

Abstract
In contexts of cultural conflict, people delegitimize the other group's perspective and lose compassion for the other group's suffering. These psychological biases have been empirically characterized in intergroup settings, but rarely in groups involved in active conflict. Similarly, the basic brain networks involved in recognizing others' narratives and misfortunes have been identified, but how these brain networks are modulated by intergroup conflict is largely untested. In the present study, we examined behavioural and neural responses in Arab, Israeli and South American participants while they considered the pain and suffering of individuals from each group. Arabs and Israelis reported feeling significantly less compassion for each other's pain and suffering (the ‘conflict outgroup’), but did not show an ingroup bias relative to South Americans (the ‘distant outgroup’). In contrast, the brain regions that respond to others' tragedies showed an ingroup bias relative to the distant outgroup but not the conflict outgroup, particularly for descriptions of emotional suffering. Over all, neural responses to conflict group members were qualitatively different from neural responses to distant group members. This is the first neuroimaging study to examine brain responses to others' suffering across both distant and conflict groups, and provides a first step towards building a foundation for the biological basis of conflict.
Keywords: functional magnetic resonance imaging, conflict, social cognition, empathy, pain, suffering
1. Introduction
Violent and apparently intractable conflicts divide many neighbouring cultures: Indians and Pakistanis, Bosnians and Serbs, Israelis and Palestinians. Members of these groups live side by side, sharing geography and history, but are divided by conflicts over resources, power, history and political institutions. Once two cultures are divided, psychological factors perpetuate the conflict [1–3]. Because at least some of these psychological drivers of conflict are stable and predictable, across geographical, historical and political contexts, they offer plausible, and promising, first targets for helping us to construct a biology of cultural conflict.
When two groups are in conflict, psychological biases operate to drive the groups apart, and prevent reconciliation. For example, members of groups in conflict come to perceive the ‘enemy’ as unreasonable and close-minded, self-interested and aggressive, and therefore undeserving of cooperative gestures [4–6]. The ideas that the other group ‘only understands the language of violence,’ and that outcomes must be zero-sum, as in ‘their loss is our gain,’ can lead to tolerating, endorsing or even perpetrating violence against members of the other group that would be unthinkable outside the context of conflict [7]. That is, if empathy includes the tendency to feel saddened by another person's misfortunes, and to feel motivated to alleviate their suffering, then cultural conflict often causes empathy to fail.
One striking possibility is that these psychological drivers of cultural conflicts could soon be traced back to different patterns of activity in specific brain regions. Over the past 10 years, the new field of social cognitive neuroscience has identified neural correlates of multiple social processes potentially relevant to intergroup conflict, including reacting to witnessing others in pain, considering their thoughts and beliefs, and reasoning about their narratives [8–15]. The emerging neuroscience of people thinking about people may thus provide a foundation for measuring the neural effects of cultural conflict.
Two ‘networks’ of brain regions may be particularly important for understanding the neural sources of failures of empathy, in cultural conflicts. First, there is a group of brain regions implicated in processing physically painful experiences—both one's own, and those of other people. Directly experiencing physical pain (PP) leads to brain activity in the so-called ‘pain matrix’, which includes regions in middle cingulate cortex (CC) extending dorsally into medial supplementary motor area, and bilateral anterior insula, as well as subcortical structures [16–18]. When observers witness other people experience PP, some of the same brain regions are activated: most reliably, the CC and insula [15,19,20]. The amount of activity in these regions is correlated with how much pain observers attribute to the victim [21]. Because these neural regions are common to first- and third-person experiences of pain, they have been hypothesized to serve as a ‘functional bridge’ between the observer and the victim [22] (but see [23]); linking other's people's painful experiences to one's own could lead observers to identify with the victim, feel for their misfortune and be motivated to alleviate it.
Second, there is a group of brain regions implicated in thinking about people's thoughts, desires and feelings (a cognitive process often called having a ‘theory of mind’ or ‘mentalizing’). These ‘mentalizing’ regions include the bilateral temporo-parietal junction (TPJ), medial prefrontal cortex (MPFC) and medial precuneus (PC). These brain regions are robustly activated when people consider what someone was thinking, in order to explain or predict or make moral judgements about their actions [9,24,25]. Although all of these regions are activated while reasoning about someone's vivid emotional experience, there is some evidence of a division of cognitive labour: regions in MPFC appear to be especially implicated in reasoning about others' affective states, while the lateral TPJ regions are more associated with reasoning about others' cognitive states [9,26–28].
Cultural conflict could modulate activity in either one or both of these groups of brain regions. In violent cultural conflicts, individuals from both groups suffer both PP, from physical injuries, and emotional losses, including losses of friends and loved ones, of freedoms and aspirations, and of dignity and security. Recognition of these individual prices of conflict would rely on activity in both ‘pain matrix’ and ‘mentalizing’ brain regions.
How could these brain networks be affected by cultural conflict? One simple possibility is that the pain and suffering of individuals from the opposing group elicits less activity than an ingroup member. On this view, we would predict reduced activity in both brain networks: in the pain matrix, for the PP of the other group, and in the mentalizing brain regions when presented with thoughts and emotions of the other group. Although these hypotheses have not yet been tested with groups in real ongoing cultural conflicts, suggestive evidence comes from studies manipulating ethnic group membership.
Ethnic differences between the observer and the target can lead to reduced activation in the ‘pain matrix’ brain regions. For example, when Chinese and Caucasian individuals watched a video of a person's face being pierced with a needle (versus touched with a Q-tip), there was less response in CC and insula regions when the ethnicity of the person in the video did not match the ethnicity of the observer [29]. Converging evidence comes from another method: when White participants watched a White hand being pierced by a needle, motor-evoked activity in the observer's hand was suppressed (similar to the reaction when anticipating experiencing the pain directly), but this suppression did not occur when participants watched the needle piercing a Black hand [30].
Plausibly, compared with ethnic differences, ongoing cultural conflict could lead to similarly, but more extremely, reduced responses to an outgroup member's pain. There is evidence that personal history with a target individual influences neural reactions to their pain. Watching one's spouse or significant other receive a painful electric shock elicits a strong response in pain matrix brain regions [15]; and among strangers, the response in these brain regions is less for an individual who had previously cheated in a competitive game against the participant than if the individual previously cooperated [31]. Also, a history of competition or conflict between groups can modulate neural responses to misfortunes of the other group. For example, for avid sports fans, failures of the rival team elicit neural responses related to pleasure and reward, including regions of the ventral striatum, rather than sharing of the other team's misfortune [32,33]. Because cultural conflict includes elements of both interpersonal and intergroup history of competition and conflict, these results suggest that neural responses to another person's pain would be strongly suppressed by group differences, for groups in conflict.
The effects of cultural conflict may also be exaggerated for attributions of complex emotions and mental states, and therefore activity in mentalizing regions. In cultural conflict, people may be willing to ascribe ‘primary’ emotions (such as fear, happiness and PP) equally to members of their own and other groups, as well as to all sentient beings included animals. In contrast, distinctively human ‘secondary’ emotions (such as remorse, embarrassment and humiliation) are sometimes reserved for members of one's own group [34–36], a psychological bias linked to ‘infrahumanization’, i.e. seeing the other group as not just different, but less human, than one's own.
Again, relevant evidence comes from studies of ethnic group differences. When Black participants looked at the photographs of Black victims of hurricane Katrina in Louisiana, there was a higher response in MPFC, a ‘mentalizing’ region, than that to similar photographs of White victims [37]. In this study, there was no group difference in the ‘pain matrix’ regions (and also no difference in MPFC for White participants). Other kinds of group membership also modulate MPFC responses: when viewing photos or making simple social judgements, targets who share the observer's political and social group membership elicit stronger responses in MPFC regions than targets from dissimilar political and social groups [38,39].
In sum, a simple starting hypothesis is that both brain regions in the ‘pain matrix’, and brain regions involved in mentalizing, will show reduced responses to depictions of people suffering when the victims are from other groups; and these group differences will be especially large (i) for misfortunes involving secondary emotions, such as humiliation and remorse, rather than primary emotions such as PP, and (ii) for groups that are involved in active conflicts.
One alternative hypothesis is that previous neural results with unfamiliar groups, such as ethnic differences, will not generalize to groups in active conflict. Rather than eliciting similar but more extreme patterns of neural responses to those of unfamiliar ethnic outgroups, conflict groups may elicit qualitatively different patterns of neural response, because members of conflict groups are highly emotionally and socially significant, and personally relevant. Neural responses to others' misfortunes may therefore depend on the emotional salience of the targets, which is low for distant unfamiliar outgroups, but possibly very high for members of the conflict outgroup. Although less supported by past research, two recent studies provide some evidence for this view: there is higher activity in MPFC for friends from the opposite political group relative to strangers from within one's own group [40], and higher activity in PC for conflict-related narratives that favour a salient conflict outgroup, compared with opinions that favour the ingroup [8].
To test these hypotheses, we asked members of three different cultural groups (Arabs, Israelis and South Americans) to read stories about each other's misfortunes. The stories introduced a protagonist (‘target’) from one of the groups, described their background, and then described a negative event that happened to that individual, resulting in either PP (e.g. breaking a leg, a bad burn, an machinery accident) or emotional suffering (e.g. loss of a home, loss of a job, humiliation and social rejection). After each story, participants were asked how much compassion they felt for the protagonist. We also measured both explicit attitudes, and implicit associations, of the participants towards the groups.
