

Abstract
Small conductance Ca2+-activated K+ (SK) and intermediate conductance Ca2+-activated K+ (IK) channels are thought to be involved in detrusor smooth muscle (DSM) excitability and contractility. Using naphtho[1,2-d]thiazol-2-ylamine (SKA-31), a novel and highly specific SK/IK channel activator, we investigated whether pharmacological activation of SK/IK channels reduced guinea pig DSM excitability and contractility. We detected the expression of all known isoforms of SK (SK1-SK3) and IK channels at mRNA and protein levels in DSM by single-cell reverse transcription-polymerase chain reaction and Western blot. Using the perforated patch-clamp technique on freshly isolated DSM cells, we observed that SKA-31 (10 μM) increased SK currents, which were blocked by apamin (1 μM), a selective SK channel inhibitor. In current-clamp mode, SKA-31 (10 μM) hyperpolarized the cell resting membrane potential, which was blocked by apamin (1 μM) but not by 1-[(2-chlorophenyl)diphenylmethyl]-1H-pyrazole (TRAM-34) (1 μM), a selective IK channel inhibitor. SKA-31 (10 nM-10 μM) significantly inhibited the spontaneous phasic contraction amplitude, frequency, duration, and muscle force in DSM isolated strips. The SKA-31 inhibitory effects on DSM contractility were blocked by apamin (1 μM) but not by TRAM-34 (1 μM), which did not per se significantly affect DSM spontaneous contractility. SK channel activation with SKA-31 reduced contractions evoked by electrical field stimulation. SKA-31 effects were reversible upon washout. In conclusion, SK channels, but not IK channels, mediate SKA-31 effects in guinea pig DSM. Pharmacological activation of SK channels reduces DSM excitability and contractility and therefore may provide a novel therapeutic approach for controlling bladder dysfunction.
Introduction
Overactive bladder (OAB), a highly prevalent disorder, is often associated with increased spontaneous contractions of the detrusor smooth muscle (DSM) (Andersson and Arner, 2004; Andersson and Wein, 2004; Fry et al., 2004). Antimuscarinic drugs are currently used for pharmacological OAB treatment, but their efficacy is compromised by adverse side effects such as dry mouth, blurred vision, and constipation (Andersson and Arner, 2004; Andersson and Wein, 2004; Epstein et al., 2006). Therefore, the identification of novel drugs that can decrease the excitability and contractility of DSM may improve OAB treatment options. We propose that pharmacological activation of small conductance Ca2+-activated K+ (SK) and/or the intermediate conductance Ca2+-activated K+ (IK) channels can be an alternative therapeutic option for modulating bladder function.
DSM exhibits spontaneous action potentials that trigger phasic contractions (Brading, 1992; Heppner et al., 1997; Herrera et al., 2000, 2003; Petkov et al., 2001b; Hashitani and Brading, 2003; Brown et al., 2008; Hristov et al., 2008; Chen et al., 2010). Among the diverse group of K+ channels, SK channels have been suggested to help modulate the spontaneous action potentials and related phasic contractions of DSM (Herrera et al., 2000; Imai et al., 2001; Herrera and Nelson, 2002; Hashitani and Brading, 2003). The three subtypes of SK channels, SK1 (KCa2.1), SK2 (KCa2.2), and SK3 (KCa2.3), and IK channels, also known as SK4 (KCa3.1) channels, are voltage-independent and are activated exclusively by intracellular Ca2+ (Xia et al., 1998; Stocker, 2004). Pharmacological blockade of the SK channel with its specific inhibitor, apamin, increases both DSM spontaneous and nerve-evoked contractions and reduces the DSM action potential after-hyperpolarization (Herrera et al., 2000, 2003, 2005; Herrera and Nelson, 2002; Hashitani and Brading, 2003). In a genetic mouse model, SK3 channel overexpression elevates SK currents and decreases the frequency of DSM phasic contractions in vitro, whereas SK3 channel expression suppression substantially increases nonvoiding contractions in vivo (Herrera et al., 2003). Likewise, studies on SK2 knockout mice suggest that the SK2 channel isoform is another essential contributor to DSM function in the mouse bladder (Thorneloe et al., 2008).
Studies suggest that pharmacological activation of SK/IK channels with specific openers may be a novel approach for controlling DSM function. 6,7-Dichloro-1H-indole-2,3-dione-3-oxime (NS309), a potent SK/IK channel activator (Strøbaek et al., 2004), increases bladder capacity and decreases voiding frequency in rats (Pandita et al., 2006), but its extremely short half-life precludes in vivo application. 4,5-Dichloro-1,3-diethyl-1,3-dihydro-benzoimidazol-2-one (NS4591), another SK/IK channel activator, inhibits DSM carbachol-induced contractions in vitro (Nielsen et al., 2011) and bladder overactivity in vivo (Hougaard et al., 2009).
A novel compound, naphthol[1,2-d]thiazol-2-ylamine (SKA-31), has been identified as the most potent activator of SK/IK channels known to date with a half-life of 12 h (Brähler et al., 2009; Sankaranarayanan et al., 2009; Damkjaer et al., 2011; Hasenau et al., 2011; Shakkottai et al., 2011). SKA-31 is 10 times more potent than its template riluzole and activates IK channels with an EC50 value of 250 nM and all three SK channels with EC50 values of 2 to 3 μM. Furthermore, SKA-31 exhibits a better selectivity over other ion channels, particularly Na+ and Ca2+ channels, but is moderately selective (7- to 10-fold) for IK over SK channels (Sankaranarayanan et al., 2009). The effects of SK/IK channel activation with SKA-31 on DSM function have not yet been explored.
The goal of this study was to test the hypothesis that the pharmacological activation of SK/IK channels with SKA-31 can reduce the excitability and contractility in guinea pig DSM. With a multidisciplinary experimental approach, including single-cell reverse transcription-polymerase chain reaction (RT-PCR), Western blot, perforated whole-cell patch-clamp technique, and isometric DSM tension recordings, we observed that SKA-31 decreases the excitability and contractility of guinea pig DSM by acting predominantly on SK channels. Although the IK channel is expressed at the mRNA and protein levels, our electrophysiological and functional studies did not confirm a functional role of the IK channel in guinea pig DSM excitability and contractility.
Materials and Methods
DSM Tissue Collection.
In this study, 106 male Hartley Albino guinea pigs (weight 424.9 ± 7 g) purchased from Charles River (Raleigh, NC) were used. Animals were euthanized with CO2 inhalation followed by thoractomy according to the animal use protocol number 1747 approved by the Institutional Animal Care and Use Committee of the University of South Carolina. DSM strips (2–3 mm wide and 5–7 mm long) devoid of urothelium and mucosa were excised from the bladder and used for single-cell isolation, isometric DSM tension recordings, and molecular biology experiments.
DSM Single-Cell Isolation.
