

Abstract
Microcystins (MCs) are toxins produced by cyanobacteria (blue-green algae), primarily Microcystis aeruginosa, forming water blooms worldwide. When an organism is exposed to environmental perturbations, alterations in normal behavioral patterns occur. Behavioral repertoire represents the consequence of a diversity of physiological and biochemical alterations. In this study, we assessed behavioral patterns and whole-body cortisol levels of adult zebrafish (Danio rerio) exposed to cell culture of the microcystin-producing cyanobacterium M. aeruginosa (MC-LR, strain RST9501). MC-LR exposure (100 μg/L) decreased by 63% the distance traveled and increased threefold the immobility time when compared to the control group. Interestingly, no significant alterations in the number of line crossings were found at the same MC-LR concentration and time of exposure. When animals were exposed to 50 and 100 μg/L, MC-LR promoted a significant increase (around 93%) in the time spent in the bottom portion of the tank, suggesting an anxiogenic effect. The results also showed that none of the MC-LR concentrations tested promoted significant alterations in absolute turn angle, path efficiency, social behavior, or whole-body cortisol level. These findings indicate that behavior is susceptible to MC-LR exposure and provide evidence for a better understanding of the ecological consequences of toxic algal blooms.
1. Introduction
Microcystis aeruginosa is a freshwater cyanobacteria, known producer of a family of toxins termed microcystins (MCs) [1, 2]. MCs are hepatotoxic cyclic heptapeptides released into water during or on senescence of cyanobacterial blooms [3]. The peptide rings of MCs contain five nonprotein amino acids, whereas the two-protein amino acids distinguish MCs from one another. MC-LR contains the amino acids leucine and arginine. MC-LR is one of the most commonly occurring [2, 4] and the most toxic microcystin [5]. The intact cells as well as the toxins released after cellular lysis can be responsible for the toxic effects observed in many organisms, from microalgae [6] to mammals [7] including human [8–10].
Exposure to toxic cyanobacteria or administration of MCs may cause hepatotoxic effects [11–13], oxidative stress [14], kidney damage [15, 16], growth inhibition [17, 18], reproductive injury [19], haematological and biochemical alterations [20–22], apoptosis [23], and even fish death [24].
Alterations in normal behavioral patterns may be the first line of defense when an animal is exposed to an environmental perturbation [25–28]. Additionally, studies have shown important interrelationships between hormones and behavior [29–33]. Thus, alteration in cortisol level may consequently alter normal fish behavior. The effects of MC on fish behavior are still unknown, but some issues have already been addressed. Baganz et al. [34, 35] reported changes in the spontaneous locomotor behavior of zebrafish (Danio rerio) and Leucaspius delineatus after MC-LR exposure, and Cazenave et al. [36] showed changes in swimming activity of Jenynsia multidentata fed with microcystin-RR (MC-RR). In addition, studies using different exposure routes (intraperitoneal injection, oral ingestion, or immersion) have demonstrated that MCs can accumulate in fish tissues, mainly in the liver [21, 36–39], intestine [37, 39–41], gills [42, 43], kidney [37, 39], muscle [40, 41, 44–46], gallbladder [47], blood [40, 41, 48], and brain [43]. Altogether, these findings indicate possible neurotoxic effects of MCs on fish, causing serious risks to the success of fish populations and changes in biodiversity, among other ecological consequences [36].
The zebrafish is rapidly becoming a popular model species in many areas of biological research. Its application includes the fields of developmental biology [49], toxicology [50], neurophysiology, biomedicine, drug discovery [51], human diseases [52–54], pharmacology and behavioral analysis [55–59]. These fish exhibit robust behavioral responses, well-characterized genome, neural and endocrine systems homologous to humans [60–62], and possess all of the “classical” vertebrate neurotransmitters [63, 64]. Additionally, zebrafish are an ideal animal model for laboratory research because they are inexpensive, require low maintenance, and produce abundant offspring [65]. Recently, this fish was also used for proteomic studies on the toxicity of MCs [66, 67].
In order to better understand the neurotoxic effects of MCs on fish and to improve the knowledge of mechanisms underlying the toxicity, the main goal of this study was to assess the effects of MC-LR on zebrafish behavioral parameters and endocrine (whole-body cortisol) response after toxin exposure.
2. Materials and Methods
2.1. Animals
Wild-type adult (<8 months old) zebrafish (Danio rerio) of both sexes were obtained from specialized supplier (Redfish Agroloja, RS, Brazil). Animals were kept in 50 L housing tanks with tap water previously treated with Tetra's AquaSafe (to neutralize chlorine, chloramines, and heavy metals present in the water that could be harmful to fish) and continuously aerated (7.20 mg O2/L) at 26 ± 2°C, under a 14–10 h light/dark photoperiod in a density of up to five animals per liter. Animals were acclimated for at least two weeks before the experiments. They were fed three times a day with TetraMin Tropical Flake fish.
The procedures were previously approved by the Animal Ethics Committee of Pontifical Catholic University of Rio Grande do Sul (PUCRS) under the protocol number 10/00142-CEUA.
2.2. Treatments
The amount of MC-LR in the cell culture of M. aeroginosa (strain RST9501) was detected by a Quantitative Antibody Immunoassay (Elisa) against MC-LR provided by Envirologix (Portland, USA), within a range of detection from 0.05 to 2.5 μg/L MCs. A suitable dilution was applied to the culture sample to provide detection within the range. Zebrafish were distributed in three groups: the first group (controls) was exposed to water containing the culture medium of M. aeroginosa for 24 hours; the second and third groups were exposed to cell culture in a final MC-LR concentration of 50 μg/L and 100 μg/L during 24 hours, respectively. Immediately after the exposure, animals were tested in tank-diving behavioral test and social interaction. After behavioral tests animals were euthanized by decapitation.
The MC-LR concentrations and the time of exposure were chosen based on a previous study using J. multidentata [43]. Besides, such concentrations are commonly encountered in cyanobacterial bloom events [68, 69].
2.3. Tank-Diving Behavioral Test
Behavioral testing took place during the light phase between 10:00 AM and 4:00 PM The animals were individually transferred to a 2.7 L tank (24 cm L × 8 cm W × 20 cm H) with laterals and bottom white covered to avoid any visual disturbances and habituated to the tank for 30 s, as previously described [70]. There was no drug exposure during behavioral experiments. The locomotor activity of the animals was video-recorded using Logitech Quickcam PRO9000 for five minutes after the habituation period and further analyzed using the ANY-Maze recording software (Stoelting Co., Wood Dale, IL, USA). The tank was divided into equal sections with four vertical lines and one horizontal line, and the following behavior patterns were measured: distance traveled (meters), immobility time (seconds), number of crossings, absolute turn angle, path efficiency, and time (seconds) spent in the bottom portion. This task exploits the natural tendency for zebrafish to spend most of the time at the bottom when introduced into a novel environment and then gradually extend the swimming range, over a period of minutes, to include the upper portions of the test tank. A longer time spent in the bottom part of the tank indicates heightened anxiety [71].
