

Abstract
The outer membrane vesicles (OMVs) from smooth B. melitensis 16 M and a derived rough mutant, VTRM1 strain, were purified and characterized with respect to protein content and induction of immune responses in mice. Proteomic analysis showed 29 proteins present in OMVs from B. melitensis 16 M; some of them are well-known Brucella immunogens such as SOD, GroES, Omp31, Omp25, Omp19, bp26, and Omp16. OMVs from a rough VTRM1 induced significantly higher expression of IL-12, TNFα, and IFNγ genes in bone marrow dendritic cells than OMVs from smooth strain 16 M. Relative to saline control group, mice immunized intramuscularly with rough and smooth OMVs were protected from challenge with virulent strain B. melitensis 16 M just as well as the group immunized with live strain B. melitensis Rev1 (P < 0.005). Additionally, the levels of serum IgG2a increased in mice vaccinated with OMVs from rough strain VTRM1 consistent with the induction of cell-mediated immunity.
1. Introduction
The release of outer membrane vesicles (OMVs) from bacteria is a phenomenon described about 40 years ago. OMVs are released spontaneously during the normal growth of Gram-negative bacteria [1–3]. OMVs have been described in both pathogenic and nonpathogenic Gram-negative bacteria such as Escherichia coli [4, 5], Shigella spp. [6, 7], Neisseria spp. [8], Porphyromonas spp. [9], Pseudomonas aeruginosa [7], Helicobacter pylori [10], Vibrio spp. [11], Salmonella spp. [12], Brucella spp. [13, 14], Actinobacillus spp. [15, 16], Xenorhabdus nematophilus [17], and Pseudoalteromonas antarctica [18].
OMVs possess a bilayer membrane and contain components such as lipoproteins, outer membrane proteins (OMP), lipopolysaccharide (LPS), and some periplasmic components [1–3]. OMVs have been implicated in many processes including the release of virulence factors such as proteases and toxins, signaling between bacterial and eukaryotic cells, DNA transfer, antibacterial activity, immunomodulation of the host, and facilitation of bacterial survival during envelope stress [2, 3, 19, 20].
Other studies have revealed that OMVs trigger the innate inflammatory response. For example, OMVs from Salmonella typhimurium are able to activate dendritic cells to secrete IL-12 and TNFα [12], and OMVs from Pseudomonas aeruginosa and Helicobacter pylori are able to elicit IL-8 production by epithelial cells [21, 22].
The use of OMVs from different Gram-negative bacteria as acellular vaccines has been explored in recent years [23–26]. OMV vaccines have been effective in the specific case of serogroup B of Neisseria meningitis [24]. More recently, OMVs from Vibrio cholerae and Bordetella pertusis were demonstrated to elicit protection in mouse model [26, 27]. The interest in OMVs as vaccine carriers is increasing, and recent reports have showed that engineered OMVs were able to harbor overexpress antigens [28].
Brucellosis is a worldwide spread zoonotic disease transmitted from domestic animals to humans. It is frequently acquired by ingestion, inhalation, or direct contact of conjunctiva or skin-lesions with infected animal products. The human disease represents an important cause of morbidity worldwide whereas animal brucellosis is associated with serious economical losses caused mainly by abortion and infertility in ruminants [29].
The first effective Brucella vaccine was based on live Brucella abortus strain 19 (S19), a smooth strain attenuated by an unknown process induced by its subculturing. This strain induces reasonable protection against B. abortus in cattle, but at the expense of persistent serological responses that confound differential serodiagnosis of vaccinated and field- infected cattle. A similar problem occurs with the B. melitensis Rev.1 strain that is still the most effective vaccine against caprine and ovine brucellosis. This problem has been overcome in cattle by the development of the rifampicin-resistant mutant B. abortus RB51 strain. This strain has been proven safe and effective in the field against bovine brucellosis and exhibits negligible interference with diagnostic serology [30].
Currently, smooth live attenuated vaccines B. abortus S19 and B. melitensis Rev1 as well rough live attenuated vaccine B. abortus RB51 are used in the control of animal brucellosis. These smooth vaccines for animals may cause disease and considered unsuitable for use in humans; the rough strain RB51 is rifampin resistant and is considered unsuitable for humans as rifampin is one of the antibiotics of choice for therapy [31]. In the last few decades much research has been done in the attempt to develop safer Brucella vaccines [32]. It is important to mention that there is no licensed vaccine for prevention of human brucellosis. A human vaccine could be useful to protect farmers, veterinarians, animal care workers, and general populations living in endemic brucellosis areas [30].
Since OMVs from other bacteria have been used for development of acellular vaccines, we were interested in assessing the protective immune response induced by Brucella OMVs. The first studies related to OMVs isolated from Brucella spp. were limited to analysis of their protein profile using one-dimensional SDS-PAGE [14, 33]. More recently, Omp25 and Omp31 were identified in B. suis OMVs using monoclonal antibodies [13]. In 2007, Lamontagne et al. performed a proteomic analysis of a fraction they called outer membrane fragments from virulent B. abortus 2308 and attenuated BvrR/BvrS mutants [34]. To date the composition of OMVs from B. melitensis has not been yet explored.
In the attempt to increase the current understanding of the composition of B. melitensis OMVs, the proteome of OMVs isolated from smooth B. melitensis 16 M is described. Because of the distinct immunological role of the Brucella O-side chain in the host, mice were immunized with OMVs purified from smooth B. melitensis 16 M and the rough mutant B. melitensis VTRM1 (lacking the side O chain of LPS). The difference in dendritic cell cytokine expression and the serum IgG subtypes levels as well as the level of protection afforded to mice is also described.
2. Materials and Methods
2.1. Ethics Statement
The mice experiments were approved and conducted by Institutional Animal Care and Use Committee (approved protocol and 07-047CVM) at Virginia Tech.
2.2. Bacterial Strains and Growth Conditions
B. melitensis 16 M (ATCC 23456) and B. melitensis VTRM1 rough mutant derived from B. melitensis 16 M [35] were used. Both strains were cultured on tryptic soy agar (TSA) plates supplemented with 0.7% yeast extract and incubated 36 h at 37°C. A bacterial suspension was obtained from both strains, adjusted each to 0.5 g of cells per mL of tryptic soy broth, of which 0.5 mL was spread onto each of 100 TSA plates (10 cm diameter) and incubated at 37°C for 48 h.
2.3. OMVs Purification
The OMVs purification was performed according to the protocol described by Gamazo et al., 1989. Briefly, the bacteria were harvested with a rubber policeman and suspended in 250 mL sterile phosphate-buffered saline (PBS 0.1 M, pH 7.3). The bacterial suspension was centrifuged at 10,000 ×g for 30 min. The supernatant was passed through a 0.22 μm filter (Millipore Corp.), and a sterility test was performed by culturing an aliquot onto a TSA plate followed by incubation for 72 h at 37°C; the filtrate was stored at 4°C during the viability check. The filtered supernatant was centrifuged at 100,000 ×g for 2 h at 4°C. The pellet was washed twice with 25 mL of sterile PBS, and the OMVs were resuspended in 1 mL of sterile PBS. The total protein concentration was determined using PIERCE-BCA (PIERCE) reagents as per manufacturer's recommendations. The OMVs samples were divided into 0.5 mL aliquots and stored at −20°C until used [33].
2.4. Bone Marrow-Derived Dendritic Cells (BMDC)
Dendritic cells were derived from 8 wk old, female BALB/c mice by in vitro culture of bone-marrow cells with 20 ng/mL rGM-CSF for 7 days as previously described [36, 37]. On day 7, cells showed differentiated morphology (BMDC) and expressed DC markers (CD11c+) in 75% of the population as assessed by flow cytometry (data not shown).