DSM single cells were freshly isolated as described previously (Brown et al., 2008; Hristov et al., 2008, 2011; Chen and Petkov, 2009; Chen et al., 2010). In brief, one to two DSM strips were incubated for 12 to 18 min in 2 ml of dissection solution (DS) supplemented with 1 mg/ml bovine serum albumin (BSA), 1 mg/ml papain (Worthington Biochemicals, Freehold, NJ), and 1 mg/ml dithioerythritol at 37°C. DSM tissues were transferred to a prewarmed 2 ml of DS supplemented with 1 mg/ml BSA, 0.5 mg/ml collagenase type II (Sigma-Aldrich, St. Louis, MO), and 100 μM CaCl2 for 12 to 15 min at 37°C. After enzymatic digestion, DSM tissue was washed two to three times with 1 mg/ml BSA-supplemented DS and then was gently triturated with a fire-blunted Pasteur pipette to disperse single DSM cells. Several drops of the DSM isolated cell suspension were placed onto a recording chamber and allowed to adhere to the glass bottom for approximately 20 min. Freshly isolated elongated, distinct, bright, and shiny DSM cells [when viewed illuminated under an Axiovert 40CFL microscope (Carl Zeiss Inc., Thornwood, NY)] were selected for patch-clamp recordings and single-cell RNA isolation.
RNA Isolation and Reverse Transcriptase-Polymerase Chain Reaction.
Fresh DSM isolated strips and enzymatically isolated single DSM cells were used to extract DSM total RNA. We collected 50 to 100 individual DSM cells in RNAlater by using an Axiovert 40CFL microscope (Carl Zeiss Inc.) with Nomarski interference contrast and a glass micropipette by using an MP-285/ROE micromanipulator (Sutter Instrument Company, Novato, CA). Total RNA was isolated from single DSM cells and whole DSM tissues by using a RNeasy Mini Kit (QIAGEN GmbH, Hilden, Germany). The extracted RNA was reverse-transcribed into cDNA by using Moloney murine leukemia virus reverse transcriptase (Promega, Madison, WI) and oligo d(T) primers. The cDNA production was amplified by using GoTaq Green Master Mix (Promega), and gene-specific primers were optimized by using a mastercycler gradient thermocycler (Eppendorf AG, Hamburg, Germany). All gene-specific primers were designed [Primer 3 (http://frodo.wi.mit.edu/primer3/) or Gene Runner (http://www.generunner.net/) software] based on the completed guinea pig mRNA sequences in Genbank and are summarized in Table 1. Guinea pig brain tissue was used as a positive control. Negative control experiments were performed in the absence of the reverse transcriptase (−RT). PCR products were purified by using the GenElute PCR Clean-Up Kit (Sigma-Aldrich) and sequenced at the University of South Carolina Environmental Genomics facility for sequence confirmation.
TABLE 1.
RT-PCR primers for the SK (1–3) and IK/SK4 channels
| Channel | Sense | Antisense | Size |
|---|---|---|---|
| bp | |||
| SK1 | ACCGTGCTGTTGGTCTTC | AACTTACGCTGGTGTTTCC | 450 |
| SK2 | TCTGGCAAAGACCCAGAACA | ATGAAGTTGGTGGCGCTGTA | 284 |
| SK3 | TGCCAAAGTCAGGAAACA | GATAGGGGCTGTCAGAGGG | 296 |
| SK4/IK | TCCAGCTGTTCATGACCGACAA | ATGTACAGCTTGGCCACGAA | 349 |
Western Blot.
Guinea pig DSM isolated strips (30–50 mg) were homogenized with standard radioimmunoprecipitation assay buffer containing protease inhibitors (Thermo Fisher Scientific, Waltham, MA). The mixture was centrifuged at 1000 rpm for 30 min. Supernatant was collected, and the pellet was resuspended in radioimmunoprecipitation assay buffer and centrifuged again. Supernatant was collected, and the protein concentration was determined with a BCA Protein Assay Kit (Thermo Fisher Scientific). Protein was mixed with 5 × Laemmli buffer (1:4) and denaturized for 5 min at 95°C. Subsequently, equal amounts of proteins (approximately 50 μg) were loaded into adjacent lanes, subjected to 4 to 20% precase SDS-polyacrylamide gel electrophoresis for 2.5 h at 20 mA, and transferred to a polyvinylidene fluoride membrane at 40 mA for 2 h using semidry blot. The membrane was blocked with 5% dry milk/Tris-buffered saline-Tween 20 buffer for 1 h at room temperature. The membrane was incubated with primary antibody anti-KCa2.1 (SK1), anti-KCa2.2 (SK2), anti-KCa2.3 (SK3), or anti-KCa3.1 (SK4) (1:200) (Alomone Labs, Jerusalem, Israel) overnight at 4°C. The membrane was washed with Tris-buffered saline-Tween 20 three to four times and incubated with goat anti-rabbit IgG conjugated with horseradish peroxidase (1:2500) in blocking buffer for 1 h at room temperature. Bound antibodies were detected by an echochemiluminescence substrate kit (Amersham, Piscataway, NJ) according to the manufacturer's instructions. Staining specificity was verified by preincubation of the antibodies with the respective competing peptide.
Electrophysiological Recordings.
The amphotericin-B perforated whole-cell configuration of the patch-clamp technique was used to conduct the electrophysiological recordings from freshly isolated single DSM cells (Petkov and Nelson, 2005; Brown et al., 2008; Hristov et al., 2008, 2011). Whole-cell currents or membrane potentials were recorded in voltage-clamp or current-clamp mode, respectively, using an Axopatch 200B amplifier, Digidata 1440A, and pCLAMP version 10.2 software (Molecular Devices, Sunnyvale, CA). Ca2+-activated K+ currents were elicited by holding the cells at −70 mV and then depolarized to +10 mV for 100 ms before stepping down to −70 mV again. This depolarization step evokes inward L-type, voltage-dependent Ca2+ channel (VDCC) currents that normally occur during an action potential, which in turn activate outward Ca2+-activated K+ currents in DSM cells (Herrera and Nelson, 2002; Petkov and Nelson, 2005). The main component of the Ca2+-activated K+ currents in DSM cells is carried by the large conductance Ca2+-activated K+ (BK) channel (Petkov et al., 2001a; Petkov and Nelson, 2005; Brown et al., 2008; Hristov et al., 2008, 2011). To study selectively the whole-cell outward SK/IK currents, the influence of BK channels was suppressed with the continuous presence of 100 nM paxilline, a specific BK channel inhibitor. Resting membrane potential was recorded in current-clamp mode (I = 0) of the patch-clamp technique in the presence of 100 nM paxilline. All patch-clamp experiments were conducted at room temperature (22–23°C).
Isometric DSM Tension Recordings.
One end of the DSM isolated strip was secured to the bottom of a temperature-controlled (37°C), water-jacketed bath (10 ml) containing physiological saline solution (PSS) aerated with 95% O2/5% CO2, pH 7.4. The other end of the DSM strip was secured to an isometric force transducer. The DSM strips were initially tensioned (1 g) and washed with fresh PSS every 15 min during an equilibration period of 45 to 60 min. To minimize the effects caused by neurotransmitter release, the experiments with myogenic contractions were performed in the presence of 1 μM tetrodotoxin (TTX), a specific blocker of neuronal Na+ channels. SKA-31 was administered in increasing concentrations every 10 min directly into the tissue baths with spontaneously contracting DSM strips.
In another experimental series, the effect of SKA-31 on nerve-evoked contractions was evaluated. Nerve-evoked contractions were induced by EFS by using a pair of platinum electrodes mounted in the tissue bath in parallel to the DSM strip. The EFS pulses were generated by using a PHM-152I stimulator (MED Associates, St. Albans, VT), and the EFS pulse parameters were as follows: 0.75-ms pulse width, 20-V pulse amplitude, and 3-s stimulus duration. Polarity was reversed for alternating pulses. For EFS studies, after the equilibration period, DSM strips were subjected to either continuous repetitive stimulation with a frequency of 20 Hz at 1-min intervals or increasing frequencies from 0.5 to 50 Hz at 3-min intervals. The contractions were recorded by using a Myomed myograph system (MED Associates).
