

Abstract
Studies of inhalational melioidosis were undertaken in the common marmoset (Callithrix jacchus). Following exposure to an inhaled challenge with aerosolized Burkholderia pseudomallei, lethal infection was observed in marmosets challenged with doses below 10 cfu; a precise LD50 determination was not possible. The model was further characterized using a target challenge dose of approximately 102 cfu. A separate pathogenesis time-course experiment was also conducted. All animals succumbed, between 27 and 78 h postchallenge. The challenge dose received and the time to the humane endpoint (1 °C below normal body temperature postfever) were correlated. The first indicator of disease was an increased core body temperature (Tc), at 22 h postchallenge. This coincided with bacteraemia and bacterial dissemination. Overt clinical signs were first observed 3–5 h later. A sharp decrease (typically within 3–6 h) in the Tc was observed prior to humanely culling the animals in the lethality study. Pathology was noted in the lung, liver and spleen. Disease progression in the common marmoset appears to be consistent with human infection in terms of bacterial spread, pathology and physiology. The common marmoset can therefore be considered a suitable animal model for further studies of inhalational melioidosis.
Keywords: animal model, Burkholderia pseudomallei, marmoset, non-human primate, pathology
Burkholderia pseudomallei is the causative agent of melioidosis that is a major cause of morbidity and mortality in Southeast Asia and northern Australia (Currie et al. 2008). Disease presentation is highly variable depending on factors such as strain of bacteria, route of transmission and health status of the individual (Poe et al. 1971). There are currently no vaccines for melioidosis, and the disease is difficult to treat using antibiotics because of an inherent resistance to many antibiotics (White 2003). The understanding of disease pathogenesis and the appropriate use of animal models is an important consideration when testing the efficacy of treatments such as vaccines or antibiotics. Many animal models have been used to study B. pseudomallei (Titball et al. 2008). However, there are no published data characterising non-human primate (NHP) models of inhalational melioidosis.
Non-human primates have been shown to be susceptible to melioidosis, and the disease has been identified in imported monkeys in the UK (Dance et al. 1992) and the USA (Kaufmann et al. 1970; Britt et al. 1981; Fritz et al. 1986). Naturally occurring melioidosis has been reported in captive animals in several places including Malaysia (Retnasabapathy & Joseph 1966; Strauss et al. 1969; Mutalib et al. 1984) and Australia (Tammemagi & Johnston 1963). There are two reports of early studies of experimental melioidosis infection of NHPs (Stanton & Fletcher 1925; Miller et al. 1948). During the 1920s, Macacus cynomolgus were infected orally with B. pseudomallei; however, no details of the infection were provided (Stanton & Fletcher 1925). Subsequently, Macacus mulata were subcutaneously infected with various doses of B. pseudomallei with only the animal receiving the highest concentration of bacteria (1.5 × 106 cfu) showing any clinical signs (Miller et al. 1948). The highest dose animal had an abscess at the site of inoculation that was self-draining after 4 days and completely healed within 2 weeks, an elevated temperature and an increase in some clinical parameters such as white blood cell count, all of which subsided with the draining of the abscess. The animal survived for the 2 month duration of the study.
Recently, the common marmoset (Callithrix jacchus), a New World NHP species, has been developed as an infection model for anthrax and tularaemia (Lever et al. 2008; Nelson et al. 2009, 2010). Marmosets offer many advantages for studying highly infectious agents including their close molecular and immunological relationship with man (‘t Hart et al. 2004), their suitability for use in the development of drugs (Orsi et al. 2011), their small size allowing ethical, safe housing within biocontainment restraints, as well as their low cost and availability.
This work focuses on developing the common marmoset as a model of experimental inhalational melioidosis. The disease has been characterized in the context of survival, core body temperature (Tc), activity, bacteriology, histopathology, immunology and clinical parameters.
Materials and Methods
Animals
Healthy sexually mature common marmosets (C. jacchus) were obtained from the Dstl Porton Down breeding colony and housed in vasectomized male and female pairs. For lethality studies, animals were aged between 24 and 39 months old and weighed between 380 and 512 g at the time of challenge. Animals used in the pathogenesis studies were aged between 24 and 58 months old and weighed between 314 and 532 g at the time of challenge. All animals were allowed free access to food and water as well as environmental enrichment. All animals were surgically implanted intra-peritoneally with a Remo 200 device under general anaesthesia (Ketamine/Isofluorane) to record Tc, as previously described (Nelson et al. 2009). Animals had Actiwatch®-Mini devices (Cambridge Neurotechnology Ltd, Cambridge, UK) attached to their collars to monitor activity. All animal studies were carried out in accordance with the UK Animals (Scientific Procedures) Act of 1986 and the Codes of Practice for the Housing and Care of Animals used in Scientific Procedures 1989. Following challenge with B. pseudomallei, all animals were handled under animal containment level 3 (CL3) conditions, within a half-suit isolator compliant with British Standard BS5726.