Based on our previous research [41], we predicted that stories about PP would lead to recruitment of the pain matrix regions, whereas stories about emotional suffering would lead to recruitment of the mentalizing brain regions. We posed two key questions: (i) Would any of these regions show a differential response to the stories, based on the relative group membership of the participant and the target? and (ii) Are neural responses to conflict outgroups qualitatively similar or dissimilar to responses to distant outgroups? People from the Middle East versus South America come from widely different cultural (as well as geographical, linguistic and political) backgrounds. Reaching across this gap to feel for someone else requires bridging a large cultural difference, but without the specific obstacle of a history of conflict. In contrast, Arabs and Israelis come from a very similar part of the world, and more similar cultures, but empathy between these groups must overcome almost a century of direct hostilities. Thus, we asked how cultural conflict, versus mere cultural difference, affects people's behavioural and neural responses to others' suffering.
2. Methods
(a). Participants
Participants were 49 adult immigrants or visitors to the United States, who were fluent speakers of English from three groups: Jewish Israeli, Arab and South American (age and gender given in table 1). All participants were born in the Middle East or South America with the exception of two Israeli and one Arab (Palestinian) participants born in the USA who self-identified as ‘Israeli’ and ‘Palestinian’, respectively, and who had spent considerable time in the region. The other Israeli participants were born in Israel; the other Arab participants were born in the Palestinian Territories, Egypt, Jordan, Bulgaria, Bahrain and the United Arab Emirates. South American participants were born in Brazil, Chile, Columbia, Peru, Venezuela and Uruguay.
Table 1.
Participant demographics, by group.
| group | age (years)min/average/max | gender male/female | highest degree high school/college/grad. | years in USA average |
|---|---|---|---|---|
| Arab | 20/25.1/35 | 5/5 | 4/2/3 | 4 |
| Israeli | 21/29.6/37 | 10/6 | 8/4/3 | 8 |
| South American | 19/27.1/40 | 9/7 | 1/9/6 | 4 |
Of the participants who responded to a question about the ‘ethnic group [they] most strongly identify with’ (n = 48), 12/13 Arab participants reported ‘Arab’ (one chose ‘other’); 16/17 Israelis reported ‘Jewish’ (one chose ‘Caucasian–American’); and 14/18 South Americans reported ‘Hispanic’ (three chose ‘other’ and one chose ‘Caucasian–American’). All groups were intermediate in their political conservatism (self-reported on a 9-point Likert scale from (1) liberal to (9) conservative): Israeli mean = 3.7; Arab mean = 3.0; South American mean = 4.6.
Participants were given the opportunity to withdraw their data from the study at any time (no participants did so), and were given compensation and fully debriefed at the end of the experiment.
(b). Design and materials
(i). Survey
Prior to the neuroimaging portion of the study, all participants filled out an online survey that included questions on demographics (age, gender, highest level of education), as well as ethnic identity, conservatism and time spent in the USA. The questionnaire also included the interpersonal reactivity index (IRI) [42], the moral foundation questionnaire (MFQ) [43] and feeling thermometers for a number of different groups (Arabs, Israelis, South Americans, White Americans, Black Americans, Africans, Mexican immigrants). Items were presented one at a time on the screen, and the order of item presentation was randomized separately for each participant.
(ii). Functional magnetic resonance imaging experiment
Seventy-two verbal scenarios involving painful experiences were written, half describing mostly PP and half describing suffering or emotional pain (EP), with normalized group identity (all typical White American names). In a pilot norming study, participants in the USA (n = 60 per story) on Amazon's Mechanical Turk rated each story on 9-point Likert type scales, along six dimensions:
Pain. ‘How much physical pain was the main character in?’
Suffering. ‘How much emotional suffering did the main character experience?’
Discomfort. ‘How uncomfortable did you feel while reading the story?’
Compassion. ‘How much compassion did you feel for the main character in the story?’
Vivid. ‘How vivid and ‘movie-like’ was the story for you?’
Familiar. ‘How familiar were the situations described in the scenario?’
If any participant did not answer a ‘check’ question (‘I have read the story completely and answered all questions honestly’) with anything other than ‘(9) completely agree’, their responses were eliminated. This resulted in the exclusion of approximately 20 per cent of the data.
Overall, the PP stories were rated to involve more physical pain (mean: 7.5 ± 0.84 s.d.) than EP stories (3.2 ± 0.98; paired-samples t-test: t(35) = 19.9, p < 0.001), and less emotional suffering (5.5 ± 0.93) than EP stories (7.8 ± 0.78; paired-samples t-test: t(35) = 10.2, p < 0.001), and more compassion was generated by EP stories (7.4 ± 0.71) than PP stories (6.7 ± 1.1; paired-samples t-test: t(35) = 2.9, p < 0.01); the three other measures (vividness, familiarity, felt discomfort) were not significantly different between conditions (p > 0.05 for all paired-samples t-tests).
The 36 stories from each condition were then divided into three sets of 12 stories such that the sets were matched on all the rated dimensions (p > 0.05 for all paired-samples t-tests between sets across all dimensions) and in average length (mean: 321 characters ± 13 s.d.). Finally, protagonist names were changed, and context was added to each story to identify the group membership (Israeli, Arab, South American) and social background of the protagonist in each story. An additional 12 stories describing South American protagonists with a neutral (non-painful) event were also presented, for a total of 84 stories in seven conditions. Sample scenarios are given in table 2; see the electronic supplementary material for full list of stimuli.
Table 2.
Sample stimuli.
| sample stimuli target group | physical pain (PP) | emotional pain (EP; suffering) |
|---|---|---|
| Arab | Amira lives with her family in Syria. Her father works for the military. Amira was putting up posters in her new room. It was early Saturday morning and she was in her pyjamas. As Amira stood back, she stepped on a thumbtack that had fallen on the ground. The thumbtack went into her heel and hit the bone. | Khalil recently began working for the Hamas government in Gaza. Khalil had worked hard to get the job he has now. He can finally take care of his son and has almost saved enough to give him the gifts he wants for his birthday. Today, Khalil was called in to the office and his boss told him that he was fired. |
| Israeli | Ariel lives with his family in a large settlement in the West Bank. After dinner one day, Ariel was doing the dishes. Ariel was talking with his friends while his hands were in the soapy water. Then his hand hit a sharp knife. The knife cut deep into the skin between his fingers and the cut burned in the dirty water. | Moshe moved from the USA to Israel; his father says it is to support the Jewish homeland. At school, Moshe was happy that some of the boys had offered him some candy. When Moshe put some in his mouth, the boys laughed and said that they had peed on the candy. All the other children started laughing and pointing at Moshe. |
| South American | Viviana lives in a city in Brazil, but works more in rural areas. Viviana was walking with some gifts to her car. She tripped over the curb and turned to avoid crushing the presents. Viviana landed on the curb with her mouth, breaking her two front teeth in half. Viviana dropped the presents and held her bleeding mouth. | Maria moved to the USA from South America to give her children a better life. Maria lives with her teenage daughter. Her daughter wants to have new friends and invites a number of people to her 14th birthday party. Nobody shows up to the birthday. Maria's daughter goes into her room to cry and Maria stands helplessly. |
During neuroimaging, each story was presented for 24 s, followed by a variable inter-stimulus interval (ISIs) ranging from 2 to 12 s. Each run contained two stories per condition (14 total), and ISIs between 2 and 12 s were pseudorandomly ordered and interleaved between each of the stories presented in each run (figure 1). The whole experiment consisted of six runs, each 7.5 min long. The order of conditions and ISIs were counterbalanced across runs and between participants. Stimuli were presented in white 24-point font on a black background in Matlab R2010a (v. 7.10), using the Psychophysics Toolbox v. 3.0 extensions [44] on an Apple MacBook Pro.
Figure 1.

Experimental design. Participants read 84 short verbal narratives while in the fMRI scanner. In all, 14 different stories were presented per run, interleaved with variable inter-stimulus intervals (ISIs) ranging from 2 to 12 s, over a total of six runs.
A response prompt appeared below each story for the final 4 s of presentation. The prompt asked participants to report how much compassion they felt for the main character on a 4-point scale: 1, none; 2, moderate; 3, a lot; 4, extreme (figure 1). Subjects made their response on an magnetic resonance-safe button box. Average responses and reaction times (RTs) for each condition were determined for each individual, and were averaged across each group within each condition.
(iii). Post-imaging behavioural measures
After the neuroimaging component of the study, participants were brought into a separate room and presented with the same stories that they had just seen in the scanner. Participants rated each story using the numbers on the computer across two dimensions (‘How much physical pain was the main character in?’ and ‘How much emotional suffering did the main character experience?’) on a 9-point scale (anchored at ‘none at all’ and ‘extreme’). For each participant, the order of story presentation was identical to the within-scanner presentation order.
Finally, each subject performed an Arab–Israeli implicit association test (IAT). The IAT consisted of eight words from each of four categories: Arab–Muslim names, Israeli–Jewish names, good words and bad words. The words and names appeared sequentially in the centre of the screen in random order, and subjects were required to sort the words and names as quickly as possible by pressing a key on the left of the keyboard (the ‘a’ key) if the word belonged to the category pair presented on the left side of the screen (e.g. Arab names/good words), and a key on the right side of the keyboard (the ‘l’ key) if the word belonged to the category pair presented on the right side of the screen (e.g. Israeli names/bad words). Participants were instructed to respond as fast and as accurately as possible. Each category pairing was tested twice in each participant. Negative attitudes towards the outgroup were measured as percentage differences in RT between pro-Israeli (Israeli/good versus Arab/bad) and pro-Arab (Israeli/bad versus Arab/good) category pairings, as follows: IAT% = 100 × (Avg_RT_Pro_Israeli − Avg_RT_Pro_Arab)/(Avg_RT_All). A positive score indicates a pro-Israeli implicit bias, and a negative score indicates a pro-Arab implicit bias. d-Scores were also computed for each participant [45].