Solutions and Drugs.
Freshly prepared DS had the following composition: 80 mM monosodium glutamate, 55 mM NaCl, 6 mM KCl, 10 mM glucose, 10 mM HEPES, and 2 mM MgCl2 with the pH adjusted to 7.3 with NaOH. The external bath solution for the patch-clamp experiments contained 134 mM NaCl, 6 mM KCl, 1 mM MgCl2, 2 mM CaCl2, 10 mM glucose, and 10 mM HEPES, with the pH adjusted to 7.4 with NaOH. The pipette solution was composed of 110 mM potassium aspartate, 30 mM KCl, 10 mM NaCl, 1 mM MgCl2, 10 mM HEPES, and 0.05 mM EGTA with the pH adjusted to 7.2 with NaOH. Pipette solution was filtered by using a 0.2-μM filter and stored at −20°C. Freshly dissolved amphotericin-B (200 μg/ml) in dimethyl sulfoxide (DMSO) was added to the pipette solution just before the experiment, and the solution was replaced every 1 to 2 h. For DSM contraction studies, freshly prepared PSS had 119 mM NaCl, 4.7 mM KCl, 24 mM NaHCO3, 1.2 mM KH2PO4, 2.5 mM CaCl2, 1.2 mM MgSO4, and 11 mM glucose and was aerated with 95% O2/5% CO2 to maintain the pH. SKA-31 was provided by Dr. Heike Wulff (University of California, Davis, CA). 1-[(2-Chlorophenyl)diphenylmethyl]-1H-pyrazole (TRAM-34), apamin, TTX, and paxilline were purchased from Sigma-Aldrich. Apamin and TTX were dissolved in double-distilled water, whereas SKA-31, TRAM-34, and paxilline were dissolved in DMSO. The maximum concentration of DMSO in the bath solution was not higher than 0.1%.
Data Analysis and Statistics.
MiniAnalysis software (Synaptosoft, Decatur, GA) was used to analyze the DSM contraction parameters. The DSM contractile activity was quantified by measuring the average maximal phasic contraction amplitude, muscle force integral (the area under the curve of the phasic contractions), the duration (defined as half-width of contraction amplitude), the frequency (contractions per minute), and the tone (phasic contraction baseline curve). To compare the phasic contraction parameters, data were normalized to the spontaneous contractions (taken to be 100%) and expressed as percentages. For the EFS-induced contractions, the maximal contraction amplitude at EFS frequency of 50 Hz under control conditions was taken to be 100%, and the data were normalized. To evaluate the effect of SKA-31 on the spontaneous phasic contractions, 10-min periods before SKA-31 application were analyzed for the control, and subsequent 10-min periods after application of each concentration of SKA-31 were analyzed. To evaluate whole-cell outward K+ currents, mean value of the last 10-ms pulse of 100-ms depolarization step of average files (6–10 files) recorded every 1 min for 6 to 10 min was calculated by using Clampfit (version 10.2; Molecular Devices). Statistical analysis was performed with Prism 4.03 software (GraphPad Software, Inc., San Diego, CA). The data are expressed as mean ± S.E.M. for the n (the number of DSM strips or cells) isolated from N (number of guinea pigs). Data were evaluated for statistical significance by using either two-tailed paired Student's t test or one-way analysis of variance followed by Newman-Keuls post test. A P value less than 0.05 was considered statistically significant. Corel Draw Graphic Suite X3 software (Corel Co., Mountain View, CA) was used to illustrate the data.
Results
Guinea Pig DSM Expresses mRNA for SK1, SK2, SK3, and IK Channels.
We detected expression of mRNA messages for SK1, SK2, SK3, and IK (SK4) channels in whole DSM tissue (Fig. 1). Detection of mRNA messages from non-DSM cells such as fibroblasts, neuronal cells, and vascular smooth muscle was avoided by performing single-DSM-cell RT-PCR. Single-cell RT-PCR results showed that mRNA messages for all isoforms of SK (SK1-SK3) and IK channels came directly from the DSM cells (Fig. 1).
Fig. 1.

RT-PCR detection of mRNA messages for SK1 (450 bp), SK2 (284 bp), SK3 (296 bp), and IK (SK4) (349 bp) channels in DSM whole tissue and single DSM cells. Illustrated gel images are selected representations of at least five independent RT-PCR experiments based on RNA extracted from five animals. Guinea pig brain was used as a positive control (+). Negative control experiments carried out with omission of RT-enzyme (−RT) in reaction mixtures showed no products.
Guinea Pig DSM Expresses Proteins for SK1, SK2, SK3, and IK Channels.
The protein expression of SK1, SK2, SK3, and IK (SK4) channels in whole DSM tissues was detected by using channel-specific antibodies (Fig. 2). The expected molecular mass values of SK1, SK2, SK3, and IK (SK4) channels were ∼60, ∼60, ∼95, and ∼56 kDa, respectively, consistent with the experimentally determined molecular mass. Preabsorption of the primary antibody with its antigenic competing peptide indicated the specificity of the antibody. These results are consistent with channel mRNA expression identified by RT-PCR on DSM tissue.
Fig. 2.
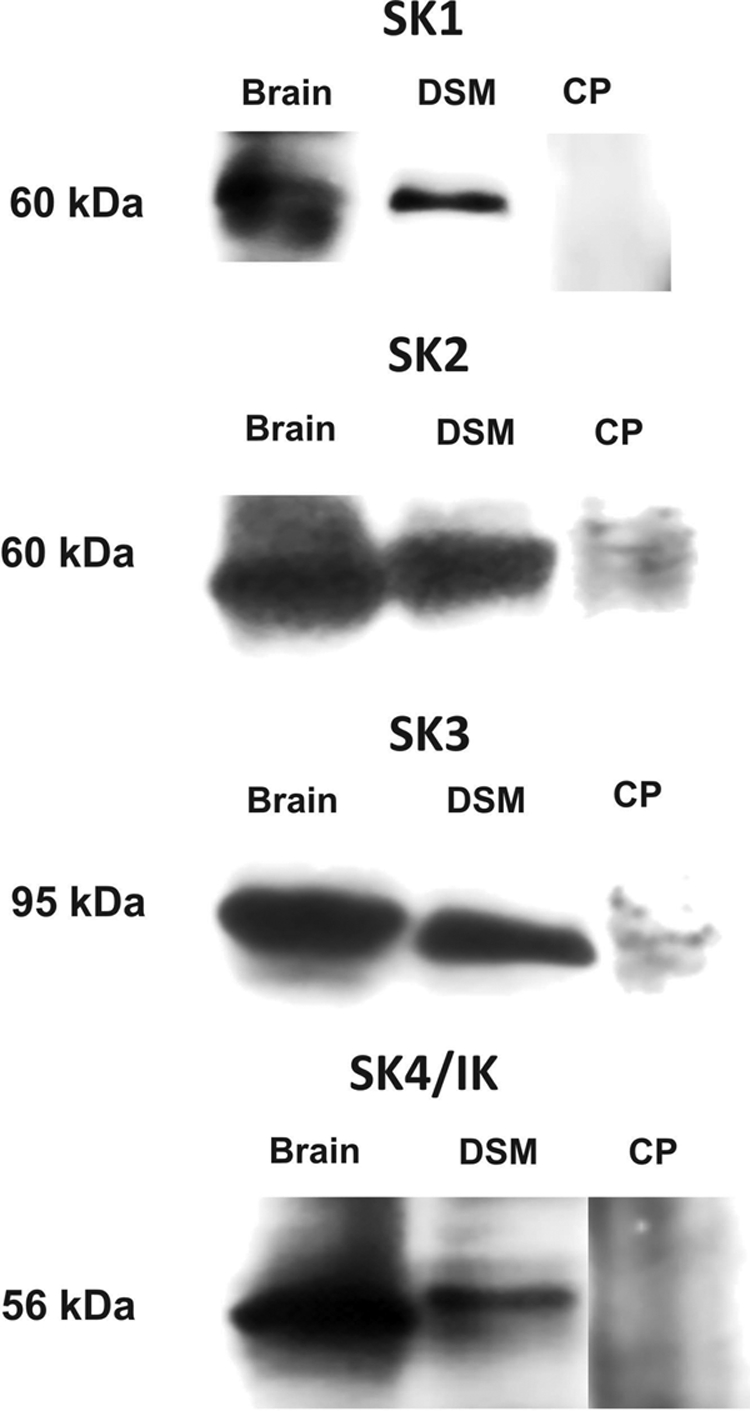
Western blot detection of SK1 (∼60 kDa), SK2 (∼60 kDa), SK3 (∼95 kDa), and IK (SK4) (∼56 kDa) channel protein expression in DSM tissues. Illustrated images are representation of at least five independent Western blot experiments based on protein extracted from five animals. Guinea pig brain was used as a positive control. The lack of immunoreactive bands in the presence of competing peptide (CP) confirmed the specificity of the primary antibodies.