2.4. Social Interaction
Zebrafish is a schooling fish that may exhibit preference for its conspecifics under certain circumstances. The social interaction analysis was based on Gerlai [72]. After 24 hours of exposition to 50 or 100 μg/L of MC-LR, fish were placed in groups of five in a small experimental tank (30 cm L × 15 cm H × 10 cm W). On one side of the experimental tank, an empty fish tank was placed, and, on the other side, there was a tank of identical size containing 15 conspecifics. The experimental fish were allowed to acclimate to the experimental tank for a 30 s period, after which their behavior was analyzed. The next 10 s of this test was analyzed as follows. The experimental tank was virtually divided into two equal sections with one vertical line. The time that all five experimental fish spent on the side of the tank closer to the conspecific school was measured using a stopwatch.
2.5. Acute Restraint Stress (ARS) Protocol
The ARS protocol was based on Piato et al. [73]. Following the habituation period, fish were submitted to the ARS protocol. This experiment consisted in keeping each animal enclosed into microcentrifuge plastic tubes of 2 mL with the cap closed and small openings in both ends to allow free water circulation inside the tube and completely avoid fish locomotion. After 90 min of confinement, animals were gently captured and immediately frozen in liquid nitrogen and stored at −80°C until cortisol extraction. Aeration (8 ppm, Labcom Test Camboriú, SC, Brazil) and water temperature (26 ± 2°C) were controlled throughout the test.
2.6. Measurement of Cortisol
The extraction and measurement of whole-body cortisol from zebrafish have been described in detail by Barcellos et al. [74]. Briefly, zebrafish were distributed in four groups: the first group, which consisted of zebrafish exposed to water containing the culture medium of M. aeroginosa for 24 hours, was considered the “negative control”; the second and third groups were exposed to cell culture in a final MC-LR concentration of 50 μg/L and 100 μg/L during 24 hours, respectively; in the fourth group, considered the “positive control,” zebrafish were submitted to the ARS protocol. After, zebrafish were captured and immediately frozen in liquid nitrogen and stored at −80°C until whole-body cortisol extraction. Each zebrafish was weighed, and a pool of three fish was minced and placed into a disposable stomacher bag with 2 mL of phosphate buffered saline (PBS, pH 7.4) for 6 min. The contents were transferred to a 10 mL screw top disposable test tube, and 5 mL of laboratory grade ethyl ether was added. The tube was vortexed for 1 min and centrifuged for 10 min at 3000 rpm. The tube was then immediately frozen at liquid nitrogen, and the unfrozen portion (ethyl ether containing cortisol) was decanted. The ethyl ether was transferred to a new tube and completely evaporated under a gentle stream of nitrogen for 2 h, yielding a lipid extract containing the cortisol. The extract was stored at −20°C until the ELISA was conducted on the samples suspended with 1 mL of PBS buffer. In order to prevent a possible stress response induced by manipulation, the time elapsed between capture and killing was less than 10 s. Whole-body cortisol was measured in duplicate samples of tissue extract with a commercially available high sensitivity salivary cortisol-enzyme immunoassay kit (Salimetrics, USA). The specificity of the test was evaluated by comparing the parallelism between the standard curve and serial dilutions of the tissue extracts in PBS (pH 7.4). The standard curve constructed with the human standards ran parallel to that obtained using serial dilutions of zebrafish tissue extracts. In the linear regression test, high positive correlation (R 2 = 0.9818) was found between the curves. The intra-assay coefficient of variation was 3.33–3.65%.
2.7. Statistical Analysis
Data of the exploratory assessment, social interaction, and cortisol levels were expressed as mean ± SEM and analyzed by one-way ANOVA, followed by Newman-Keuls post hoc test. A significant difference was attributed to P < 0.05. All data were evaluated by SPSS 18.0 for Windows.
3. Results
Distinct parameters of zebrafish swimming activity were evaluated in the 5-min tank-diving behavioral test. MC-LR exposure at 100 μg/L significantly (one-way ANOVA/Newman-Keuls, P < 0.0081, n = 10) decreased the distance traveled (3.7 ± 0.6 meters) in relation to control animals (10.3 ± 1.7 meters) (Figure 1(a)) and significantly (one-way ANOVA/Newman-Keuls, P < 0.039, n = 10) increased the immobility time (137.6 ± 27.6 seconds) when compared to the control group (41.5 ± 17.3 seconds) (Figure 1(b)) whereas MC-LR exposure at 50 μg/L did not alter both parameters. No differences in the number of line crossings, absolute turn angle, and path efficiency were observed in both concentrations tested (Figure 1(c), 1D and 1E, resp.).
Figure 1.
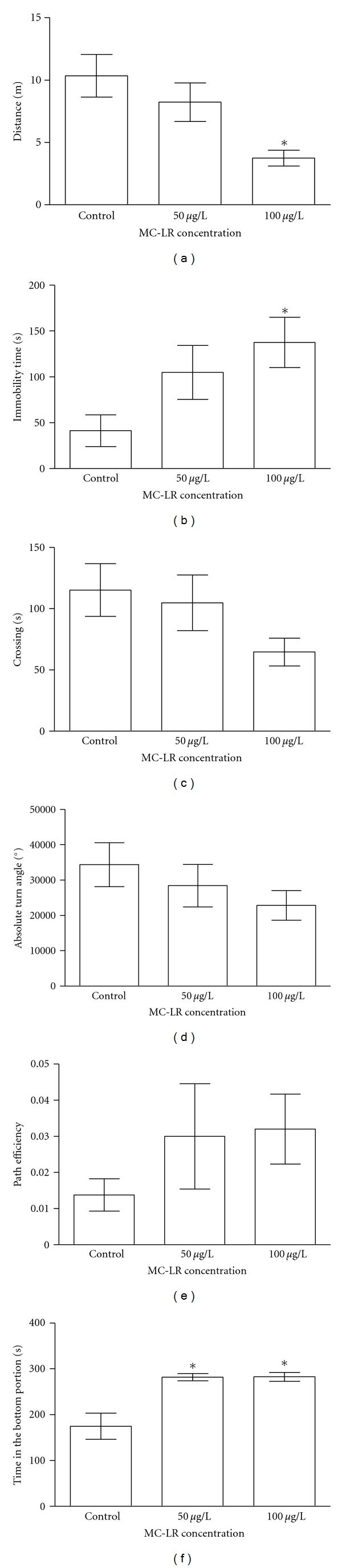
Effect of microcystin-LR exposure on the distance traveled (a), immobility time (b), number of line crossings (c), absolute turn angle (d), path efficiency (e), and time in the bottom portion (f) determined during 5 min of video recording in the tank-diving behavioral test. Data expressed as mean ± SEM. n = 10. One-way ANOVA/Newman-Keuls post hoc test. *: P < 0.05 compared to control group.
