

Abstract
As pulmonary emphysema and diabetes mellitus are common diseases, concomitance of both is correspondingly expected to occur frequently. To examine whether insulin influences the development of inflammation in the alveolar septa, diabetic male Wistar rats (alloxan, 42 mg/kg, i.v., n = 37) and matching controls (n = 31) were used. Ten days after alloxan injection, diabetic and control rats were instilled with physiologic saline solution containing porcine pancreatic elastase (PPE, 0.25 IU/0.2 ml, right lung) or saline only (left lung). The following analyses were performed: (i) number of leucocytes in the bronchoalveolar lavage (BAL) fluid of the animals, 6 h after PPE/saline instillation (early time point); and (ii) mean alveolar diameter (μm) and quantification of elastic and collagen fibres (%) 50 days after PPE/saline instillation (late time point). Relative to controls, alloxan-induced diabetic rats showed a 42% reduction in the number of neutrophils in BAL fluid, a 20% increase in the mean alveolar diameter and a 33% decrease in elastic fibre density in the alveolar septa. Treatment of diabetic rats with 4 IU neutral protamine Hagedorn (NPH) insulin, 2 h before elastase instillation, restored the number of neutrophils in the BAL fluid. The mean alveolar diameter and elastic fibre content in alveolar septa matched the values observed in control rats if diabetic rats were treated with 4 IU NPH insulin 2 h before instillation followed by 2 IU/day for the next 50 days. Density of collagen fibres did not differ between the various groups. Thus, the data presented suggest that insulin modulates the inflammatory and repair responses in elastase-induced emphysema, and assures normal repair and tissue remodelling.
Keywords: diabetes mellitus, insulin, porcine pancreatic elastase, pulmonary emphysema
Pulmonary emphysema, a distinctive type of chronic obstructive pulmonary disease often associated with smoking, is characterized by abnormal and permanent destruction of the alveolar septa by inflammation (Barnes 2008) and degeneration of the elastic component of the extracellular matrix (Black et al. 2008). A diagnosis of emphysema has important prognostic implications. Unlike other lung diseases, emphysema does not respond to anti-inflammatory therapies, and its clinical course is marked by inexorable deterioration. Currently, no medical therapies have been proven to prolong life expectancy. Thus, there is great interest in understanding the mechanisms of injury from normal and remodelling areas, and for the treatment to be effective, we must identify these pathogenic mechanisms to avoid remodelling and destruction of the lungs.
Many studies have debated the importance of inflammation in the pathogenesis of emphysema (reviewed by Taraseviciene-Stewart & Voelkel 2008) and have examined other mechanisms to unravel what might relate to lung repair and remodelling (Chung & Adcock 2008; Taraseviciene-Stewart & Voelkel 2008). A prevailing hypothesis in the generation of emphysema is that there is a protease–antiprotease imbalance. Derangements of this balance may result in acute and chronic inflammation leading to extracellular matrix degradation and remodelling over time. Major experimental approaches to mimic chronic obstructive pulmonary disease (COPD) include inhalation of tobacco smoke and tracheal instillation of tissue-degrading enzymes to induce emphysema-like lesions and gene-modifying techniques leading to COPD-like murine phenotypes (Groneberg & Chung 2004). Instillation of elastases remains a useful tool to study the effects of proteolytic injury and inflammation. Therefore, the effectiveness of a single challenge with porcine pancreatic elastase (PPE) in the development of emphysema has been directly related to the elastolytic activity of the enzyme and severity of the disease (Wright et al. 2008).
Clinical and experimental evidence suggests the occurrence of abnormalities in the course of the inflammatory response in diabetes mellitus (Garcia-Leme & Farsky 1993; Alba-Loureiro et al. 2007). These include reduced number of leucocytes in inflammatory lesions (Pereira et al. 1987; Vianna & Garcia-Leme 1995; Anjos-Valotta et al. 2006), a reduction in mast cell degranulation (Cavalher-Machado et al. 2004), cytokine release (Martins et al. 2006, 2009) and lymph node retention capacity (Moriguchi et al. 2005), which might contribute to increased susceptibility and severity of infections in the diabetic host. Most prior reports regarding the impact of smoking on health did not discuss separately results on subsets of individuals with diabetes, and several of the studies do not distinguish between type 1 and type 2 diabetes. On the other hand, evidence reviewed by Haire-Joshu et al. (1999) indicated that there are consistent results from both cross-sectional and prospective studies showing enhanced risk for micro and macrovascular disease and premature mortality from the combination of smoking and diabetes. In addition, insulin receptors can be identified on granulocytes and endothelial cells (Fussganger et al. 1976; Bar et al. 1978). Therefore, it is reasonable to presume that the activity of the reacting structures in inflammation is under the control of the ambient concentration of insulin. In this context, we hypothesized that the continuing deficiency of insulin associated with diabetes mellitus influences the development of the inflammatory and reparative responses to PPE instillation, thus impairing remodelling of the alveolar walls following emphysema. This study was undertaken to investigate the role of insulin on the course of the inflammatory and reparative responses to PPE instillation into the lungs of alloxan-induced diabetic rats.