2.5. In Vitro Stimulation of BMDC, RNA Extraction, and Reverse-Transcription Polymerase Chain Reaction
Aliquots of 2.5 × 106 BMDC per well were plated in a 6-well flat-bottomed plate by triplicate and incubated overnight. Then 40 μg of purified OMVs from smooth B. melitensis 16 M or OMVs from rough B. melitensis VTRM1 were added to each well by triplicate. Total RNA (RNAeasy Qiagen) was extracted from BMDC (stimulated and unstimulated) at 1, 3, 6, and 12 h after induction. The DNA was removed with DNase I (DNA-free Kit Ambion). Then cDNA was prepared from 1 μg of total RNA (Promega, A2500 kit). To verify the complete elimination of DNA, PCR for glyceraldehyde-3-phosphate dehydrogenase (GAPDH) gene amplification was performed (data not shown).
2.6. Real Time-PCR
Templates cDNA were analyzed for IL-2, IL-6, IL-12p40 (IL-12), IL-10, IL-17, IL-23, INF-γ, TNF-α, and TGF-β (SABioscienes) expression using the PCR Array and the RT2 SYBR Green/Fluorescein qPCR Master Mix (SABiosciences) on the iCycler PCR System (Bio-Rad) as per recommendations of the manufacturer. Fold changes in gene expression were calculated using the ΔΔCt method in the PCR Array Data Analysis template. The amplification of house-keeping gapdh gene was used to normalize the fold changes in the cytokine expression.
2.7. Mice Immunizations
Female BALB/c mice of 6 weeks of age (5 per group) were vaccinated by two intramuscular inoculations, at day 0 and day 30, with 5 μg of purified OMVs from B. melitensis 16 M and B. melitensis VTRM1. Before the first dose, mice were prebled by puncturing retro-orbital plexus under anesthesia. Two weeks after boosting, the mice were bled by the same route. The serum was separated from the clotted blood and stored at −20°C until use for detection of IgG subtypes. As a positive control, a group of mice was vaccinated with 1.5 × 104 CFU of vaccine strain, B. melitensis Rev1. As a negative control, one group of mice was injected with saline. Mice were challenged at 6 weeks after the first vaccination dose with 5 × 104 CFUs of virulent strain B. melitensis 16 M by intraperitoneal route. At 2 weeks after challenge, all the mice were euthanized by CO2 asphyxiation followed by cervical dislocation, spleens were collected aseptically, and colony-forming units (CFU) were determined.
2.8. Indirect ELISA
Levels of serum immunoglobulin IgG1 and IgG2a isotypes with specificity to OMVs from smooth B. melitensis 16 M and rough strain VTRM1 were determined by indirect ELISA. Sera from mice immunized with OMVs purified from B. melitensis 16 M were tested with OMVs purified from B. melitensis 16 M, and sera from mice immunized with OMVs purified from rough B. melitensis VTRM1 were tested with OMVs purified from rough strain. The OMVs were diluted in carbonate buffer, pH 9.6. The wells of polystyrene plates (Nunc-Immunoplate with maxisorp surface) were coated with 100 μL/well of the diluted antigens (2.5 μg/mL of protein from OMVs). Following overnight incubation at 4°C, plates were washed four times in wash buffer (Tris-buffered saline at pH 7.4, .05% Tween 20) and blocked with 2% bovine serum albumin (BSA) in Tris-buffered saline. After 1 h incubation at 37°C, mice sera with appropriate dilution in blocking buffer were added to the wells (50 μL/well). Each serum sample was tested in triplicate wells; the plates were incubated for 4 h at room temperature and washed four times. Horse radish peroxidase-labeled anti-mouse isotype-specific conjugates (Southern Biotechnology Associates Inc, Birmingham, Alabama) were added (50 μL/well) at an appropriate dilution. After 1 h incubation at room temperature, the plates were washed four times. A 100 μL of substrate solution (TMB Microwell peroxidase substrate; Kirkegaard and Perry Laboratories, Gaithersburg, Md) was applied to each well. After 20 min incubation at room temperature, the enzyme reaction was stopped by adding 100 μL of stop solution (0.185 M sulfuric acid), and the absorbance at 492 nm was recorded using microplate reader (Molecular Devices, Sunnyvale, Calif).
2.9. Electron Microscopy
20 μL of purified OMVs (25 μg of protein) or intact bacteria were placed onto copper grids coated with formvar and dried using filter paper. 40 μL of 1% phosphotungstic acid was added, and the grids were allowed to stand overnight at room temperature; they were observed under the transmission electron microscope (JEOL model JEM 10-10).
2.10. Denaturing Gel Electrophoresis
SDS-PAGE was performed in 15% acrylamide slab gels by the method of Laemmli [38]; the gels were stained with Coomassie blue. The apparent molecular masses of the OMV proteins were determined by comparing their electrophoretic mobility with that of the wide-range molecular mass markers [SigmaMarker (Sigma)] using the computer program SigmaGel V. 1.0.
2.11. Enzymatic Digestion
After the separation of OMV proteins by denaturing electrophoresis, the acrylamide gel was cut into six sections. The excised samples were reduced with 50 mM dithiothreitol, alkylated with iodoacetamide and then “in gel” digested with trypsin. The peptides were desalted using a ZipTip (Millipore Corp) and then concentrated in a Speed-Vac SPD 1010 ThermoElectron (Instituto Nacional de Biotecnologia-UNAM, Cuernavaca, México).
2.12. LC-MS/MS
The samples were reconstituted to approximately 0.1–0.5 μg/μL in 50% acetonitrile containing 1% acetic acid and placed directly into a Finnigan LCQ ion trap mass spectrometer (Instituto Nacional de Biotecnologia-UNAM, Cuernavaca, Mexico), using a Surveyor MS syringe pump delivery system. The eluate at 10 μL/min was split to allow only 5% of the sample to enter the nanospray source (0.5 μL/min). LC-MS/MS analyses were carried out using a PicoFrit needle/column RP C18 from New Objective (Woburn, Mass, USA), with a fast gradient system from 5% to 60% of solution B (100% acetonitrile containing 1% acetic acid) for a period of 45 min.
The electrospray ionization source voltage was set at 1.8 kV and the capillary temperature at 130°C. Collision-Induced Dissociation (CID) was performed using 25 V of collision energy, 35–45% (arbitrary units) of normalized collision energy and the scan had the wide band activated.
All spectra were obtained in the positive-ion mode. Data acquisition and the deconvolution of data were carried out using Xcalibur software on a Windows XP PC system. The MS/MS spectra from enzymatically generated peptides were analyzed by Sequest software from Finnigan (Palo Alto, Calif) and MASCOT search engine from Matrix Science Ltd (Boston, Mass).
2.13. In Silico Analysis
Once the proteins in the OMVs were identified, an in silico analysis was performed. Initially the amino acid sequences of the identified proteins were analyzed by BLAST comparing them to similar sequences from species of Brucella and other bacteria (http://www.ncbi.nlm.nih.gov/BLAST/). The isoelectric point and molecular weight were determined using Antheprot 2000 V. 5.2.
The prediction of “motif” sequences was performed by searching My Hits Motif Scan database (http://hits.isb-sib.ch/cgi-bin/PFSCAN) that uses different databases such as PROSITE, HAMAP, and Pfam. The prediction of the subcellular location of the proteins was carried out on the PSORT server available at http://www.psort.org [39] and Softberry database. The MatGAT V. 2.02 [40] program was used to determinate the similitude grade of the homologous sequences found by the BLAST search. The ProLinks database (http://dip.doe-mbi.ucla.edu/pronav/) [41] and Gene Ontology (http://www.geneontology.org) [42] were used to determinate the hypothetical function of proteins into OMVs.