Control animals spent 58.4% of time (175.0 ± 28.6) in the bottom portion of the test tank. Animals exposed to 50 and 100 μg/L MC-LR significantly (one-way ANOVA/Newman-Keuls, P < 0.0003, n = 10) increased (93%) the time spent in the bottom portion of the test tank (282.1 ± 7.90 and 282.7 ± 9.7, resp.) when compared with control group (175.0 ± 28.6) (Figure 1(f)).
In relation to social interaction test, the results showed that 50 and 100 μg/L of MC-LR concentrations did not promote any alteration in the animals regarding this behavior (Figure 2).
Figure 2.
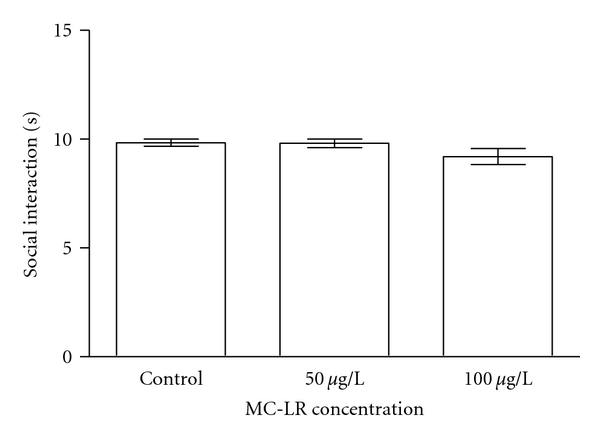
Effect of exposure to microcystin-LR on social interaction. Data expressed as mean ± SEM. n = 10. One-way ANOVA/Newman-Keuls post hoc test.
Levels of whole-body cortisol also were measured. The ARS protocol (positive control) resulted in enhanced whole-body cortisol in relation to control group (one-way ANOVA/Newman-Keuls, P < 0.005, n = 7; 10.7 ± 1.4 and 6.7 ± 0.7, resp.). Zebrafish treated with both concentrations of MC-LR did not present altered levels of cortisol in relation to control group (Figure 3).
Figure 3.
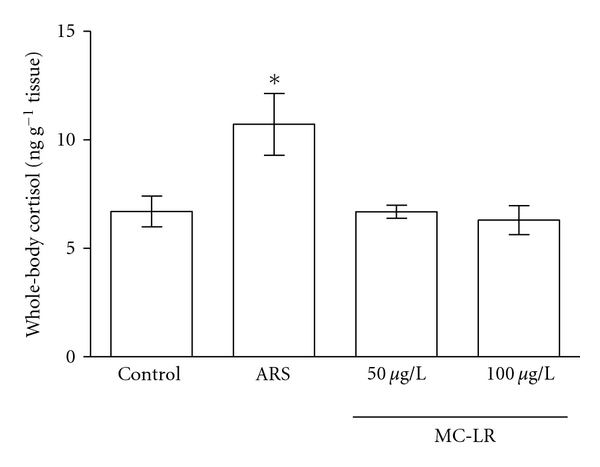
Effect of exposure to microcystin-LR on whole-body cortisol levels. Data expressed as men ± SEM. n = 7. One-way ANOVA/Newman-Keuls post hoc test. *P < 0.05 compared to control group.
4. Discussion
Behavioral alterations reflect how an animal senses and responds to its environment and is the first line of defense when an animal is exposed to an environmental perturbation [28]. Since it was already demonstrated that the effects promoted by cyanobacterial crude extracts on aquatic organisms were either more pronounced or different from those observed using pure toxins [75, 76], we used cell culture of the microcystin-producing cyanobacterium M. aeruginosa (MC-LR) in order to evaluate the effects of MCs on zebrafish behavior.
The toxin concentration and time period of animals' exposure were chosen based on previous studies that showed MCs accumulation in fish tissues [21, 37–48]. The results presented herein demonstrated that 100 μg/L MC-LR decreased the distance traveled and increased the immobility time. However, no significant alterations were found in the number of line crossings with both concentrations. When animals were exposed to 50 and 100 μg/L, MC-LR led to a significant increase in the time spent in the bottom portion. The results also showed that none of the MC-LR concentrations tested promoted significant alterations in the absolute turn angle, path efficiency, or social interaction.
Since behavior links physiological function with ecological processes for a given species, it might provide a useful indicator or biomarker for detecting harmful chemical pollutants [77]. The potential of the zebrafish as a model in neurobehavioral research has emerged only recently. Studies have examined behavior in zebrafish larvae [78–81], as well as their responses to different drugs, such as ethanol [82, 83] and fluoxetine [84]. Studies on adult zebrafish include social behavior [85–87], olfactory-related behaviors [88, 89], anxiety [74], addiction [90–92], sleep [93], learning and memory [94, 95].
There are still only few studies evaluating the effects of MCs on fish behavior. Baganz et al. [34] verified that exposure to MC-LR caused dose-effect-related changes in spontaneous locomotor activity in zebrafish. Whereas exposure to lower concentrations (0.5 and 5 μg/L) caused an increase in daytime mobility, elevated exposures (15 and 50 μg/L) led to significantly increased immobility. The highest exposure (50 μg/L) also reduced the spawning activity and reduced spawning success. In contrast to daytime activities, night-time swimming activity was significantly greater at the higher MC-LR exposures. In another study, Baganz et al. [35] showed changes in the spontaneous locomotor behavior of zebrafish and L. delineatus after exposure to MC-LR in concentrations of 0.5, 5, and 15 μg/L for 17 days and 50 μg/L for six days. During the daytime, the mobility of zebrafish as well as L. delineatus increased significantly by exposure to the lowest concentrations, whereas higher concentrations led to significantly decreased mobility. Influenced by MC-LR, the swimming time of L. delineatus reversed, going from a prominently diurnal activity to a nocturnal one; zebrafish remained active during the daytime. Additionally, Cazenave et al. [36] reported changes in the swimming activity of J. multidentata fed with contaminated food pellets containing MC-RR. Low levels (0.01 μg/g) increased the swimming activity, while the highest dose (1 μg/g) used produced significant changes with respect to control group (only since approximately 20 hours of exposure), when the swimming activity was decreased.
In this sense, our findings demonstrate that MC-LR at the highest concentration (100 μg/L) caused a decrease in the distance traveled and an increase in the immobility time in zebrafish. Interestingly, no significant alterations in the number of line crossings were found at the same MC-LR concentration and time of exposure, despite the tendency to decrease the number of crossings in greater concentration. It is important to emphasize that these findings are similar to the results published previously by Baganz et al. [35]. However, these authors have used purified toxin whereas a cell culture of the microcystin-producing cyanobacterium M. aeruginosa (MC-LR) was used in our experiments. Reduction in swimming capability, resulting in a reduction in the rate of activity, may decrease the ability to gather food and make the fish more vulnerable to predation [96]. Under natural conditions, this reduced overall level of activity will eventually cause disadvantages to the organisms in the ecosystem, and, therefore, influence the biocoenotic structures and functions [34].