Material and methods
Animals
Sixty-eight male Wistar rats weighing 200 ± 20 g (about 2 months of age) at the beginning of the experiments were used. The animals were maintained at 23 ± 2 °C under a cycle of 12 h light/darkness (light on at 06:00 a.m.) and were allowed access to food and water ad libitum. All experiments were in adherence to ethical principles in animal research adopted by the Brazilian College of Animal Experimentation.
Induction of diabetes mellitus and insulin treatment
Diabetes mellitus was induced by an intravenous injection of 42 mg/kg of alloxan monohydrate (Sigma Chemical Co., St. Louis, MO, USA) dissolved in physiologic saline after an overnight fasting period. Control rats were injected with physiologic saline only. Ten days thereafter, the presence of diabetes was verified by blood glucose concentrations (>11.2 mmol/l) determined by a blood glucose monitor (Eli Lilly, São Paulo, SP, Brazil) in samples obtained from the cut tip of the rat tail. Body weight was concomitantly recorded. A group of diabetic rats received 4 IU of neutral protamine Hagedorn (NPH) insulin (Eli Lilly) subcutaneously, 2 h before PPE instillation, and responses were evaluated 6 h thereafter (early time point). A second group of diabetic rats received 4 IU of NPH insulin subcutaneously, 2 h before PPE instillation, followed by 2 IU/day at 05:00 p.m. for the next 50 days. Analyses were evaluated 16 h after the last dose of insulin (late time point).
Porcine Pancreatic Elastase (PPE) instillation
The animals were anaesthetized with an intraperitoneal injection of 300 mg/kg chloral hydrate, and a polyethylene cannula was inserted into the right main bronchus. Physiologic saline solution (0.9% NaCl; 0.2 ml) containing 0.25 IU of PPE (Sigma Chemical Co.) was instilled into the right lung. Control left lung was instilled with physiologic saline through a polyethylene cannula inserted into the left main bronchus.
Bronchoalveolar lavage
Bronchoalveolar lavage (BAL) was performed 6 h after intrabronchial administration of PPE or saline. The animals were anaesthetized as described earlier, and the abdominal cavity was opened for exsanguination from the abdominal aorta. Each lung was lavaged by instillation of 15 ml (three times 5 ml) of phosphate-buffered saline, at room temperature, through a polyethylene tube (1 mm in diameter) inserted into the right and left main bronchi to harvest leucocytes. The lavage fluid was not used if the retrieved volume was <85% of the 15 ml instilled. Total cell counts were determined by using an automatic haemocytometer (CC510; CELM, São Paulo, SP, Brazil). Differential cell counts were carried out on stained slides under oil immersion microscopy. A total of 200 cells were counted and classified as neutrophils, eosinophils or mononuclear cells based on morphological criteria.
Morphometric analysis
The animals were sacrificed 50 days after intrabronchial administration of PPE or saline. The animals were anaesthetized with chloral hydrate (300 mg/kg, i.p.), and lungs were removed by median sternotomy. The lungs were prepared for morphometric measurements by fixation with 10% neutral buffered formaldehyde solution at an inflation pressure of 20 cm H2O for 12 h. After fixation, midcoronal blocks of the right (PPE) and left (saline) lungs were cut and embedded in paraffin, and 5-μm sections were then prepared and stained with haematoxylin and eosin. To evaluate late reparative response, morphometric analysis was carried out with an integrated eyepiece with a coherent system made of a 100-point and 50-line grid of known length coupled to a light microscope (Axioplan; Zeiss, Oberkochen, Germany). Sections were examined at 400× magnification, and the mean alveolar diameter of lung parenchyma (μm) was determined by the point-counting technique (Weibel 1990). The number of alveolar intercepts was counted and divided by the total length of straight lines. This analysis was performed in 20 random, non-overlapping fields in each lung. Points falling on alveolar air spaces, blood vessel lumen and bronchial lumen were excluded. Aiming to investigate the number of macrophages in alveolar space, 20 non-successive microscopic fields of each slide (randomly chosen and coded) were examined with the help of a reticulated eyepiece coupled to an optical microscope at 400× magnification. Results are presented as number of macrophages/mm2. Tissue slices also underwent specific staining methods to compute the amount of collagen and elastic fibres in the alveolar septa. For collagen, the tissue was stained in a solution of Sirius red dissolved in aqueous saturated picric acid and observed under polarized light microscopy (Montes 1996). For elastic fibres, the Weigert's resorcin fuchsin method modified with oxidation was used (Fullmer et al. 1974). In each lung, 10 different microscopic fields were randomly selected to quantify collagen and elastic fibres. Quantification (400× magnification) was carried out with the aid of a digital analysis system consisting of a digital camera (Coolsnap Pro Color; Nikon, Tokyo, Japan) attached to a triocular microscope (Nikon) and a computer. The images were processed using the software Pro-Plus version 4.1 (Media Cybernetics, Silver Spring, MD, USA). The thresholds for collagen and elastic fibres were established after enhancement of the contrast up to the point where the fibres were easily identified as either black (elastic) or birefringent (collagen) bands. Bronchi and blood vessels were carefully avoided during the measurements. Results are presented as the fraction of area (%) occupied by elastic and collagen fibres.