2.14. Statistical Analysis
Statistical analysis, Two-Way-ANOVA, was carried out with SigmaStat statistical package V. 2.0 (SYSTAT).
3. Results
3.1. Isolation of OMVs and Electron Microscopy
OMVs were isolated from cell-free culture medium by differential centrifugation. In order to confirm purification of the OMVs from both strains, electron micrographs were performed using negative staining with phosphotungstic acid. In the micrographs (Figure 1), it is possible to see the bleb formation leading to the liberation of OMVs from the outer surface of the Brucella. In addition, the spherical morphology (average diameter 60–90 nm) of the purified OMVs including a double membrane can be observed as previously described [14, 33]. Moreover, no membrane debris bigger than 100 nm were observed by electron microscopy. In general, no differences, at least in the shape or in the number of OMVs released, were observed for the smooth or rough Brucella strains.
Figure 1.

Electron microscopy of OMVs from smooth B. melitensis M16 and rough B. melitensis VTRM1. Negative stain of OMVs released from surface B. melitensis VTRM1 (a) and B. melitensis 16 M (b); negative stain of OMV purified by differential centrifugation from both strains; B. melitensis VTRM1 (c) and B. melitensis 16 M (d). The arrows point to the apparent shedding of the OMVs from the cell surface in (a) and (b); while the arrows in (c) and (d) point to spherical OMVs purified from both strains. The bars correspond to 100 nm.
3.2. Cytokine Expression
In order to explore if OMVs could induce an immune response in antigen-presenting cells, we used BMDC exposed to OMVs either from smooth or rough B. melitensis. At increasing times following exposure, the cytokines associated with the DC1-mediated Th1 (IFN-γ, IL-2, IL-6, IL-12, and TNF-α), DC2-mediated Th2 (IL-4 and IL-10), and DC17-mediated Th17 (IL-17, IL-23, and TGF-β) responses were measured by qRT-PCR. The results are shown in Table S1 (see in table S1 Supplementary Material available online at doi: 10.1155/2012/352493). The cytokine profiles elicited in the BMDC stimulated with smooth B. melitensis 16 M OMVs reached a maximum at 12 hours following stimulation with the highest production of IL-6, IL-4, IL-10, and IL-17 (Figure 2(a)). In contrast, the induction of cytokines by rough OMVs reached maximum expression at 1 h after stimulation and decreased over time, except for IL-10 (maximum expression at 3 hours) and for TNF-α (maximum expression at 3 hours) (Figure 2(b)). Using statistical analysis we compared the cytokine expression obtained from BMDC stimulated with OMVs from smooth and rough Brucella strains (Two-way-ANOVA analysis). Results showed significant differences between the cytokine profile induced by OMVs purified from rough and smooth B. melitensis (P < 0.05). OMVs from rough B. melitensis VTRM1 induced three cytokines that were significantly higher: IFN-γ, TNF-α, and IL-12 (P < 0.05, P < 0.01 and P < 0.001, resp.).
Figure 2.
Cytokine expression of BMDC stimulated with OMVs. (a) BMDC stimulated with OMVs from smooth B. melitensis 16 M. (b) BMDC stimulated with OMVs from rough B. melitensis VTRM1. Two-way ANOVA analysis was performed to compare results. *(P < 0.05); **(P < 0.01); ***(P < 0.001).
3.3. Protection against Challenge with Virulent B. melitensis 16 M
Mice were challenged with virulent strain B. melitensis 16 M to examine the protection induced by OMVs. In this experiment, protection was defined as a significant reduction in the number of bacteria in the spleens of immunized mice compared to the mice receiving saline. The B. melitensis Rev1 vaccine induced 2.64 log units of protection compared to saline control (Figure 3). In the case of mice vaccinated with OMVs, we observed that smooth OMVs induced 1.9 log units, and rough OMVs induced 3.08 log units of protection compared to saline control (P < 0.005).
Figure 3.
Analysis of IgG1 (a) and IgG2a (b) antibody responses of BALB/c mice to outer membrane vesicles from Brucella. Outer membrane vesicles (OMVs) were purified from B. melitensis 16 M and B. melitensis VTRM1, and mice were immunized. Sera from each mouse were collected and were assayed individually by ELISA. Antibody levels are expressed as optical density (OD) at 492 nm.
3.4. Serology
Presence of antibodies specific to OMVs in serum of the mice vaccinated was determined by ELISA. The induction of IgG2a subclass during immune response should give an idea about Th1 or Th2 balance. As illustrated in Figures 4(a) and 4(b) OMVs purified from rough B. melitensis VTRM1 induced a higher IgG2a response than IgG1, suggesting a shift toward a Th1 response. In contrast, OMVs from smooth strain B. melitensis 16 M induced the same levels of IgG1 and IgG2a.
Figure 4.
Level of protection against B. melitensis 16 M conferred by outer membrane vesicles purified from B. melitensis 16 M and B. melitensis VTRM1. In this experiment vaccine strain B. melitensis Rev1 was used as a positive control of vaccination, as negative control was used saline. n = 5 (*P ≤ 0.005 comparisons were OMVs versus saline, and B. melitensis Rev1 versus saline).
3.5. SDS-PAGE and Proteomic Analysis
The denatured electrophoretic protein profiles observed from OMVs obtained from rough B. melitensis VTRM1 and smooth B. melitensis 16 M show no discernable differences (Figure 5). The trypsin-generated peptide masses, as well as their fragment ions, were analyzed by LC-MS/MS. The resulting peptides sequences were used to query databases that led to the identification of 29 unique proteins (Table 1). A query result was only considered as significant if the overall score was higher than 25 and more than two tryptic peptides as well as their fragment ions matched to the protein and the calculated molecular weight corresponded to molecular weight in the original gel section [43].
Figure 5.
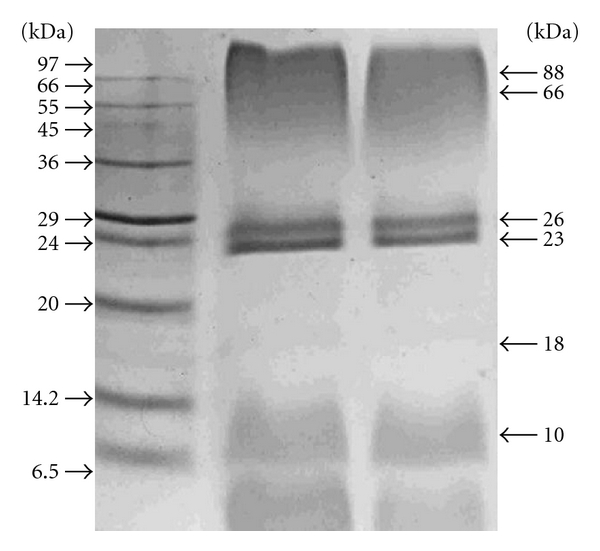
Electrophoretic profiles of OMVs purified from B. melitensis. Lane 1, molecular weight markers; lane 2, OMVs from smooth B. melitensis 16 M (80 μg); lane 3, OMVs from rough B. melitensis VTRM1 (80 μg). SDS-PAGE gels were stained with Coomassie blue.
Table 1.