Exposure to a novel environment evokes a robust anxiety response in zebrafish [97], as they dive to the bottom (geotaxis) until they feel safe to swim in the upper regions of the tank [58]. Here, MC-LR at 50 and 100 μg/L promoted an increase in the time spent in the bottom portion, suggesting an anxiety behavior.
The zebrafish is a social species and exhibits group preference as well as aggression. Shoaling behavior commences soon after hatching and fish reared in isolation quickly form shoals when placed together [98]. One study has demonstrated that exposure to nonylphenol over a 5-day period decreased shoaling tendency in juvenile rainbow trout (Oncorhynchus mykiss) [99]. Similarly, herbicide-exposed goldfish also showed a decreased aggregation [100]. Locomotor activity, aggressive behavior, and group preference of the male zebrafish and group preference of the females were clearly inhibited when zebrafish were exposed for 60 days to 100 μg/L nonylphenol concentration [101]. For this reason, we evaluated the effect of MC-LR exposure on zebrafish social interaction. However, no significant alteration was found between control and MC-LR-exposed animals.
Studies have shown important interrelationships between stress hormones and behavior [29–33]. An elevated plasma cortisol level is a primary indicator of a stress response in fish [102]. Zebrafish, like humans, employ cortisol as a primary stress response hormone [74]. Considering this, we measured whole-body cortisol in zebrafish to verify if MC could elicit a stress response in treated fish. However, no significant alterations were found in whole-body cortisol levels in animals exposed to both concentrations of MC-LR compared to the control group. Barcellos et al. [74] demonstrated that whole-body cortisol level of zebrafish increases after visual contact with a predator species. Cortisol levels were significantly higher in zebrafish submitted to unpredictable chronic stress (UCS) protocol when compared to control group [103]. Bury et al. [104] reported a significant increase in plasma cortisol levels of the brown trout (Salmo trutta) after 1 h and returned to the control level after 24 h of exposure to lysed toxic Microcystis aeruginosa cells. Crucian carp (Carassius auratus) exposed to sublethal and lethal doses (150 μg/kg and 600 μg/kg, resp.) of Microcystis extracts exhibited a significant acute increase in plasma cortisol levels, which suggested that MC elicited a stress response in treated fish. The profiles of cortisol changes in fish treated with MC appeared to be dose dependent, indicating that fish in the high-dose group experienced greater MC-induced disturbance [105].
In summary, behavioral response of fish may be a promising biomarker of sublethal toxicity and water contamination. Several behavioral endpoint measurements, especially locomotor activity and the time spent in the bottom portion, may provide an effective assessment of MCs in aquatic ecosystem.
Acknowledgments
This work was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (Proc. 472298/2010-0) and DECIT/SCTIEMS through CNPq and Fundação de Amparo à Pesquisa do Estado do Rio Grande do Sul (FAPERGS) (Proc. FAPERGS 10/0036-5-PRONEX). L. W. Kist, J. G. S. da Rosa, and G. Koakoski were recipient of fellowships from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES).
References
- 1.Carmichael WW. Cyanobacteria secondary metabolites—the cyanotoxins. Journal of Applied Bacteriology. 1992;72(6):445–459. doi: 10.1111/j.1365-2672.1992.tb01858.x. [DOI] [PubMed] [Google Scholar]
- 2.Dawson RM. The toxicology of microcystins. Toxicon. 1998;36(7):953–962. doi: 10.1016/s0041-0101(97)00102-5. [DOI] [PubMed] [Google Scholar]
- 3.Malbrouck C, Kestemont P. Effects of microcystins on fish. Environmental Toxicology and Chemistry. 2006;25(1):72–86. doi: 10.1897/05-029r.1. [DOI] [PubMed] [Google Scholar]
- 4.Watanabe MF, Harada KI, Carmichael WW, Fujiki H. Toxic Microcystis. Boca Raton, Fla, USA: CRC Press; 1996. [Google Scholar]
- 5.Rinehart KL, Namikoshi M, Choi BW. Structure and biosynthesis of toxins from blue-green algae (cyanobacteria) Journal of Applied Phycology. 1994;6(2):159–176. [Google Scholar]
- 6.Kearns KD, Hunter MD. Toxin-producing Anabaena flos-aquae induces settling of Chlamydomonas reinhardtii, a competing motile alga. Microbial Ecology. 2001;42(1):80–86. doi: 10.1007/s002480000086. [DOI] [PubMed] [Google Scholar]
- 7.Puschner B, Galey FD, Johnson B, et al. Blue-green algae toxicosis in cattle. Journal of the American Veterinary Medical Association. 1998;213(11):1605–1607. [PubMed] [Google Scholar]
- 8.Ueno Y, Nagata S, Tsutsumi T, et al. Detection of microcystins, a blue-green algal hepatotoxin, in drinking water sampled in Haimen and Fusui, endemic areas of primary liver cancer in China, by highly sensitive immunoassay. Carcinogenesis. 1996;17(6):1317–1321. doi: 10.1093/carcin/17.6.1317. [DOI] [PubMed] [Google Scholar]
- 9.Carmichael WW, Azevedo SM, An JS, et al. Human fatalities form cyanobacteria: chemical and biological evidence for cyanotoxins. Environmental Health Perspectives. 2001;109(7):663–668. doi: 10.1289/ehp.01109663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Chen J, Xie P, Li L, Xu J. First identification of the hepatotoxic microcystins in the serum of a chronically exposed human population together with indication of hepatocellular damage. Toxicological Sciences. 2009;108(1):81–89. doi: 10.1093/toxsci/kfp009. [DOI] [PubMed] [Google Scholar]
- 11.Solter PF, Wollenberg GK, Huang X, Chu FS, Runnegar MT. Prolonged sublethal exposure to protein phosphatase inhibitor microcystin-LR results in multiple dose-dependent hepatotoxic effects. Toxicological Sciences. 1998;44(1):87–96. doi: 10.1006/toxs.1998.2478. [DOI] [PubMed] [Google Scholar]