Statistical analysis
All results are expressed as means ± standard error of the mean. The significance of differences between treatment groups was assessed by Student's t-test or analysis of variance followed by Tukey–Kramer multiple comparisons test when appropriate. Wilcoxon test or Kruskal–Wallis test followed by the Dunn's multiple comparison test was used when data were not normally distributed. P<0.05 was considered to be significant.
Results
Body weight gain and blood glucose levels
Relative to controls, animals rendered diabetic by the injection of alloxan 10 days before exhibited a significant reduction in body weight during the experimental period and sharply elevated blood glucose levels. Results are summarized in Table 1. Treatment of diabetic rats with a single dose of NPH insulin, 2 h before PPE/saline instillation, significantly reduced glycaemia from 30.87 ± 1.45 to 18.94 ± 2.12 mmol/l (n = 9, P < 0.001). Table 2 summarizes data from diabetic rats and matching controls that were submitted to instillation 10 days after alloxan injection and evaluated 50 days thereafter. Relative to controls, diabetic rats exhibited hyperglycaemia and a decrease in body weight. Treatment of diabetic rats with 4 IU before instillation followed by 2 IU/day NPH insulin, for the next 50 days, significantly reduced blood glucose levels compared to non-treated diabetic rats. Body weight, however, did not improve.
Table 1.
Characteristics of the animals in the inflammatory phase of pulmonary emphysema
| Body weight (g) at 10-day interval | ||||
|---|---|---|---|---|
| Groups | n | Blood glucose (mmol/l) | Initial | Final |
| Matching control | 20 | 5.07 ± 0.08 | 222 ± 5 | 282 ± 5 |
| Diabetic | 23 | 26.49 ± 1.45* | 234 ± 4 | 212 ± 8* |
Rats were rendered diabetic by the injection of alloxan (42 mg/kg, i.v.) 10 days before porcine pancreatic elastase instillation to evaluate bronchoalveolar response 6 h thereafter. Data are presented as means ± SEM and analysed by Student's t-test as follows
P < 0.001 compared with corresponding values in control group.
Table 2.
Characteristics of the animals in the reparative phase and mean alveolar diameter of lung parenchyma
| Body weight (g) at 60-day interval | |||||
|---|---|---|---|---|---|
| Groups | Blood glucose (mmol/l) | Initial | Final | Instillation | Mean alveolar diameter (μm) |
| Matching control (11) | 4.70 ± 0.17 | 221 ± 6 | 413 ± 15 | PPE | 59.65 ± 1.44‡ |
| Saline | 51.50 ± 1.22 | ||||
| Diabetic (7) | 28.73 ± 1.62* | 234 ± 5 | 230 ± 19* | PPE | 70.99 ± 2.34‡θδ |
| Saline | 54.24 ± 1.43 | ||||
| Diabetic + Insulin (7) | 19.50 ± 2.27*† | 229 ± 6 | 257 ± 4* | PPE | 61.41 ± 1.05γ |
| Saline | 55.48 ± 1.59 | ||||
Rats were rendered diabetic by the injection of alloxan (42 mg/kg, i.v.) 10 days before porcine pancreatic elastase (PPE, right lung) and saline (left lung) instillation. Diabetic rats were treated with 4 IU neutral protamine Hagedorn (NPH) insulin 2 h before instillation, followed by 2 IU/day NPH insulin for the next 50 days. Number of animals is indicated in parenthesis. Data are presented as means ± SEM and analysed by one-way anova followed by the Tukey–Kramer multiple comparisons test as follows
P < 0.001 compared with corresponding values in control group
P < 0.01 compared with values in diabetic group
P < 0.001
P < 0.01 compared with values in saline-treated lungs
P < 0.001 compared with PPE-treated lungs in control group
P < 0.01 compared with PPE-treated lungs in diabetic + insulin group.
PPE-induced leucocyte migration: role of insulin
Compared with saline-instilled lungs, instillation of PPE into the lungs significantly increased the total cell number in the BAL fluid of control rats. Neutrophil counts were elevated accounting for 79% of total cells. There were no differences in the number of mononuclear leucocytes, and eosinophils were absent. Results are illustrated in Figure 1. In contrast, a 42% reduction in the number of neutrophils was observed in the BAL fluid of diabetic rats after instillation of PPE compared with PPE-instilled lungs of control animals. Values obtained for mononuclear leucocytes in PPE-instilled lungs of diabetic rats matched those observed in PPE-instilled lungs of controls. Treatment of diabetic rats with a single dose of NPH insulin, 2 h before PPE instillation, completely restored the capacity of the animals to respond to the proteolytic enzyme. Number of neutrophils harvested from the BAL fluid of these animals matched those observed in control rats. There were no differences in the number of mononuclear cells between groups.
Figure 1.
