGCCAATACCCTACCGTCT-3′; GenBank no. AF164102). The reaction was performed in a Bio-Rad iCycler iQ multicolor PCR Detection System using iCycler software (version 3.0). Amplification consisted of 15 min at 95 C followed by 40 cycles of 15 sec at 95 C, 15 sec at 52 C, and 20 sec at 72 C, followed by 0.5-degree increments for 10 sec starting at 75 C and ending with 95 C for the Melt Curve. Fluorescence was measured during the annealing step of each cycle. Ct values of each run were compared to standards with known amounts of C. parvum DNA and transformed into number of organisms per mg of stool sample.
RNA extraction and reverse transcriptase PCR for toll-like receptors expression
Ileal and colon samples were taken immediately from necropsied animals and frozen in liquid nitrogen. After freezing, the samples were stored at −80 C until analysis. Samples were thawed and total RNA was extracted using Qiagen RNeasy mini Kit according to the manufacturer’s instructions. RNA concentration was quantified and checked for purity (A260:280 ratio) by standard spectrophotometry (Biophotometer; Eppendorf, Hamburg, Germany). Synthesis of cDNA by Reverse Transcriptase PCR was performed using SuperScript III First-Strand Synthesis System SuperMix (Invitrogen) with the use of oligo (dT) < as primers. cDNA was used in quantitative PCR for measuring TLR2, 4, and 9 expression compared to GAPDH expression (used as housekeeping gene) and using the nourished uninfected group as control. The primers for murine TLR2 (forward: 5′-TCT GCT GTG CCC TTC TCC TGT TGA-3; reverse: 5′-GGC CGC GTC GTT GTT CTC GT-3′) (GenBank no. NM_011905), TLR4 (forward: 5′-AGC CGG AAG GTT ATT GTG GTA GT-3′; reverse: 5′-TGC CGT TTC TTG TTC TTC CTC T-3′; GenBank no. NM_021297.2) and TLR9 (forward: 5′-GCA ATG GAA AGG ACT GTC CAC TTT GTG-3′; reverse: 5′-ATC GCC TTC TAT CGC CTT CTT GAC GAG-3′; GenBank no. AY649791) were purchased from Invitrogen. Amplification consisted of 10 min at 95 C, followed by 40 cycles of 25 sec at 95 C, 25 sec at the respective annealing temperature to each pair of primers (60 C for TLR2 and 4, 45 C for TLR9, and 54 C for GAPDH), and 20 sec at 72 C, followed by 40 cycles of 10 sec starting at 75 C with 0.5 C increment for the Melt Curve. Fluorescence was measured during the annealing step of each cycle. Reagents and equipments were the same as what were used in C. parvum quantification above. The relative gene expression was determined using the 2−ΔΔCt method (Pfaffl, 2001).
Cytokine concentrations in ileal homogenates
Ileal tissue was homogenized using cold lysis buffer (50 mM HEPES, 1% Triton ×100 and 1:100 protease inhibitor), centrifuged at 4 C at full speed for 10 min, then supernatant was collected and stored at −80 C. Levels of IL-2, IL-4. IL-5, IL-6, IL-10, IL-12, GM-CSF, IFN-γ, and TNF-α were measured using a Bio-Plex Pro Mouse Cytokine Assay (Pro Mo Cyto TH1/TH2 1 × 96, M6000003J7; Bio-Rad) according to the manufacturer’s instructions. A dual-laser, flow-based microplate reader system was used to report the level of target protein in each sample, employing a Bio-Plex 200 reader System (Bio-Rad).
Histopathology analyses
Digital micrographs of H&E sections were taken using a high-resolution microscope Aperio ScanScope Slide Scanner and analyzed with ImageScope software (Aperio Technologies, Vista, California). For villus height and crypt depth, well-oriented sections of ileum for 4 mice at day 3 or 4 post-challenge were analyzed.
Statistical analyses
Analyses of weight loss/increase were expressed as a percent change in baseline body weight. The analyses of stool shedding (number of parasites per mg of stool) and tissue burden (number of parasites per mg of tissue) were performed using SPSS 17.0 software (Chicago, Illinois). Statistical analyses were performed using ANOVA with Bonferroni post-hoc correction. A P value less than 0.05 was considered significant. Data were presented as mean ± SEM.
RESULTS
Malnutrition is critical for generating a murine model of excysted C. parvum infection
Protein deprivation and C. parvum challenge influence growth
The inability to attain an expected rate of growth has been proposed as a biological marker of malnutrition, or enteric infection, or both, in humans and in small animal models (Coutinho et al., 2008; Mondal et al., 2009). Following up to 12 days of a special diet containing 2% protein, 2 of 4 groups of 36-day-old C57BL/6 mice had a profound failure to gain weight (day 26–36; P < 0.01) (Fig. 1). Challenge with excysted C. parvum oocysts (5 × 107/mouse) was associated with a rapid further impairment in the rate of weight gain of malnourished mice, compared to iso-calorically nourished mice, which steadily gained weight despite infection (P < 0.001). Malnourished infected mice had about 20% reduction in weight gain over 14 days caused by protein deprivation, and an additional 20% weight loss caused by C. parvum challenge. Similar results were observed in a second independent study. These findings suggest that malnutrition in association with C. parvum infection in weaned C57BL/6 mouse results in disease of substantial severity in which there is profound weight loss.
Figure 1.