SKA-31 Hyperpolarizes the Resting Membrane Potential in Freshly Isolated Guinea Pig DSM Cells.
The average whole-cell capacitance of all studied guinea pig DSM cells was 28.8 ± 0.7 pF (n = 267; N = 106). In current-clamp mode, we evaluated the effects of SKA-31 (10 μM) on the resting membrane potential of freshly isolated guinea pig DSM cells. All experiments were performed in the presence of paxilline to eliminate the influence of BK channel on the resting membrane potential. Membrane potential was recorded for at least 10 min in the presence of paxilline (100 nM) before the addition of any compounds. SKA-31 (10 μM) significantly hyperpolarized the resting membrane potential from −24.7 ± 1.2 to −27.2 ± 1.3 mV (n = 11; N = 9; P < 0.05). SKA-31 (10 μM)-induced DSM cell membrane hyperpolarization was reversed back to the control values after application of 1 μM apamin (n = 5; N = 4; P < 0.05; Fig. 3A). We also evaluated the effect of apamin (1 μM) alone and in the presence of SKA-31 (10 μM) on DSM cell resting membrane potential. Pretreatment of DSM cells with apamin (1 μM) did not affect the resting membrane potential but it significantly suppressed the hyperpolarizing effect of SKA-31 (10 μM) (n = 8; N = 5; Fig. 3B). In another series of experiments, we explored whether 1 μM TRAM-34, a selective IK channel inhibitor, can reverse SKA-31-induced hyperpolarization. Unlike apamin, TRAM-34 (1 μM) failed to reverse the SKA-31 (10 μM)-induced membrane hyperpolarization (n = 6; N = 5; Fig. 3C). Likewise, we also examined the effect of TRAM-34 (1 μM) alone and in the presence of SKA-31 (10 μM) to evaluate whether the pharmacological inhibition of IK channels with TRAM-34 can affect the DSM cell resting membrane potential. Pretreatment of DSM cells with TRAM-34 (1 μM) did not alter the DSM cell resting membrane potential (n = 11; N = 6). Further TRAM-34 did not suppress the hyperpolarizing effect of SKA-31 (10 μM) on the resting membrane potential (n = 11; N = 6; Fig. 3D). These results suggest that pharmacological activation of SK channels, but not IK channels, can hyperpolarize the guinea pig DSM cell resting membrane potential.
Fig. 3.

SKA-31 hyperpolarizes the resting membrane potential in freshly isolated guinea pig DSM cells. A, representative recording illustrating that SKA-31 (10 μM) hyperpolarized the resting membrane potential (RMP) in a guinea pig DSM cell. Apamin (1 μM) reversed the hyperpolarizing effect of SKA-31 (10 μM) on the resting membrane potential. Bar graph shows the effect of SKA-31 (10 μM) on the resting membrane potential in the absence and after application of 1 μM apamin (n = 5; N = 4; *, P < 0.05). B, representative recording illustrating that apamin (1 μM) pretreatment for 10 min prevented the SKA-31 (10 μM)-induced hyperpolarizing effect on the resting membrane potential in a guinea pig DSM cell. Bar graph shows the effect of apamin (1 μM) and the lack of SKA-31 (10 μM) effect in the presence of apamin on the resting membrane potential (n = 8; N = 5; P > 0.05). C, representative recording illustrating that TRAM-34 (1 μM) did not reverse the hyperpolarizing effect of SKA-31 (10 μM) on the resting membrane potential. Bar graph shows the effect of SKA-31 (10 μM) on the resting membrane potential in the absence and after application of TRAM-34 (1 μM) (n = 6; N = 5; *, P < 0.05). D, representative recording showing that pretreatment of DSM cells with TRAM-34 (1 μM) for 10 min did not change the SKA-31 (10 μM)-induced hyperpolarizing effect on the resting membrane potential in guinea pig DSM cells. Bar graph shows the lack of TRAM-34 (1 μM) effect alone and the hyperpolarizing effect of SKA-31 (10 μM) in the presence of TRAM-34 (1 μM) (n = 11; N = 6; *, P < 0.05). All experiments were performed in the presence of paxilline (100 nM). NS, nonsignificant.
SKA-31 Activates Whole-Cell Outward SK Currents in Freshly Isolated Guinea Pig DSM Cells.
A voltage step depolarization protocol of perforated whole-cell patch-clamp configuration was applied to evoke VDCC currents, and thus activate SK/IK currents from guinea pig DSM cells. Voltage step depolarization from a holding potential of −70 to +10 mV for 100 ms evoked small VDCC currents followed by large outward K+ currents (Petkov and Nelson, 2005). We used this protocol because the depolarization of +10 mV facilitates the Ca2+ entry through VDCC to activate outward Ca2+-activated K+ currents, such as SK, IK, and BK currents. Paxilline (100 nM) was present in the bath solution throughout the recordings to block the BK current and dissect the BK from the SK/IK currents. Exposure of DSM cells to SKA-31 (10 μM) in bath solution significantly increased the whole-cell Ca2+-activated outward K+ currents from the control mean value of 2.6 ± 0.4 to 3.0 ± 0.4 pA/pF (n = 10; N = 9; P < 0.005; Fig. 4A).
Fig. 4.

SKA-31 increases the whole-cell SK current amplitude in freshly isolated guinea pig DSM cells. A, representative recording illustrating that SKA-31 (10 μM) increased the whole-cell SK currents evoked by activation of VDCCs caused by membrane depolarization from −70 to +10 mV for 100 ms. Bar graph shows the summary data for the stimulatory effect of SKA-31 on whole-cell Ca2+-activated outward K+ currents (n = 10; N = 9; **, P < 0.005). B, representative recording illustrating that SKA-31 (10 μM)-induced whole-cell Ca2+-activated outward K+ currents were suppressed by 1 μM apamin. Bar graph shows the summary data for the stimulatory effect of SKA-31 on whole-cell outward K+ currents that were inhibited by 1 μM apamin (n = 10; N = 9; **, P < 0.005).