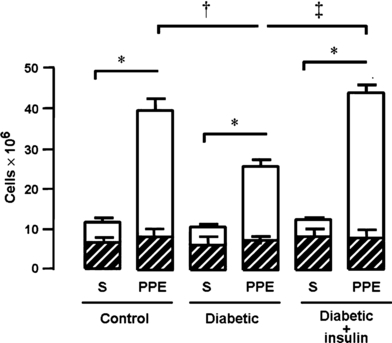
Number of neutrophils (open area) and mononuclear cells (hatched area) in bronchoalveolar lavage fluid of control (n = 20), diabetic (n = 14) and insulin-treated diabetic (n = 9) rats 6 h after porcine pancreatic elastase (PPE, right lung) or saline (S, left lung) instillation. Neutral protamine Hagedorn insulin (4 IU/rat) was given 2 h before instillation. Data are presented as means ± SEM and analysed by one-way anova followed by the Tukey–Kramer multiple comparisons test as follows: *P < 0.001 compared with corresponding values in saline-treated lungs; ‡P < 0.001 and †P < 0.01 compared with PPE-treated lung from diabetic rats.
Morphometric analysis of lung parenchyma: role of insulin
In PPE-instilled lungs, there were characteristic morphologic alterations of emphysematous lesions with a pattern of panacinar emphysema. There was destruction of septa and increase in the alveolar spaces, with formation of structures at the ends of these ruptures known as ‘drum sticks’. Representative photomicrographs are illustrated in Figure 2(a–d). Results, summarized in Table 2, show that in control rats, instillation of PPE into the lungs increased the mean alveolar diameter with consequent reduction in the alveolar surface, compared with saline-instilled lungs. In contrast, in PPE-instilled lungs of diabetic rats, there was a distortion of the acinar architecture because of hyperdistension of the alveolar ducts associated with the rupture of alveolar septa, resulting in 20% increase in alveolar diameter values compared with PPE-instilled lungs of control rats. When diabetic rats were treated with 4 IU NPH insulin 2 h before instillation, followed by 2 IU/day for the next 50 days, the observed histological pattern and values of alveolar diameter in PPE-instilled lungs matched the values attained in PPE-instilled lungs of control rats. No differences were observed in saline-instilled lungs among groups (Table 2), showing the maintenance of lobular and acinar architecture (Figure 2a). An additional analysis was performed in haematoxylin–eosin-stained lung samples to evaluate the number of infiltrating cells in the emphysema-like lesions, 50 days after PPE instillation. Number of macrophages in either PPE-instilled lungs or saline-instilled lungs did not differ among the groups (P=0.0870, Kruskal–Wallis test). Values (mean ± SEM) in PPE-instilled lungs were 1.24 ± 0.32, 1.27 ± 0.24 and 2.01 ± 0.50 cells/mm2, and in saline-instilled lungs were 0.99 ± 0.17, 0.92 ± 0.13 and 1.64 ± 0.36 cells/mm2 in control (n = 11), diabetic (n = 8) and insulin-treated diabetic rats (n = 6) respectively. Neutrophils were rarely seen at this stage.
Figure 2.
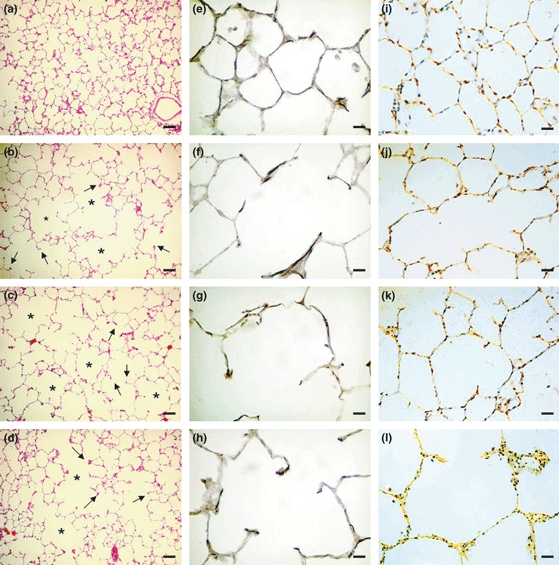
Photomicrographs of lung parenchyma histology 50 days after instillation of porcine pancreatic elastase (PPE, right lung) or saline (left lung). Haematoxylin–eosin staining (a–d) magnification ×100 (bar 200 μm): (a) saline-instilled lung showing the maintenance of the lobular and acinar architecture; (b) PPE-instilled lung from a control rat; note the hyperdistension of the alveolar ducts (*) with rupture of the alveolar septa (arrows); (c) PPE-instilled lung from a diabetic rat with distortion of the acinar architecture because of hyperdistension of the alveolar ducts (*) associated with the rupture of alveolar septa (arrows); and (d) PPE-instilled lung from insulin-treated diabetic rat showing similar histological pattern to PPE-instilled lung from control rat. Resorcin–fuchsin staining (e–h) magnification ×400 (bar 50 μm): (e) saline-instilled lung showing integrity of the elastic component in the alveolar walls, contrasting with disruption and thickening distribution of the fibres along the alveolar wall of PPE-instilled lung from a control rat (f); and decrease in PPE-instilled lung from a diabetic rat (g); note the restoration of the elastic system fibres pattern in PPE-instilled lung from insulin-treated diabetic rat (h). Picrosirius staining under polarized light (i–l) magnification ×400 (bar 50 μm): (i) note the weak orange-green birefringence of collagen fibres in saline-instilled lung compared to (j) PPE-instilled lung from a control rat; (k) PPE-instilled lung from a diabetic rat; and (l) PPE-instilled lung from insulin-treated rat.