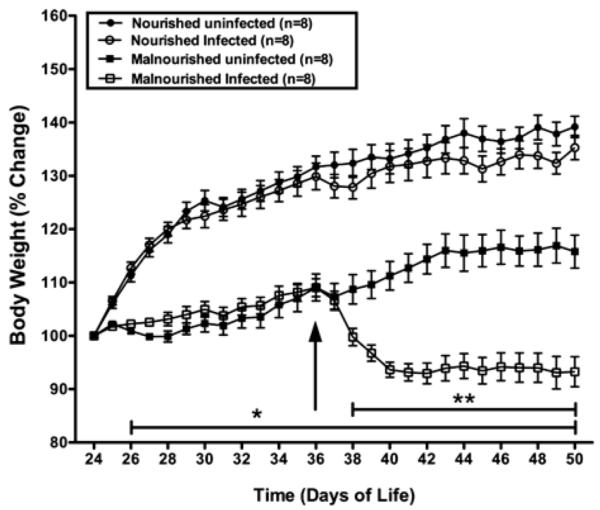
Growth of mice as altered by nutritional status and by challenge with C. parvum oocysts. Female C57BL/6 mice at 24 days of life were assigned to either a 2% (malnourished) or 20% (nourished) protein diet. Groups to be infected were then challenged 12 days later (day 36 of life) with 5 × 107 excysted oocysts/mouse. Shown is the body weight change (percentage of initial body weight) from each group starting at day 24 until day 50 of life; *P < 0.01 for both malnourished groups compared with both nourished groups; **P < 0.01 for malnourished infected mice compared with malnourished uninfected mice. Results are shown as mean ± SEM.
Stool shedding of parasites
We reasoned that the intensity of enteric infection in mice might be best reflected in the number of and duration over which challenge organisms are shed into the stool. To test this hypothesis, we collected stool from each oocyst-challenged mouse for 14 days and quantified the number of organisms per mg of stool by quantitative PCR (Fig. 2). Malnourished mice shed more organisms than nourished mice (by 1–3 logs on days 1–7 PI), with shedding gradually abating over time. This result suggested that malnourished and nourished mice can both be infected orally, but that the intensity of infection, as mirrored by shedding of organisms into the stool, is markedly increased in those that are malnourished. Findings were similar in a second independent experiment with a shorter duration of observation.
Figure 2.
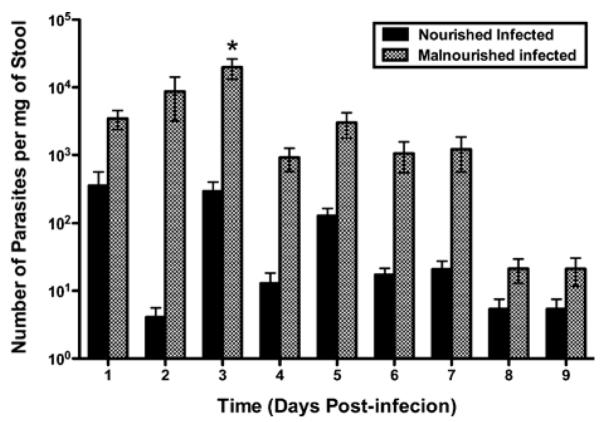
Malnutrition increases stool shedding of oocyst-challenged mice. Parasite shedding was increased in malnourished infected relative to the nourished infected mice over the first 9 days PI. Shown is the number of parasites per mg of stool determined by quantitative real-time PCR. *P < 0.05 at day 3 PI (n = 11). Results are shown in log scale as mean ± SEM.
Tissue burden of organisms
While stool shedding may reflect the rate of generation of new intra-luminal parasites, the ability of the organism to infect the epithelium may be more indicative of disease activity. Therefore, stool-free colon and distal ileum were taken from C57BL/6 mice 3 days after challenge with 5 × 107 excysted oocysts/mouse and analyzed for C. parvum, by quantifying the amount of C. parvum-specific DNA. Normalized by wet weight of tissue, malnourished mice were observed to have 3.5-fold more C. parvum organisms per mg of tissue, in both the ileum and the colon, than nourished mice (P < 0.05) (Fig. 3).
Figure 3.
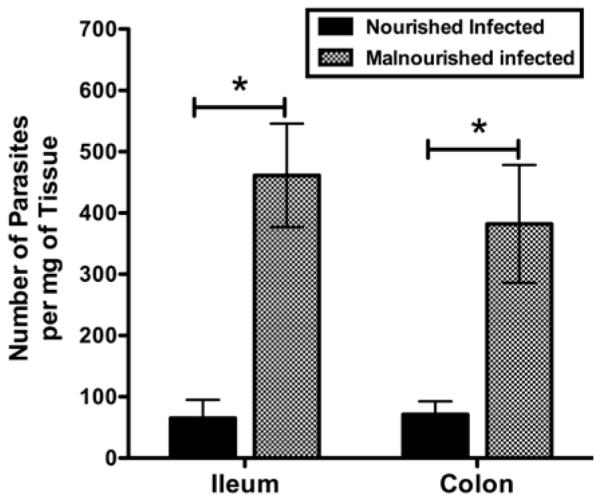
Tissue burden of C. parvum organisms in the ileum and colon of challenged mice, as altered by nutritional status. Female C57BL/6 mice, nourished or malnourished (12 days), were challenged with 5 × 107 excysted oocysts/mouse and then killed 3 days later in order to determine the level of ileal and colonic tissue burden of organisms. The number of parasites per mg of tissue was determined by quantitative PCR. Data are shown as mean ± SEM. n = 5 mice per group. For ileum and colon, *P < 0.05 for malnourished infected vs. nourished infected mice.