To examine whether whole-cell Ca2+-activated outward K+ currents were increased because of SK channel activation with SKA-31, we applied 1 μM apamin to the bath solution in the presence of SKA-31. As shown in Fig. 4B, apamin significantly suppressed the whole-cell Ca2+-activated outward K+ currents induced by 10 μM SKA-31 (3.0 ± 0.4 to 2.4 ± 0.4 pA/pF; n = 10; N = 9; P < 0.005). This suggests that the increase in the whole-cell outward K+ currents is carried out by the activation of the SK channels. In subsequent experiments, the effect of SKA-31 on the whole-cell Ca2+-activated K+ currents was recorded in cells preincubated with 1 μM apamin. In the presence of 1 μM apamin, SKA-31 (10 μM) failed to increase the whole-cell Ca2+-activated K+ currents (2.6 ± 0.4 to 2.5 ± 0.4 pA/pF; n = 5; N = 3, P > 0.05; data not illustrated). These results further confirm that SKA-31 targets exclusively the SK channels.
Activation of the SK Channels with SKA-31 Reduces Spontaneous Phasic Contractions in Guinea Pig DSM Isolated Strips.
DSM isolated strips exhibiting spontaneous contractions were exposed to increasing concentrations of SKA-31 (10 nM-10 μM). SKA-31 caused a concentration-dependent inhibition of the spontaneous phasic contraction amplitude, frequency, duration, and muscle force integral without significant effects on muscle tone (n = 10; N = 8; Fig. 5A). The role of SK channels was examined by testing the effects of apamin (1 μM), a selective blocker of SK channels. SK channel inhibition with apamin dramatically increased the contraction amplitude by 52.2 ± 6.1%, muscle integrated force by 65.4 ± 10.3% and contraction duration by 43.6 ± 9.6%. These data are consistent with previous reports (Herrera et al., 2000; Herrera and Nelson, 2002).
Fig. 5.

SKA-31 decreases the spontaneous phasic contractions in a concentration-dependent manner in guinea pig DSM isolated strips. A, representative recordings showing the concentration-response inhibitory effects for SKA-31 (10 nM-10 μM) on the DSM spontaneous phasic contractions. B, representative recordings depicting the concentration-response inhibitory effects for SKA-31 (10 nM-10 μM) on the DSM spontaneous phasic contractions in the presence of apamin (1 μM). C, representative recordings showing the concentration-response effects for SKA-31 (10 nM-10 μM) on the DSM spontaneous phasic contraction in the presence of TRAM-34 (1 μM). SKA-31 was added in increasing concentrations every 10 min to the bath solution. All recordings were performed in the presence of TTX (1 μM). D, concentration-response curves for the inhibitory effects of SKA-31 alone (■) and after preincubation with 1 μM apamin (▾) or 1 μM TRAM-34 (▴). X-axis in D is log transformation of SKA-31 molar concentration. SKA-31 decreased the spontaneous phasic contraction amplitude, muscle force integral, phasic contraction frequency, and duration in a concentration-dependent manner (n = 10; N = 8). Preincubation of DSM isolated strips with 1 μM apamin significantly decreased the inhibitory effects of SKA-31 and caused a significant inhibition of the concentration-response curves (n = 9; N = 8; **, P < 0.01; *, P < 0.05). TRAM-34 (1 μM) did not block the inhibitory effect of SKA-31 (n = 7; N = 7; P > 0.05). Data are means ± S.E.M.
Pretreatment of DSM strips with 1 μM apamin blocked the SKA-31-induced inhibition of spontaneous phasic contractions (Fig. 5B), whereas pretreatment with 1 μM TRAM-34 did not prevent SKA-31-induced inhibition of spontaneous phasic contractions (Fig. 5C). Figure 5D shows the concentration-response curves for the effect of SKA-31 (10 nM-10 μM) in the absence or presence of either apamin (1 μM) or TRAM-34 (1 μM). In the presence of SKA-31 (10 μM) the spontaneous phasic contraction amplitude was reduced by 73.8 ± 4.0%, muscle force integral by 67.4 ± 1.9%, phasic contraction duration by 65.4 ± 1.3%, and phasic contraction frequency by 51.8 ± 3.0%. The average EC50 values for SKA-31 calculated for contraction parameters were: contraction amplitude, 0.5 μM; muscle force integral, 1 μM; contraction duration, 3.4 μM; and contraction frequency, 40 nM (n = 10; N = 8). Application of apamin (1 μM) inhibited SKA-31 effects on all contraction parameters (n = 9; N = 8; P < 0.05). Pretreatment of the DSM strips with 1 μM apamin (for 20–30 min) led to a significant reduction in the concentration-responses to SKA-31 (n = 9; N = 8). Pretreatment of the DSM strips with 1 μM TRAM-34 did not change the inhibitory effect of SKA-31 on any of the phasic contraction parameters (n = 7; N = 7; P > 0.05).
To explore the potential functional role of IK channels in DSM contractility, a concentration-response curve for TRAM-34 (1–30 μM) was performed on spontaneous phasic contractions in DSM strips. However, no significant change in any of the contraction parameters was observed even at 30 μM TRAM-34 (n = 5; N = 5; P > 0.05, data not illustrated), suggesting that IK channels do not play a role in regulating guinea pig DSM spontaneous phasic contractions.
Pharmacological SK Channel Activation with SKA-31 Inhibits EFS-Induced Contractions of Guinea Pig DSM Isolated Strips.
Next, we investigated the effect of SKA-31 on guinea pig DSM nerve-evoked contractions. In the absence of TTX, contractions were evoked by repetitive EFS (20 Hz/min; Fig. 6). SKA-31 reduced the amplitude of EFS-induced contractions in a concentration-dependent fashion (EC50 = 0.4 μM). The highest concentration of SKA-31 (10 μM) reduced the EFS-induced contraction amplitude by 84.3 ± 3.1% (n = 8; N = 7; P < 0.05; Fig. 6B).
Fig. 6.

SK channel pharmacological activation with SKA-31 significantly reduces the EFS-induced contraction amplitude in a concentration-dependent manner in guinea pig DSM isolated strips. A, representative recording showing the inhibitory effects of SKA-31 (10 nM-10 μM) on EFS-induced contractions (20 Hz/min). B, concentration-response curve for the inhibitory effect of SKA-31 on the amplitude of EFS-induced contractions (n = 8; N = 7). X-axis represents log transformation of SKA-31 molar concentration. Data are means ± S.E.M.