B. melitensis 16 M OMVs proteins identified by 1D-SDS-PAGE coupled to LC-MS/MS.
| Protein | B. melitensis denomination | Molecular weight (kDa) | Motif, subcellular localization | Closest ortholog, other than in Brucellae (% similarity) |
|---|---|---|---|---|
| Bacterial surface antigen | BMEI0830 | 85.90 | Surface antigen, OM | Outer membrane protein Bartonella henselae str. Houston-1, (75.3%) |
| Iron-regulated outer membrane protein FRPB | BMEII0105 | 72.05 | TonB-dependent receptor Plug Domain, OM | TonB-dependent receptor Escherichia coli, (41.3%) |
| Metal chelate outer membrane receptor | BMEI0657 | 64.80 | TonB-dependent receptor Plug Domain, OM | Chain A, Outer Membrane Cobalamin Transporter (Btub) Escherichia coli, (46.2%) |
| Sugar-binding protein | BMEII0590 | 43.20 | Bacterial sugar-binding extracellular protein, P | Probable sugar ABC transporter substrate binding protein Rhizobium etli, (75.7%) |
| Outer surface protein | BMEII0376 | 31.55 | Surface antigen, OM | Probable heat-resistant agglutinin 1 protein Rhizobium leguminosarum bv. viciae, (54.9%) |
| D-Ribose-binding periplasmic protein precursor | BMEII0435 | 30.99 | Periplasmic binding protein and sugar-binding domain of LacI family, P | Porin Rhizobium leguminosarum bv. trifolii, (59.2%) |
| Hypothetical protein BMEI0542 | BMEI0542 | 30.04 | Unknown, EC | Hypothetical protein Rhizobium sp., (48.3%) |
| 25 kDa outer-membrane immunogenic protein precursor | BMEI1007 | 25.24 | Porin type 2, OM | Hemin-binding C protein Bartonella tricoborum, (58.7%) |
| 25 kDa outer-membrane immunogenic protein precursor | BMEI1249 | 23.18 | Porin type 2, OM | Hemin-binding B protein Bartonella henselae str. Houston-1, (58.7%) |
| 25 kDa outer-membrane immunogenic protein precursor | BMEI1829 | 24.58 | Porin type 2, OM | Hemin-binding C protein Bartonella tricoborum, (45.4%) |
| 25 kDa outer-membrane immunogenic protein precursor | BMEI1830 | 24.74 | Porin type 2, OM | Outer membrane protein Rhizobium etli, (60.1%) |
| 31 kDa outer-membrane immunogenic protein precursor | BMEII0844 | 23.27 | OmpA-like domain profile, OM | Porin Rhizobium leguminosarum bv. trifolii, (52.5%) |
| BP26 | BMEI0536 | 24.77 | Protein of unknown function (DUF541), P | Unknown function protein DUF541 Rhizobium leguminosarum bv. trifolii, (64.9%) |
| Precursor YBIS protein | BMEI1369 | 23.51 | Domain YkuD, C | Hypothetical protein Rhizobium leguminosarum bv. viciae, (67.2%) |
| OmpA family protein | BMEI0786 | 22.96 | OmpA-like domain, OM | OmpA family protein Rhizobium leguminosarum bv. viciae, (81.4%) |
| Hypothetical lipoprotein | BMEI0785 | 21.91 | Prokaryotic membrane lipoprotein lipid attachment profile, IM | Hypothetical protein Rhizobium leguminosarum bv. viciae, (74.7%) |
| Ribosome recycling factor | BMEI0826 | 20.66 | Ribosome recycling factor (RRF-frr), C | Ribosome recycling factor Bartonella henselae str. Houston-1, (90.3%) |
| Hypothetical membrane-associated protein (BMEII0692) | BMEII0692 | 20.42 | Invasion-associated locus B (IalB) protein, IM | Invasion-associated locus B family protein Rhizobium leguminosarum bv. trifolii, (62.4%) |
| 22 kDa outer membrane protein precursor | BMEI0717 | 19.44 | Unknown, OM | Outer membrane protein putative precursor Rhizobium leguminosarum bv. viciae, (48.6%) |
| DNA starvation/stationary phase protection protein Dps | BMEI1980 | 18.25 | Dps protein family Ferritin-like domain, C | DNA starvation/stationary phase protection protein Dps Escherichia coli, (72.5%) |
| Peptidoglycan-associated lipoprotein | BMEI0340 | 18.23 | OmpA family protein, OM | Outer membrane lipoprotein Rhizobium etli, (82.6%) |
| Invasion protein B | BMEI1584 | 18.03 | Invasion-associated locus B (IalB) protein, P | Invasion-associated locus B protein Bartonella quintana str. Tolouse, (55.4%) |
| Outer membrane lipoprotein | BMEI0135 | 17.60 | Bacterial outer membrane lipoprotein; Omp19, OM | Outer membrane lipoprotein Bartonella henselae str. Houston-1, (59.6%) |
| Chain A, Cu-Zn superoxide dismutase | BMEII0581 | 16.07 | Cu-Zn superoxide dismutase, P | Cu-Zn superoxide dismutase Escherichia coli, (68.7%) |
| Thioredoxin C-1 | BMEI2022 | 11.42 | Thioredoxin active site, C | Putative thioredoxin Rhizobium leguminosarum bv. viciae, (88.8%) |
| Cochaperonin GroES | BMEII1047 | 10.39 | 10 kDa chaperonin protein Cnp10, C | Cochaperonin GroES Bartonella tricoborum, (91.8%) |
| DNA-binding protein HU | BMEI0877 | 9.07 | Bacterial histone-like DNA binding protein signature, C | DNA-binding protein Rhizobium sp., (83.5%) |
| Hypothetical lipoprotein | Unknown | 8.286 | Prokaryotic membrane lipoprotein lipid attachment profile, OM | 17 kDa surface antigen Rhizobium leguminosarum bv. trifolii, (61.2%) |
| Hypothetical protein BMEI0287 | BMEI0287 | 8.596 | Prokaryotic membrane lipoprotein lipid attachment profile, OM | Hypothetical protein Rhizobium etli, (56.8%) |
3.6. In Silico Analysis
The results of the proteomic characterization of the B. melitensis vesicular proteins are summarized in Table 1. In addition, the complete results are available in supporting information Table S2 and Table S3. The identification score varied from 55 to 100%, with coverage from 3 to 50%. Besides the identification of each spot and calculation of its basic biochemical characteristics, the isoelectric point and molecular weight was obtained by means of analysis and search in databases. Subcellular location, protein motifs, immunogenic regions, signal peptide prediction, and closest homologues were also analyzed. Of the 29 proteins identified, approximately 52% belonged to the outer membrane, 17% to the periplasm, 20.6% to the cytoplasm, 2 proteins were from inner membrane, and 1 protein was predicted to be an extracellular protein. Using the Motif databases these proteins were classified into five groups: (i) structural and transport proteins (such as the outer membrane proteins), (ii) antigenic proteins, (iii) involved in metabolic processes (e.g., Frr, HU, GroES), (iv) involved in stress response (e.g., Dps, TrxC, and SOD), and v) invasion proteins (e.g., InvB and IalB). About 60% of the proteins were predicted to contain signal peptides and thus capable to be exported or targeted by the cellular machinery to the periplasm or outer membrane. Only twelve proteins did not contain signal peptides, five of these were predicted as cytoplasmic or mature periplasmic proteins in which the signal sequence was processed.
Additionally, the analysis of sequences using ProLinks [41] showed that with the exception of two proteins, the possible functions of 27 proteins were predicted. While the analysis of sequences using Gene Ontology terms showed that one half (14/29) of the proteins appeared to be involved in transport and/or integrity of the membrane.
All the proteins found in the B. melitensis OMVs were highly related to homologous proteins in other Brucella species (from 88 to 100% in similarity, data not shown). For most proteins, the closest non-Brucella homologues were found in Rhizobia, such as Rhizobium and Bartonella (50% to 90% in similarity). Interestingly, four proteins with a high degree of homology to those in Escherichia coli were found: FrpB (an iron-regulated outer membrane protein), a metal chelate outer membrane receptor, Dps (involved in DNA protection due to starvation or stationary phase), and SOD (a Cu/Zn superoxide dismutase).