Duration of protein deprivation
While the malnourished C. parvum–challenged weaned mouse does manifest enhanced infection as assessed by prolonged stool shedding and a substantial tissue burden (Figs. 2, 3), the duration and extent of protein deprivation needed to enable parasite colonization and stool shedding in these mice is not clear. Therefore, we subjected to the 2% protein diet identical groups of mice for 3, 7, or 10 days, then challenged them with 5 × 107 excysted oocysts per mouse (Fig. 4). Stool shedding through days 7–11 was observed, with a tendency to prolonged shedding with longer periods of protein deprivation. Similarly, challenge of mice with half as many excysted oocysts (2.5 × 107) was associated with modestly (but not significantly) fewer shed oocysts than in mice given the higher inoculum challenge (data not shown). Thus, while short periods of protein deprivation and challenge with as few as 2.5 × 107 excysted oocysts/mouse appear sufficient for enabling infection in a weaned C57BL/6 mice model, longer duration (10 days) was associated with more prolonged stool shedding through day 11.
Figure 4.
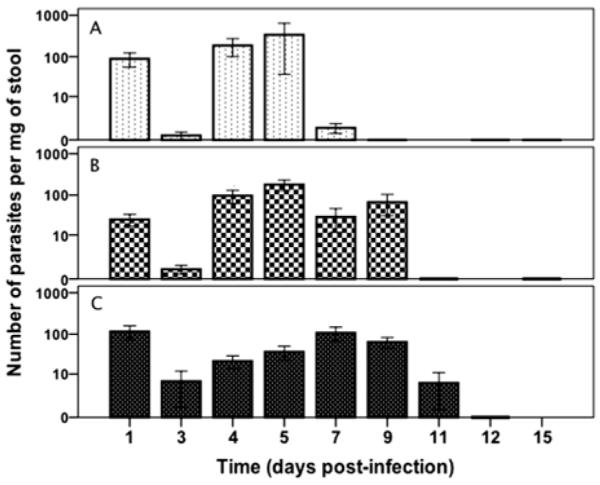
Stool shedding as affected by the duration of protein deprivation. Mice were malnourished for 3, 7, or 10 days (A, B, C, respectively), then challenged with 5 × 107 excysted oocysts per mouse. Shown is the number of parasites per mg of stool, determined by qPCR over time after C. parvum challenge. Results are shown in log scale as mean ± SEM. n = 4 per group.
Taken together, these data (Figs. 1–4) suggest that C. parvum challenge of C57BL/6 mice that are made protein deficient results in a loss of growth velocity, more intense infection as reflected in stool shedding of organisms, and heavier epithelial infection compared to nourished mice.
Cryptosporidial infection causes intestinal crypt changes in malnourished mice
Histopathological changes
Cryptosporidium parvum infection in humans is associated with villus blunting and crypt hyperplasia (Orenstein, 1997; Alcantara et al., 2008); hence, we examined whether there would be similar changes in the excysted oocyst–challenged mouse. On day 3 following oocyst challenge, ileal segments were examined for villous height and crypt depth. A reduction in villous height was noted in the malnourished mice (Fig. 5A), and crypt depth was observed to be significantly greater in the ileum of malnourished infected animals compared with any other group (P < 0.05) (Fig. 5B). Nourished infected mice did not show significant differences in villous height or crypt depth compared with nourished and malnourished uninfected animals. The ratio of villous height to crypt depth was significantly lower in malnourished infected animals compared with any other group (P < 0.01) (Fig. 5C). Representative ileal histopathology from infected and uninfected nourished and malnourished mice is shown in Figure 6.
Figure 5.

Impact of malnutrition and infection on ileal morphology. Ileal sections were harvested from C57BL/6 mice 3 days after C. parvum challenge with 5 × 107 oocysts/mouse. (A) Villus height was diminished in malnourished mice but was not statistically different from other groups. (B) Crypt depth was increased in malnourished infected mice compared with any other group (*P < 0.05). (C) Villus height:crypt depth was lower in malnourished infected vs. any other group (*P < 0.01). n = 3–5 per group. Bars show mean ± SEM.
Figure 6.

Ileal morphology in malnourished mice. Tissue from uninfected control (A) and infected (B) mice were examined at 4 days PI, demonstrating diminished villous height (*) and increased crypt depth (**) in malnourished, infected mice. Brackets mark the length of villi and crypts in representative specimens.
TLR mRNA expression
Previous studies have shown that TLR2 and TLR4, through activation of NFκB, mediate defense against cryptosporidiosis in human cholangiocytes (Chen et al., 2005, 2007) while in C. parvum–infected neonatal mice, activation of TLR9 by CpG-ODN decreased intestinal parasite load by 80–95% (Barrier et al., 2006). To begin to explore innate immune events underlying the effects of infection and malnutrition, ileal tissues were harvested from C57BL/6 mice 3 days after challenge with 5 × 107 excysted oocysts/mouse and analyzed for mRNA expression for TLR2, 4, and 9. Malnourished uninfected mice showed higher TLR2 and TLR4 expression compared with nourished controls (Fig. 7; P < 0.05 for TLR2). Malnourished infected mice had lower TLR2 and TLR4 expression compared with malnourished uninfected controls (P < 0.05 for TLR2). No difference was observed on TLR9 expression. These data suggest that infection with C. parvum is associated a marked reduction in expression of TLR2 mRNA from that induced by malnutrition alone. Similar, but not statistically significant, effects were seen with TLR4.
Figure 7.