- 12.Li L, Xie P, Chen J. In vivo studies on toxin accumulation in liver and ultrastructural changes of hepatocytes of the phytoplanktivorous bighead carp i.p.-injected with extracted microcystins. Toxicon. 2005;46(5):533–545. doi: 10.1016/j.toxicon.2005.06.025. [DOI] [PubMed] [Google Scholar]
- 13.Gupta US, Guha S. Microcystin toxicity in a freshwater fish, Heteropneustes fossilis (Bloch) Current Science. 2006;91(9):1261–1271. [Google Scholar]
- 14.Ángeles J, Pichardo S, Prieto AI, et al. Toxic cyanobacterial cells containing microcystins induce oxidative stress in exposed tilapia fish (Oreochromis sp.) under laboratory conditions. Aquatic Toxicology. 2005;72(3):261–271. doi: 10.1016/j.aquatox.2005.01.003. [DOI] [PubMed] [Google Scholar]
- 15.Kotak BG, Semalulu S, Fritz DL, Prepas EE, Hrudey SE, Coppock RW. Hepatic and renal pathology of intraperitoneally administered microcystin-LR in rainbow trout (Oncorhynchus mykiss) Toxicon. 1996;34(5):517–525. doi: 10.1016/0041-0101(96)00009-8. [DOI] [PubMed] [Google Scholar]
- 16.Fischer WJ, Dietrich DR. Pathological and biochemical characterization of microcystin-induced hepatopancreas and kidney damage in carp (Cyprinus carpio) Toxicology and Applied Pharmacology. 2000;164(1):73–81. doi: 10.1006/taap.1999.8861. [DOI] [PubMed] [Google Scholar]
- 17.Bury NR, Eddy FB, Codd GA. The effects of the cyanobacterium Microcystis aeruginosa, the cyanobacterial hepatotoxin microcystin-LR, and ammonia on growth rate and ionic regulation of brown trout. Journal of Fish Biology. 1995;46(6):1042–1054. [Google Scholar]
- 18.Zhao M, Xie S, Zhu X, Yang Y, Gan L, Song L. Effect of inclusion of blue-green algae meal on growth and accumulation of microcystins in gibel carp (Carassius auratus gibelio) Journal of Applied Ichthyology. 2006;22(1):72–78. [Google Scholar]
- 19.Ding XS, Li XY, Duan HY, Chung IK, Lee JA. Toxic effects of Microcystis cell extracts on the reproductive system of male mice. Toxicon. 2006;48(8):973–979. doi: 10.1016/j.toxicon.2006.07.039. [DOI] [PubMed] [Google Scholar]
- 20.Vajcova V, Navrátil S, Palíková M. The effect of intraperitoneally applied pure microcystin LR on haematological, biochemical and morphological indices of silver carp (Hypophthalmichthys molitrix Val.) Acta Veterinaria. 1998;67(4):281–287. [Google Scholar]
- 21.Malbrouck C, Trausch G, Devos P, Kestemont P. Hepatic accumulation and effects of microcystin-LR on juvenile goldfish Carassius auratus L. Comparative Biochemistry and Physiology: C. 2003;135(1):39–48. doi: 10.1016/s1532-0456(03)00047-4. [DOI] [PubMed] [Google Scholar]
- 22.Zhang X, Xie P, Li D, Shi Z. Hematological and plasma biochemical responses of crucian carp (Carassius auratus) to intraperitoneal injection of extracted microcystins with the possible mechanisms of anemia. Toxicon. 2007;49(8):1150–1157. doi: 10.1016/j.toxicon.2007.02.009. [DOI] [PubMed] [Google Scholar]
- 23.McDermott CM, Nho CW, Howard W, Holton B. The cyanobacterial toxin, microcystin-LR, can induce apoptosis in a variety of cell types. Toxicon. 1998;36(12):1981–1996. doi: 10.1016/s0041-0101(98)00128-7. [DOI] [PubMed] [Google Scholar]
- 24.Lindholm T, Öhman P, Kurki-Helasmo K, Kincaid B, Meriluoto J. Toxic algae and fish mortality in a brackish-water lake in Aland, SW Finland. Hydrobiologia. 1999;397:109–120. [Google Scholar]
- 25.Scherer E. Behavioral responses as indicators of environmental alterations, approaches, results, developments. Journal of Applied Ichthyology. 1992;8(1–4):122–131. [Google Scholar]
- 26.Paul EA, Simonin HA. Effects of Naled, synergized,and non-synergized resmethrin on the swimming performance of young trout. Bulletin of Environmental Contamination and Toxicology. 1996;57(3):495–502. doi: 10.1007/s001289900217. [DOI] [PubMed] [Google Scholar]
- 27.Campbell HA, Handy RD, Sims DW. Increased metabolic cost of swimming and consequent alterations to circadian activity in rainbow trout (Oncorhynchus mykiss) exposed to dietary copper. Canadian Journal of Fisheries and Aquatic Sciences. 2002;59(5):768–777. [Google Scholar]
- 28.Begout Anras ML, Lagardère JP. Measuring cultured fish swimming behaviour: first results on rainbow trout using acoustic telemetry in tanks. Aquaculture. 2004;240(1–4):175–186. [Google Scholar]
- 29.Oliveira RF, Almada VC, Canario AVM. Social modulation of sex steroid concentrations in the urine of male cichlid fish Oreochromis mossambicus . Hormones and Behavior. 1996;30(1):2–12. doi: 10.1006/hbeh.1996.0002. [DOI] [PubMed] [Google Scholar]
- 30.Contreras-Sánchez WM, Schreck CB, Fitzpatrick MS, Pereira CB. Effects of stress on the reproductive performance of rainbow trout (Oncorhynchus mykiss) Biology of Reproduction. 1998;58(2):439–447. doi: 10.1095/biolreprod58.2.439. [DOI] [PubMed] [Google Scholar]
- 31.Pottinger TG, Carrick TR. Stress responsiveness affects dominant-subordinate relationships in rainbow trout. Hormones and Behavior. 2001;40(3):419–427. doi: 10.1006/hbeh.2001.1707. [DOI] [PubMed] [Google Scholar]
- 32.Sloman KA, Metcalfe NB, Taylor AC, Gilmour KM. Plasma cortisol concentrations before and after social stress in rainbow trout and brown trout. Physiological and Biochemical Zoology. 2001;74(3):383–389. doi: 10.1086/320426. [DOI] [PubMed] [Google Scholar]
- 33.Øverli Ø, Pottinger TG, Carrick TR, Øverli E, Winberg S. Differences in behaviour between rainbow trout selected for high- and low-stress responsiveness. Journal of Experimental Biology. 2002;205(3):391–395. doi: 10.1242/jeb.205.3.391. [DOI] [PubMed] [Google Scholar]
- 34.Baganz D, Staaks G, Steinberg C. Impact of the cyanobacteria toxin, microcystin-LR on behaviour of zebrafish, Danio rerio . Water Research. 1998;32(3):948–952. [Google Scholar]