4. Discussion
As has been described in other Gram-negative bacteria, OMVs are released from the Brucella's outer membrane as we observed by electron microscopy (Figure 1). The OMVs are shed from both rough and smooth Brucella strains, grown in liquid or on solid media, and spontaneously released during the growth as observed previously [13, 14, 33].
OMVs can also strongly activate the host innate and acquired immune response pathways [23]. Based on this previous evidence, we stimulated BMDC with OMVs isolated from smooth and rough Brucella strains. At different time points, cytokines expression for DC1-mediated Th1 (IFN-γ, IL-2, IL-6, IL-12, and TNF-α), DC2-mediated Th2 (IL-4 and IL-10), and DC-mediated Th17 (IL-17, IL-23 and TGF-β) was analyzed by qRT-PCR. Interestingly, we determined that OMVs from rough B. melitensis VTRM1 induced significantly higher expression of IFN-γ, TNF-α, and IL-12.
TNF-α is necessary for full expression of the macrophage anti-Brucella activities. It also plays an important role in the triggering of specific immunity against several intracellular pathogens and positively controls early expression of IL-12 and IFN-γ in Brucella-infected mice [44]. Our results demonstrated an earlier expression of IL-12 and IFN-γ by OMVs from rough B. melitensis VTRM1 (Figure 2(b)). Clearly TNF-α participates in the establishment of acquired immunity of the Th1 response, with the generation of IFN-γ-producing CD4+ cells and CD8+ cytotoxic cells, two outcomes crucial for the complete killing of the intracellular Brucella. TNF-α produced a synergistic effect in presence of IFN-γ for the ultimate clearance of the infection [45].
The early expression of IL-6, IL-23, and TGF-β genes was observed in BMDC stimulated with OMVs from rough B. melitensis VTRM1. In contrast, in the case of smooth B. melitensis 16 M OMVs, the same cytokines genes were expressed after a longer time with a maximum expression at 12 h following induction. The early induction of the cytokine expression could offer a fast immune response against Brucella, for example, if IL-6 promotes a proinflammatory environment. However, the role of the IL-23 and TGF-β as part of the Th17 response in Brucella infection is not very clear to date. Some data suggest that a Th17 response is favored when IL-6 is present in high quantities, especially in aged mice [46].
The determination of whether Th17 response is merely present as epiphenomena or truly playing a role in the host defense is a focus of current research. We speculate that these differences could be the effect of the O-side chain in the LPS present in the OMVs from the smooth strain but not in the OMVs derived from rough B. melitensis VTRM1. In contrast to the infection with smooth B. abortus and B. suis strains and purified smooth LPS, infection of human DCs with rough mutants of Brucella leads to both phenotypic and functional maturation of infected cells [47].
Recently, Surendran et al. observed phenotypic maturation and production of IL-12 and TNF-α when murine BMDC were stimulated with live B. abortus RB51, a rough vaccine strain approved by the USDA for use in cattle and with B. abortus RB51SOD, which overexpressed a Cu-Zn superoxide dismutase (SOD) [37]. In contrast no maturation or secretion of either cytokine occurred when B. abortus strain 2308 was used to stimulate murine BMDC [47]. While Billard et al., [47] observed higher production of TNF-α and human DC maturation using rough mutants but not with smooth strains of B. suis, our cytokine expression results are in agreement with the works published by Billard et al., and Surendran et al. Those are in conflict with the reports by Zwerdling et al. and Macedo et al., where it was observed that smooth B. abortus exposure induced human and murine DC maturation, respectively [48]. Zwerdling et al. could only speculate the reasons for these discrepancies and some considerations were made such as: the different method for cell isolation, the concentration of the DC and the interactions between themselves in each experiment, and the type of strains used for those experiments [48].
The direct effect of purified rough or smooth LPS molecules on human DC maturation also has been explored [47]. However no difference could be determined between DC responses to rough or smooth LPS of B. abortus. These results are in line with the very low endotoxic properties of Brucella LPS and with the equivalent stimulation of macrophages by rough and smooth LPS [47]. This means that the ability of rough Brucella strains to induce human or murine DC maturation is not related to a direct effect of their LPS but maybe the absence of the O-side chain could allow the exposure of bacterial surface molecules that should normally be hidden. It is also possible that the absence of O-side chain in the rough OMVs permits a higher exposure to the outer membrane proteins. For example, it is well known that Omp16 and Omp19 interact with the TLR2 receptor and induce the production of IL-12, which is important for the control of infection [44].
Recently, it was demonstrated that Omp16 requires TLR4 interaction for the activation of DC and macrophages and elicits a Th1 and protective immune response [49]. On the other hand TLR4 is not involved in the Th1 and protective response induced by Omp19 [32].
After intramuscular administration of OMVs from B. melitensis 16 M and rough B. melitensis VTRM1, challenge with virulent B. melitensis 16 M was performed in BALB/c mice. Relative to saline control group, mice immunized intramuscularly with rough OMVs were protected from challenge with strain virulent B. melitensis 16 M just as well as the groups immunized with live strains B. melitensis Rev1 or OMVs from rough strain B. melitensis VTRM1 (P < 0.005) (Figure 3).
In addition to protection, in this study, we analyzed the humoral immune response induced in mice by OMVs isolated from smooth and rough Brucella strains. Since the subclass of IgG response is determined by the pattern of cytokines secreted by CD4 helper T cells, we measured the titers of both the IgG1 and IgG2a antibodies produced against OMVs. As has been observed (Figure 4(b)), OMVs from the rough strain were able to induce a higher IgG2a subclass. This IgG2a isotype is important because of the binding of their Fc portion to Fc receptors on the surface of phagocytes that activates a broad spectrum of antimicrobial responses (e.g., phagocytosis, cytokine synthesis, release of inflammatory mediators, and generation of reactive oxidant species) [50].
The information related to the immune response of OMVs from Brucella is limited to observe the antigenicity of these entities in rabbits as reported previously [33]; on the other hand using monoclonal antibodies other authors were able to identify two proteins in OMVs from B. suis [13]. In the same line, there are no reports regarding the immune protection of vaccines based on OMVs against Brucella infection in mouse model.
Many vaccine candidates for human brucellosis involve live attenuated Brucella strains, subunits vaccines, recombinant proteins, and DNA vaccines which have shown to be protective in a mouse model [31].
The biophysical properties of vesicles, as heterogeneous, proteinaceous, amphipathic structures, may allow greater movement through tissues. As a result, vesicles could travel deeper into tissues where resident phagocytes are located [2]. As mentioned previously, the practical application of OMVs as acellular vaccines has been exploited in other pathogens [24, 26, 27]. The advantage to make vaccines based on Brucella OMVs could be that they are able to carry many antigens exposed naturally in the outer membrane and periplasm of Brucella; on the other hand, OMVs are acellular entities that could give an alternative to make safer vaccines instead of using live Brucella vaccines, which have the additional potential complications of replication, virulence, and side effects due to induction of a strong immune response (i.e., fever).
Proteomics approaches have been used to identify the protein components of vesicles in attempts to provide clues to the mechanisms of vesicles production and cargo loading [51]. In this study conventional SDS-PAGE coupled to LC-MS/MS was used to identify the composition of B. melitensis 16 M OMVs. The denatured electrophoretic protein profiles observed from OMVs obtained from rough B. melitensis VTRM1 and smooth B. melitensis 16 M showed no discernable differences (Figure 5). The method selected for OMVs purification was that one reported for Brucella spp by Gamazo et al. in 1989, in which the authors observed a range of sizes in OMVs when Brucella was grown in solid media [33]. We also observed a slightly better yield of OMVs when solid medium was used instead of liquid medium to grow Brucella.