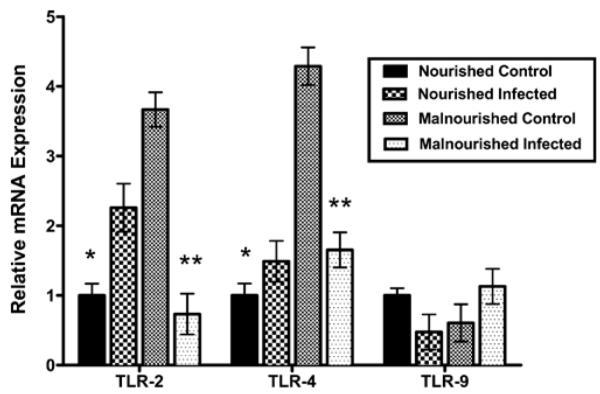
Relative mRNA expression for TLR2, TLR4, and TLR9 measured with RT-qPCR. C57BL/6 mice were malnourished for 10 days, challenged with 5 × 107 excysted C. parvum oocysts, and killed 3 days later for ileal tissue. Age-matched nourished untreated mice were studied as controls. Malnourished uninfected mice showed significantly higher TLR2 and TLR4 expressions compared with nourished controls (*). Malnourished infected mice have significantly lower TLR2 and TLR4 expressions compared to malnourished control (**). Ct values were normalized for GAPDH (used as reference gene) in the same sample. Expression ratio calculated using the 2−ΔΔCt method. n = 4 per group. Bars show mean ± SEM.
Cryptosporidium parvum generation of cytokines
Given studies above showing that C. parvum elicits morphological changes and an increased tissue burden of organisms in the ileum of malnourished mice, we examined whether local (mucosal) generation of pro-inflammatory cytokines might play a role. Therefore, ileal tissues from nourished (n = 4) and malnourished (n = 4) weaned mice challenged with 5 × 107 excysted oocysts/mouse, were examined for cytokine concentrations 4 days post-challenge. IFN-γ, shown previously to be important for resistance to C. parvum infection (Coutinho et al., 2008), was 292 ± 72 pg/ml in normal uninfected ileum, but was reduced with malnutrition (37 ± 32 pg/ml), with infection (103 ± 13 pg/ml), as well as in the setting of malnutrition and infection (107 ± 14 pg/ml) (P < 0.02, compared to normal uninfected tissue). The TNF-α concentrations determined in ileum followed a similar pattern (929 ± 413 pg/ml dropping to 693, 236, and 584 pg/ml, respectively) (P < 0.03, comparing nourished control and nourished infected tissue). IL-2 was also highest in concentration in nourished, uninfected control tissue (5.1 ± 3.9 pg/ml), while IL-4 and IL-5 were essentially unchanged. These findings suggest that malnutrition, as well as infection with C. parvum, is associated with marked reductions in at least 2 important Th1 cytokines in affected tissue, compared to control, and may contribute to inability of the host to combat the parasite after challenge.
Nitazoxanide is not effective against cryptosporidiosis in malnourished mice
Prior reports suggested that nitazoxanide may be therapeutic for C. parvum infection in pediatric populations (Diaz et al., 2003). Therefore, nitazoxanide was given by gavage (2 mg/mouse/day) on days 1, 2, and 3 after challenge with 5 × 107 excysted C. parvum oocysts, to C57BL/6 mice malnourished for 10–12 days with a 2% protein diet. Nitazoxanide did not ameliorate weight loss in malnourished infected mice, relative to malnourished non-infected mice, studied over 14 days post-challenge (Fig. 8). Furthermore, nitazoxanide did not diminish the intensity of stool shedding of parasites relative to controls (malnourished infected but non-treated mice; Fig. 9). Nourished challenged mice continued to shed the fewest oocysts. These findings suggest that, at oral doses similar on a weight basis to those used in human pediatric populations, nitazoxanide had no effect on preserving weight gain or on reducing the intensity of infection measured by stool shedding in C. parvum–challenged malnourished mice.
Figure 8.
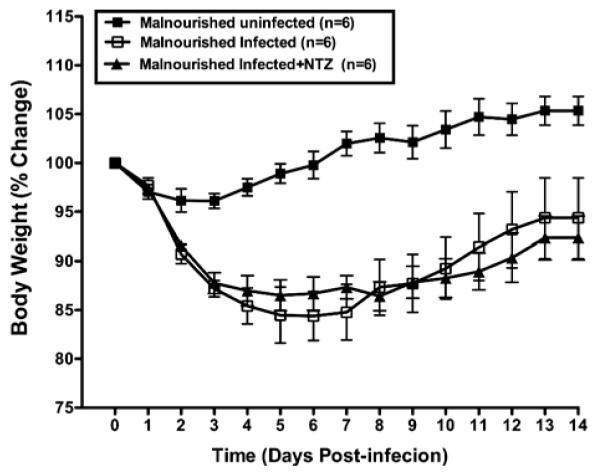
Effect of three consecutive days of treatment with NTZ (days 1 to 3 PI) on body weight change in malnourished female C57BL/6 mice challenged with 5 × 107 excysted oocysts/mouse at day 36 of life. Results are shown as mean ± SEM. P < 0.05 for malnourished uninfected compared to any infected group. NTZ had no significant effects on weight change.
Figure 9.
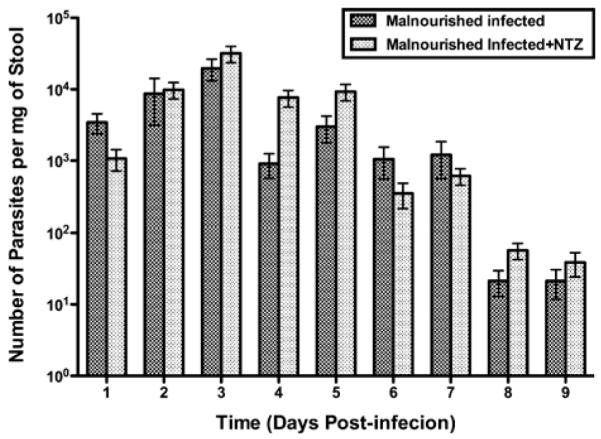
Lack of effect of NTZ on fecal shedding of parasites in malnourished mice described in Figure 8 (procedures for malnutrition and administration of NTZ were the same). Shown is the number of parasites per mg of stool determined by quantitative PCR. Results are shown in log scale as mean ± SEM.