- 35.Baganz D, Staaks G, Pflugmacher S, Steinberg CEW. Comparative study of microcystin-LR-induced behavioral changes of two fish species, Danio rerio and Leucaspius delineatus . Environmental Toxicology. 2004;19(6):564–570. doi: 10.1002/tox.20063. [DOI] [PubMed] [Google Scholar]
- 36.Cazenave J, Nores ML, Miceli M, Díaz MP, Wunderlin DA, Bistoni MA. Changes in the swimming activity and the glutathione S-transferase activity of Jenynsia multidentata fed with microcystin-RR. Water Research. 2008;42(4-5):1299–1307. doi: 10.1016/j.watres.2007.09.025. [DOI] [PubMed] [Google Scholar]
- 37.Williams DE, Kent ML, Andersen RJ, Klix H, Holmes CFB. Tissue distribution and clearance of tritium-labeled dihydromicrocystin-LR epimers administered to Atlantic salmon via intraperitoneal injection. Toxicon. 1995;33(2):125–131. doi: 10.1016/0041-0101(94)00154-z. [DOI] [PubMed] [Google Scholar]
- 38.Sipiä VO, Kankaanpää HT, Flinkman J, Lahti K, Meriluoto JAO. Time-dependent accumulation of cyanobacterial hepatotoxins in flounders (Platichthys flesus) and mussels (Mytilus edulis) from the Northern Baltic Sea. Environmental Toxicology. 2001;16(4):330–336. doi: 10.1002/tox.1040. [DOI] [PubMed] [Google Scholar]
- 39.Mohamed ZA, Carmichael WW, Hussein AA. Estimation of microcystins in the freshwater fish Oreochromis niloticus in an Egyptian fish farm containing a Microcystis bloom. Environmental Toxicology. 2003;18(2):137–141. doi: 10.1002/tox.10111. [DOI] [PubMed] [Google Scholar]
- 40.Xie L, Xie P, Ozawa K, Honma T, Yokoyama A, Park HD. Dynamics of microcystins-LR and -RR in the phytoplanktivorous silver carp in a sub-chronic toxicity experiment. Environmental Pollution. 2004;127(3):431–439. doi: 10.1016/j.envpol.2003.08.011. [DOI] [PubMed] [Google Scholar]
- 41.Xie L, Xie P, Guo L, Li L, Miyabara Y, Park HD. Organ distribution and bioaccumulation of microcystins in freshwater fish at different trophic levels from the eutrophic Lake Chaohu, China. Environmental Toxicology. 2005;20(3):293–300. doi: 10.1002/tox.20120. [DOI] [PubMed] [Google Scholar]
- 42.Carbis CR, Simons JA, Grant P, Mitchell GF, Anderson JW, McCauley I. A study of feral carp, Cyprinus carpio L., exposed to Microcystis aeruginosa at Lake Mokoan, Australia, and possible implications for fish health. Journal of Fish Diseases. 1997;20(2):81–91. [Google Scholar]
- 43.Cazenave J, Wunderlin DA, Bistoni MDLA, et al. Uptake, tissue distribution and accumulation of microcystin-RR in Corydoras paleatus, Jenynsia multidentata and Odontesthes bonariensis: a field and laboratory study. Aquatic Toxicology. 2005;75(2):178–190. doi: 10.1016/j.aquatox.2005.08.002. [DOI] [PubMed] [Google Scholar]
- 44.Bury NR, Newlands AD, Eddy FB, Codd GA. In vivo and in vitro intestinal transport of 3H-microcystin-LR, a cyanobacterial toxin, in rainbow trout (Oncorhynchus mykiss) Aquatic Toxicology. 1998;42(2):139–148. [Google Scholar]
- 45.Magalhães VF, Moraes Soares R, Azevedo SMFO. Microcystin contamination in fish from the Jacarepaguá Lagoon (Rio de Janeiro, Brazil): ecological implication and human health risk. Toxicon. 2001;39(7):1077–1085. doi: 10.1016/s0041-0101(00)00251-8. [DOI] [PubMed] [Google Scholar]
- 46.Magalhães VF, Marinho MM, Domingos P, et al. Microcystins (cyanobacteria hepatotoxins) bioaccumulation in fish and crustaceans from Sepetiba Bay (Brasil, RJ) Toxicon. 2003;42(3):289–295. doi: 10.1016/s0041-0101(03)00144-2. [DOI] [PubMed] [Google Scholar]
- 47.Sahin A, Tencalla FG, Dietrich DR, Naegeli H. Biliary excretion of biochemically active cyanobacteria (blue-green algae) hepatotoxins in fish. Toxicology. 1996;106(1–3):123–130. doi: 10.1016/0300-483x(95)03173-d. [DOI] [PubMed] [Google Scholar]
- 48.Tencalla F, Dietrich D. Biochemical characterization of microcystin toxicity in rainbow trout (Oncorhynchus mykiss) Toxicon. 1997;35(4):583–595. doi: 10.1016/s0041-0101(96)00153-5. [DOI] [PubMed] [Google Scholar]
- 49.Zon LI, Peterson RT. In vivo drug discovery in the zebrafish. Nature Reviews Drug Discovery. 2005;4(1):35–44. doi: 10.1038/nrd1606. [DOI] [PubMed] [Google Scholar]
- 50.Spitsbergen JM, Kent ML. The state of the art of the zebrafish model for toxicology and toxicologic pathology research—advantages and current limitations. Toxicologic Pathology. 2003;31:62–87. doi: 10.1080/01926230390174959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Rubinstein AL. Zebrafish: from disease modeling to drug discovery. Current Opinion in Drug Discovery and Development. 2003;6(2):218–223. [PubMed] [Google Scholar]
- 52.Dooley K, Zon LI. Zebrafish: a model system for the study of human disease. Current Opinion in Genetics and Development. 2000;10(3):252–256. doi: 10.1016/s0959-437x(00)00074-5. [DOI] [PubMed] [Google Scholar]
- 53.Lieschke GJ, Currie PD. Animal models of human disease: zebrafish swim into view. Nature Reviews Genetics. 2007;8(5):353–367. doi: 10.1038/nrg2091. [DOI] [PubMed] [Google Scholar]
- 54.Best JD, Alderton WK. Zebrafish: an In vivo model for the study of neurological diseases. Neuropsychiatric Disease and Treatment. 2008;4(3):567–576. doi: 10.2147/ndt.s2056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Egan RJ, Bergner CL, Hart PC, et al. Understanding behavioral and physiological phenotypes of stress and anxiety in zebrafish. Behavioural Brain Research. 2009;205(1):38–44. doi: 10.1016/j.bbr.2009.06.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Cachat J, Canavello P, Elegante M, et al. Modeling withdrawal syndrome in zebrafish. Behavioural Brain Research. 2010;208(2):371–376. doi: 10.1016/j.bbr.2009.12.004. [DOI] [PubMed] [Google Scholar]
- 57.Maximino C, de Brito TM, Dias CA, Gouveia A, Morato S. Scototaxis as anxietylike behavior in fish. Nature Protocols. 2010;5(2):209–216. doi: 10.1038/nprot.2009.225. [DOI] [PubMed] [Google Scholar]