Quantification of elastic and collagen fibres: role of insulin
Results, illustrated in Figure 3, show a reduction in the proportion of elastic fibres and an increase in the proportion of collagen fibres in the alveolar septa of PPE-instilled lungs. Saline-treated lungs showed a preserved architectural pattern of the elastic component. In contrast, instillation of PPE into the lungs of control rats demonstrated a large, sparse and fragmented bundle of elastic system fibres indicative of significant reduction in the percentage of alveolar septa elastin (Figure 3a). However, in PPE-treated lungs of diabetic rats, there was a further (30%) reduction in the proportion of elastin in lung parenchyma, compared with PPE-treated lungs of control rats. When diabetic rats were treated with 4 IU NPH insulin 2 h before instillation, followed by 2 IU/day for the next 50 days, the elastic system fibres pattern was restored in PPE-instilled lungs of insulin-treated diabetic rats to values attained in PPE-instilled lungs of control rats. No differences were observed in saline-instilled lungs among groups, showing a preserved pattern of the elastic component. Instillation of PPE into the lungs of control rats induced a significant increase in the percentage of collagen fibres in alveolar septa and diffuse increase of birefringence in pulmonary parenchyma when compared with saline-treated lungs. Similar results were observed in PPE-instilled lungs obtained from diabetic rats and from insulin-treated diabetic rats compared with the corresponding saline-treated lungs (Figure 3b). Representative photomicrographs of lung parenchyma show the pattern distribution of elastic (Figure 2e–h) and collagen (Figure 2i–l) fibres after instillation of saline or PPE into the lungs of the animals.
Figure 3.
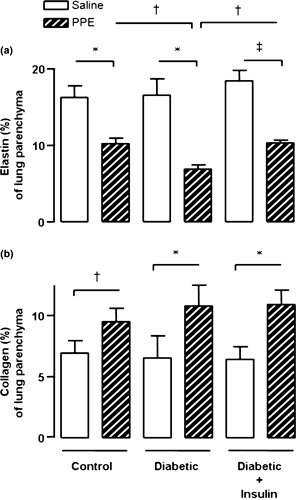
Fraction of area (%) occupied by elastin (a) and collagen (b) in the lung parenchyma of control (n = 8), diabetic (n = 7) and insulin-treated diabetic (n = 7) rats, 50 days after porcine pancreatic elastase (PPE, right lung) or saline (left lung) instillation. Diabetic rats were treated with 4 IU neutral protamine Hagedorn insulin, 2 h before instillation, followed by 2 IU/day for the next 50 days. Data are presented as means ± SEM and analysed by one-way anova followed by the Tukey–Kramer multiple comparisons test as follows: ‡P < 0.001, †P < 0.01 and *P < 0.05 compared with corresponding values in saline-treated lungs; †P < 0.01 compared with values in PPE-treated lungs from diabetic rats.
Discussion
Data presented here suggest that insulin modulates the inflammatory and repair responses of elastase-induced emphysema. The suggestion is supported by the following observations: (i) neutrophil infiltration into the lungs, which characterizes the inflammatory phase of pulmonary emphysema, was significantly reduced in alloxan-induced diabetic rats; (ii) measurements of the mean alveolar diameter, and quantification of elastic fibres, which characterize the reparative phase of emphysema, showed that the magnitude of the lesion was greater in diabetic animals; (iii) treatment of diabetic rats with insulin completely restored the magnitude of elastase-induced inflammation and restricted the size of the ongoing emphysematous lesion.
In the present study, pulmonary emphysema was obtained 50 days after instillation of PPE into the right lung of the animals, whereas the left lung instilled with saline was used as a control of the instillation procedure, without effects on the integrity of the lungs among diabetic, diabetic treated with insulin and matching control groups. Histological analysis of the lungs showed clear differences between PPE- and saline-instilled lungs including an increase in alveolar spaces consequent to the rupture of interalveolar septa, then satisfying the morphologic criteria that characterize emphysema (Groneberg & Chung 2004; Emami et al. 2008).