DISCUSSION
Cryptosporidiosis is a diarrheal illness that is self-limited for many healthy travelers but is frequently devastating and persistent for young malnourished children living in many areas, as well as immune-compromised patients such as those with AIDS (Dillingham et al., 2002; Huang et al., 2004; Tzipori and Widmer, 2008). Several animal models of C. parvum infection have been published, primarily in the neonatal host or in the immune-suppressed adult mouse (Ungar et al., 1990). Here we report a new model in the weaned malnourished mouse, unique in its ability to be easily generated (change in diet), require minimal manipulation, be free of the need for inborn genetic defects in protective cytokines, and have robust quantifiable end points (growth rate, stool shedding) that can be followed longitudinally, associated with tissue invasion and morphometric changes, and whose size allows a wealth of tissue to be available for study in vitro. This new malnourished weaned mouse model should provide an excellent tool to elucidate disease mechanisms contributing to the infection-malnutrition cycle, as well as putative therapies, with several distinct advantages compared to other models. The time interval before disease onset is one such area, particularly in comparison with genetically manipulated models. SCID mice require a minimum of 2–3 wk before shedding is detected (Mead et al., 1991; Tzipori et al., 1995). In the interferon gamma knockout adult mouse infected with small doses of oocysts, body weight change does not occur for 7 days, while high doses of oocysts (~107/mouse) are needed to observe profound weight loss (Griffiths et al., 1998). Models dependent on steroid administration require continuous immune suppression, as well as a high challenge dose of oocysts in the adult mouse (Rasmussen and Healey, 1992; Cheng et al., 1996; Surl and Kim, 2006). In contrast, the current model we report demonstrated weight loss and robust shedding of oocysts on the first day post-challenge, with a time course that extended for at least 11 days. Our finding of prolonged stool shedding, detected by PCR, is consistent with that clinically observed in humans and strongly supports the notion that nutritional status affects host susceptibility to cryptosporidiosis (Agnew et al., 1998; Bushen et al., 2007). The absence of genetic manipulation of the immune system/response in the current model enables it to be used to explore the naturally occurring innate and acquired immune response to cryptosporidiosis. While parasite attachment/invasion of the ileal and colonic mucosa was marked in the malnourished challenged animal, atrophy of villi and elongation of crypts were seen in the ileum when the host was both malnourished and infected with C. parvum.
The new malnourished weaned mouse model can be useful to study already established chemotherapies and new potential therapeutics against C. parvum. NTZ is the drug of choice to treat cryptosporidiosis (Rossignol, 2010). The effect of NTZ has been studied in immunocompetent and immunodeficient people, including young children and patients with AIDS and highly active antiretroviral therapy (HAART) (Rossignol et al., 2001; Smith and Corcoran, 2004; Anderson and Curran, 2007), in many animal models as well as in vitro (Gargala et al., 2000). In the present study, we analyzed for the first time the effect of the treatment with NTZ on malnutrition and C. parvum infection using a weaned mouse model. We observed that malnourished infected mice treated with NTZ did not show significant differences in stool shedding of parasites or body weight loss compared to malnourished infected mice that did not receive the treatment. These findings contrast with previous studies showing that NTZ can reduce C. parvum infection in neonatal mice (Blagburn et al., 1998) and SCID adult mice (Theodos et al., 1998). This discrepancy with the current study likely reflects the effects of malnutrition that intensifies cryptosporidiosis as was previously observed in neonatal mice (Coutinho et al., 2008) and, potentially, of wasting among HIV-infected pediatric populations in developing countries, contributing to a lack of efficacy of NTZ (Zulu et al., 2005; Abubakar et al., 2007; Amadi et al., 2009), indicating the need for improved chemotherapy and alternative approaches to treatment.
Intestinal epithelial cells play an important role in the activation of immune responses to C. parvum infection. Epithelial cells can trigger innate and adaptive immune responses via production of cytokines and activation of TLRs, for example, TLR2, 4, and 9 (Abreu et al., 2005). We analyzed the mRNA expressions of these TLRs from ileal tissue of nourished and malnourished mice that were infected and uninfected. Compared with control (nourished, uninfected), malnourished uninfected mice exhibited up-regulation of TLR2 and TLR4 mRNA expression. However, in the presence of infection at day 4 post-challenge, both were diminished to near control levels. TLR9 message expression did not change with malnutrition or with infection (Fig. 7). Prior in vitro studies using epithelial cells showed that TLR2 and TLR4 mediate defense to C. parvum infection (Chen et al., 2005, 2007; O’Hara et al., 2009) and that MyD88 (signaling adaptor of most TLRs) mediates protective responses against cryptosporidiosis in adult mice (Rogers et al., 2006). Moreover, malnutrition and infection in weaned mice resulted to inadequate local humoral response as evident in the reduced ileal TNFα and IFNγ levels. Our findings suggest that the intensified and prolonged shedding in the current model may be due to the inability to sustain protective innate immune responses when the malnourished host is challenged with C. parvum in vitro.