(iv). Functional magnetic resonance imaging image acquisition and data analysis
Participants were scanned using a Siemens Magnetom Tim Trio 3T system (Siemens Solutions, Erlangen, Germany) in the Athinoula A. Martinos Imagining Centre at the McGovern Institute for Brain Research at MIT. Data were collected during six runs with 200 volumes collected per run using a gradient/spin echo pulse sequence (GRAPPA acceleration factor of 2×) on a near axial plane with whole brain coverage (TR = 2 s, TE = 30 ms, flip angle = 90°). Each volume had a field of view 192 × 192 × 120 mm consisting of 30 2 mm slices with a 1 mm skip and an interleaved acquisition order.
MRI data were analysed using SPM8 (http://www.fil.ion.ucl.ac.uk/spm/software/spm8/), SnPM5 (http://www2.warwick.ac.uk/fac/sci/statistics/staff/research/nichols/software/snpm/) and custom software. Each participant's data were motion corrected using the first functional image as a reference and then resliced using first-degree B-spline interpolation. The data were normalized onto a common brain space (Montreal Neurological Institute (MNI), echo planar imaging (EPI) template) using a combined affine and nonlinear transformation model (using discrete cosine basis). Data were smoothed using a Gaussian filter (full width half maximum = 5 mm).
Participants were dropped from the functional magnetic resonance imaging (fMRI) analysis if greater than 20 functional volumes included more than 5 mm of motion or a global mean signal that was greater than 2 s.d. from the session mean. Five participants' data (two Israelis and three Arabs) were dropped from the analyses using these criteria, resulting in 14 Israeli and 10 Arab participants.
Functional images were analysed using both whole brain random effects analyses and group-level-independent functional regions of interest. For whole brain analyses, we first built a modified linear model of the experimental design, and used this model to analyse the blood-oxygen-level dependent (BOLD) response in each voxel. The model included both covariates of interest (the experimental conditions) and nuisance covariates (run effects, an intercept term and global signal). We modelled the conditions as a box-car (matching the onset and duration of each block) convolved with SPM's canonical difference of gammas haemodynamic response function. Time-series data were subjected to a high-pass filter (greater than 0.0078 Hz). To identify voxels in which effects of condition were reliable across participants, BOLD signal differences between conditions (linear combinations of the beta parameters for condition covariates) were submitted to second-level, random effects analysis. All group analyses were conducted using SnPM and used corrected p thresholds, at p < 0.05, based on Monte Carlo simulations of the false positive rate in these data [46].
To identify brain regions in which the magnitude of response was related to individual differences, we performed second-level regression analyses to determine the correlation between brain activity in Arab and Israeli participant to ingroup versus conflict outgroup targets (i.e. Arab protagonists for Israeli participants and Israeli protagonists for Arab participants) across all conditions (ingroup PP and EP > conflict outgroup PP and EP) and three separate behavioural measures: trait empathy (the empathic concern subscale of the IRI), feeling thermometer ratings (warmth towards the ingroup—warmth towards the conflict outgroup) and compassion (average compassion reported across all stories with ingroup protagonists—average compassion reported across all stories with conflict outgroup protagonists). All regressions were examined at p < 0.001, uncorrected and k > 10.
Regions of interest (ROIs) were defined using data from a previous study in which participants read stories describing people experiencing various levels of PP and EP. Regions involved in representing and responding to PP were identified using a continuous regressor, across individual items, quantifying the amount of PP described in each story (based on independent ratings from an online survey). Similarly, regions involved in representing and responding to emotional suffering were identified using a regressor quantifying the amount of emotional suffering described in each story. Coordinates of the peak voxel in each ROI were identified as local maxima in the contrast image, thresholded at p < 0.00001 uncorrected. All supra-threshold voxels within a 9 mm radius from the peak voxel were included in the ROI.
Because group ROIs may only approximately match the relevant functional regions in each individual subjects [47], we also repeated the same analyses in individually tailored functional ROIs. That is, these ROIs were composed of only those voxels in each individual subject that showed a significant preference for emotional or physical stories (all emotional > all physical, or all physical > all emotional) both in the random effects group analysis, and in the individual subject's own contrast. The ROIs were defined as the conjunction of each group ROI and the individual's functional contrast thresholded at p < 0.001 uncorrected. The response of these voxels was then extracted for only the preferred categories (e.g. only emotional stories), and the response was compared across target group (e.g. Israeli, Arab). Because the target group was orthogonal to the contrast used to define these regions, the resulting analyses are not subject to the ‘non-independence’ error [48].
The response at each time point for each story condition was calculated as the average BOLD response across all voxels in each ROI, across all participants; this response was then converted to percentage signal change as follows: PSC = 100 × average_BOLD_response(condition,time)/average_BOLD_response(rest). The BOLD response at rest was calculated as the average signal in each ROI during the rest periods between trials, excluding the 6 s immediately following a trial to account for haemodynamic lag. The data extracted from the ROIs were subject to the preprocessing steps described earlier but were not filtered beyond averaging. All peak voxels are reported in MNI coordinates.
For the purpose of statistics, the average response from 16 to 28 s post-story onset was averaged to create a single average response per subject, per ROI.
Statistical analysis (behavioural and fMRI experiment) used post hoc paired-samples t-tests and repeated-measures ANOVAs, both conducted with an alpha level of 0.05. When the significance level of the Mauchly's test was p < 0.05, we corrected for sphericity using the Greenhouse–Geisser correction, and we report corrected degrees of freedom.
3. Results
(a). Behavioural results
(i). General attitudes and associations about the other groups
Consistent with predictions, Arab and Israeli participants felt warmer towards their ingroup than towards the conflict outgroup (on a scale from 0 to 1: ingroup = 0.80, conflict outgroup = 0.42, t(31) = 6.4, p < 0.001; figure 2), and than towards the distant outgroup (South Americans: mean = 0.68, t(31) = 2.8, p < 0.01). South Americans also showed an ingroup bias in warmth (0.90), but did not distinguish between Arabs (0.73) and Israelis (0.74, t(16) = 0.3, n.s.) on this measure.
Figure 2.

Behavioural results. Each dot shows a single participant; dot shape reflects participant group. (a) The correlation between explicitly reported warmth towards Arabs and Israelis and the difference in reaction time on the IAT while pairing Israeli names with good words and Arab names with bad words, versus the reverse (Pearson's r = 0.66, p < 0.001). (b) The correlation between reported warmth towards the groups as a whole and reported compassion for individuals who are members of those groups experiencing emotional suffering (Pearson's r = 0.64, p < 0.01).
Arab and Israeli participants displayed an ingroup bias in the implicit measure: Israeli RT on the IAT was 24 per cent slower (d = 0.6) for the bad word-Israeli/good word-Arab category pairings than the good word-Israeli/bad word-Arab category pairings, while Arab RT was 10 per cent faster for the same comparison (d = −0.3; difference between Arab and Israeli participants, t(24) = 4.8, p < 0.001); South American participants were 10 per cent slower for the bad word-Israeli/good word-Arab (i.e. an intermediate pro-Israeli bias, d = 0.2, significantly greater than zero, t(16) = 2.5, p < 0.05). Across all participants, explicit and implicit measures of attitudes towards Arabs and Israelis were positively correlated (Pearson's r = 0.66, p < 0.001; figure 2).
(ii). Responses to narrative stimuli
Simply belonging to different or unfamiliar groups did not lead participants to report less compassion for PP or emotional suffering. South American participants did not differ in the compassion they reported for South American versus Middle Eastern targets (rating from 1 to 4: PP stories, ingroup = 2.61, outgroup = 2.61, t(17) = 0.1, n.s.; emotional suffering stories, ingroup = 2.99, mean outgroup = 2.90, t(17) = 1.5, n.s; figure 3, top). Likewise, Israeli and Arab participants did not report less compassion for South American than for ingroup targets (PP stories: ingroup = 2.5, outgroup = 2.4, t(24) = 1.2, n.s.; emotional suffering stories: ingroup = 2.9, outgroup = 2.9, t(24) = 0.15, n.s.; figure 4, top).
Figure 3.

Reactions to stories about physical pain (PP). Top panel, participants' ratings of the compassion they felt for the protagonist in each kind of story, divided by the group membership of the target and of the participant. Bottom panel, average percentage signal change while reading stories about PP and no-pain (control) stories, 8–20 s post-story onset, in the same participants, extracted from six group functional regions of interest (ROIs). Images on the left show the corresponding ROIs, identified based on independent data.
Figure 4.

Reactions to stories about emotional suffering. Top panel, participants' ratings of the compassion they felt for the protagonist (target) in the stories, divided across the group membership of the target and of the participant. Bottom panel, average percentage signal change while reading stories about emotional suffering and no-pain (control) stories, 8–20 s post-story onset, in the same participants, extracted from six group functional regions of interest (identified in independent data). DMPFC and VMPFC, dorsal and ventral medial prefrontal cortex; PC, precuneus; L and R TPJ, left and right temporoparietal junction; LAT, left anterior temporal.