Bacterial strain and culture
Glycerol stocks of B. pseudomallei strain K96243 were provided by the Health Protection Agency (HPA), UK. Bacteria were recovered into Luria-Bertani (LB) broth and incubated at 37 °C with shaking at 180 rpm for 24 h, prior to recovery into phosphate buffered saline (PBS), pH 7.3. The optical density reading (OD590) of the suspension was adjusted to 0.35, equivalent to approximately 1 × 108 cfu/ml. The suspension was serially diluted to the appropriate concentration for challenge. Viable counts were performed on LB-agar plates retrospectively to determine the actual value. All procedures were carried out at Laboratory CL3, in Class 3 microbiological safety cabinets.
Challenge
Marmosets were anaesthetized with 25 mg/kg ketamine intra-muscularly prior to exposure and were challenged in pairs by the airborne route, as previously described (Lever et al. 2008; Nelson et al. 2009). Conditioned aerosol particles of approximately 1–3 μm were generated using a collison nebuliser (containing 20 ml B. pseudomallei and three drops of Antifoam 289 (Sigma, Gillingham, Dorset, UK) in conjunction with a modified Henderson apparatus.
Lethality studies
The Dixon staircase method was used as previously described (Lever et al. 2008). A log decrease in colony-forming units (cfu) was used in four sequential studies, each using two animals, starting from a target estimated challenge dose of approximately 103 cfu, followed by approximately 102, 10 and 1 cfu. For reproducibility, two further studies were carried out at the 102 cfu target dose.
Pathogenesis studies
Ten marmosets received an inhaled dose range of 135–440 cfu of B. pseudomallei strain K96243. Pairs of animals were culled at 0, 10, 22, 36 and 46 h postchallenge (p.c.).
Postmortem analysis
Postmortem examinations were performed on all animals in both studies; organs removed were assessed for bacteriology, immunology, gross pathology and histopathology. Blood was removed from anaesthetized marmosets by cardiac puncture for assessment of bacteraemia, clinical chemistry, haematology and immunology.
Bacteriology
Bacterial loads were determined in blood, liver, spleen, kidneys, brain. Organs were removed aseptically and processed as previously described (Nelson et al. 2009). Appropriate dilutions were sub-cultured onto LB-agar plates and incubated at 37 °C for 24 h. Counts are expressed as cfu/g of tissue or cfu/ml of blood.
Cell type determination
Cell type was assessed as previously described (Nelson et al. 2009). Cell type levels are expressed as a percentage of viable population.
Histopathology
Tissues were fixed in 10% neutral buffered formalin and processed for paraffin wax embedding using standard techniques. Thin sections (5 μm) were cut and stained with haematoxylin and eosin for histopathological analysis.
Immunohistochemistry
Sections of formalin fixed wax embedded tissues were prepared and dried onto polylysine slides (VWR Ltd., East Grinstead, West Sussex, UK). Sections were rehydrated in xylene, graded alcohol solutions and water before incubating at 37 °C for 10 min in antigen retrieval solution consisting of 0.05% (w/v) pronase XIV (5.2 U/mg Sigma P5147-1G) in PBS. Sections were incubated (20 °C for 30 min) to block for secondary antibody binding using rabbit serum (Abcam, Cambridge, Cambridgeshire, UK ab7487 diluted 1:10 in PBS) followed by 10 min in 3% H2O2 to block endogenous peroxidase activity. Slides were incubated at 4 °C overnight in a humidifier tray with a B. pseudomallei anti-capsular antibody (3VIE5 at 2 μg/ml) (Jones et al. 2002) then incubated at 20 °C with 7 μg/ml biotinylated polyclonal secondary rabbit anti-mouse IgG antibody (Abcam ab6727-1). Slides were treated with streptavidin-HRP conjugate (Abcam ab64269-125) and antigen binding visualized using diamino benzidine (DAB) as Horse-radish peroxidase (HRP) substrate (Sigma staining kit D-4293). To test for primary antibody non-specific binding, the primary 3VIE5 antibody was replaced with a mouse IgG anti-Francisella tularensis antibody, a pathogen not anticipated to be present in any of the tissues. Absence of non-specific secondary antibody binding was confirmed by omission of the 3VIE5 antibody in the staining procedure. All DAB stained tissue sections were counterstained in haematoxylin for the visualization of cell nuclei then dehydrated in graded alcohols and xylene and coverslipped with DPX mountant (Bios Europe, Skelmersdale, Lancashire, UK).
Haematology, clinical chemistry and electrolytes
Blood was collected at defined time-points into tubes containing EDTA, and key parameters were measured by use of a laser-flow cytometry-based haematological analyser (LaserCyte; IDEXX Laboratories Ltd, Bucks, UK). Clinical chemistry parameters were analysed using a ‘dry-slide’ technology biochemistry analyser (VetTest; IDEXX). Electrolyte concentrations were measured using an ion-selective electrode-based electrolyte analyser (VetLyte; IDEXX). Postchallenge data were compared with mean prechallenge results.