To investigate the influence of diabetes mellitus on the course of the emphysematous lesion, the initial inflammatory response of the animals to PPE instillation was evaluated by the presence of inflammatory cells in the BAL fluid. In diabetic rats, the number of neutrophils was reduced to 58% the values attained in PPE-treated lungs from control rats. Number of neutrophils was completely restored after treatment of diabetic rats with insulin. Despite a mean of 39% reduction in the levels of blood glucose after treatment with a single dose of insulin, diabetic rats still were hyperglycaemic compared with controls. It should be considered the choice of the dose and timing of insulin treatment used in the present study. Because the maximum serum concentration of NPH insulin is reached between 6 and 8 h after subcutaneous administration, we decided to give insulin to the animals 2 h before PPE instillation, as in previous studies (Martins et al. 2006, 2009; Alba-Loureiro et al. 2006). The dose of insulin was chosen based on its ability to restore the initial inflammatory response to elastase instillation into the lung of diabetic rats. Furthermore, increases in corticosterone levels in the blood of animals with an ongoing inflammatory reaction occur apparently without detectable change in plasma glucose (Moraes & Garcia-Leme 1982), and levels of serum corticosterone does not differ significantly between alloxan-induced diabetic rats and controls (Martins et al. 2006). Accordingly, the reduced neutrophil infiltration into the lungs in response to elastase instillation might be primarily linked to a continuing insulin deficiency rather than to the hyperglycaemic state of diabetic rats, or to a prevalence of glucocorticoids. Clinical and experimental evidence indicates that an overall reduced inflammatory reaction is observed whenever a relative lack of insulin occurs in an organism (Alba-Loureiro et al. 2007). In previously published studies on alloxan-induced diabetes, it was demonstrated that the local exudative cellular reaction in an inflammatory lesion, including carrageenan or tumour necrosis factor-α-induced injury (Pereira et al. 1987; Anjos-Valotta et al. 2006), the allergic airway inflammation (Vianna & Garcia-Leme 1995; Cavalher-Machado et al. 2004), lipopolysaccharide-induced acute lung inflammation (Martins et al. 2006), as well as the uptake of macromolecules into lymph nodes and lymphocyte recirculation (Moriguchi et al. 2005), depends on the availability of insulin. As demonstrated in the present study, a complete recovery of the response to PPE instillation was observed when diabetic rats were treated with a single dose of insulin that, nevertheless, was not sufficient to normalize the level of blood glucose.
To evaluate the action of insulin on the repair process of the emphysematous lesion, diabetic rats were treated with the same single dose of NPH insulin before elastase instillation, followed by daily doses of 2 IU NPH insulin for 50 days. At the end of the treatment, diabetic rats exhibited a reduction (32%) in the levels of blood glucose compared to non-treated diabetic rats, but animals still were hyperglycaemic compared to their matching controls. Values of mean alveolar diameter finding in saline-instilled lungs of all groups were similar and compatible with values described for Wistar rats (Faffe et al. 2002). Compared with saline instillation, the mean alveolar diameter increased in PPE-instilled lungs of all groups, accompanied by rupture of interalveolar septa that characterizes pulmonary emphysema (Monteiro et al. 2004). However, the magnitude of alveolar diameter in the emphysematous lung of diabetic rats was greater, and reduction of elastic fibres density was more evident compared with the other groups. Studies have demonstrated that degradation of elastin plays a key role in the initiation of pulmonary emphysema (Snider 1992; Houghton et al. 2006). Elastin fragments are important chemotactic factors for macrophages in the interstitium, septum and alveolar airspace of the lung, ultimately leading to enhanced lung tissue destruction by placing more macrophages, and thus more macrophage elastase within close proximity of the airspace (Houghton et al. 2006). However, data presented herein showed that there were no differences in the number of macrophages in the emphysema-like lesions among groups. Because neutrophils were rarely seen at this stage, other factors might be involved in the repair process associated with diabetes. The mechanisms by which insulin modulates progression of the disease need further investigation.
The breakdown and the degradation of collagen fibres have also been reported in both human patients (Stone et al. 1995) and animal models of emphysema (Wright & Churg 1995; Dhami et al. 2000). This degradation was accompanied by a remodelling of collagen fibres resulting in an abnormal repair process that contributes to the pathogenesis of the disease (Finlay et al. 1996; Lucey et al. 1998; Itu et al. 2005; Taraseviciene-Stewart & Voelkel 2008). In the present study, collagen fibres were increased in PPE-instilled lungs without differences between groups of animals. Indeed, PPE-instilled rodents exhibit a decrease in elastin content and an increase in collagen fibres content in alveolar septa (Itu et al. 2005), as demonstrated herein.
The capacity of the host to respond appropriately to pathogenic challenges appears to reside in the integrative function of endogenous glucocorticoids and insulin. When such regulatory activity fails, the control and neutralization of the noxious stimulus is impaired and the transitory reaction is prone to become a sustained and often deleterious event (Garcia-Leme & Farsky 1993; Alba-Loureiro et al. 2007). Indeed, the magnitude of the emphysematous lesion in diabetic rats was greater than that observed in matching controls, as depicted by the increase in mean alveolar diameter and the decrease in elastin in alveolar septa. When diabetic rats were daily treated with insulin, the final size of the lesion was reduced to values observed in control rats. These data obtained through histomorphometric analyses validate the current experimental model of pulmonary emphysema and emphasize the severity of the ongoing emphysematous lesion in insulin-deficient states. It should be considered that the presence of oxidative stress, endothelial dysfunction and cell apoptosis, common features of both diseases emphysema (Morissette et al. 2009) and diabetes mellitus (Van den Oever et al. 2010), might be implicated in the association of smoking and diabetes.
In conclusion, the data presented suggest that insulin modulates inflammatory and repair responses of elastase-induced emphysema, assuring normal repair and tissue remodelling. These findings strongly suggest that an early adequate response is essential to prevent or limit the intensity of a future lesion.