- 58.Stewart A, Kadri F, DiLeo J. The developing utility of zebrafish in modeling neurobehavioral disorders. International Journal of Comparative Psychology. 2010;23:104–121. [Google Scholar]
- 59.Wong K, Elegante M, Bartels B, et al. Analyzing habituation responses to novelty in zebrafish (Danio rerio) Behavioural Brain Research. 2010;208(2):450–457. doi: 10.1016/j.bbr.2009.12.023. [DOI] [PubMed] [Google Scholar]
- 60.Kaslin J, Panula P. Comparative anatomy of the histaminergic and other aminergic systems in zebrafish (Danio rerio) Journal of Comparative Neurology. 2001;440(4):342–377. doi: 10.1002/cne.1390. [DOI] [PubMed] [Google Scholar]
- 61.Shin JT, Fishman MC. From zebrafish to human: modular medical models. Annual Review of Genomics and Human Genetics. 2002;3:311–340. doi: 10.1146/annurev.genom.3.031402.131506. [DOI] [PubMed] [Google Scholar]
- 62.Gabriel JP, Mahmood R, Kyriakatos A, et al. Serotonergic modulation of locomotion in zebrafish: endogenous release and synaptic mechanisms. Journal of Neuroscience. 2009;29(33):10387–10395. doi: 10.1523/JNEUROSCI.1978-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Mueller T, Vernier P, Wullimann MF. The adult central nervous cholinergic system of a neurogenetic model animal, the zebrafish Danio rerio . Brain Research. 2004;1011(2):156–169. doi: 10.1016/j.brainres.2004.02.073. [DOI] [PubMed] [Google Scholar]
- 64.Panula P, Sallinen V, Sundvik M, et al. Modulatory neurotransmitter systems and behavior: towards zebrafish models of neurodegenerative diseases. Zebrafish. 2006;3(2):235–247. doi: 10.1089/zeb.2006.3.235. [DOI] [PubMed] [Google Scholar]
- 65.Gerlai R, Lee V, Blaser R. Effects of acute and chronic ethanol exposure on the behavior of adult zebrafish (Danio rerio) Pharmacology Biochemistry and Behavior. 2006;85(4):752–761. doi: 10.1016/j.pbb.2006.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Li G, Chen J, Xie P, Jiang Y, Wu L, Zhang X. Protein expression profiling in the zebrafish (Danio rerio) embryos exposed to the microcystin-LR. Proteomics. 2011;11(10):2003–2018. doi: 10.1002/pmic.201000442. [DOI] [PubMed] [Google Scholar]
- 67.Zhao Y, Xiong Q, Xie P. Analysis of microRNA expression in embryonic developmental toxicity induced by MC-RR. PLoS One. 2011;6(7) doi: 10.1371/journal.pone.0022676. Article ID e22676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Oberholster PJ, Myburgh JG, Govender D, Bengis R, Botha AM. Identification of toxigenic Microcystis strains after incidents of wild animal mortalities in the Kruger National Park, South Africa. Ecotoxicology and Environmental Safety. 2009;72(4):1177–1182. doi: 10.1016/j.ecoenv.2008.12.014. [DOI] [PubMed] [Google Scholar]
- 69.Backer LC, McNeel SV, Barber T, et al. Recreational exposure to microcystins during algal blooms in two California lakes. Toxicon. 2010;55(5):909–921. doi: 10.1016/j.toxicon.2009.07.006. [DOI] [PubMed] [Google Scholar]
- 70.Gerlai R, Lahav M, Guo S, Rosenthal A. Drinks like a fish: zebra fish (Danio rerio) as a behavior genetic model to study alcohol effects. Pharmacology Biochemistry and Behavior. 2000;67(4):773–782. doi: 10.1016/s0091-3057(00)00422-6. [DOI] [PubMed] [Google Scholar]
- 71.Levin ED, Bencan Z, Cerutti DT. Anxiolytic effects of nicotine in zebrafish. Physiology & Behavior. 2007;90(1):54–58. doi: 10.1016/j.physbeh.2006.08.026. [DOI] [PubMed] [Google Scholar]
- 72.Gerlai R. Zebra fish: an uncharted behavior genetic model. Behavior Genetics. 2003;33(5):461–468. doi: 10.1023/a:1025762314250. [DOI] [PubMed] [Google Scholar]
- 73.Piato AL, Rosemberg DB, Capiotti KM, et al. Acute restraint stress in zebrafish: behavioral parameters and purinergic signaling. Neurochemical Research. 2011;36(10):1876–1886. doi: 10.1007/s11064-011-0509-z. [DOI] [PubMed] [Google Scholar]
- 74.Barcellos LJG, Ritter F, Kreutz LC, et al. Whole-body cortisol increases after direct and visual contact with a predator in zebrafish Danio rerio . Aquaculture. 2007;272(1–4):774–778. [Google Scholar]
- 75.Oberemm A, Becker J, Codd GA, Steinberg C. Effects of cyanobacterial toxins and aqueous crude extracts of cyanobacteria on the development of fish and amphibians. Environmental Toxicology. 1999;14(1):77–88. [Google Scholar]
- 76.Pietsch C, Wiegand C, Amé MV, Nicklisch A, Wunderlin D, Pflugmacher S. The effects of a cyanobacterial crude extract on different aquatic organisms: evidence for cyanobacterial toxin modulating factors. Environmental Toxicology. 2001;16(6):535–542. doi: 10.1002/tox.10014. [DOI] [PubMed] [Google Scholar]
- 77.Zala SM, Penn DJ. Abnormal behaviours induced by chemical pollution: a review of the evidence and new challenges. Animal Behaviour. 2004;68(4):649–664. [Google Scholar]
- 78.Saint-Amant L, Drapeau P. Time course of the development of motor behaviors in the zebrafish embryo. Journal of Neurobiology. 1998;37(4):622–632. doi: 10.1002/(sici)1097-4695(199812)37:4<622::aid-neu10>3.0.co;2-s. [DOI] [PubMed] [Google Scholar]
- 79.Saint-Amant L, Drapeau P. Synchronization of an embryonic network of identified spinal interneurons solely by electrical coupling. Neuron. 2001;31(6):1035–1046. doi: 10.1016/s0896-6273(01)00416-0. [DOI] [PubMed] [Google Scholar]
- 80.Borla MA, Palecek B, Budick S, O’Malley DM. Prey capture by larval zebrafish: evidence for fine axial motor control. Brain, Behavior and Evolution. 2002;60(4):207–229. doi: 10.1159/000066699. [DOI] [PubMed] [Google Scholar]
- 81.Watkins J, Miklosi A, Andrew RJ. Early asymmetries in the behaviour of zebrafish larvae. Behavioural Brain Research. 2004;151(1-2):177–183. doi: 10.1016/j.bbr.2003.08.012. [DOI] [PubMed] [Google Scholar]