Statistics
Pearson's correlation analysis was used to determine the relationship of gender, body weight, time to death and inhaled dose. Comparative analysis of activity, bacteriology, immunological and blood chemistry data was performed using two-way anova. Regression analysis using natural logged data was performed.
Results
There were no correlations between clinical outcome compared with weight, age or gender and either time to death.
Lethality studies
Six pairs of animals were challenged with various doses of aerosolized B. pseudomallei. In an initial susceptibility study (Step 1), two animals were exposed to a calculated inhaled dose of 2.6 × 103 cfu [male (M)] or 7.7 × 103 cfu [female (F)]. Animals were culled based on clinical signs at 53 and 48 h postchallenge (p.c.) respectively. A second pair of animals received a log lower challenge dose [1.8 × 102 (M) and 1.5 × 102 (F) cfu] and were culled at 55 and 57 h respectively (Step 2). A third pair received 25 (M) and 63 (F) cfu (Step 3); they were culled at 59 and 64 h respectively. A fourth pair received a further log lower challenge dose [2 (M) and 5 (F) cfu] (Step 4) and were culled at 78 and 74 h p.c. (Figure 1).
Figure 1.
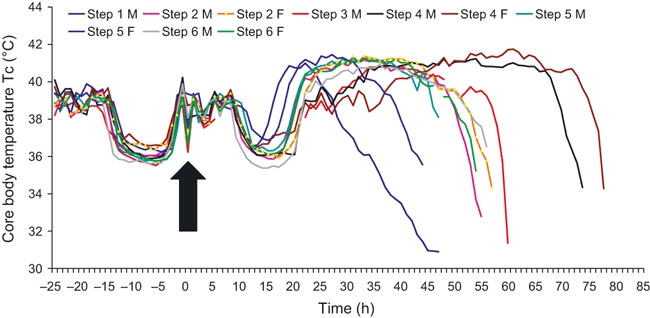
Core body temperature of individual marmosets challenged with Burkholderia pseudomallei by the airborne route (Step 1 F and Step 3 F data not presented because of failure of implant prior to the study). Temperature was measured using a Remo 200 telemetry device surgically implanted into the peritoneum of the animals.
As it was not possible to establish precisely an LD50 value, repeat studies were performed to determine the reproducibility of the 102 cfu target challenge dose (Steps 5 and 6). An average of 334 ± 78 cfu was achieved. Time to death was 44–57 h (mean 52 ± 2).
Clinical signs and temperature profile
Noticeable features were the uniform lethality, temperature profile and clinical signs. All animals receiving a dose above 10 cfu exhibited clinical signs within 24 h, following a temperature increase. Animals became more subdued and somnolent, with increasing dyspnoea, particularly by 48 h. Piloerection, hunching and an increased respiratory rate were also noted at this time. Clinical signs appeared 24 h later in animals receiving less than 10 cfu. All animals were culled based on the severity of the above signs, (specifically major dyspnoea) and reluctance to respond to external stimuli.
All animals exhibited a normal Tc profile prechallenge (Figure 1). Postchallenge all animals followed a similar Tc trend: a rapid increase, a plateau at approximately 41 °C, followed by a rapid temperature decline in temperature. Time to onset and subsequent decline of the febrile response varied with challenge dose. Animals challenged with 103 cfu showed an increase in Tc at approximately 14 h and a decline at 28 h. In the six animals challenged with approximately 102 cfu, fever occurred at 18–25 h (mean 21) p.c. and the Tc drop at 45–48 h (mean 47). Fever and subsequent Tc drop occurred at 29 and 59 h (10 cfu challenge) and at 32–35 and 71–78 h (>10 cfu) respectively.
Data were standardized so that the study endpoint was the time when, after a period of fever, a 1 °C decrease in Tc below normal was observed. This was also indicative of a rapid decline in the animals’ condition (specifically clinical signs, behaviour and continuing temperature decline) and was considered to be the humane endpoint for future studies.
There was a highly significant linear correlation (R2 = 0.927, P = 0.03) between dose and time to the humane endpoint.
Pathogenesis studies
The first two animals were culled immediately postexposure (t = 0 h). At t = 10, animals appeared normal with no apparent clinical signs or changes in behaviour. At t = 22, the Tc was increasing (approximately 1 °C above normal, either 40.9 or 40 °C). At t = 36, Tc had plateaued and some clinical signs were apparent. By t = 46, Tc had decreased by 1 °C compared with baseline (36.4 °C).
Activity
Daily activity, compared with the three days prechallenge, decreased with time postchallenge. A slower decline in activity was noted with lower challenge doses (P < 0.0001) (Figure 2).