Next, EFS frequency-response curves were generated by measuring the amplitude of the EFS-induced DSM contractions at increasing stimulation frequencies (0.5, 2, 3, 5, 7.5, 10, 12.5, 15, 20, 30, 40, and 50 Hz; Fig. 7A). After generating a control frequency-response curve, a second curve was generated by using the same EFS parameters after preincubation with SKA-31 (10 μM) for 30 min. A final curve was generated after five to seven washouts of SKA-31 with fresh PSS at 2-min intervals. EFS-induced contractions (0.5–50 Hz) of guinea pig DSM isolated strips were significantly decreased in the presence of 10 μM SKA-31 at all stimulation frequencies greater than 3 Hz (Fig. 7). The amplitude of EFS-induced contraction at 50 Hz was reduced by 65.6 ± 6.5% in the presence of 10 μM SKA-31 (n = 7; N = 7; P < 0.005; Fig. 7). Washout of SKA-31 with fresh PSS significantly recovered the contraction amplitude at stimulation frequencies greater than 3 Hz (n = 7; N = 7; P < 0.05; Fig. 7). These findings suggest that pharmacological activation of the SK channel with SKA-31 can effectively inhibit nerve-evoked contractions.
Fig. 7.

SK channel pharmacological activation with SKA-31 significantly reduces the nerve-evoked contractions in guinea pig DSM isolated strips. A, representative recordings of EFS-induced contraction (stimulation frequency 0.5–50 Hz at 3-min intervals) in the absence of SKA-31 (control), 35 min after application of SKA-31, and after washout of SKA-31. B, frequency-response curves illustrating a decrease in the amplitude of EFS-induced contractions after application of 10 μM SKA-31 (n = 7; N = 7; ***, P < 0.005; **, P < 0.01; *, P < 0.05). Washout of SKA-31 caused a partial recovery on the amplitude of EFS-induced contractions (n = 6; N = 6; ###, P < 0.005; ##, P < 0.01; #, P < 0.05). The maximal contraction amplitude at EFS frequency of 50 Hz under control conditions was taken to be 100%. Data are means ± S.E.M.
Discussion
Here, using a combined experimental approach involving RT-PCR, Western blot, perforated whole-cell patch-clamp, and isometric DSM tension recordings, we revealed that pharmacological activation of SK channels with SKA-31 can reduce the excitability and contractility of guinea pig DSM. For the first time, we report mRNA message expression of all known isoforms of SK and IK channels in native guinea pig DSM isolated cells by using single-cell RT-PCR (Fig. 1). We also confirmed the expression of all SK (SK1-SK3) and IK channel proteins at the DSM tissue level by Western blot (Fig. 2). In freshly isolated guinea pig DSM cells, SKA-31 increased the apamin-sensitive whole-cell outward SK currents in voltage-clamp mode and hyperpolarized the resting membrane potential in current-clamp mode (Figs. 3 and 4). Furthermore, this novel compound remarkably inhibited the myogenic and nerve-evoked contractions of guinea pig DSM isolated strips (Figs. 5–7).
We measured a pharmacologically activated whole-cell SK current of ∼0.6 pA/pF at +10 mV (Fig. 4). This observation is consistent with studies that reported low SK channel density in guinea pig DSM cells and calculated that these cells have ∼0.01 SK channels/μm2 or ∼50 SK channels/cell (Herrera and Nelson, 2002). The low SK channel density along with their pronounced Ca2+ sensitivity uniquely suits these channels as sensors for changes in global Ca2+ induced by activation of VDCC during the DSM action potential. Our results also demonstrate that activation of SK channels with SKA-31 caused relatively small, but statistically significant, DSM cell membrane hyperpolarization (Fig. 3), which was sufficient to inhibit DSM spontaneous phasic contractions (Fig. 5). Using KATP channel openers, we previously established that modest cell membrane hyperpolarization can substantially inhibit guinea pig DSM action potentials and related spontaneous phasic contractions (Petkov et al., 2001b). Likewise, the present study clearly indicates that small increases in the whole-cell K+ conductance caused by SK channel activation with SKA-31 substantially inhibit guinea pig DSM contractility. Our data also indicate that the dramatic inhibitory effects of SKA-31 on DSM contractility (Fig. 5) are not caused by SKA-31 inhibition of VDCC (Fig. 4), findings that are consistent with a previous report indicating that SKA-31 does not affect VDCC (Sankaranarayanan et al., 2009).
In guinea pig and mouse DSM, SK channel inhibition increases the spontaneous phasic contraction amplitude and decreases the contraction frequency (Herrera et al., 2000, 2003; Imai et al., 2001; Thorneloe et al., 2008). A recent report indicated that NS4591, an activator of SK/IK channels, inhibits EFS- and carbachol-induced contractions in rat, pig, and human DSM (Nielsen et al., 2011). However, to our knowledge, no patch-clamp study has been performed to elucidate the role of DSM SK and IK channels by using SK/IK channel pharmacological activators. Our work shows, for the first time, that activation of SK channels with SKA-31 leads to a significant inhibition of DSM spontaneous phasic contractions. This novel SK channel opener potently decreased the phasic contraction amplitude, muscle force integral, contraction duration, and frequency of guinea pig DSM strips with EC50 values in the submicromolar range (Fig. 5). Similar EC50 values of SKA-31 have been reported for activation of cloned and native SK (SK1-SK3) channels (Sankaranarayanan et al., 2009). Our data showed that apamin did not change the resting membrane potential in DSM isolated cells (Fig. 3B). Because SK channels control only the action potential after-hyperpolarization phase (Hashitani and Brading, 2003), SK channel pharmacological activation with SKA-31 would prolong the action potential after-hyperpolarization phase, delaying the next action potential generation, thus reducing the phasic contractions.
The physiological role of IK channels in DSM function has not been explored, although they may function similarly to the BK and SK channels to facilitate bladder relaxation. Expression of mRNA message for the IK channel in mouse DSM has been reported previously (Ohya et al., 2000). However, the functional role of IK channels in DSM excitability and contractility has been controversial. Under normal physiological conditions, we observed no significant effects of TRAM-34 (1–30 μM) on the spontaneous DSM contractions. TRAM-34 (1 μM) did not change the SKA-31 inhibitory effects on DSM phasic contractions either (Fig. 5). The lack of IK channel functional effect on DSM contractility is consistent with our electrophysiological data showing that TRAM-34 had no effect on the DSM cell resting membrane potential and could not block SKA-31-induced cell membrane hyperpolarization (Fig. 3). IK channels are major players in endothelial cell function, but their role in native smooth muscle is not yet established. Possibly, in guinea pig DSM, IK channels are expressed in the cytoplasm but normally are not trafficking to the cell membrane. A recent study shows that the IK channel has two splice variants, one expressed in the plasma membrane and the other one in the cytoplasm, and further suggests that alternative splicing events are responsible for the fine-tuning of IK channel activity under physiological and pathophysiological conditions (Ohya et al., 2011). Indeed, a role for IK channels has been demonstrated in some vascular smooth muscle cells under pathological conditions (Tharp and Bowles, 2009). Our findings do not rule out a potential functional role of the IK channels in DSM contractility under certain pathophysiological conditions.