Our results have several limitations. Inherent to the nature of animal studies, it is uncertain whether our findings would hold true in other mouse strains or in humans. Although weight loss and intestinal histopathology were noted, infected mice, unlike in human disease, were not observed to develop diarrhea. Only selected TLRs and cytokines were examined, and no time course experiments were performed, possibly missing changes that occur at different time points after infection and during recovery of the epithelium. Besides malnutrition, there may be other host, parasite, or environmental factors, not examined by us, that could have resulted in poor response to nitazoxanide.
In conclusion, we described cryptosporidiosis in a novel weaned mouse model with biological end points (weight loss, stool shedding, tissue burden of organisms, and morphometry) that are worsened by malnutrition. In this model, NTZ did not change the natural history of the disease. TLR2 and TLR4 were down-regulated, and fewer Th1 cytokines were seen with malnutrition, potentially helping to explain the increased infection and histopathology in the intestinal epithelium. The model may be useful in elucidating mechanisms that intersect malnutrition and C. parvum infection, as well as developing new therapies to better control the disease in vulnerable human populations.
ACKNOWLEDGMENTS
This study was partially funded by the National Institutes of Health/National Institute of Allergy and Infectious Diseases grants U54 AI057168 (Middle Atlantic Research Center of Excellence for Biodefense and Emerging Infectious Diseases Research) and U01 AI75520. LBC, JES, and RF were supported by the NIH/Fogarty International Center Global Infectious Disease Research Training grant D43 TW0006578. We acknowledge the valuable assistance of Edgar Martin for cytokine assay, Dr. Relana Pinkerton for statistical analyses, the University of Virginia Research Histology Core, and Yamen N. Hama for preparation of figures.
LITERATURE CITED
- Abreu MT, Fukata M, Arditi M. TLR signaling in the gut in health and disease. Journal of Immunology. 2005;174:4453–4460. doi: 10.4049/jimmunol.174.8.4453. [DOI] [PubMed] [Google Scholar]
- Abubakar I, Aliyu SH, Arumugam C, Hunter PR, Usman NK. Prevention and treatment of cryptosporidiosis in immunocompromised patients. Cochrane Database Systems Review. 2007 doi: 10.1002/14651858.CD004932.pub2. CD004932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Agnew DG, Lima AA, Newman RD, Wuhib T, Moore RD, Guerrant RL, Sears CL. Cryptosporidiosis in northeastern Brazilian children: Association with increased diarrhea morbidity. Journal of Infectious Disease. 1998;177:754–760. doi: 10.1086/514247. [DOI] [PubMed] [Google Scholar]
- Alcantara WC, Destura RV, Sevilleja JE, Barroso LF, Carvalho H, Barrett LJ, O’Brien AD, Guerrant RL. Detection of epithelial-cell injury, and quantification of infection, in the HCT-8 organoid model of cryptosporidiosis. Journal of Infectious Diseases. 2008;198:143–149. doi: 10.1086/588819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amadi B, Mwiya M, Sianongo S, Payne L, Watuka A, Katubulushi M, Kelly P. High dose prolonged treatment with nitazoxanide is not effective for cryptosporidiosis in HIV positive Zambian children: A randomised controlled trial. BMC Infectious Diseases. 2009;9:195. doi: 10.1186/1471-2334-9-195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson VR, Curran MP. Nitazoxanide: A review of its use in the treatment of gastrointestinal infections. Drugs. 2007;67:1947–1967. doi: 10.2165/00003495-200767130-00015. [DOI] [PubMed] [Google Scholar]
- Armson A, Thompson RC, Reynoldson JA. A review of chemotherapeutic approaches to the treatment of cryptosporidiosis. Expert Review of Anti-Infective Therapy. 2003;1:297–305. doi: 10.1586/14787210.1.2.297. [DOI] [PubMed] [Google Scholar]
- Barrier M, Lacroix-Lamande S, Mancassola R, Auray G, Bernardet N, Chausse AM, Uematsu S, Akira S, Laurent F. Oral and intraperitoneal administration of phosphorothioate oligodeoxynucleotides leads to control of Cryptosporidium parvum infection in neonatal mice. Journal of Infectious Diseases. 2006;193:1400–1407. doi: 10.1086/503748. [DOI] [PubMed] [Google Scholar]
- Blagburn BL, Drain KL, Land TM, Kinard RG, Moore PH, Lindsay DS, Patrick DA, Boykin DW, Tidwell RR. Comparative efficacy evaluation of dicationic carbazole compounds, nitazoxanide, and paromomycin against Cryptosporidium parvum infections in a neonatal mouse model. Antimicrobial Agents and Chemothery. 1998;42:2877–2882. doi: 10.1128/aac.42.11.2877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borad A, Ward H. Human immune responses in cryptosporidiosis. Future Microbiology. 2010;5:507–519. doi: 10.2217/fmb.09.128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brantley RK, Williams KR, Silva TM, Sistrom M, Thielman NM, Ward H, Lima AA, Guerrant RL. AIDS-associated diarrhea and wasting in northeast Brazil is associated with subtherapeutic plasma levels of antiretroviral medications and with both bovine and human subtypes of Cryptosporidium parvum. Brazilian Journal of Infectious Diseases. 2003;7:16–22. doi: 10.1590/s1413-86702003000100003. [DOI] [PubMed] [Google Scholar]