However, being involved in a direct conflict did lead Israeli and Arab participants to report less compassion for targets from the opposite group, both for stories describing PP (ingroup = 2.4, conflict outgroup = 2.2, t(24) = 2.9, p < 0.01; figure 3, top), and for stories describing emotional suffering (ingroup = 2.9, conflict outgroup = 2.4, t(24) = 3.8, p < 0.001; figure 4, top).
In the post-scan ratings, confirming our pilot data, participants from South America judged that the stories about Arab versus Israeli targets depicted equivalent physical pain in PP stories (rated from 1 to 9: Arab = 6.6, Israeli = 6.5), and emotional pain in EP stories (Arab = 7.6, Israeli = 7.5). Israeli and Arab participants also judged that the Israeli and Arab protagonists experienced equivalent physical pain in PP stories (ingroup = 6.1, conflict outgroup = 6.1.), but judged that members of their own group experienced marginally more emotional suffering than members of the other group in EP stories (mean ingroup = 7.4, conflict outgroup = 7.0, t(24) = 2.0, p < 0.06).
(iii). Attitudes towards the other group predict responses to the stimuli
The difference in warmth participants reported towards Arabs and Israelis, in general, was correlated with the difference in compassion participants felt towards specific targets from those groups, both for stories describing PP (Pearson's r = 0.63, p < 0.001) and for stories describing emotional suffering (Pearson's r = 0.64, p < 0.001; figure 2). These correlations were marginally stronger when considering only Arab and Israeli participants (PP r = 0.70; EP r = 0.72, both p-values < 0.001).
(b). Neuroimaging results
(i). Whole brain analysis
First, to find the main effect for the type of misfortune experienced, we contrasted all the stories that focused on PP with all the stories that focused on emotional suffering (EP). Consistent with previous studies using similar stimuli [41], contrasting PP > EP revealed activity in brain regions associated with perceiving human movement and physical sensations (left lateral occipital and bilateral secondary sensory (SII) regions), as well as the most commonly activated components of the ‘pain matrix’ (anterior middle CC, and bilateral insula cortex; table 3a and figure 5a). We also compared stories (describing South American targets) about PP with the no-pain control stories; this contrast revealed activity in the left lateral occipital and bilateral SII (table 3a and figure 5b). Although the insula and CC regions were not visible in this contrast, the ROI analyses (below) suggest that the activity was present, but simply below the conservative statistical threshold used in the random effects contrasts.
Table 3.
(a) Brain regions with higher responses to stories about physical pain (PP). Peak voxels, in MNI coordinates, from six regions, in three contrasts: (i) brain regions correlated with the amount of pain depicted in the stimuli, from independent data, used to define regions of interest; (ii) regions with a higher response to stories about PP than emotional suffering, in the current stories, across all groups; and (iii) regions responding more to stories about South American targets in PP than neutral, no-pain control stories. Left lat occ., left lateral occipital. (b) Brain regions with higher responses to stories about emotional suffering. Peak voxels from seven regions, in three contrasts: (i) brain regions correlated with the amount of emotional suffering depicted in the stimuli, from independent data, used to define regions of interest; (ii) regions with a higher response to stories about emotional suffering than PP, in the current stories, across all groups; and (iii) regions responding more to stories about South American targets in emotional suffering than neutral, no-pain control stories. DMPFC and VMPFC, dorsal and ventral medial prefrontal cortex; PC, precuneus; L and R TPJ, left and right temporo-parietal junction. L and R ATL, left and right anterior temporal lobe.
| region | X | Y | Z | X | Y | Z | X | Y | Z |
|---|---|---|---|---|---|---|---|---|---|
| (a) | rated PP (independent data) | all physical > all emotional | South American PP > control | ||||||
| cingulate | 0 | 6 | 30 | −2 | 4 | 34 | |||
| left insula | −38 | −6 | −10 | −38 | −8 | −12 | |||
| right insula | 40 | 8 | −18 | 38 | −2 | 16 | |||
| left SII | −62 | −24 | 30 | −62 | −34 | 38 | −64 | −30 | 30 |
| right SII | 68 | −26 | 32 | 62 | −34 | 36 | 66 | −26 | 34 |
| left lat. occ. | −50 | −66 | 0 | −58 | −66 | −6 | −58 | −58 | −4 |
| (b) | rated EP (independent data) | all emotional > all physical | South American EP > control | ||||||
| DMPFC | −10 | 48 | 20 | −8 | 48 | 20a | −4 | 54 | 28 |
| VMPFC | −2 | 56 | −10 | −4 | 54 | −6a | −2 | 56 | −10 |
| PC | −8 | −50 | 40 | 0 | −56 | 32 | −2 | −56 | 32 |
| LTPJ | −56 | −58 | 30 | −48 | −62 | 28 | −54 | −60 | 24 |
| RTPJ | 54 | −60 | 30 | 50 | −64 | 24 | 48 | −62 | 24 |
| left ATL | −58 | −20 | −14 | −62 | −12 | −20 | −64 | −12 | −18 |
| right ATL | 58 | −18 | −16 | 58 | −6 | −18 | 52 | −16 | −12a |
aLocal maximum, within a larger cluster.
Figure 5.

Whole brain random effects analyses: brain regions involved in representing physical pain (PP). (a) Regions showing higher responses to stories about PP than emotional suffering, for all participants and targets, including (1) cingulate, (2) left lateral occipital, (3) left insula, (4) left secondary sensory, (5) right insula and (6) right secondary sensory regions. Functional activations corrected for multiple comparisons, p < 0.05; shown on a canonical template brain. (b) Many of the same regions are recruited more during stories about South American targets' PP, than for neutral control stories about South American targets, the no-pain control condition.
The opposite contrast of EP > PP revealed activity in a group of brain regions most commonly associated with mentalizing and ‘theory of mind’, including bilateral TPJ, anterior temporal lobe extending to the temporal poles, PC and ventral and dorsal regions of the MPFC (table 3b and figure 6a). We also compared stories (describing South American targets) about emotional suffering with the no-pain control stories; this contrast revealed activity in dorsal medial prefrontal cortex (DMPFC), ventral medial prefrontal cortex (VMPFC), PC, bilateral TPJ and bilateral lateral temporal regions (table 3b and figure 6b).
Figure 6.

Whole brain random effects analyses: brain regions involved in representing emotional suffering. (a) Regions showing higher responses to stories about emotional suffering than physical pain, for all participants and targets, including (1) DMPFC, (2) VMPFC, (3) PC, (4) LTPJ, (5) RTPJ, (6) left anterior temporal and (7) right anterior temporal regions. Functional activations corrected for multiple comparisons, p < 0.05; shown on a canonical template brain. DMPFC and VMPFC, dorsal and ventral medial prefrontal cortex; PC, precuneus; L- and RTPJ, left and right temporo-parietal junction. (b) The same regions are recruited more during stories about South American targets' emotional suffering, than for neutral control stories about South American targets, the no-pain control condition.
To identify any effects of cultural conflict on these patterns of brain activity, we contrasted the brain responses to stories describing PP in an ingroup target (e.g. Arab target for Arab participants) versus a conflict group target (e.g. Israeli target for Arab participants) and the brain responses for stories describing emotional suffering in an ingroup target versus a conflict group target. For these analyses, we looked only at Arab and Israeli participants. For the stories describing PP, no brain regions showed a significantly different response to ingroup > conflict group, or conflict group > ingroup targets. Similarly, none of the regions typically involved in representing emotional suffering were active for ingroup > conflict group targets. Stories involving emotional suffering of conflict group > ingroup targets led to a small region of increased activity in a left lateral PC/parietal region (peak voxel [−26, −66, 32], p < 0.05 corrected).
Over all, conflict groups did not elicit differential responses in either the pain matrix or the mentalizing regions, in whole brain analyses. We then compared the response in Israeli and Arab participants to ingroup targets versus distant outgroup targets (i.e. South Americans). For stories about PP, there was again no difference in the neural response to ingroup versus distant outgroup targets. However, for stories about emotional suffering, ingroup targets elicited increased responses in five regions involved in processing others' emotions: the DMPFC and VMPFC, PC, right temporoparietal junction (RTPJ) and left anterior temporal regions (electronic supplementary material, figure S1).
We also looked for brain regions showing differential responses to the three target groups (ingroup, conflict outgroup and distant outgroup), averaged across both kinds of misfortune (physical and emotional). In Arab and Israeli participants, reading about an individual from the conflict outgroup, compared with an ingroup target, again resulted in no brain activity at the threshold of p < 0.05, corrected, for either ingroup > conflict group or conflict group > ingroup contrasts. By contrast, the ingroup > distant group contrast resulted in robust activity, localized to ‘theory of mind’ brain regions: bilateral TPJ, PC, VMPFC and bilateral anterior temporal regions; the reverse contrast of distant group > ingroup also resulted in activity, in bilateral sensory-motor regions and dorsal regions of the posterior cingulate (figure 7).
Figure 7.