Nerve-evoked contractions reflect the activation of DSM purinergic and muscarinic receptors (Hashitani and Suzuki, 1995; Hashitani et al., 2000; Andersson and Arner, 2004). It has been suggested that the inhibition of DSM excitability has a significant influence on EFS-induced contractions by decreasing the Ca2+ entry through the VDCC (Herrera et al., 2005). Our results showed that SKA-31 reduced the amplitude of nerve-evoked contractions in a concentration-dependent manner with an EC50 value of 0.4 μM (Fig. 6). SKA-31 also reduces the amplitude of the nerve-evoked contractions under a whole range of EFS stimulation frequencies (Fig. 7). This inhibition of EFS-induced contraction amplitude is favored by the activation of SK channels, which leads to a closure of VDCC caused by DSM cell hyperpolarization (Fig. 3). Previous studies have shown that the pharmacological block of the VDCC eliminates SK currents and decreases action potentials and spontaneous and nerve-evoked contractions in DSM (Herrera et al., 2000, 2005; Herrera and Nelson, 2002). By sensing elevations in global cell Ca2+ concentration induced by the activation of VDCC during the DSM action potential, SK channels limit the cell excitability and thus DSM contractility. As shown here, the pharmacological activation of the DSM SK channels further facilitates this physiological process. Our findings on the pharmacological activation of the SK channel in DSM are in line with previous studies using conditional and conventional SK2 and SK3 channel knockout mouse models (Herrera et al., 2003; Thorneloe et al., 2008). Selective SK3 channel overexpression reduces DSM spontaneous phasic contraction frequency and increases bladder capacity, whereas selective suppression of SK3 channel expression results in DSM overactivity (Herrera et al., 2003). Another study with SK2 knockout mice showed a loss of apamin sensitivity on spontaneous phasic and nerve-evoked contractions, demonstrating a vital role of SK2 channels in DSM function (Thorneloe et al., 2008).
In conclusion, three key points can be drawn from our study: 1) SK1–3 and IK channels are expressed in guinea pig DSM cells at mRNA and protein levels; 2) SK channel activation with SKA-31 moderately decreases cell excitability but dramatically decreases the contractility of guinea pig DSM; and 3) IK channels do not play a role in guinea pig DSM excitability and contractility although they are expressed in these cells. Our findings suggest that the pharmacological activation of SK channels with SKA-31 may have a therapeutic potential for the treatment of OAB. SK channel openers could, in the future, replace the commonly prescribed therapy for OAB antimuscarinic agents. However, future clinical trials are needed to evaluate the therapeutic potential of the SK channel openers and identify any potential adverse effects. Most importantly, this study is a foundation for developing novel strategies for OAB treatment.
Acknowledgments
We thank Dr. Heike Wulff for kindly providing SKA-31 and Dr. Jennifer Schnellmann, Dr. John Malysz, Dr. Wenkuan Xin, Qiuping Cheng, Serge Afeli, and Amy Smith for critically evaluating the manuscript.
This study was supported by the National Institutes of Health National Institute of Diabetes and Digestive and Kidney Diseases [Grants DK084284, DK083687] (to G.V.P.).
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
- OAB
- overactive bladder
- DSM
- detrusor smooth muscle
- DS
- dissection solution
- PSS
- physiological salt solution
- VDCC
- L-type voltage-gated Ca2+ channel
- SK
- small conductance Ca2+-activated K+
- IK
- intermediate conductance Ca2+-activated K+
- BK
- large conductance voltage- and Ca2+-activated K+
- RT-PCR
- reverse transcription-polymerase chain reaction
- TTX
- tetrodotoxin
- BSA
- bovine serum albumin
- DMSO
- dimethyl sulfoxide
- EFS
- electrical field stimulation
- RMP
- resting membrane potential
- NS
- nonsignificant
- SKA-31
- naphtho[1,2-d]thiazol-2-ylamine
- TRAM-34
- 1-[(2-chlorophenyl)diphenylmethyl]-1H-pyrazole
- bp
- base pairs
- NS309
- 6,7-dichloro-1H-indole-2,3-dione-3-oxime
- NS4591
- 4,5-dichloro-1,3-diethyl-1,3-dihydro-benzoimidazol-2-one
- n
- number of DSM strips or cells
- N
- number of guinea pigs
- CP
- competing peptide.
Authorship Contributions
Participated in research design: Petkov.
Conducted experiments: Parajuli, Soder, and Hristov.
Performed data analysis: Parajuli, Soder, and Hristov.
Wrote or contributed to the writing of the manuscript: Parajuli, Soder, Hristov, and Petkov.
References
- Andersson KE, Arner A. (2004) Urinary bladder contraction and relaxation: physiology and pathophysiology. Physiol Rev 84:935–986 [DOI] [PubMed] [Google Scholar]
- Andersson KE, Wein AJ. (2004) Pharmacology of the lower urinary tract: basis for current and future treatments of urinary incontinence. Pharmacol Rev 56:581–631 [DOI] [PubMed] [Google Scholar]
- Brading AF. (1992) Ion channels and control of contractile activity in urinary bladder smooth muscle. Jpn J Pharmacol 58 (Suppl 2):120P–127P [PubMed] [Google Scholar]
- Brähler S, Kaistha A, Schmidt VJ, Wölfle SE, Busch C, Kaistha BP, Kacik M, Hasenau AL, Grgic I, Si H, et al. (2009) Genetic deficit of SK3 and IK1 channels disrupts the endothelium-derived hyperpolarizing factor vasodilator pathway and causes hypertension. Circulation 119:2323–2332 [DOI] [PubMed] [Google Scholar]
- Brown SM, Bentcheva-Petkova LM, Liu L, Hristov KL, Chen M, Kellett WF, Meredith AL, Aldrich RW, Nelson MT, Petkov GV. (2008) β-Adrenergic relaxation of mouse urinary bladder smooth muscle in the absence of large-conductance Ca2+-activated K+ channel. Am J Physiol Renal Physiol 295:F1149–F1157 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen M, Kellett WF, Petkov GV. (2010) Voltage-gated K+ channels sensitive to stromatoxin-1 regulate myogenic and neurogenic contractions of rat urinary bladder smooth muscle. Am J Physiol Regul Integr Comp Physiol 299:R177–R184 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen M, Petkov GV. (2009) Identification of large conductance calcium activated potassium channel accessory β4 subunit in rat and mouse bladder smooth muscle. J Urol 182:374–381 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Damkjaer M, Nielsen G, Bodendiek S, Staehr M, Gramsbergen JB, de Wit C, Jensen BL, Simonsen U, Bie P, Wulff H, et al. (2011) Pharmacological activation of KCa3.1/KCa2.3 channels produces endothelial hyperpolarisation and lowers blood pressure in conscious dogs. Br J Pharmacol http://dx.doi.org/10.1111/j.1476-5381.2011.01546.x [DOI] [PMC free article] [PubMed]
- Epstein BJ, Gums JG, Molina E. (2006) Newer agents for the management of overactive bladder. Am Fam Physician 74:2061–2068 [PubMed] [Google Scholar]
- Fry CH, Sui GP, Severs NJ, Wu C. (2004) Spontaneous activity and electrical coupling in human detrusor smooth muscle: implications for detrusor overactivity? Urology 63:3–10 [DOI] [PubMed] [Google Scholar]