In comparing our results with Gamazo and Moriyon [33], in 1987, they observed an electrophoretic profile that showed two major bands (25 and 30 kDa) and several minor bands (18, 22, and 84 kDa) in the OMVs of both smooth B. melitensis 16 M and a rough strain B. melitensis B115. Our results showed bands similar to those observed by Gamazo and Moriyon, [14] in both smooth and rough OMVs. However, the same research group in 1989 working with OMVs purified from field strains of B. melitensis and B. ovis observed different electrophoretic profiles divided in four groups of proteins: A (25.5–32 kDa), B (21.5–22.5 kDa), C (18–19.5 kDa), and D (13–15.5 kDa); these profiles were different from that reported in 1987. The differences may be due to inherent differences between field strains compared with the reference strain.
After electrophoretic separation, the OMVs proteins from B. melitensis 16 M were excised and digested with trypsin. The trypsin-generated peptide masses, as well as their fragment ions, were analyzed by LC-MS/MS. The resulting peptides sequences were used to query databases led to the identification of 29 unique proteins (Table 1). Our results showed that the outer membrane proteins are the principal components of Brucella OMVs as has been reported previously for other Gram-negative bacteria [23, 41, 52]. However, Lamontagne et al. (2007) found that periplasmic and cytoplasmic proteins as the principal components of OMVs from B. abortus while outer membrane proteins were present as a smaller proportion.
In OMVs from B. melitensis 16 M we identified Omp25 and Omp31 that belong to the major outer membrane protein family [53]. Boigegrain et al. (2004) identified Omp25 and Omp31 in OMVs from B. suis using monoclonal antibodies. Also, Lamontagne et al. (2007) using mass spectrometry were able to identify the Omp31b and Omp25 (Omp3a) in OMVs from B. abortus. The spontaneous release of the OMVs and gentle isolation procedures should minimize cytoplasmic leakage and prevent the contamination that follows cell disruption [51]. Our analyses did not show the presence of inner membrane markers in OMVs composition including NADH-cytochrome C-oxidoreductase or succinate dehydrogenase.
In our study, outer membrane proteins with an OmpA motif were identified; these have been involved in immunostimulatory activities and induce leukocyte migration [54]. The presence of the family Ton B-dependent receptor proteins could be an alternative mechanism for Brucella survival in nutrient-limiting conditions such as found in macrophages [18, 55]. Additionally, Ton B-dependent receptors have been involved in siderophore internalization [18].
Omp16, and the Omp19 are lipoproteins that induce immunological protection very similarly to that elicited by the live vaccine B. abortus S19 with the induction of IFN-γ and CD4+ as well as CD8+ T-cells [29, 48, 56]. Also, the Omp16 shows significant similarity to the peptidoglycan-associated lipoprotein (PALs) of many Gram-negative bacteria [57].
Lipoproteins Omp10, Omp16 and Omp19 were previously identified in OMVs from B. abortus by Western blot [58].
OMVs from B. melitensis 16 M contain Cu-Zn SOD, Dps, and GroES that are part of the antioxidant defense system that protects bacteria from the toxic effects of reactive oxygen intermediates (ROIs) [59, 60]. The Dps protein has been reported in other pathogens (Escherichia coli, Campylobacter jejuni, and Salmonella enterica) to be responsible for resistance to oxidative stress and protecting the DNA against ROIs. The Dps protein has a ferritin-like domain (Table 1) and is thought to nullify the toxic combination of Fe (II) and peroxide [60, 61].
5. Conclusion
In summary, we identified 29 proteins in OMVs released by B. melitensis 16 M, some of them are well-known Brucella immunogens such as SOD, GroES, Omp31, Omp25, Omp19, bp26, and Omp16. Additionally, we determined that rough OMVs both stimulate a stronger innate response, as well as protective immunity against B. melitensis 16 M challenge. Based on these data, the potential of using rough OMVs of Brucella as an acellular vaccine should be considered.
Supplementary Material
Table S1. Results or the real time PCR values for each cytokine and each time point
Table S2. List of all proteins identified in OMVs
Table S3. In silico analysis and annotation of identified proteins based on hypothetical functions
Acknowledgments
This work was funded by CONACYT 76395, and SIP-IPN 20101327 and 20110891 grants. E. D. Avilla-Calderón was supported by CONACYT and PIFI-IPN scholarships. A. Contreras-Rodríguez, A. Lopez-Merino, and E. O. Lopez-Villegas were supported by fellowships from COFAA, SIP-EDI, and SNI-CONACYT. The authors thank Dr. Luz María Ramírez for technical support.
References
- 1.Beveridge TJ. Structures of gram-negative cell walls and their derived membrane vesicles. Journal of Bacteriology. 1999;181(16):4725–4733. doi: 10.1128/jb.181.16.4725-4733.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Kuehn MJ, Kesty NC. Bacterial outer membrane vesicles and the host-pathogen interaction. Genes and Development. 2005;19(22):2645–2655. doi: 10.1101/gad.1299905. [DOI] [PubMed] [Google Scholar]
- 3.Mashburn-Warren LM, Whiteley M. Special delivery: vesicle trafficking in prokaryotes. Molecular Microbiology. 2006;61(4):839–846. doi: 10.1111/j.1365-2958.2006.05272.x. [DOI] [PubMed] [Google Scholar]
- 4.Davis JM, Carvalho HM, Rasmussen SB, O’Brien AD. Cytotoxic necrotizing factor type 1 delivered by outer membrane vesicles of uropathogenic Escherichia coli attenuates polymorphonuclear leukocyte antimicrobial activity and chemotaxis. Infection and Immunity. 2006;74(8):4401–4408. doi: 10.1128/IAI.00637-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Horstman AL, Kuehn MJ. Enterotoxigenic Escherichia coli secretes active heat-labile enterotoxin via outer membrane vesicles. Journal of Biological Chemistry. 2000;275(17):12489–12496. doi: 10.1074/jbc.275.17.12489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Dutta S, Iida KI, Takade A, Meno Y, Nair GB, Yoshida SI. Release of Shiga toxin by membrane vesicles in Shigella dysenteriae serotype 1 strains and in vitro effects of antimicrobials on toxin production and release. Microbiology and Immunology. 2004;48(12):965–969. doi: 10.1111/j.1348-0421.2004.tb03626.x. [DOI] [PubMed] [Google Scholar]
- 7.Kadurugamuwa JL, Beveridge TJ. Membrane vesicles derived from Pseudomonas aeruginosa and Shigella flexneri can be integrated into the surfaces of other Gram-negative bacteria. Microbiology. 1999;145(8):2051–2060. doi: 10.1099/13500872-145-8-2051. [DOI] [PubMed] [Google Scholar]
- 8.Post DMB, Zhang D, Eastvold JS, Teghanemt A, Gibson BW, Weiss JP. Biochemical and functional characterization of membrane blebs purified from Neisseria meningitidis serogroup B. Journal of Biological Chemistry. 2005;280(46):38383–38394. doi: 10.1074/jbc.M508063200. [DOI] [PubMed] [Google Scholar]
- 9.Sharma A, Novak EK, Sojar HT, Swank RT, Kuramitsu HK, Genco RJ. Porphyromonas gingivalis platelet aggregation activity: outer membrane vesicles are potent activators of murine platelets. Oral Microbiology and Immunology. 2000;15(6):393–396. doi: 10.1034/j.1399-302x.2000.150610.x. [DOI] [PubMed] [Google Scholar]