Acknowledgments
This work was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo and Pronex/CNPq. The authors thank Irene M. Gouveia for expert technical help.
References
- Alba-Loureiro TC, Martins EF, Landgraf RG, Jancar S, Curi R, Sannomiya P. Role of insulin on PGE2 generation during LPS-induced lung inflammation in rats. Life Sci. 2006;78:578–585. doi: 10.1016/j.lfs.2005.05.057. [DOI] [PubMed] [Google Scholar]
- Alba-Loureiro TC, Munhoz CD, Martins JO, et al. Neutrophil function and metabolism in individuals with diabetes mellitus. Braz. J. Med. Biol. Res. 2007;40:1037–1044. doi: 10.1590/s0100-879x2006005000143. [DOI] [PubMed] [Google Scholar]
- Anjos-Valotta EA, Martins JO, Oliveira MA, et al. Inhibition of tumor necrosis factor–α-induced intercellular adhesion molecule-1 expression in diabetic rats: role of insulin. Inflamm. Res. 2006;55:16–22. doi: 10.1007/s00011-005-0003-7. [DOI] [PubMed] [Google Scholar]
- Bar RS, Hoack JC, Peacock ML. Insulin receptors in human endothelial cells: identification and characterization. J. Clin. Endocrinol. Metab. 1978;47:699–702. doi: 10.1210/jcem-47-3-699. [DOI] [PubMed] [Google Scholar]
- Barnes PJ. Immunology of asthma and chronic obstructive pulmonary disease. Nat. Rev. Immunol. 2008;8:183–192. doi: 10.1038/nri2254. [DOI] [PubMed] [Google Scholar]
- Black PN, Ching PST, Beaumont B, Ranasinghe S, Taylor G, Merrilees MJ. Changes in elastic fibers in the small airways and alveoli in COPD. Eur. Respir. J. 2008;31:998–1004. doi: 10.1183/09031936.00017207. [DOI] [PubMed] [Google Scholar]
- Cavalher-Machado SC, de Lima WT, Damazo AS, et al. Down-regulation of mast cell activation and airway reactivity in diabetic rats: role of insulin. Eur. Respir. J. 2004;24:552–558. doi: 10.1183/09031936.04.00130803. [DOI] [PubMed] [Google Scholar]
- Chung KF, Adcock IM. Multifaceted mechanisms in COPD: inflammation, immunity, and tissue repair and destruction. Eur. Respir. J. 2008;31:1334–1356. doi: 10.1183/09031936.00018908. [DOI] [PubMed] [Google Scholar]
- Dhami R, Gilks B, Xie C, Zay K, Wright J, Churg A. Acute cigarette smoke-induced connective tissue breakdown is mediated by neutrophils and prevented by α1-antitrypsin. Am. J. Respir. Cell Mol. Biol. 2000;22:244–252. doi: 10.1165/ajrcmb.22.2.3809. [DOI] [PubMed] [Google Scholar]
- Emami K, Cadman RV, Woodburn JM, et al. Early changes of lung function and structure in an elastase model of emphysema—a hyperpolarized 3He MRI study. J. Appl. Physiol. 2008;104:773–786. doi: 10.1152/japplphysiol.00482.2007. [DOI] [PubMed] [Google Scholar]
- Faffe DS, Rocco MRP, Negri EM, Zin WA. Comparison of rat and mouse pulmonary tissue mechanical properties and histology. J. Appl. Physiol. 2002;92:230–234. doi: 10.1152/japplphysiol.01214.2000. [DOI] [PubMed] [Google Scholar]
- Finlay GA, O'Donnell MD, O'Connor CM, Hayes JP, Fitz Gerald MX. Elastin and collagen remodeling in emphysema: a scanning electron microscopy study. Am. J. Pathol. 1996;149:1405–1415. [PMC free article] [PubMed] [Google Scholar]
- Fullmer HM, Sheetz JH, Narkates AJ. Oxytalan connective tissue fibers: a review. J. Oral Pathol. 1974;3:291–316. doi: 10.1111/j.1600-0714.1974.tb01724.x. [DOI] [PubMed] [Google Scholar]
- Fussganger RD, Kahn CR, Roth J, De Meyts P. Binding and degradation of insulin by human peripheral granulocytes. Demonstration of specific receptors with high affinity. J. Biol. Chem. 1976;251:2761–2769. [PubMed] [Google Scholar]
- Garcia-Leme J, Farsky SP. Hormonal control of inflammatory responses. Mediat. Inflamm. 1993;2:181–198. doi: 10.1155/S0962935193000250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Groneberg DA, Chung KF. Models of chronic obstructive pulmonary disease. Respir. Res. 2004;5:18–34. doi: 10.1186/1465-9921-5-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haire-Joshu D, Glasgow RE, Tibbs TL. Smoking and diabetes. Diabetes Care. 1999;22:1887–1898. doi: 10.2337/diacare.22.11.1887. [DOI] [PubMed] [Google Scholar]