Whole brain random effects analyses: regions showing differential activity in response to stories about individuals from the ingroup and the distant outgroup. For Arab and Israeli participants only, regions responding to stories about ingroup > distant outgroup (red-yellow), and distant outgroup > ingroup (blue-green), including (i) VMPFC, (ii) PC, (iii) RTPJ, (iv) RIFG, (v) right anterior temporal lobe, (vi) left anterior temporal lobe, (vii) left sensory-motor regions, (viii) right sensory-motor regions and (ix) dorsal posterior cingulate. Functional activations corrected for multiple comparisons, p < 0.05; shown on a canonical template brain. VMPFC, ventral medial prefrontal cortex; PC, precuneus; RTPJ, right temporo-parietal junction; RIFG, right inferior frontal gyrus.
Finally, we asked whether the magnitude of neural response to these stimuli was predicted by individual differences in trait empathy (measured by the IRI empathic concern subscale) or attitudes towards the conflict group (measured by the feeling thermometers, and compassion for outgroup misfortunes). We found no significant correlation, in whole brain analyses, between any of the three measures and the magnitude of response to ingroup versus conflict group targets.
(ii). Regions of interest analyses
Whole brain analyses are conservative tests of activity in a region, and therefore not very sensitive. As a more sensitive test of the effect of target group membership on activity in these brain regions, we next performed an unbiased ROI analysis using functional group ROIs generated from a separate dataset. The ROIs represented the brain regions that were most strongly correlated with ratings of the amount of PP, and of emotional suffering, depicted in a set of verbal narratives describing protagonists with no specific group membership.
We tested the response to stories involving PP in six ROIs: the middle cingulate, left lateral occipital, bilateral insula and bilateral SII regions (figure 3). To confirm that we had identified regions involved in representing PP, we compared the response in these regions to stories about South American participants experiencing emotional suffering, PP or no-pain control (table 4). Bilateral insula and bilateral SII regions showed robustly higher responses to PP stories than the no-pain control (all t(41) > 5.5, all p < 0.001), and no difference between EP stories and the controls. The middle cingulate and left lateral occipital regions both showed significantly higher responses in PP stories, compared with the no-pain control (both t(41) > 2.5, p < 0.05), and significantly lower responses in EP stories (both t(41) > 2, p < 0.05).
Table 4.
Statistical tests in regions of interest. (a) ROIs for representing physical pain (PP). All six ROIs show a higher response to stories about PP than the no-pain control stories. In general, the response is not affected by target group membership. Left and right insula show a higher response to distant outgroup than ingroup stimuli. (b) ROIs for representing emotional suffering. All seven ROIs show robustly higher responses to stories about emotional suffering than to the no-pain control stories (in all participants combined); and differentiate between ingroup and distant group targets, but not between ingroup and conflict group targets (Arab and Israeli participants only).
| region | physical > no pain (South American) | ingroup > conflict group | ingroup > distant group |
|---|---|---|---|
| (a) | |||
| cingulate | t(41) = 2.6, p < 0.01 | t(23) = 1.1, p = 0.3 | T(23) = 3.1, p < 0.01 |
| left insula | t(41) = 6.8, p < 0.001 | t(23) = 1.6, p = 0.1 | t(23) = 3.0, p < 0.01 |
| right insula | t(41) = 3.5, p < 0.01 | t(23) = 1.0, p = 0.3 | t(23) = 3.3, p < 0.01 |
| left SII | t(41) = 7.6, p < 0.001 | t(23) = 1.3, p = 0.2 | t(23) = 1.7, p = 0.1 |
| right SII | t(41) = 7.2, p < 0.001 | t(23) = 1.6, p = 0.1 | t(23) = 2.0, p = 0.07 |
| left lat. occ. | t(41) = 5.6, p < 0.001 | t(23) = 2.1, p < 0.05 | t(23) = 0.6, p = 0.5 |
| (b) | emotional > no pain (South American) | ingroup > conflict group | ingroup > distant group |
| DMPFC | t(41) = 6.6, p < 0.001 | t(23) = 1.7, p = 0.1 | t(23) = 3.1, p < 0.01 |
| VMPFC | t(41) = 8.7, p < 0.001 | t(23) = 0.9, p = 0.4 | t(23) = 6.0 p < 0.001 |
| PC | t(41) = 8.5, p < 0.001 | t(23) = 0.4, p = 0.7 | t(23) = 5.1, p < 0.001 |
| LTPJ | t(41) = 10.3, p < 0.001 | t(23) = 0.2, p = 0.9 | t(23) = 5.1, p < 0.001 |
| RTPJ | t(41) = 6.3, p < 0.001 | t(23) = 0.2, p = 0.9 | t(23) = 9.9, p < 0.001 |
| left ATL | t(41) = 10.3, p < 0.001 | t(23) = 0.9, p = 0.4 | t(23) = 4.3, p < 0.001 |
| right ATL | t(41) = 6.9, p < 0.001 | t(23) = 1.0, p = 0.3 | t(23) = 4.4, p < 0.001 |
Then, we compared the response in these regions to PP stories, separated by the group membership of the target (i.e. protagonist). We found a distinct response to ingroup versus conflict group targets in only one region: the left lateral occipital area response was higher for ingroup targets in PP (0.05% SC) than conflict group targets in PP (0.01% SC, t(23) = 2.1, p < 0.05). Comparing the response to ingroup versus distant outgroup targets, for Arab and Israeli participants, led to a surprising result: both left and right insula, and middle cingulate showed significantly higher responses to stories about distant outgroup (South American) than ingroup targets. However, this difference did not replicate in South American participants. In fact, in South American participants, none of the ROIs differentiated between ingroup and Middle Eastern targets in PP.
Next, we tested the response to stories describing emotional suffering in seven ROIs: bilateral TPJ, bilateral anterior temporal lobe, posterior cingulate, DMPFC and VMPFC (figure 4). To confirm that we had identified regions involved in representing emotional suffering, we compared the response in these regions to stories about South American participants experiencing emotional suffering, PP or no-pain (control). All seven regions showed a robustly higher response to EP stories than no-pain control stories (all t(41) > 6.5, all p < 0.001; table 4). Six of the regions did not differentiate between stories about PP and the no-pain control condition; only the VMPFC showed a significantly lower response to PP than to the control condition (t(41) = 3.5, p < 0.01).
Then, we compared the response in these regions to EP stories, separated by the group membership of the protagonist (table 4). In Arab and Israeli participants, none of these regions differentiated between stories about the emotional suffering of ingroup and conflict group targets (all t(23) < 2, all p > 0.1). By contrast, all seven regions showed robustly higher response to ingroup than distant outgroup (South American) targets (all t(23) > 3, all p < 0.01; figures 4 and 8). Conversely, in South American participants, there was a higher response to stories about emotional suffering of South American than Middle Eastern targets in the DMPFC, left anterior temporal region and left TPJ (all t(23) > 2, p < 0.05).
Figure 8.
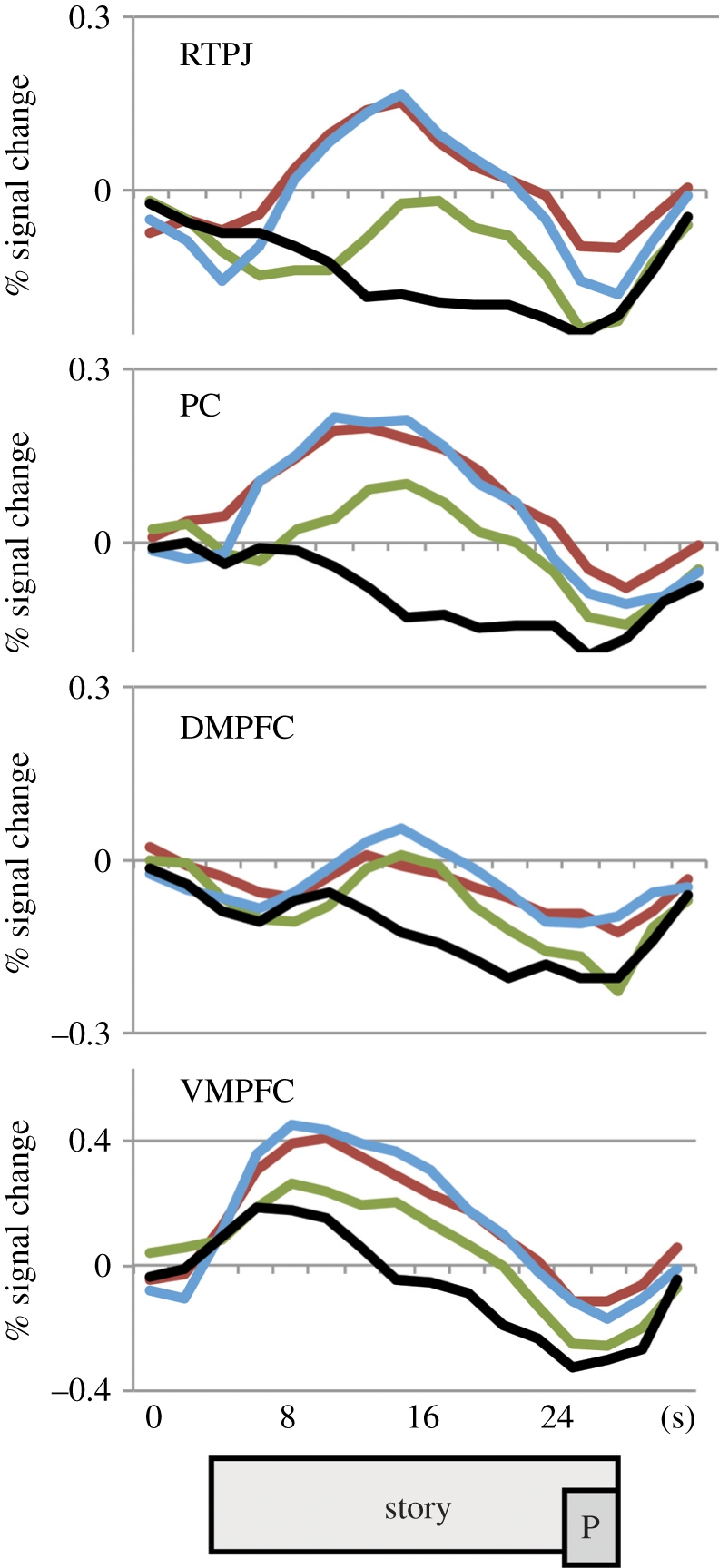
Time course of responses to stories about emotional suffering. The response over the duration of the story, in four representative regions, for Arab and Israeli participants (combined). Colours indicate the relative group membership of the story protagonist: the conflict outgroup (red), the distant outgroup (green) or the participant's ingroup (blue). Black lines show the time course for the no-pain control stories. The rectangles at the bottom show approximate timing of the story, and the compassion question prompt (P; shifted to account for haemodynamic lag).