Figure 2.
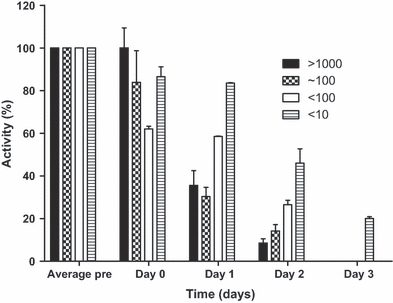
The activity of marmosets challenged with various doses of Burkholderia pseudomallei strain K96243 during lethality studies. Activity was measured using a collar-based Actiwatch®-Mini device. n = 2 in all groups except the 102 cfu group where n = 6. Error bars represent the SE of the mean.
Bacteriology
In the lethality studies, higher counts were recovered from the spleens and lungs (Figure 3a). There was no statistically significant relationship between the bacterial load and either the initial challenge dose or time to death.
Figure 3.
Bacterial counts recovered from marmoset organs. (a) lethality studies where n = 2 in all groups except the 102 cfu group where n = 6, (b) pathogenesis studies where two animals were killed at various times postchallenge. Error bars represent the SE of the mean.
Bacterial counts in the pathogenesis study rose with time and are shown in Figure 3(b). Bacteraemia was detected at t = 22.
Haematology and blood chemistry
Results are shown in Figure 4. Decreased platelet (P < 0.001), red (P < 0.01) and white blood cell counts (P < 0.05) (Figure 4b) were observed from t = 36. Potassium concentrations were elevated at t = 10–10.2 mmol/l (P < 0.001) and at t = 22–8.3 mmol/l (P < 0.001). Aspartate transferase concentrations rose by t = 10, (P < 0.001) and remained elevated. Alanine transferase (ALT) and alkaline phosphatase (ALKP) concentrations were increased at t = 36 (both P < 0.01), whilst creatine kinase and lactate concentrations were raised by t = 22 (P < 0.001 and P < 0.01 respectively).
Figure 4.
Key blood parameters from marmosets infected with approximately 102 cfu of Burkholderia pseudomallei strain K96243 and culled at specific times postchallenge. (a) Liver function enzymes (alanine transferase, aspartate transferase and alkaline phosphatase), measured using the VetTest Analsyser, IDEXX Laboratories Ltd (b) Haematology parameters (Red blood cell count × 106/μl, White blood cell and Platelet counts × 103/μl) measured using LaserCyte Analyser, IDEXX Laboratories Ltd.
Immunology
Changes in the proportions of NK cells, neutrophils and macrophages in the blood, lung and spleen were compared with baseline (naïve) levels (t = 0) (Figure 5). There was an increase in the peripheral neutrophil count (P < 0.001) by t = 36, followed by a rapid decrease by t = 46 (P < 0.01). This was associated with an increase in the levels in the lungs (P < 0.05) at this time (Figure 5a). Peripheral blood macrophage counts gradually increased until t = 46 (P < 0.001).
Figure 5.
The progression of the cellular populations in the lungs, spleen and blood of marmoset organs infected with approximately 102 cfu of inhalational Burkholderia pseudomallei strain K96243 (n = 2 for all time-points) measured using flow cytometry. (a) neutrophils, (b) macrophages, (c) NK cells. Cell type levels are expressed as a percentage of the viable cell population.
Lung NK cell numbers peaked at t = 10 and declined to below baseline by t = 36 (not significant). There was a decrease in T-cell numbers in the spleen at t = 36 to 8.8% (P < 0.01) followed by an increase in the B-cell population at t = 46 to 32% (P < 0.01).
Pathology and immunohistochemistry
Slight haemorrhage was apparent in the lungs at t= 10. This increased in severity and extensive haemorrhaging was observed at t = 46, at which time the lungs were enlarged and firmer than normal. By t = 46, numerous small hepatic necrotic foci were evident. Most animals had splenomegaly (83%), with small necrotic foci observed in 50%. No gross pathology was observed in the kidneys or the brain.
The primary histopathology was a necrotising pneumonia followed by local lymphadenitis. Multifocal necrotising pneumonia with degenerated and viable neutrophils and some epithelioid macrophages, lymphocytes and plasma cells was evident in all animals from t = 22 (Figure 6a). This tended to progress to coalescent necrotising pneumonia in the lethality study. Diffuse extensive alveolar oedema and haemorrhages were evident from t = 46.
Figure 6.
Pathological features from marmosets challenged with approximately 102 cfu of Burkholderia pseudomallei strain K96243 by the airborne route. (a) Typical necrotising pneumonia and oedema exhibited in the lungs of marmosets (H & E, 40×), (b) Typical necrotising hepatitis exhibited in the livers of marmosets (H & E, 100×) (c) Typical necrotising lymphadenitis in the lymph nodes of marmosets (H & E, 40×), (d) Typical necrotising splenitis in the spleens of marmosets (H & E, 100 × 10× magnification), (e) Immunohistochemistry of section (a) using an B. pseudomallei anti-capsular antibody (40×), (f) Immunohistochemistry of section (c) using an B. pseudomallei anti-capsular antibody (40×).