- Houghton AM, Quintero PA, Perkins DL, et al. Elastin fragments drive disease progression in a murine model of emphysema. J. Clin. Invest. 2006;116:753–759. doi: 10.1172/JCI25617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Itu S, Ingenito EP, Brewer KK, et al. Mechanics, nonlinearity, and failure strength of lung tissue in a mouse model of emphysema: possible role of collagen remodeling. J. Appl. Physiol. 2005;98:503–511. doi: 10.1152/japplphysiol.00590.2004. [DOI] [PubMed] [Google Scholar]
- Lucey EC, Goldstein RH, Stone PJ, Snider GL. Remodeling of alveolar walls after elastase treatment of hamsters: results of elastin and collagen mRNA in situ hybridization. Am. J. Respir. Crit. Care Med. 1998;158:555–564. doi: 10.1164/ajrccm.158.2.9705021. [DOI] [PubMed] [Google Scholar]
- Martins JO, Meyer-Pflug AR, Alba-Loureiro TC, et al. Modulation of lipopolysaccharide-induced acute lung inflammation: role of insulin. Shock. 2006;25:260–266. doi: 10.1097/01.shk.0000194042.18699.b4. [DOI] [PubMed] [Google Scholar]
- Martins JO, Zanoni FL, Martins DO, et al. Insulin regulates cytokines and intercellular adhesion molecule-1 gene expression through NF-kappaB activation in lipopolysaccharide-induced acute lung injury in rats. Shock. 2009;31:404–409. doi: 10.1097/SHK.0b013e318186275e. [DOI] [PubMed] [Google Scholar]
- Monteiro R, Jatene FB, Pazetti R, et al. Evaluation of the cardiac morphological alterations secondary to the pulmonary emphysema: experimental study in rats. Braz. J. Cardiovasc. Surg. 2004;19:341–347. [Google Scholar]
- Montes GS. Structural biology of the fibers of the collagenous and elastic system. Cell Biol. Int. 1996;20:15–27. doi: 10.1006/cbir.1996.0004. [DOI] [PubMed] [Google Scholar]
- Moraes FR, Garcia-Leme J. Endogenous corticosteroids and insulin in acute inflammation. Microvasc. Res. 1982;23:281–293. doi: 10.1016/s0026-2862(82)80001-0. [DOI] [PubMed] [Google Scholar]
- Moriguchi P, Sannomiya P, Lara PF, Oliveira-Filho RM, Greco KV, Sudo Hayashi LS. Lymphatic system changes in diabetes mellitus: role of insulin and hyperglycemia. Diabetes Metab. Res. Rev. 2005;21:150–157. doi: 10.1002/dmrr.500. [DOI] [PubMed] [Google Scholar]
- Morissette M, Parent J, Milot J. Alveolar epithelial cell apoptosis in emphysema: what we know and what we need to know. Int. J. Chron. Obstruct. Pulmon. Dis. 2009;4:19–31. [PMC free article] [PubMed] [Google Scholar]
- Pereira MA, Sannomiya P, Garcia-Leme J. Inhibition of leukocyte chemotaxis by factor in alloxan-induced diabetic rat plasma. Diabetes. 1987;36:1307–1314. doi: 10.2337/diab.36.11.1307. [DOI] [PubMed] [Google Scholar]
- Snider GL. Emphysema: the first two centuries-and beyond. A historical overview, with suggestion for future research: part 2. Am. Rev. Respir. Dis. 1992;146:1615–1622. doi: 10.1164/ajrccm/146.6.1615. [DOI] [PubMed] [Google Scholar]
- Stone PJ, Gottlier DJ, O'Connor GT, et al. Elastin and collagen degradation products in urine of smokers with and without chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 1995;151:952–959. doi: 10.1164/ajrccm.151.4.7697272. [DOI] [PubMed] [Google Scholar]
- Taraseviciene-Stewart L, Voelkel NF. Molecular pathogenesis of emphysema. J. Clin. Invest. 2008;118:394–402. doi: 10.1172/JCI31811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van den Oever IAM, Raterman HG, Nurmohamed MT, Simsek S. Endothelial dysfunction, inflammation, and apoptosis in diabetes mellitus. Mediat. Inflamm. 2010;2010:792393–792408. doi: 10.1155/2010/792393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vianna EO, Garcia-Leme J. Allergen-induced airway inflammation in rats: role of insulin. Am. J. Respir. Crit. Care Med. 1995;151:809–814. doi: 10.1164/ajrccm/151.3_Pt_1.809. [DOI] [PubMed] [Google Scholar]
- Weibel ER. Morphometry: stereological theory and practical methods. In: Gil J, editor. Models of Lung Disease-Microscopy and Structural Methods. New York: Marcel Dekker; 1990. pp. 199–247. [Google Scholar]
- Wright JL, Churg A. Smoke-induced emphysema in guinea pig is associated with morphometric evidence of collagen breakdown and repair. Am. J. Physiol. Lung Cell. Mol. Physiol. 1995;268:L17–L20. doi: 10.1152/ajplung.1995.268.1.L17. [DOI] [PubMed] [Google Scholar]
- Wright JL, Cosio M, Churg A. Animal models of chronic obstructive pulmonary disease. Am. J. Physiol. Lung Cell. Mol. Physiol. 2008;295:L1–L15. doi: 10.1152/ajplung.90200.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]