Overall response magnitudes were higher in the individually tailored ROIs, but the patterns of response across conditions were unchanged (electronic supplementary material, figures S2–S4).
4. Discussion
The current behavioural results provide an illustration of the psychological biases that drive cultural conflict. The participants expressed a specific lack of compassion for individuals in the conflict group, especially in situations eliciting secondary emotions, such as embarrassment, shame and humiliation. The reduction in compassion applied only to the conflict outgroup, not to a distant unfamiliar outgroup, and was related (across individuals) to overall warmth towards the conflict outgroup.
However, the neural responses showed a different pattern. First, we replicated our previous studies, finding that the ‘pain matrix’ regions responded to stories about people in PP, and the ‘mentalizing’ regions responded to stories about emotional suffering. This neural division of labour may be an important clue to the overall neural structure of empathy. Second, we looked for differences in the response of these brain regions based on the group membership of the story's protagonist (the ‘target’). The largest observed neural difference was reduced recruitment of brain regions for understanding emotions and mental states when reading stories about distant outgroup targets (South Americans, for Middle Eastern participants). On the whole, neural responses in mentalizing regions were robust, and equally high, for Arab and Israeli participants reading about Israeli and Arab targets.
The robust response of mentalizing regions for conflict group targets is consistent with other recent evidence that activity in (some of) these regions depends on the relevance or personal significance of the target for the observer. The MPFC, for example, was initially thought to respond more during social inferences about similar (versus dissimilar) others [38]. Subsequent investigations, however, found that the MPFC response depends on personal closeness, or self-relevance, rather than similarity. The response was higher for a dissimilar friend from an opposing political group, than for a similar stranger from the participants' own political group [40].
Conflict groups, in some sense, provide an even stronger test of the same idea: individuals from a conflict group are highly significant and emotionally salient for the participants, although they are also perceived as cold and hostile. Unlike strangers from distant or unfamiliar groups, such as the South Americans in the current study, and perhaps racial outgroup members in previous studies, members of conflict groups elicit strong and familiar emotional reactions.
Relatedly, our previous neuroimaging study of Arabs and Israelis thinking about each others' thoughts found not decreased but increased responses specifically in the PC, one of the mentalizing regions [8]. Activity in the PC was higher for arguments expressing the conflict group's perspective on the conflict, and lower for opinions expressing the ingroup's perspective. We interpreted that result as evidence of participants' own emotional reactions to the expressed opinions and arguments, which they judged ‘unreasonable’.
Nevertheless, the absence of any region whose neural response mirrors the pattern of behavioural judgements is puzzling. Participants report a significant difference in the compassion that they feel for ingroup versus conflict group targets, but no brain regions show a similarly suppressed response. We also did not observe clear evidence of Schadenfreude (pleasure at the outgroup's suffering), as indexed by activity in ventral striatum [32].
To some extent, the absence of differences between ingroup and conflict targets may be related to lack of sensitivity in the experimental design and procedures. The Arab and Israeli populations studied here were living far from the Middle East, away from a daily experience of conflict. Participants were fluent in English and interested in participating in a scientific study; thus, the population in the study may be biased towards more moderate and cosmopolitan members of each group. Still, we observed robust group differences in the explicit warmth, implicit associations and reported compassion felt towards each group; so a moderate population cannot entirely account for the absence of group effects in the neural response.
Another possibility is that the neural response measured here reflects a complex combination of responses to the presence of an ‘enemy’, and responses to the specific misfortune described in the story. The current design did not include neutral or positive events experienced by ingroup or conflict group members; so we cannot distinguish responses to the event from responses to the target individual as a whole.
One key result of the current experiment is the clear and robust difference between neural responses to the conflict group versus the distant outgroup. While stories about distant outgroup members elicited reduced responses in standard ‘mentalizing’ regions, compared with ingroup targets, stories about conflict group members were indistinguishable from ingroup targets in these brain regions. These results illustrate the importance of the current design, using both distant and conflict groups, for future studies. Past work both in social psychology and neuroimaging has often focused on ethnic groups whose relationship with each other is characterized more by distance, ambivalence and ignorance than open hostility. In order for this literature to inform our growing understanding of the psychological processes involved in the context of protracted conflict, we must better understand how these processes compare across distant and conflict groups.
In addition to specific conclusions, these results illustrate the possibility of a biology of cultural conflict. The current experiment focused on one psychological bias that can influence and perpetuate conflicts: failures of empathy towards the suffering of outgroup members. However, there are many other psychological biases that contribute to conflict, and that could be investigated using cognitive neuroscience. Here, we discuss three topics that are ripe for future investigation.
First, future studies should investigate the interaction between cultural conflict and neural correlates of moral judgement. Nearly universally, people judge that it is wrong to harm another person. Nevertheless, in the context of intergroup conflict, people often excuse and even endorse deliberately harming members of the other group. Even preschool children say that it is more OK to hurt another person, if that person is from another group [49]. Neuroimaging studies are beginning to identify a complex network of brain regions involved in moral judgements: for example, brain regions involved in calculating costs and benefits of actions involving moral tradeoffs, in reasoning about the intentions of the perpetrators, and in reacting emotionally to negative outcomes [25,50–53]. Any or all of these processes could be influenced by intergroup conflicts.
Second, future studies should investigate the influence of cultural conflict on neural correlates of economic decision-making; for example, about fair distribution of resources. When all other factors are equal, people have strong preferences for fair distributions of resources that give each individual an equal (or proportional) share of any available good. In fact, the preference for fair outcomes can trump self-interest: people will punish unfair outcomes, at their own expense and with no likelihood of personal gain [54]. However, when assigned to one of multiple competing groups, people's preferences shift to a strong ingroup bias, preferring distributions that favour the ingroup [55]. Again, even very young children show this pattern: a general preference for equal shares, trumped by an ingroup bias in competitive contexts [56]. Although many fMRI studies have characterized neural mechanisms for calculating value, for preferring fair outcomes and for altruistic punishment [57–59], none have yet looked at the effects on these neural mechanisms of active conflict.
Finally, a key future direction will be to use the tools of cognitive neuroscience to help design and evaluate conflict resolution programmes. For example, alternative strategies for conflict resolution could be quantitatively compared in their ability to reduce or mitigate neural correlates of intergroup conflict (‘neuro-evaluation’). Alternatively, neural measures could identify signatures that predict who will benefit, and who will not, from a particular intervention [60].
As a practical assessment tool, neuro-evaluation would share a number of characteristics with neuro-marketing, which uses brain imaging to assess consumer preferences. Neuro-marketing assumes that consumer behaviour is caused at least in part by subconscious motives that are undetectable by questionnaires or focus groups. Neuro-marketers aim to look ‘under the hood’ at these motives, and thus hope to outperform surveys and focus groups in predicting subsequent consumer behaviour [61]. Similarly, there is considerable evidence that the causes of intergroup behaviour and attitudes are at least partially inaccessible to the participant themselves (and thus missed by standard explicit measures), and more differentiated than a single positive–negative dimension (and thus missed by standard implicit measures [62,63]. Like neuro-marketing, neuro-evaluation offers the chance to look ‘under the hood’ at these causes of intergroup behaviour.
Even more importantly, in both neuro-marketing and neuro-evaluation, actual behaviour provides a ground-truth for comparing alternative behavioural and neural predictors. Neuro-marketers will be evaluated by their ability to predict actual buying behaviour outside the laboratory. Likewise, an ideal neural measure of intergroup hostility should not only be correlated with explicit attitudes and implicit associations, but specifically should outperform (cheaper and faster) behavioural assessments in predicting long-term pro-social and anti-social intergroup behaviour outside the laboratory (e.g. intergroup friendships, or voting for or participating in negotiations rather than violent conflict). In principle, we believe that neuro-evaluation could outperform behavioural assessments in exactly this way. For both neuro-evaluation and neuro-marketing, however, this horizon remains a long way off.
Acknowledgements
Funding for this work was provided by the Air Force Office of Scientific Research, managed through the Office of Naval Research, grant number N000140910845.