- Bushen OY, Kohli A, Pinkerton RC, Dupnik K, Newman RD, Sears CL, Fayer R, Lima AA, Guerrant RL. Heavy cryptosporidial infections in children in northeast Brazil: Comparison of Cryptosporidium hominis and Cryptosporidium parvum. Transactions of the Royal Society of Tropical Medicine and Hygiene. 2007;101:378–384. doi: 10.1016/j.trstmh.2006.06.005. [DOI] [PubMed] [Google Scholar]
- Chen XM, O’Hara SP, Nelson JB, Splinter PL, Small AJ, Tietz PS, Limper AH, Larusso NF. Multiple TLRs are expressed in human cholangiocytes and mediate host epithelial defense responses to Cryptosporidium parvum via activation of NF-kappaB. Journal of Immunology. 2005;175:7447–7456. doi: 10.4049/jimmunol.175.11.7447. [DOI] [PubMed] [Google Scholar]
- – – – , Splinter PL, O’Hara SP, Larusso NF. A cellular micro-RNA, let-7i, regulates Toll-like receptor 4 expression and contributes to cholangiocyte immune responses against Cryptosporidium parvum infection. Journal of Biological Chemistry. 2007;282:28929–28938. doi: 10.1074/jbc.M702633200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng L, Rasmussen KR, Healey MC, Yang S. Primary and secondary infections with Cryptosporidium parvum in immuno-suppressed adult mice. American Journal of Tropical Medicine and Hygiene. 1996;55:324–329. doi: 10.4269/ajtmh.1996.55.324. [DOI] [PubMed] [Google Scholar]
- Coutinho BP, Oria RB, Vieira CM, Sevilleja JE, Warren CA, Maciel JG, Thompson MR, Pinkerton RC, Lima AA, Guerrant RL. Cryptosporidium infection causes undernutrition and, conversely, weanling undernutrition intensifies infection. Journal of Parasitology. 2008;94:1225–1232. doi: 10.1645/GE-1411.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diaz E, Mondragon J, Ramirez E, Bernal R. Epidemiology and control of intestinal parasites with nitazoxanide in children in Mexico. American Journal of Tropical Medicine and Hygiene. 2003;68:384–385. [PubMed] [Google Scholar]
- Dillingham RA, Lima AA, Guerrant RL. Cryptosporidiosis: Epidemiology and impact. Microbes and Infection. 2002;4:1059–1066. doi: 10.1016/s1286-4579(02)01630-1. [DOI] [PubMed] [Google Scholar]
- Gargala G. Drug treatment and novel drug target against Cryptosporidium. Parasite. 2008;15:275–281. doi: 10.1051/parasite/2008153275. [DOI] [PubMed] [Google Scholar]
- – – – , Delaunay A, Li X, Brasseur P, Favennec L, Ballet JJ. Efficacy of nitazoxanide, tizoxanide and tizoxanide glucuronide against Cryptosporidium parvum development in sporozoite-infected HCT-8 enterocytic cells. Journal of Antimicrobial Chemotherapy. 2000;46:57–60. doi: 10.1093/jac/46.1.57. [DOI] [PubMed] [Google Scholar]
- Gendrel D, Treluyer JM, Richard-Lenoble D. Parasitic diarrhea in normal and malnourished children. Fundamental and Clinical Pharmacology. 2003;17:189–197. doi: 10.1046/j.1472-8206.2003.00169.x. [DOI] [PubMed] [Google Scholar]
- Griffiths JK, Theodos C, Paris M, Tzipori S. The gamma interferon gene knockout mouse: A highly sensitive model for evaluation of therapeutic agents against Cryptosporidium parvum. Journal of Clinical Microbiology. 1998;36:2503–2508. doi: 10.1128/jcm.36.9.2503-2508.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guerrant DI, Lima AA, Davidson F. Micronutrients and infection: Interactions and implications with enteric and other infections and future priorities. Journal of Infecious Diseases. 2000;182(Suppl. 1):S134–S138. doi: 10.1086/315924. [DOI] [PubMed] [Google Scholar]
- – – – , Moore SR, Lima AA, Patrick PD, Schorling JB, Guerrant RL. Association of early childhood diarrhea and cryptosporidiosis with impaired physical fitness and cognitive function four-seven years later in a poor urban community in northeast Brazil. American Journal of Tropical Medicine and Hygiene. 1999;61:707–713. doi: 10.4269/ajtmh.1999.61.707. [DOI] [PubMed] [Google Scholar]
- Huang DB, Chappell C, Okhuysen PC. Cryptosporidiosis in children. Seminars in Pediatric Infectious Disease. 2004;15:253–259. doi: 10.1053/j.spid.2004.07.006. [DOI] [PubMed] [Google Scholar]
- Katona P, Katona-Apte J. The interaction between nutrition and infection. Clinical Infectious Diseases. 2008;46:1582–1588. doi: 10.1086/587658. [DOI] [PubMed] [Google Scholar]
- Maggi P, Larocca AM, Quarto M, Serio G, Brandonisio O, Angarano G, Pastorel G. Effect of antiretroviral therapy on cryptosporidiosis and microsporidiosis in patients infected with human immunodeficiency virus type 1. European Journal of Clinical Microbiology and Infectious Diseases. 2000;19:213–217. doi: 10.1007/s100960050461. [DOI] [PubMed] [Google Scholar]
- Mead JR, Arrowood MJ, Sidwell RW, Healey MC. Chronic Cryptosporidium parvum infections in congenitally immuno-deficient SCID and nude mice. Journal of Infectious Diseases. 1991;163:1297–1304. doi: 10.1093/infdis/163.6.1297. [DOI] [PubMed] [Google Scholar]
- Mondal D, Haque R, Sack RB, Kirkpatrick BD, Petri WA., JR. Attribution of malnutrition to cause-specific diarrheal illness: Evidence from a prospective study of preschool children in Mirpur, Dhaka, Bangladesh. American Journal of Tropical Medicine and Hygiene. 2009;80:824–836. [PMC free article] [PubMed] [Google Scholar]