Multifocal necrotising hepatitis with multiple degenerated and viable neutrophils, lymphocytes, epitheloid macrophages and plasma cells occurred by t = 46 (Figure 6b). All animals in the lethality study exhibited multifocal to coalescent necrotising lymphadenitis affecting the majority of the tissue (Figure 6c) and multifocal necrotising splenitis with multiple foci showing degenerated neutrophils and necrotic debris, mainly within the splenic red pulp (Figure 6d). Bacteria were associated with necrotic centres in the lungs, lymph tissue and liver (Figure 6e,f).
No brain histopathology was observed and mild interstitial mononuclear cell infiltrates in the kidney, which was deemed background renal pathology.
Discussion
The susceptibility of the common marmoset to airborne infection with B. pseudomallei, including the lethal nature of the infection, has been demonstrated. Marmosets appear highly susceptible to the bacterium and exhibit a rapid acute disease. It was not possible to determine a precise LD50 in this study because of the high infectivity, although data suggest it is <10 cfu.
There are very few published data on experimental infection of NHPs with B. pseudomallei. Macaques are relatively resistant to oral and subcutaneous infection (Stanton & Fletcher 1925; Miller et al. 1948). Naturally occurring cases have been described in NHPs in endemic regions and in imported monkeys (Sprague & Neubauer 2004). Clinical findings include skin abscesses and organ lesions, with a variable incubation period (6 months to 3 years) (Sprague & Neubauer 2004).
In the marmoset, illness was acute onset (within 24 h) and rapidly progressive. The data are consistent with acute-onset sepsis. All animals succumbed to infection after becoming bacteraemic, with widespread bacterial dissemination. This concurs with murine and human data (White 2003; Lever et al. 2009). Melioidosis in the marmoset was associated with high fever, pulmonary distress and hepatic dysfunction, consistent with human disease (White 2003; Leelarasamee 2004). The high mortality in this presentation of the disease, as well as hepatosplenomegaly, hepatic necrosis and lymphadenopathy, is all features of acute human melioidosis (Dance 1996; Bossi et al. 2004; Cheng & Currie 2005; Jansen et al. 2005). Poor prognostic indicators in humans, including fever, leucopoenia, abnormal liver function enzymes and pneumonia (Jansen et al. 2005), were all present in the later stages of infection in the marmoset. Leucopoenia (and specifically, neutropenia) was observed in the marmosets from 46 h postchallenge. Another indicator of poor prognosis in humans is a high activation of the coagulation system (Wiersinga et al. 2008). In that study, human septic melioidosis was associated with a significantly enhanced coagulation as determined by, amongst other things, a prolonged prothrombin time and activated partial thromboplastin time. This has also been observed in marmosets with severe melioidosis (Nelson, unpublished observation). These features are associated with disseminated intravascular coagulation that is a severe condition causing a rapid utilization of clotting factors, including platelets. Thrombocytopenia was evident in the current marmoset study from around 36 h postchallenge.
Melioidosis has been called ‘The Remarkable Imitator’ (Poe et al. 1971) because of its variability in clinical presentation, which can occur for many reasons including the route of infection and strain of infecting bacteria. The disease is more prevalent during the wet seasons in Australia and Thailand (Currie & Jacups 2003; White 2003) and also is commonly associated with natural disasters such as typhoons and tsunamis (Chierakul et al. 2005; Chou et al. 2007). Interestingly, after Typhoon Haitang struck Southern Taiwan in July 2005, there were a significantly higher number of cases of melioidosis presenting with bacteraemia and pneumonia and a significant increase in the number of deaths from this disease (Currie & Jacups 2003).
In humans, the incubation period varies greatly, usually 1–21 days, although overt disease may occur many years after exposure (Howe et al. 1971). The incubation period is a factor of both the route of infection and the inoculum size. Disease outcome, including survival, severity and time to death, also varies in experimental murine infections, depending on the route of administration, the dose and the strain of bacteria. Murine disease presentation also varies with the strain of mice; C57Bl/6 mice are more resistant to intra-peritoneal infection than Balb/c mice (Hoppe et al. 1999). However, when the pathogen is administered by the intra-nasal or inhalational routes, the pathogen is more virulent, resulting in death after 2 and 3 days and having a low LD50 of between 5–30 cfu regardless of the strain of mouse (Jeddeloh et al. 2003; Titball et al. 2008; Lever et al. 2009). This is consistent with the acute inhalational marmoset model developed here.