References
- 1.Bar-Tal D. 2007. Sociopsychological foundations of intractable conflicts. Am. Behav. Sci. 50, 1430–1453 10.1177/0002764207302462 (doi:10.1177/0002764207302462) [DOI] [Google Scholar]
- 2.Krueger J. I., Funder D. C. 2004. Towards a balanced social psychology: causes, consequences, and cures for the problem-seeking approach to social behavior and cognition. Behav. Brain Sci. 27, 313–327 [DOI] [PubMed] [Google Scholar]
- 3.Ross L., Stillinger C. 1991. Barriers to conflict resolution. Negot. J. 7, 389–404 10.1111/j.1571-9979.1991.tb00634.x (doi:10.1111/j.1571-9979.1991.tb00634.x) [DOI] [Google Scholar]
- 4.Ross L. 1995. Reactive devaluation in negotiation and conflict resolution. In Barriers to Conflict Resolution (eds Arrow K., Mnookin R. H., Ross L., Tversky A., Wilson R.), pp. 26–43 New York, NY: W. W. Norton [Google Scholar]
- 5.Kennedy K. A., Pronin E. 2008. When disagreement gets ugly: perceptions of bias and the escalation of conflict. Pers. Soc. Psychol. Bull. 34, 833–848 10.1177/0146167208315158 (doi:10.1177/0146167208315158) [DOI] [PubMed] [Google Scholar]
- 6.Maoz I., McCauley C. 2008. Threat, dehumanization, and support for retaliatory aggressive policies in asymmetric conflict. J. Confl. Resolut. 52, 93–116 10.1177/0022002707308597 (doi:10.1177/0022002707308597) [DOI] [Google Scholar]
- 7.Cikara M., Bruneau E. G., Saxe R. R. 2011. Us and them. Curr. Dir. Psychol. Sci. 20, 149–153 10.1177/0963721411408713 (doi:10.1177/0963721411408713) [DOI] [Google Scholar]
- 8.Bruneau E. G., Saxe R. 2010. Attitudes towards the outgroup are predicted by activity in the precuneus in Arabs and Israelis. Neuroimage 52, 1704–1711 10.1016/j.neuroimage.2010.05.057 (doi:10.1016/j.neuroimage.2010.05.057) [DOI] [PubMed] [Google Scholar]
- 9.Saxe R., Kanwisher N. 2003. People thinking about thinking people: the role of the temporo-parietal junction in ‘theory of mind’. Neuroimage 19, 1835–1842 10.1016/S1053-8119(03)00230-1 (doi:10.1016/S1053-8119(03)00230-1) [DOI] [PubMed] [Google Scholar]
- 10.Saxe R., Powell L. J. 2006. It's the thought that counts: specific brain regions for one component of theory of mind. Psychol. Sci. 17, 692–699 10.1111/j.1467-9280.2006.01768.x (doi:10.1111/j.1467-9280.2006.01768.x) [DOI] [PubMed] [Google Scholar]
- 11.Chiao J., Mathur V. 2010. Intergroup empathy: how does race affect empathic neural responses? Curr. Biol. 20, R478–R480 10.1016/j.cub.2010.04.001 (doi:10.1016/j.cub.2010.04.001) [DOI] [PubMed] [Google Scholar]
- 12.Decety J., Jackson P. 2006. A social-neuroscience perspective on empathy. Curr. Dir. Psychol. Sci. 15, 54–58 10.1111/j.0963-7214.2006.00406.x (doi:10.1111/j.0963-7214.2006.00406.x) [DOI] [Google Scholar]
- 13.Jackson P., Meltzoff A., Decety J. 2005. How do we perceive the pain of others? A window into the neural processes involved in empathy. Neuroimage 24, 771–779 10.1016/j.neuroimage.2004.09.006 (doi:10.1016/j.neuroimage.2004.09.006) [DOI] [PubMed] [Google Scholar]
- 14.Singer T., Lamm C. 2009. The social neuroscience of empathy. Ann. N.Y. Acad. Sci. 1156, 81–96 10.1111/j.1749-6632.2009.04418.x (doi:10.1111/j.1749-6632.2009.04418.x) [DOI] [PubMed] [Google Scholar]
- 15.Singer T., Seymour B., O'Doherty J., Kaube H., Dolan R., Frith C. 2004. Empathy for pain involves the affective but not sensory components of pain. Science 303, 1157–1162 10.1126/science.1093535 (doi:10.1126/science.1093535) [DOI] [PubMed] [Google Scholar]
- 16.Hutchison W. D., Davis K., Lozano A., Tasker R., Dostrovsky J. 1999. Pain-related neurons in the human cingulate cortex. Nat. Neurosci. 2, 403–405 10.1038/8065 (doi:10.1038/8065) [DOI] [PubMed] [Google Scholar]
- 17.Jackson P., Meltzoff A., Decety J. 2006. Neural circuits involved in imitation and perspective-taking. Neuroimage 31, 429–439 10.1016/j.neuroimage.2005.11.026 (doi:10.1016/j.neuroimage.2005.11.026) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lamm C., Decety J., Singer T. 2010. Meta-analytic evidence for common and distinct neural networks associated with directly experienced pain and empathy for pain. Neuroimage 54, 2492–2502 10.1016/j.neuroimage.2010.10.014 (doi:10.1016/j.neuroimage.2010.10.014) [DOI] [PubMed] [Google Scholar]
- 19.Botvinick M., Jha A., Bylsma L., Fabian S., Solomon P., Prkachin K. 2005. Viewing facial expressions of pain engages cortical areas involved in the direct experience of pain. Neuroimage 25, 312–319 10.1016/j.neuroimage.2004.11.043 (doi:10.1016/j.neuroimage.2004.11.043) [DOI] [PubMed] [Google Scholar]
- 20.Lamm C., Batson C., Decety J. 2007. The neural substrate of human empathy: effects of perspective-taking and cognitive appraisal. J. Cogn. Neurosci. 19, 42–58 10.1162/jocn.2007.19.1.42 (doi:10.1162/jocn.2007.19.1.42) [DOI] [PubMed] [Google Scholar]
- 21.Saarela M., Hlushchuk Y., Williams A., Schurmann M., Kalso E., Hari R. 2007. The compassionate brain: humans detect intensity of pain from another's face. Cereb. Cortex. 17, 230–237 10.1093/cercor/bhj141 (doi:10.1093/cercor/bhj141) [DOI] [PubMed] [Google Scholar]
- 22.Preston S. D., De Waal F. B. M. 2002. Empathy: its ultimate and proximate bases. Behav. Brain Sci. 25, 1–20 [DOI] [PubMed] [Google Scholar]
- 23.Morrison I., Downing P. 2007. Organization of felt and seen pain responses in anterior cingulate cortex. Neuroimage 37, 642–651 10.1016/j.neuroimage.2007.03.079 (doi:10.1016/j.neuroimage.2007.03.079) [DOI] [PubMed] [Google Scholar]
- 24.Young L., Cushman F., Hauser M., Saxe R. 2007. The neural basis of the interaction between theory of mind and moral judgment. Proc. Natl Acad. Sci. 104, 8235–8240 10.1073/pnas.0701408104 (doi:10.1073/pnas.0701408104) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Young L., Saxe R. 2008. The neural basis of belief encoding and integration in moral judgment. Neuroimage 40, 1912–1920 10.1016/j.neuroimage.2008.01.057 (doi:10.1016/j.neuroimage.2008.01.057) [DOI] [PubMed] [Google Scholar]
- 26.Shamay-Tsoory S., Tibi-Elhanany Y., Aharon-Peretz J. 2006. The ventromedial prefrontal cortex is involved in understanding affective but not cognitive theory of mind stories. Soc. Neurosci. 1, 149–166 10.1080/17470910600985589 (doi:10.1080/17470910600985589) [DOI] [PubMed] [Google Scholar]
- 27.Shamay-Tsoory S., Tomer R., Berger B., Aharon-Peretz J. 2003. Characterization of empathy deficits following prefrontal brain damage: the role of the right ventromedial prefrontal cortex. J. Cogn. Neurosci. 15, 324–337 10.1162/089892903321593063 (doi:10.1162/089892903321593063) [DOI] [PubMed] [Google Scholar]
- 28.Jenkins A. C., Macrae C. N., Mitchell J. P. 2008. Repetition suppression of ventromedial prefrontal activity during judgments of self and others. Proc. Natl Acad. Sci. 105, 4507–4512 10.1073/pnas.0708785105 (doi:10.1073/pnas.0708785105) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Xu X., Zuo X., Wang X., Han S. 2009. Do you feel my pain? Racial group membership modulates empathic neural responses. J. Neurosci. 29, 8525–8529 10.1523/jneurosci.2418-09.2009 (doi:10.1523/jneurosci.2418-09.2009) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Avenanti A., Sirigu A., Aglioti S. M. 2010. Racial bias reduces empathic sensorimotor resonance with other-race pain. Curr. Biol. 20, 1018–1022 10.1016/j.cub.2010.03.071 (doi:10.1016/j.cub.2010.03.071) [DOI] [PubMed] [Google Scholar]