- Newman RD, Sears CL, Moore SR, Nataro JP, Wuhib T, Agnew DA, Guerrant RL, Lima AA. Longitudinal study of Cryptosporidium infection in children in northeastern Brazil. Journal of Infectious Diseases. 1999;180:167–175. doi: 10.1086/314820. [DOI] [PubMed] [Google Scholar]
- O’Hara SP, Small AJ, Gajdos GB, Badley AD, Chen XM, Larusso NF. HIV-1 Tat protein suppresses cholangiocyte toll-like receptor 4 expression and defense against Cryptosporidium parvum. Journal of Infectious Diseases. 2009;199:1195–1204. doi: 10.1086/597387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orenstein JM. Cryptosporidiosis. In: Connor DH, Chandler FW, Manz HJ, Schwartz DA, Lack EE, editors. Pathology of infectious diseases. Appleton and Lange; Stamford, Connecticut: 1997. pp. 1147–1168. [Google Scholar]
- Pantenburg B, Dann SM, Wang HC, Robinson P, Castellanos-Gonzalez A, Lewis DE, White AC., Jr. Intestinal immune response to human Cryptosporidium sp. infection. Infection and Immunity. 2008;76:23–29. doi: 10.1128/IAI.00960-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfaffl MW. A new mathematical model for relative quantifica-tion in real-time RT-PCR. Nucleic Acids Research. 2001;29:e45. doi: 10.1093/nar/29.9.e45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pozio E, Rezza G, Boschini A, Pezzotti P, Tamburrini A, Rossi P, Di Fine Smacchia C, Schiesari A, Gattei E, Zucconi R, Ballarini P. Clinical cryptosporidiosis and human immunodeficiency virus (HIV)-induced immunosuppression: Findings from a longitudinal study of HIV-positive and HIV-negative former injection drug users. Journal of Infectious Diseases. 1997;176:969–975. doi: 10.1086/516498. [DOI] [PubMed] [Google Scholar]
- Rasmussen KR, Healey MC. Experimental Cryptosporidium parvum infections in immunosuppressed adult mice. Infection and Immunity. 1992;60:1648–1652. doi: 10.1128/iai.60.4.1648-1652.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogers KA, Rogers AB, Leav BA, Sanchez A, Vannier E, Uematsu S, Akira S, Golenbock D, Ward HD. MyD88-dependent pathways mediate resistance to Cryptosporidium parvum infection in mice. Infection and Immunity. 2006;74:549–556. doi: 10.1128/IAI.74.1.549-556.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rossignol JF. Cryptosporidium and Giardia: Treatment options and prospects for new drugs. Experimental Parasitology. 2010;124:45–53. doi: 10.1016/j.exppara.2009.07.005. [DOI] [PubMed] [Google Scholar]
- – – – , Ayoub A, Ayers MS. Treatment of diarrhea caused by Cryptosporidium parvum: A prospective randomized, double-blind, placebo-controlled study of nitazoxanide. Journal of Infectious Diseases. 2001;184:103–106. doi: 10.1086/321008. [DOI] [PubMed] [Google Scholar]
- Sasahara T, Maruyama H, Aoki M, Kikuno R, Sekiguchi T, Takahashi A, Satoh Y, Kitasato H, Takayama Y, Inoue M. Apoptosis of intestinal crypt epithelium after Cryptosporidium parvum infection. Journal of Infection and Chemotherapy. 2003;9:278–281. doi: 10.1007/s10156-003-0259-1. [DOI] [PubMed] [Google Scholar]
- Smith HV, Corcoran GD. New drugs and treatment for cryptosporidiosis. Current Opinion in Infectious Diseases. 2004;17:557–564. doi: 10.1097/00001432-200412000-00008. [DOI] [PubMed] [Google Scholar]
- Surl CG, Kim HC. Concurrent response to challenge infection with Cryptosporidium parvum in immunosuppressed C57BL/6N mice. Journal of Veterinary Science. 2006;7:47–51. doi: 10.4142/jvs.2006.7.1.47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Theodos CM, Griffiths JK, D’Onfro J, Fairfield A, Tzipori S. Efficacy of nitazoxanide against Cryptosporidium parvum in cell culture and in animal models. Antimicrobial Agents and Chemotherapy. 1998;42:1959–1965. doi: 10.1128/aac.42.8.1959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tzipori S, Rand W, Theodos C. Evaluation of a two-phase scid mouse model preconditioned with anti-interferon-gamma monoclonal antibody for drug testing against Cryptosporidium parvum. Journal of Infectious Diseases. 1995;172:1160–1164. doi: 10.1093/infdis/172.4.1160. [DOI] [PubMed] [Google Scholar]
- – – – , Ward H. Cryptosporidiosis: Biology, pathogenesis, and disease. Microbes and Infection. 2002;4:1047–1058. doi: 10.1016/s1286-4579(02)01629-5. [DOI] [PubMed] [Google Scholar]
- – – – , Widmer G. A hundred-year retrospective on cryptosporidiosis. Trends in Parasitology. 2008;24:184–189. doi: 10.1016/j.pt.2008.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ungar BL, Burris JA, Quinn CA, Finkelman FD. New mouse models for chronic Cryptosporidium infection in immunodeficient hosts. Infection and Immunity. 1990;58:961–969. doi: 10.1128/iai.58.4.961-969.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zulu I, Kelly P, Njobvu L, Sianongo S, Kaonga K, Mcdonald V, Farthing M, Pollok R. Nitazoxanide for persistent diarrhoea in Zambian acquired immune deficiency syndrome patients: A randomized-controlled trial. Alimentary Pharmacology and Therapeutics. 2005;21:757–563. doi: 10.1111/j.1365-2036.2005.02394.x. [DOI] [PubMed] [Google Scholar]