Naturally occurring airborne B. pseudomallei infections have been reported, usually associated with the inhalation of dust or water particles (as reported in Vietnam War helicopter crews) or in near-drowning cases (Howe et al. 1971; Lee et al. 1985). However, the infectious dose for humans by the inhalation route remains undetermined, although there is some evidence to suggest that in near-drowning incidents, the higher the dose the shorter the incubation period (Chierakul et al. 2005). Interestingly, in marmosets, there was a correlation between challenge dose and the time to death. This phenomenon has also been retrospectively analysed in a variety of small animal models of melioidosis (Tammemagi & Johnston 1963).
In this study, marmosets did not exhibit clinical signs until the rapid increase in temperature between 20 and 24 h postchallenge; this coincided with an increase in the number of bacteria isolated from all organs and the presence of bacteraemia. There was haematogenous spread from the lungs to the liver and spleen, followed by kidney and brain, which is consistent with murine inhalational data and human disease (Jeddeloh et al. 2003; White 2003; Lever et al. 2009). At all times, the highest number of bacteria was present in the lungs, again consistent with the murine inhalational model (Liu et al. 2002; Lever et al. 2009). The lung is the organ involved most frequently in human B. pseudomallei infection, regardless of the route of entry (at least 50% of cases) (White 2003).
An interesting finding was the dynamics of the neutrophil population during the study. There was a rapid increase in the blood at 36 h followed by neutropenia and recruitment to the lungs at 46 h p.c. This has also been observed in murine models of melioidosis (Dr R. Lukaszeski, Personal Communication). This may be a result of normal neutrophil dynamics, resulting in increased recruitment of the neutrophils following early cytokine activation leading to neutropenia because of acute massive sequestration to the lung tissue. However, numerous immature neutrophils were observed under the light microscope in the lung, liver and spleen of these animals suggesting premature release of the cells from the bone marrow because of high demand. Future studies will examine the bone marrow to confirm this hypothesis. The importance of neutrophils in resistance to B. pseudomallei infection in mice has been demonstrated resulting in controlling bacterial growth (Liu et al. 2002; Easton et al. 2007). They have also been shown to have a critical role in the generation of an early pro-inflammatory cytokine response in the lungs (Barnes et al. 2001). The susceptibility of the marmoset to B. pseudomallei infection may, in part, be due to the host's inability to produce mature neutrophils at an appropriate rate. Therefore, allowing bacterial growth potentially leads to the immunopathology that finally overwhelms the host.
Histopathological changes, typically necrotizing inflammation, were evident in the lung, liver, spleen and lymph nodes, as also observed in human and murine studies (Piggott & Hochholzer 1970; Wong 1995; Lever et al. 2009). In agreement with human melioidosis and murine studies, pathology was not observed in the kidney or brain of marmosets. In contrast to earlier murine studies (Lever et al. 2009), bacteria associated with necrotising lesions were identified in all organ types by immunohistochemistry, despite using the same monoclonal antibody raised against the bacterial capsule.
In conclusion, the common marmoset is highly susceptible to challenge with inhaled B. pseudomallei, and the disease is consistent with human acute-onset melioidosis (White 2003). The clinical presentation, bacterial spread, immunology and pathology of melioidosis in the marmoset mimic human infection. Therefore, the authors suggest that the common marmoset may be a useful animal model for the investigation of therapeutics for treatment of inhalational melioidosis.
Acknowledgments
The authors would like to thank Dr Kristin DeBord and Allen Roberts for their support. The authors would like to thank all Dstl employees that helped support this work, including Jan Platt for histopathological support. We also would like to thank Donna Hopkins and Graham Hatch at the HPA for performing the clinical analyses of blood samples. Funding: This project has been funded with Federal funds from the National Institute of Allergy and Infectious Diseases (NIAID), under contract No. N01-AI-30062 through the Prime Contractor, the Health Protection Agency, Porton Down, UK.