- 31.Singer T., Seymour B., O'Doherty J., Stephan K., Dolan R., Frith C. 2006. Empathic neural responses are modulated by the perceived fairness of others. Nature 439, 466–469 10.1038/nature04271 (doi:10.1038/nature04271) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Cikara M., Botvinick M. M., Fiske S. T. 2011. Us versus them. Psychol. Sci. 22, 306–313 10.1177/0956797610397667 (doi:10.1177/0956797610397667) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Hein G., Silani G., Preuschoff K., Batson C. D., Singer T. 2010. Neural responses to ingroup and outgroup members' suffering predict individual differences in costly helping. Neuron 68, 149–160 10.1016/j.neuron.2010.09.003 (doi:10.1016/j.neuron.2010.09.003) [DOI] [PubMed] [Google Scholar]
- 34.Demoulin S., Leyens J., Paladino M., Rodriguez-Torres R., Rodriguez-Perez A., Dovidio J. 2004. Dimensions of ‘uniquely’ and ‘non-uniquely’ human emotions. Cogn. Emot. 18, 71–96 10.1080/02699930244000444 (doi:10.1080/02699930244000444) [DOI] [Google Scholar]
- 35.Leyens J., Rodriguez-Perez A., Rodriguez-Torres R., Gaunt R., Paladino M., Vaes J., Demoulin S. 2001. Psychological essentialism and the differential attribution of uniquely human emotions to ingroups and outgroups. Eur. J. Soc. Psychol. 31, 395–411 10.1002/ejsp.50 (doi:10.1002/ejsp.50) [DOI] [Google Scholar]
- 36.Paladino M., Leyens J., Rodriguez R., Rodriguez A., Gaunt R., Demoulin S. 2002. Differential association of uniquely and non uniquely human emotions with the ingroup and the outgroup. Group Process. Intergroup Relat. 5, 105–117 10.1177/1368430202005002539 (doi:10.1177/1368430202005002539) [DOI] [Google Scholar]
- 37.Mathur V., Harada T., Lipke T., Chiao J. 2010. Neural basis of extraordinary empathy and altruistic motivation. Neuroimage 51, 1468–1475 10.1016/j.neuroimage.2010.03.025 (doi:10.1016/j.neuroimage.2010.03.025) [DOI] [PubMed] [Google Scholar]
- 38.Mitchell J. P., Macrae C. N., Banaji M. R. 2006. Dissociable medial prefrontal contributions to judgments of similar and dissimilar others. Neuron 50, 655–663 10.1016/j.neuron.2006.03.040 (doi:10.1016/j.neuron.2006.03.040) [DOI] [PubMed] [Google Scholar]
- 39.Harris L. T., Fiske S. T. 2006. Dehumanizing the lowest of the low. Psychol. Sci. 17, 847–853 10.1111/j.1467-9280.2006.01793.x (doi:10.1111/j.1467-9280.2006.01793.x) [DOI] [PubMed] [Google Scholar]
- 40.Krienen F. M., Tu P. C., Buckner R. L. 2010. Clan mentality: evidence that the medial prefrontal cortex responds to close others. J. Neurosci. 30, 13 906–13 915 10.1523/JNEUROSCI.2180-10.2010 (doi:10.1523/JNEUROSCI.2180-10.2010) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Bruneau E. G., Pluta A., Saxe R. 2011. In press Distinct roles of the ‘shared pain’ and ‘theory of mind’ networks in processing others’ emotional suffering. Neuropsychologia. 10.1016/j.neuropsychologia.2011.11.008 (doi:10.1016/j.neuropsychologia.2011.11.008) [DOI] [PubMed] [Google Scholar]
- 42.Davis M. H. 1983. Measuring individual differences in empathy: evidence for a multidimensional approach. J. Pers. Soc. Psychol. 44, 113–126 10.1037/0022-3514.44.1.113 (doi:10.1037/0022-3514.44.1.113) [DOI] [Google Scholar]
- 43.Graham J., Haidt J., Nosek B. 2009. Liberals and conservatives rely on different sets of moral foundations. J. Pers. Soc. Psychol. 96, 1029–1046 10.1037/a0015141 (doi:10.1037/a0015141) [DOI] [PubMed] [Google Scholar]
- 44.Kleiner M., Brainard D., Pelli D. 2007. What's new in Psychtoolbox-3. Perception 36 (ECVP abstr. suppl.) [Google Scholar]
- 45.Greenwald A. G., Nosck B. A., Banaji M. R. 2003. Understanding and using the implicit association test: I. An improved scoring algorithm. J. Pers. Soc. Psychol. 85, 197–216 10.1037/0022-3514.85.2.197 (doi:10.1037/0022-3514.85.2.197) [DOI] [PubMed] [Google Scholar]
- 46.Nichols T., Holmes A. 2004. Nonparametric permutation tests for functional neuroimaging. Hum. Brain Funct. 2, 887–910 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Fedorenko E., Hsieh P. J., Nieto-Castañon A., Whitfield-Gabrieli S., Kanwisher N. 2010. New method for fMRI investigations of language: defining ROIs functionally in individual subjects. J. Neurophysiol. 104, 1177–1194 10.1152/jn.00032.2010 (doi:10.1152/jn.00032.2010) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Vul E., Harris C., Winkielman P., Pashler H. 2009. Voodoo correlations in social neuroscience. Perspect. Psychol. Sci. 4, 274–290 10.1111/j.1745-6924.2009.01125.x (doi:10.1111/j.1745-6924.2009.01125.x) [DOI] [PubMed] [Google Scholar]
- 49.Rhodes M., Brickman D. 2011. The influence of competition on children's social categories. J. Cognit. Dev. 12, 194–221 10.1080/15248372.2010.535230 (doi:10.1080/15248372.2010.535230) [DOI] [Google Scholar]
- 50.Greene J. D., Nystrom L. E., Engell A. D., Darley J. M., Cohen J. D. 2004. The neural bases of cognitive conflict and control in moral judgment. Neuron 44, 389–400 10.1016/j.neuron.2004.09.027 (doi:10.1016/j.neuron.2004.09.027) [DOI] [PubMed] [Google Scholar]
- 51.Greene J. D., Sommerville R. B., Nystrom L. E., Darley J. M., Cohen J. D. 2001. An fMRI investigation of emotional engagement in moral judgment. Science 293, 2105–2108 10.1126/science.1062872 (doi:10.1126/science.1062872) [DOI] [PubMed] [Google Scholar]
- 52.Moll J., de Oliveira-Souza R. 2007. Moral judgments, emotions and the utilitarian brain. Trends Cogn. Sci. 11, 319–321 10.1016/j.tics.2007.06.001 (doi:10.1016/j.tics.2007.06.001) [DOI] [PubMed] [Google Scholar]
- 53.Young L., Saxe R. 2009. An fMRI investigation of spontaneous mental state inference for moral judgment. J. Cogn. Neurosci. 21, 1396–1405 10.1162/jocn.2009.21137 (doi:10.1162/jocn.2009.21137) [DOI] [PubMed] [Google Scholar]
- 54.Fehr E., Gachter S. 2002. Altruistic punishment in humans. Nature 415, 137–140 10.1038/415137a (doi:10.1038/415137a) [DOI] [PubMed] [Google Scholar]
- 55.Tajfel H., Billig M. G., Bundy R. P., Flament C. 1971. Social categorization and intergroup behaviour. Eur. J. Soc. Psychol. 1, 149–178 10.1002/ejsp.2420010202 (doi:10.1002/ejsp.2420010202) [DOI] [Google Scholar]
- 56.Fehr E., Bernhard H., Rockenbach B. 2008. Egalitarianism in young children. Nature 454, 1079–1084 10.1038/nature07155 (doi:10.1038/nature07155) [DOI] [PubMed] [Google Scholar]
- 57.Rilling J. K., Sanfey A. G., Aronson J. A., Nystrom L. E., Cohen J. D. 2004. Opposing BOLD responses to reciprocated and unreciprocated altruism in putative reward pathways. Neuroreport 15, 2539–2543 10.1097/00001756-200411150-00022 (doi:10.1097/00001756-200411150-00022) [DOI] [PubMed] [Google Scholar]
- 58.King-Casas B., Tomlin D., Anen C., Camerer C. F., Quartz S. R., Montague P. R. 2005. Getting to know you: reputation and trust in a two-person economic exchange. Science 308, 78–83 10.1126/science.1108062 (doi:10.1126/science.1108062) [DOI] [PubMed] [Google Scholar]
- 59.Plassmann H., O'Doherty J., Rangel A. 2007. Orbitofrontal cortex encodes willingness to pay in everyday economic transactions. J. Neurosci. 27, 9984. 10.1523/JNEUROSCI.2131-07.2007 (doi:10.1523/JNEUROSCI.2131-07.2007) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Gordon E. 2007. Integrating genomics and neuromarkers for the era of brain-related personalized medicine. Pers. Med. 4, 201–215 10.2217/17410541.4.2.201 (doi:10.2217/17410541.4.2.201) [DOI] [PubMed] [Google Scholar]
- 61.Fugate D. L. 2008. Marketing services more effectively with neuromarketing research: a look into the future. J. Serv. Mark. 22, 170–173 10.1108/08876040810862903 (doi:10.1108/08876040810862903) [DOI] [Google Scholar]
- 62.Fazio R. H., Olson M. A. 2003. Implicit measures in social cognition research: their meaning and use. Annu. Rev. Psychol. 54, 297–327 10.1146/annurev.psych.54.101601.145225 (doi:10.1146/annurev.psych.54.101601.145225) [DOI] [PubMed] [Google Scholar]
- 63.Greenwald A. G., Banaji M. R. 1995. Implicit social cognition: attitudes, self-esteem, and stereotypes. Psychol. Rev. 102, 4–27 10.1037/0033-295X.102.1.4 (doi:10.1037/0033-295X.102.1.4) [DOI] [PubMed] [Google Scholar]