- 82.Loucks E, Carvan MJ. Strain-dependent effects of developmental ethanol exposure in zebrafish. Neurotoxicology and Teratology. 2004;26(6):745–755. doi: 10.1016/j.ntt.2004.06.017. [DOI] [PubMed] [Google Scholar]
- 83.Damodaran S, Dlugos CA, Wood TD, Rabin RA. Effects of chronic ethanol administration on brain protein levels: a proteomic investigation using 2-D DIGE system. European Journal of Pharmacology. 2006;547(1–3):75–82. doi: 10.1016/j.ejphar.2006.08.005. [DOI] [PubMed] [Google Scholar]
- 84.Airhart MJ, Lee DH, Wilson TD, Miller BE, Miller MN, Skalko RG. Movement disorders and neurochemical changes in zebrafish larvae after bath exposure to fluoxetine (PROZAC) Neurotoxicology and Teratology. 2007;29(6):652–664. doi: 10.1016/j.ntt.2007.07.005. [DOI] [PubMed] [Google Scholar]
- 85.Pyron M. Female preferences and male-male interactions in zebrafish (Danio rerio) Canadian Journal of Zoology. 2003;81(1):122–125. [Google Scholar]
- 86.Engeszer RE, Ryan MJ, Parichy DM. Learned social preference in zebrafish. Current Biology. 2004;14(10):881–884. doi: 10.1016/j.cub.2004.04.042. [DOI] [PubMed] [Google Scholar]
- 87.Bass SLS, Gerlai R. Zebrafish (Danio rerio) responds differentially to stimulus fish: the effects of sympatric and allopatric predators and harmless fish. Behavioural Brain Research. 2008;186(1):107–117. doi: 10.1016/j.bbr.2007.07.037. [DOI] [PubMed] [Google Scholar]
- 88.Mann KD, Turnell ER, Atema J, Gerlach G. Kin recognition in juvenile zebrafish (Danio rerio) based on olfactory cues. The Biological Bulletin. 2003;205(2):224–225. doi: 10.2307/1543264. [DOI] [PubMed] [Google Scholar]
- 89.Vitebsky A, Reyes R, Sanderson MJ, Michel WC, Whitlock KE. Isolation and characterization of the laure olfactory behavioral mutant in the zebrafish, Danio rerio . Developmental Dynamics. 2005;234(1):229–242. doi: 10.1002/dvdy.20530. [DOI] [PubMed] [Google Scholar]
- 90.Darland T, Dowling JE. Behavioral screening for cocaine sensitivity in mutagenized zebrafish. Proceedings of the National Academy of Sciences of the United States of America. 2001;98(20):11691–11696. doi: 10.1073/pnas.191380698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Ninkovic J, Bally-Cuif L. The zebrafish as a model system for assessing the reinforcing properties of drugs of abuse. Methods. 2006;39(3):262–274. doi: 10.1016/j.ymeth.2005.12.007. [DOI] [PubMed] [Google Scholar]
- 92.Kily LJ, Cowe YC, Hussain O, et al. Gene expression changes in a zebrafish model of drug dependency suggest conservation of neuro-adaptation pathways. Journal of Experimental Biology. 2008;211(10):1623–1634. doi: 10.1242/jeb.014399. [DOI] [PubMed] [Google Scholar]
- 93.Cirelli C, Tononi G. Differential expression of plasticity-related genes in waking and sleep and their regulation by the noradrenergic system. Journal of Neuroscience. 2000;20(24):9187–9194. doi: 10.1523/JNEUROSCI.20-24-09187.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Williams FE, White D, Messer WS. A simple spatial alternation task for assessing memory function in zebrafish. Behavioural Processes. 2002;58(3):125–132. doi: 10.1016/s0376-6357(02)00025-6. [DOI] [PubMed] [Google Scholar]
- 95.Colwill RM, Raymond MP, Ferreira L, Escudero H. Visual discrimination learning in zebrafish (Danio rerio) Behavioural Processes. 2005;70(1):19–31. doi: 10.1016/j.beproc.2005.03.001. [DOI] [PubMed] [Google Scholar]
- 96.Plaut I. Effects of fin size on swimming performance, swimming behaviour and routine activity of zebrafish Danio rerio . Journal of Experimental Biology. 2000;203(4):813–820. doi: 10.1242/jeb.203.4.813. [DOI] [PubMed] [Google Scholar]
- 97.Blaser R, Gerlai R. Behavioral phenotyping in zebrafish: comparison of three behavioral quantification methods. Behavior Research Methods. 2006;38(3):456–469. doi: 10.3758/bf03192800. [DOI] [PubMed] [Google Scholar]
- 98.Spence R, Gerlach G, Lawrence C, Smith C. The behavior and ecology of the zebrafish, Danio rerio . Biological Reviews of the Cambridge Philosofical Society. 2008;83(1):13–34. doi: 10.1111/j.1469-185X.2007.00030.x. [DOI] [PubMed] [Google Scholar]
- 99.Ward AJW, Duff AJ, Currie S. The effects of the endocrine disrupter 4-nonylphenol on the behaviour of juvenile rainbow trout (Oncorhynchus mykiss) Canadian Journal of Fisheries and Aquatic Sciences. 2006;63(2):377–382. [Google Scholar]
- 100.Saglio P, Trijasse S. Behavioral responses to atrazine and diuron in goldfish. Archives of Environmental Contamination and Toxicology. 1998;35(3):484–491. doi: 10.1007/s002449900406. [DOI] [PubMed] [Google Scholar]
- 101.Xia J, Niu C, Pei X. Effects of chronic exposure to nonylphenol on locomotor activity and social behavior in zebrafish (Danio rerio) Journal of Environmental Sciences. 2010;22(9):1435–1440. doi: 10.1016/s1001-0742(09)60272-2. [DOI] [PubMed] [Google Scholar]
- 102.Barton BA. Stress in fishes: a diversity of responses with particular reference to changes in circulating corticosteroids. Integrative and Comparative Biology. 2002;42(3):517–525. doi: 10.1093/icb/42.3.517. [DOI] [PubMed] [Google Scholar]
- 103.Piato AL, Capiotti KM, Tamborski AR, et al. Unpredictable chronic stress model in zebrafish (Danio rerio): behavioral and physiological responses. Progress in Neuro-Psychopharmacology & Biological Psychiatry. 2011;35(2):561–567. doi: 10.1016/j.pnpbp.2010.12.018. [DOI] [PubMed] [Google Scholar]
- 104.Bury NR, Eddy FB, Codd GA. The stress responses of brown trout, Salmo truffa L, to the cyanobacterium, Microcystis aeruginosa . Environmental Toxicology and Water Quality. 1996;11(3):187–193. [Google Scholar]
- 105.Li D, Xie P, Zhang X. Changes in plasma thyroid hormones and cortisol levels in crucian carp (Carassius auratus) exposed to the extracted microcystins. Chemosphere. 2008;74(1):13–18. doi: 10.1016/j.chemosphere.2008.09.065. [DOI] [PubMed] [Google Scholar]