- 10.Fiocca R, Necchi V, Sommi P, et al. Release of Helicobacter pylori vacuolating cytotoxin by both a specific secretion pathway and budding of outer membrane vesicles. Uptake of released toxin and vesicles by gastric epithelium. Journal of Pathology. 1999;188(2):220–226. doi: 10.1002/(SICI)1096-9896(199906)188:2<220::AID-PATH307>3.0.CO;2-C. [DOI] [PubMed] [Google Scholar]
- 11.Hong GE, Kim DG, Park EM, Nam BH, Kim YO, Kong IS. Identification of Vibrio anguillarum outer membrane vesicles related to immunostimulation in the japanese flounder, Paralichthys olivaceus . Bioscience, Biotechnology and Biochemistry. 2009;73(2):437–439. doi: 10.1271/bbb.80580. [DOI] [PubMed] [Google Scholar]
- 12.Alaniz RC, Deatherage BL, Lara JC, Cookson BT. Membrane vesicles are immunogenic facsimiles of Salmonella typhimurium that potently activate dendritic cells, prime B and T cell responses, and stimulate protective immunity in vivo . Journal of Immunology. 2007;179(11):7692–7701. doi: 10.4049/jimmunol.179.11.7692. [DOI] [PubMed] [Google Scholar]
- 13.Boigegrain RA, Salhi I, Alvarez-Martinez MT, et al. Release of periplasmic proteins of Brucella suis upon acidic shock involves the outer membrane protein Omp25. Infection and Immunity. 2004;72(10):5693–5703. doi: 10.1128/IAI.72.10.5693-5703.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Gamazo C, Moriyon I. Release of outer membrane fragments by exponentially growing Brucella melitensis cells. Infection and Immunity. 1987;55(3):609–615. doi: 10.1128/iai.55.3.609-615.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Demuth DR, James D, Kowashi Y, Kato S. Interaction of Actinobacillus actinomycetemcomitans outer membrane vesicles with HL60 cells does not require leukotoxin. Cellular Microbiology. 2003;5(2):111–121. doi: 10.1046/j.1462-5822.2003.00259.x. [DOI] [PubMed] [Google Scholar]
- 16.Núñez-del Arco A, Salas-Téllez E, de la Garza M, Díaz-Aparicio E, Tenorio-Gutiérrez V. Identification of an immunogenic protein of Actinobacillus seminis that is present in microvesicles. Canadian Journal of Veterinary Research. 2006;70(1):43–49. [PMC free article] [PubMed] [Google Scholar]
- 17.Khandelwal P, Banerjee-Bhatnagar N. Insecticidal activity associated with the outer membrane vesicles of Xenorhabdus nematophilus . Applied and Environmental Microbiology. 2003;69(4):2032–2037. doi: 10.1128/AEM.69.4.2032-2037.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Nevot M, Deroncelé V, Messner P, Guinea J, Mercadé E. Characterization of outer membrane vesicles released by the psychrotolerant bacterium Pseudoalteromonas antarctica NF3. Environmental Microbiology. 2006;8(9):1523–1533. doi: 10.1111/j.1462-2920.2006.01043.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Klieve AV, Yokoyama MT, Forster RJ, Ouwerkerk D, Bain PA, Mawhinney EL. Naturally occurring DNA transfer system associated with membrane vesicles in cellulolytic Ruminococcus spp. of ruminal origin. Applied and Environmental Microbiology. 2005;71(8):4248–4253. doi: 10.1128/AEM.71.8.4248-4253.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.McBroom AJ, Kuehn MJ. Release of outer membrane vesicles by Gram-negative bacteria is a novel envelope stress response. Molecular Microbiology. 2007;63(2):545–558. doi: 10.1111/j.1365-2958.2006.05522.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bauman SJ, Kuehn MJ. Purification of outer membrane vesicles from Pseudomonas aeruginosa and their activation of an IL-8 response. Microbes and Infection. 2006;8(9-10):2400–2408. doi: 10.1016/j.micinf.2006.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ismail S, Hampton MB, Keenan JI. Helicobacter pylori outer membrane vesicles modulate proliferation and interleukin-8 production by gastric epithelial cells. Infection and Immunity. 2003;71(10):5670–5675. doi: 10.1128/IAI.71.10.5670-5675.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Ellis TN, Kuehn MJ. Virulence and immunomodulatory roles of bacterial outer membrane vesicles. Microbiology and Molecular Biology Reviews. 2010;74(1):81–94. doi: 10.1128/MMBR.00031-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Holst J, Martin D, Arnold R, et al. Properties and clinical performance of vaccines containing outer membrane vesicles from Neisseria meningitidis . Vaccine. 2009;27(2):B3–B12. doi: 10.1016/j.vaccine.2009.04.071. [DOI] [PubMed] [Google Scholar]
- 25.Katial RK, Brandt BL, Moran EE, Marks S, Agnello V, Zollinger WD. Immunogenicity and safety testing of a group B intranasal meningococcal native outer membrane vesicle vaccine. Infection and Immunity. 2002;70(2):702–707. doi: 10.1128/iai.70.2.702-707.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Roberts R, Moreno G, Bottero D, et al. Outer membrane vesicles as acellular vaccine against pertussis. Vaccine. 2008;26(36):4639–4646. doi: 10.1016/j.vaccine.2008.07.004. [DOI] [PubMed] [Google Scholar]
- 27.Schild S, Nelson EJ, Camilli A. Immunization with Vibrio cholerae outer membrane vesicles induces protective immunity in mice. Infection and Immunity. 2008;76(10):4554–4563. doi: 10.1128/IAI.00532-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Collins BS. Gram-negative outer membrane vesicles in vaccine development. Discovery Medicine. 2011;12(62):7–15. [PubMed] [Google Scholar]
- 29.Pappas G, Akritidis N, Bosilkovski M, Tsianos E. Medical progress Brucellosis. New England Journal of Medicine. 2005;352(22):2325–2367. doi: 10.1056/NEJMra050570. [DOI] [PubMed] [Google Scholar]
- 30.Schurig GG, Sriranganathan N, Corbel MJ. Brucellosis vaccines: past, present and future. Veterinary Microbiology. 2002;90(1–4):479–496. doi: 10.1016/s0378-1135(02)00255-9. [DOI] [PubMed] [Google Scholar]
- 31.Perkins SD, Smither SJ, Atkins HS. Towards a Brucella vaccine for humans. FEMS Microbiology Reviews. 2010;34(3):379–394. doi: 10.1111/j.1574-6976.2010.00211.x. [DOI] [PubMed] [Google Scholar]
- 32.Pasquevich KA, Ibañez AE, Coria LM, et al. An oral vaccine based on U-Omp19 induces protection against B. abortus mucosal challenge by inducing an adaptive IL-17 immune response in mice. PLoS ONE. 2011;6(1) doi: 10.1371/journal.pone.0016203. Article ID e16203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Gamazo C, Winter AJ, Moriyon I, Riezu-Boj JI, Blasco JM, Diaz R. Comparative analyses of proteins extracted by hot saline or released spontaneously into outer membrane blebs from field strains of Brucella ovis and Brucella melitensis . Infection and Immunity. 1989;57(5):1419–1426. doi: 10.1128/iai.57.5.1419-1426.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lamontagne J, Butler H, Chaves-Olarte E, et al. Extensive cell envelope modulation is associated with virulence in Brucella abortus . Journal of Proteome Research. 2007;6(4):1519–1529. doi: 10.1021/pr060636a. [DOI] [PubMed] [Google Scholar]
- 35.Winter AJ, Schurig GG, Boyle SM, et al. Protection of BALB/c mice against homologous and heterologous species of Brucella by rough strain vaccines derived from Brucella melitensis and Brucella suis biovar 4. American Journal of Veterinary Research. 1996;57(5):677–683. [PubMed] [Google Scholar]