- Hasenau AL, Nielsen G, Morisseau C, Hammock BD, Wulff H, Kohler R. (2011) Improvement of endothelium-dependent vasodilations by SKA-31 and SKA-20, activators of small- and intermediate-conductance Ca2+-activated K+ -channels. Acta Physiol (Oxf) 203:117–126 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hashitani H, Brading AF. (2003) Ionic basis for the regulation of spontaneous excitation in detrusor smooth muscle cells of the guinea-pig urinary bladder. Br J Pharmacol 140:159–169 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hashitani H, Bramich NJ, Hirst GD. (2000) Mechanisms of excitatory neuromuscular transmission in the guinea-pig urinary bladder. J Physiol 524:565–579 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hashitani H, Suzuki H. (1995) Electrical and mechanical responses produced by nerve stimulation in detrusor smooth muscle of the guinea-pig. Eur J Pharmacol 284:177–183 [DOI] [PubMed] [Google Scholar]
- Heppner TJ, Bonev AD, Nelson MT. (1997) Ca2+-activated K+ channels regulate action potential repolarization in urinary bladder smooth muscle. Am J Physiol Cell Physiol 273:C110–C117 [DOI] [PubMed] [Google Scholar]
- Herrera GM, Etherton B, Nausch B, Nelson MT. (2005) Negative feedback regulation of nerve-mediated contractions by KCa channels in mouse urinary bladder smooth muscle. Am J Physiol Regul Integr Comp Physiol 289:R402–R409 [DOI] [PubMed] [Google Scholar]
- Herrera GM, Heppner TJ, Nelson MT. (2000) Regulation of urinary bladder smooth muscle contractions by ryanodine receptors and BK and SK channels. Am J Physiol Regul Integr Comp Physiol 279:R60–R68 [DOI] [PubMed] [Google Scholar]
- Herrera GM, Nelson MT. (2002) Differential regulation of SK and BK channels by Ca2+ signals from Ca2+ channels and ryanodine receptors in guinea-pig urinary bladder myocytes. J Physiol 541:483–492 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herrera GM, Pozo MJ, Zvara P, Petkov GV, Bond CT, Adelman JP, Nelson MT. (2003) Urinary bladder instability induced by selective suppression of the murine small conductance calcium-activated potassium (SK3) channel. J Physiol 551:893–903 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hougaard C, Fraser MO, Chien C, Bookout A, Katofiasc M, Jensen BS, Rode F, Bitsch-Nørhave J, Teuber L, Thor KB, et al. (2009) A positive modulator of KCa 2 and KCa 3 channels, 4,5-dichloro-1,3-diethyl-1,3-dihydro-benzoimidazol-2-one (NS4591), inhibits bladder afferent firing in vitro and bladder overactivity in vivo. J Pharmacol Exp Ther 328:28–39 [DOI] [PubMed] [Google Scholar]
- Hristov KL, Chen M, Kellett WF, Rovner ES, Petkov GV. (2011) Large conductance voltage- and Ca2+-activated K+ channels regulate human detrusor smooth muscle function. Am J Physiol Cell Physiol 301:C903–C912 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hristov KL, Cui X, Brown SM, Liu L, Kellett WF, Petkov GV. (2008) Stimulation of β3-adrenoceptors relaxes rat urinary bladder smooth muscle via activation of the large-conductance Ca2+-activated K+ channels. Am J Physiol Cell Physiol 295:C1344–C1353 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Imai T, Okamoto T, Yamamoto Y, Tanaka H, Koike K, Shigenobu K, Tanaka Y. (2001) Effects of different types of K+ channel modulators on the spontaneous myogenic contraction of guinea-pig urinary bladder smooth muscle. Acta Physiol Scand 173:323–333 [DOI] [PubMed] [Google Scholar]
- Nielsen JS, Rode F, Rahbek M, Andersson KE, Rønn LC, Bouchelouche K, Nordling J, Bouchelouche P. (2011) Effect of the SK/IK channel modulator 4,5-dichloro-1,3-diethyl-1,3-dihydro-benzoimidazol-2-one (NS4591) on contractile force in rat, pig and human detrusor smooth muscle. BJU Int 108:771–777 [DOI] [PubMed] [Google Scholar]
- Ohya S, Kimura S, Kitsukawa M, Muraki K, Watanabe M, Imaizumi Y. (2000) SK4 encodes intermediate conductance Ca2+-activated K+ channels in mouse urinary bladder smooth muscle cells. Jpn J Pharmacol 84:97–100 [DOI] [PubMed] [Google Scholar]
- Ohya S, Niwa S, Yanagi A, Fukuyo Y, Yamamura H, Imaizumi Y. (2011) Involvement of dominant-negative spliced variants of the intermediate conductance Ca2+-activated K+ channel, K(Ca)3.1, in immune function of lymphoid cells. J Biol Chem 286:16940–16952 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pandita RK, Rønn LC, Jensen BS, Andersson KE. (2006) Urodynamic effects of intravesical administration of the new small/intermediate conductance calcium activated potassium channel activator NS309 in freely moving, conscious rats. J Urol 176:1220–1224 [DOI] [PubMed] [Google Scholar]
- Petkov GV, Bonev AD, Heppner TJ, Brenner R, Aldrich RW, Nelson MT. (2001a) β1-Subunit of the Ca2+-activated K+ channel regulates contractile activity of mouse urinary bladder smooth muscle. J Physiol 537:443–452 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petkov GV, Heppner TJ, Bonev AD, Herrera GM, Nelson MT. (2001b) Low levels of KATP channel activation decrease excitability and contractility of urinary bladder. Am J Physiol Regul Integr Comp Physiol 280:R1427–R1433 [DOI] [PubMed] [Google Scholar]
- Petkov GV, Nelson MT. (2005) Differential regulation of Ca2+-activated K+ channels by β-adrenoceptors in guinea pig urinary bladder smooth muscle. Am J Physiol Cell Physiol 288:C1255–C1263 [DOI] [PubMed] [Google Scholar]
- Sankaranarayanan A, Raman G, Busch C, Schultz T, Zimin PI, Hoyer J, Köhler R, Wulff H. (2009) Naphtho[1,2-d]thiazol-2-ylamine (SKA-31), a new activator of KCa2 and KCa3.1 potassium channels, potentiates the endothelium-derived hyperpolarizing factor response and lowers blood pressure. Mol Pharmacol 75:281–295 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shakkottai VG, do Carmo Costa M, Dell'Orco JM, Sankaranarayanan A, Wulff H, Paulson HL. (2011) Early changes in cerebellar physiology accompany motor dysfunction in the polyglutamine disease spinocerebellar ataxia type 3. J Neurosci 31:13002–13014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stocker M. (2004) Ca2+-activated K+ channels: molecular determinants and function of the SK family. Nat Rev Neurosci 5:758–770 [DOI] [PubMed] [Google Scholar]
- Strøbaek D, Teuber L, Jørgensen TD, Ahring PK, Kjaer K, Hansen RS, Olesen SP, Christophersen P, Skaaning-Jensen B. (2004) Activation of human IK and SK Ca2+ -activated K+ channels by NS309 (6,7-dichloro-1H-indole-2,3-dione 3-oxime). Biochim Biophys Acta 1665:1–5 [DOI] [PubMed] [Google Scholar]
- Tharp DL, Bowles DK. (2009) The intermediate-conductance Ca2+ -activated K+ channel (KCa3.1) in vascular disease. Cardiovasc Hematol Agents Med Chem 7:1–11 [DOI] [PubMed] [Google Scholar]
- Thorneloe KS, Knorn AM, Doetsch PE, Lashinger ES, Liu AX, Bond CT, Adelman JP, Nelson MT. (2008) Small-conductance, Ca2+ -activated K+ channel 2 is the key functional component of SK channels in mouse urinary bladder. Am J Physiol Regul Integr Comp Physiol 294:R1737–R1743 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xia XM, Fakler B, Rivard A, Wayman G, Johnson-Pais T, Keen JE, Ishii T, Hirschberg B, Bond CT, Lutsenko S, et al. (1998) Mechanism of calcium gating in small-conductance calcium-activated potassium channels. Nature 395:503–507 [DOI] [PubMed] [Google Scholar]