References
- Barnes JL, Ulett GC, Ketheesan N, Clair T, Summers PM, Hirst RG. Induction of multiple chemokine and colony-stimulating factor genes in experimental Burkholderia pseudomallei infection. Immunol. Cell Biol. 2001;79:490–501. doi: 10.1046/j.1440-1711.2001.01038.x. [DOI] [PubMed] [Google Scholar]
- Bossi P, Tegnell A, Baka A, et al. Bichat guidelines for the clinical management of glanders and melioidosis and bioterrorism-related glanders and melioidosis. Euro. Surveill. 2004;9:E17–E18. [PubMed] [Google Scholar]
- Britt JO, Howard EB, Kean CJ, Jones J. Melioidotic osteomyelitis in an imported primate. J. Am. Vet. Med. Assoc. 1981;179:1303–1305. [PubMed] [Google Scholar]
- Cheng AC, Currie BJ. Melioidosis: epidemiology, pathophysiology, and management. Clin. Microbiol. Rev. 2005;18:383–416. doi: 10.1128/CMR.18.2.383-416.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chierakul W, Winothai W, Wattanawaitunechai C, et al. Melioidosis in 6 tsunami survivors in southern Thailand. Clin. Infect. Dis. 2005;41:982–990. doi: 10.1086/432942. [DOI] [PubMed] [Google Scholar]
- Chou DW, Chung KM, Chen CH, Cheung BM. Bacteremic melioidosis in southern Taiwan: clinical characteristics and outcome. J. Formos. Med. Assoc. 2007;106:1013–1022. doi: 10.1016/S0929-6646(08)60077-7. [DOI] [PubMed] [Google Scholar]
- Currie BJ, Jacups SP. Intensity of rainfall and severity of melioidosis, Australia. Emerg. Infect. Dis. 2003;9:1538–1542. doi: 10.3201/eid0912.020750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Currie BJ, Dance DA, Cheng AC. The global distribution of Burkholderia pseudomallei and melioidosis: an update. Trans. R. Soc. Trop. Med. Hyg. 2008;102(Suppl. 1):S1–S4. doi: 10.1016/S0035-9203(08)70002-6. [DOI] [PubMed] [Google Scholar]
- Dance DAB. Melioidosis and glanders. In: Weatherall DJ, Ledingham CG, Warrell DA, editors. Oxford Textbook of Medicine. Oxford: Oxford Medical Publications; 1996. pp. 590–605. [Google Scholar]
- Dance DA, King C, Aucken H, Knott CD, West PG, Pitt TL. An outbreak of melioidosis in imported primates in Britain. Vet. Rec. 1992;130:525–529. doi: 10.1136/vr.130.24.525. [DOI] [PubMed] [Google Scholar]
- Easton A, Haque A, Chu K, Lukaszewski R, Bancroft GJ. A critical role for neutrophils in resistance to experimental infection with Burkholderia pseudomallei. J. Infect. Dis. 2007;195:99–107. doi: 10.1086/509810. [DOI] [PubMed] [Google Scholar]
- Fritz PE, Miller JG, Slayter M, Smith TJ. Naturally occurring melioidosis in a colonized rhesus monkey (Macaca mulatta) Lab. Anim. 1986;20:281–285. doi: 10.1258/002367786780808749. [DOI] [PubMed] [Google Scholar]
- ‘t Hart BA, Laman JD, Bauer J, Blezer E, van Kooyk Y, Hintzen RQ. Modelling of multiple sclerosis: lessons learned in a non-human primate. Lancet Neurol. 2004;3:588–597. doi: 10.1016/S1474-4422(04)00879-8. [DOI] [PubMed] [Google Scholar]
- Hoppe I, Brenneke B, Rohde M, et al. Characterisation of a murine model of melioidosis: comparison of different strains of mice. Infect. Immun. 1999;67:2891–2900. doi: 10.1128/iai.67.6.2891-2900.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howe C, Sampath A, Spotnitz M. The pseudomallei group: a review. J. Infect. Dis. 1971;124:598–606. doi: 10.1093/infdis/124.6.598. [DOI] [PubMed] [Google Scholar]
- Jansen A, Hagen RM, Schmidt W, et al. Pulmonary melioidosis associated with bronchiolitis obliterans organizing pneumonia. J. Infect. 2005;50:68–71. doi: 10.1016/j.jinf.2003.10.012. [DOI] [PubMed] [Google Scholar]
- Jeddeloh JA, Fritz DL, Waag DM, Hartings JM, Andrews GP. Biodefense-driven murine model of pneumonic melioidosis. Infect. Immun. 2003;71:584–587. doi: 10.1128/IAI.71.1.584-587.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones SM, Ellis JF, Russell P, Griffin KF, Oyston PC. Passive protection against Burkholderia pseudomallei infection in my by monoclonal antibodies against capsular polysaccharide, lipopolysaccharide or proteins. J. Med. Microbiol. 2002;52:1055–1062. doi: 10.1099/0022-1317-51-12-1055. [DOI] [PubMed] [Google Scholar]
- Kaufmann AF, Alexander AD, Allen MA, et al. Melioidosis in imported non-human primates. J. Wildl. Dis. 1970;6:211–219. doi: 10.7589/0090-3558-6.4.211. [DOI] [PubMed] [Google Scholar]