- 36.Inaba K, Inaba M, Romani N, et al. Generation of large numbers of dendritic cells from mouse bone marrow cultures supplemented with granulocyte/macrophage colony-stimulating factor. Journal of Experimental Medicine. 1992;176(6):1693–1702. doi: 10.1084/jem.176.6.1693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Surendran N, Hiltbold EM, Heid B, et al. Live Brucella abortus rough vaccine strain RB51 stimulates enhanced innate immune response in vitro compared to rough vaccine strain RB51SOD and virulent smooth strain 2308 in murine bone marrow-derived dendritic cells. Veterinary Microbiology. 2011;147(1-2):75–82. doi: 10.1016/j.vetmic.2010.06.001. [DOI] [PubMed] [Google Scholar]
- 38.Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227(5259):680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- 39.Gardy JL, Laird MR, Chen F, et al. PSORTb v.2.0: expanded prediction of bacterial protein subcellular localization and insights gained from comparative proteome analysis. Bioinformatics. 2005;21(5):617–623. doi: 10.1093/bioinformatics/bti057. [DOI] [PubMed] [Google Scholar]
- 40.Campanella JJ, Bitincka L, Smalley J. MatGAT: an application that generates similarity/identity matrices using protein or DNA sequences. BMC Bioinformatics. 2003;4, article 29 doi: 10.1186/1471-2105-4-29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Bowers PM, Pellegrini M, Thompson MJ, Fierro J, Yeates TO, Eisenberg D. Prolinks: a database of protein functional linkages derived from coevolution. Genome Biology. 2004;5(5):p. R35. doi: 10.1186/gb-2004-5-5-r35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Ashburner M, Ball CA, Blake JA, et al. Gene ontology: tool for the unification of biology. Nature Genetics. 2000;25(1):25–29. doi: 10.1038/75556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Sidhu VK, Vorhölter FJ, Niehaus K, Watt SA. Analysis of outer membrane vesicle associated proteins isolated from the plant pathogenic bacterium Xanthomonas campestris pv. campestris. BMC Microbiology. 2008;8, article 87 doi: 10.1186/1471-2180-8-87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Oliveira SC, de Oliveira FS, Macedo GC, de Almeida LA, Carvalho NB. The role of innate immune receptors in the control of Brucella abortus infection: toll-like receptors and beyond. Microbes and Infection. 2008;10(9):1005–1009. doi: 10.1016/j.micinf.2008.07.005. [DOI] [PubMed] [Google Scholar]
- 45.Dornand J, Gross A, Lafont V, Liautard J, Oliaro J, Liautard JP. The innate immune response against Brucella in humans. Veterinary Microbiology. 2002;90(1–4):383–394. doi: 10.1016/s0378-1135(02)00223-7. [DOI] [PubMed] [Google Scholar]
- 46.High KP, Prasad R, Marion CR, Schurig GG, Boyle SM, Sriranganathan N. Outcome and immune responses after Brucella abortus infection in young adult and aged mice. Biogerontology. 2007;8(5):583–593. doi: 10.1007/s10522-007-9106-6. [DOI] [PubMed] [Google Scholar]
- 47.Billard E, Dornand J, Gross A. Interaction of Brucella suis and Brucella abortus rough strains with human dendritic cells. Infection and Immunity. 2007;75(12):5916–5923. doi: 10.1128/IAI.00931-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Zwerdling A, Delpino MV, Barrionuevo P, et al. Brucella lipoproteins mimic dendritic cell maturation induced by Brucella abortus . Microbes and Infection. 2008;10(12-13):1346–1354. doi: 10.1016/j.micinf.2008.07.035. [DOI] [PubMed] [Google Scholar]
- 49.Pasquevich KA, Samartino CG, Coria LM, et al. The protein moiety of Brucella abortus outer membrane protein 16 is a new bacterial pathogen-associated molecular pattern that activates dendritic cells in vivo, induces a Th1 immune response, and is a promising self-adjuvanting vaccine against systemic and oral acquired brucellosis. Journal of Immunology. 2010;184(9):5200–5212. doi: 10.4049/jimmunol.0902209. [DOI] [PubMed] [Google Scholar]
- 50.Al-Mariri A, Tibor A, Mertens P, et al. Protection of BALB/c mice against Brucella abortus 544 challenge by vaccination with bacterioferritin or P39 recombinant proteins with CpG oligodeoxynucleotides as adjuvant. Infection and Immunity. 2001;69(8):4816–4822. doi: 10.1128/IAI.69.8.4816-4822.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Lee EY, Choi DS, Kim KP, Gho YS. Proteomics in Gram-negative bacterial outer membrane vesicles. Mass Spectrometry Reviews. 2008;27(6):535–555. doi: 10.1002/mas.20175. [DOI] [PubMed] [Google Scholar]
- 52.Lee EY, Joo YB, Gun WP, et al. Global proteomic profiling of native outer membrane vesicles derived from Escherichia coli . Proteomics. 2007;7(17):3143–3153. doi: 10.1002/pmic.200700196. [DOI] [PubMed] [Google Scholar]
- 53.Salhi I, Boigegrain RA, Machold J, Weise C, Cloeckaert A, Rouot B. Characterization of new members of the group 3 outer membrane protein family of Brucella spp . Infection and Immunity. 2003;71(8):4326–4332. doi: 10.1128/IAI.71.8.4326-4332.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Prasadarao NV, Wass CA, Weiser JN, Stins MF, Huang SHE, Kim KS. Outer membrane protein A of Escherichia coli contributes to invasion of brain microvascular endothelial cells. Infection and Immunity. 1996;64(1):146–153. doi: 10.1128/iai.64.1.146-153.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Lamontagne J, Forest A, Marazzo E, et al. Intracellular adaptation of Brucella abortus . Journal of Proteome Research. 2009;8(3):1594–1609. doi: 10.1021/pr800978p. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Pasquevich KA, Estein SM, Samartino CG, et al. Immunization with recombinant Brucella species outer membrane protein Omp16 or Omp19 in adjuvant induces specific CD4+ and CD8+ T cells as well as systemic and oral protection against Brucella abortus infection. Infection and Immunity. 2009;77(1):436–445. doi: 10.1128/IAI.01151-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Tibor A, Decelle B, Letesson JJ. Outer membrane proteins Omp10, Omp16, and Omp19 of Brucella spp. are lipoproteins. Infection and Immunity. 1999;67(9):4960–4962. doi: 10.1128/iai.67.9.4960-4962.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Barquero-Calvo E, Chaves-Olarte E, Weiss DS, et al. Brucella abortus uses a stealthy strategy to avoid activation of the innate immune system during the onset of infection. PLoS ONE. 2007;2(7) doi: 10.1371/journal.pone.0000631. Article ID e631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Gee JM, Valderas MW, Kovach ME, et al. The Brucella abortus Cu,Zn superoxide dismutase is required for optimal resistance to oxidative killing by murine macrophages and wild-type virulence in experimentally infected mice. Infection and Immunity. 2005;73(5):2873–2880. doi: 10.1128/IAI.73.5.2873-2880.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Halsey TA, Vazquez-Torres A, Gravdahl DJ, Fang FC, Libby SJ. The Ferritin-like Dps protein is required for Salmonella enterica serovar typhimurium oxidative stress resistance and virulence. Infection and Immunity. 2004;72(2):1155–1158. doi: 10.1128/IAI.72.2.1155-1158.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Ishikawa T, Mizunoe Y, Kawabata SI, et al. The iron-binding protein Dps confers hydrogen peroxide stress resistance to Campylobacter jejuni . Journal of Bacteriology. 2003;185(3):1010–1017. doi: 10.1128/JB.185.3.1010-1017.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1. Results or the real time PCR values for each cytokine and each time point
Table S2. List of all proteins identified in OMVs
Table S3. In silico analysis and annotation of identified proteins based on hypothetical functions