- Lee N, Wu JL, Lee CH, Tsai WC. Pseudomonas pseudomallei infection from drowning: the first reported case in Taiwan. J. Clin. Microbiol. 1985;22:352–354. doi: 10.1128/jcm.22.3.352-354.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leelarasamee A. Recent development in melioidosis. Curr. Opin. Infect. Dis. 2004;17:131–136. doi: 10.1097/00001432-200404000-00011. [DOI] [PubMed] [Google Scholar]
- Lever MS, Stagg AJ, Nelson M, et al. Experimental respiratory anthrax infection in the common marmoset (Callithrix jacchus) Int. J. Exp. Pathol. 2008;89:171–179. doi: 10.1111/j.1365-2613.2008.00581.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lever MS, Nelson M, Stagg AJ, Beedham RJ, Simpson AJ. Experimental acute respiratory Burkholderia pseudomallei infection in BALB/c mice. Int. J. Exp. Pathol. 2009;90:16–25. doi: 10.1111/j.1365-2613.2008.00619.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu B, Koo GC, Yap EH, Chua KL, Gan YH. Model of differential susceptibility to mucosal Burkholderia pseudomallei infection. Infect. Immun. 2002;70:504–511. doi: 10.1128/IAI.70.2.504-511.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller WR, Pannell L, Cravitz L, Tanner WA, Rosebury T. Studies on certain biological characteristics of Malleocyes mallei and Malleocyes pseudomallei. II. Virulence and Infectivity for animals. J. Bacteriol. 1948;55:127–135. [PMC free article] [PubMed] [Google Scholar]
- Mutalib AR, Sheikh-Omar AR, Zamri M. Melioidosis in a banded leaf-monkey (Presbytis melalophos) Vet. Rec. 1984;115:438–439. doi: 10.1136/vr.115.17.438. [DOI] [PubMed] [Google Scholar]
- Nelson M, Lever MS, Savage VL, et al. Establishment of lethal inhalational infection with Francisella tularensis (tularemia) in the common marmoset (Callithrix jacchus) Int. J. Exp. Pathol. 2009;90:109–118. doi: 10.1111/j.1365-2613.2008.00631.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nelson M, Lever MS, Dean RE, et al. Characterization of lethal inhalational infection with Francisella tularensis in the common marmoset (Callithrix jacchus) J. Med. Microbiol. 2010;59:1107–1113. doi: 10.1099/jmm.0.020669-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orsi A, Rees D, Andreini I, Venturella S, Cinelli S, Oberto G. Overview of the marmoset as a model in nonclinical development of pharmaceutical products. Regul. Toxicol. Pharmacol. 2011;59:19–27. doi: 10.1016/j.yrtph.2010.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piggott JA, Hochholzer L. Human melioidosis. A histopathologic study of acute and chronic melioidosis. Arch. Pathol. 1970;90:101–111. [PubMed] [Google Scholar]
- Poe RH, Vassallo CL, Domm BM. Melioidosis: the remarkable imitator. Am. Rev. Respir. Dis. 1971;104:427–431. doi: 10.1164/arrd.1971.104.3.427. [DOI] [PubMed] [Google Scholar]
- Retnasabapathy A, Joseph PG. A case of melioidosis in a macaque monkey. Vet. Rec. 1966;79:72–73. doi: 10.1136/vr.79.3.72. [DOI] [PubMed] [Google Scholar]
- Sprague LD, Neubauer H. Melioidosis in animals: a review on epizootiology, diagnosis and clinical presentation. J. Vet. Med. B. Infect. Dis. Vet. Public Health. 2004;51:305–320. doi: 10.1111/j.1439-0450.2004.00797.x. [DOI] [PubMed] [Google Scholar]
- Stanton AT, Fletcher W. Melioidosis and its relation to glanders. J. Hyg. (Lond). 1925;23:33–35. doi: 10.1017/s0022172400034252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strauss JM, Jason S, Lee H, Gan E. Melioidosis with spontaneous remission of osteomyelitis in a macaque (Macaca nemestrina) J. Am. Vet. Med. Assoc. 1969;155:1169–1175. [PubMed] [Google Scholar]
- Tammemagi L, Johnston AY. Melioidosis in an Orang-utan in north Queensland. Aust. Vet. J. 1963;39:241–242. [Google Scholar]
- Titball RW, Russell P, Cuccui J, et al. Burkholderia pseudomallei: animal models of infection. Trans. R. Soc. Trop. Med. Hyg. 2008;102(Suppl. 1):S111–S116. doi: 10.1016/S0035-9203(08)70026-9. [DOI] [PubMed] [Google Scholar]
- White NJ. Melioidosis. Lancet. 2003;361:1715–1722. doi: 10.1016/s0140-6736(03)13374-0. [DOI] [PubMed] [Google Scholar]
- Wiersinga WJ, Meijers JC, Levi M, et al. Activation of coagulation with concurrent impairment of anticoagulant mechanisms correlates with a poor outcome in severe melioidosis. J. Thromb. Haemost. 2008;6:32–39. doi: 10.1111/j.1538-7836.2007.02796.x. [DOI] [PubMed] [Google Scholar]
- Wong KT. The histopathology of human melioidosis. Histopathology. 1995;26:51–55. doi: 10.1111/j.1365-2559.1995.tb00620.x. [DOI] [PubMed] [Google Scholar]