

Abstract
Colorectal cancer (CRC) has become one of the most common fatal cancers. CRC tumorigenesis is a complex process involving multiple genetic changes to several sequential mutations or molecular alterations. P53 is one of the most significant genes; its mutations account for more than half of all CRC. Therefore, understanding the cellular genes that are directly or indirectly related to p53 is particularly crucial for investigating CRC tumorigenesis. In this study, a p53-related ribosomal protein, ribosomal protein S19 (RPS19), obtained from the feces of CRC patients is evaluated by using specifically quantitative real-time PCR and knocked down in the colonic cell line by gene silencing. This study found that CRC patients with higher expressions of RPS19 in their feces had a better prognosis and consistent expressions of RPS19 and BAX in their colonic cells. In conclusion, the potential mechanism of RPS19 in CRC possibly involves cellular apoptosis through the BAX/p53 pathway, and the levels of fecal RPS19 may function as a prognostic predictor for CRC patients.
1. Introduction
Despite progress in reducing the incidence and mortality rate and improving patient survival, human cancers still account for numerous deaths [1]. Colorectal cancer (CRC) has become one of the most common fatal cancers, involving a complex process with multiple genetic changes [2–4]. This molecular heterogeneity possibly results from multiple sequential mutations or molecular alterations during tumorigenesis [5]. Therefore, the identification of CRC-related genes will assist in cancer prevention, detection, and prognostic prediction [6–8].
One important tumor suppressor, p53, is known to prevent cancer, but is also involved in CRC progression [9, 10]. Mutations of p53 account for more than half of all CRCs, particularly in patients at the more advanced stages [11]. Numerous cellular genes are also out of control because of the abnormal p53 expression during tumor progression [12, 13]. For example, the p53-related ribosomal proteins (RPs) were identified as cancer-related molecules [14, 15], indicating that the oncogenic potential of RPs result from the relationship between p53 and RPs [16–18]. Moreover, the p53-inducible modulator RPS27-like (RPS27L), which responds to genotoxic stress, was recently evaluated in CRC [19].
Feces can serve as the material for detecting genetic alterations in CRC [20–22]. Numerous ribosomal proteins are significantly expressed in the feces of CRC patients [23]. In this study, p53-related RPS19 of CRC was further evaluated. Other studies have reported on the developmental abnormalities resulting from a RPS19 deficiency through the activation of the p53 protein family [24]. First, the clinical significance of RPS19 in feces was evaluated from the stool samples of CRC patients using specifically, quantitative real-time PCR (qRT-PCR). Then, the functional importance of RPS19 was addressed by silencing its expression in colonic cells. In this context, we explored the possible cell fate of changing the RPS19 expression in colonic cells, which could affect CRC patients' survival.
2. Materials and Methods
2.1. Patients
Solid fecal samples of 101 CRC patients (n male = 69; n female = 32) from the Cathay General Hospital and the Taipei Veterans General Hospital were obtained before surgery or application of chemotherapy, with IRB-approved informed consent. Follow-up data were obtained prospectively, and the mean follow-up time was 44.1 months (SD, 29.0; median, 37.8). Patients' initial tumor stage and additional clinical information are listed in Table 1. Patients with distant metastasis were routinely confirmed by abdominal computed tomography.
Table 1.
Clinical characteristics of CRC patients.
| Variable | No. of cases | Level of fecal RPS19 (%) (≥2.76 × 10−5) |
P |
|---|---|---|---|
| Age (yr)* | 0.448 | ||
| <64.7 | 47 | 27 (57.4) | |
| ≥64.7 | 54 | 35 (64.8) | |
| Gender | 0.140 | ||
| Male | 69 | 39 (56.5) | |
| Female | 32 | 23 (71.9) | |
| Depth of invasion | 0.303 | ||
| T1 + T2 | 15 | 11 (73.3) | |
| T3 + T4 | 86 | 51 (59.3) | |
| Lymphatic invasion | 0.068 | ||
| N0 | 53 | 37 (69.8) | |
| N1 + N2 + N3 | 48 | 25 (52.1) | |
| Distant metastasis | 0.183 | ||
| No | 84 | 54 (64.3) | |
| Yes | 17 | 8 (47.1) | |
| Tumor size (cm)** | 0.682 | ||
| <4.7 | 61 | 36 (59.0) | |
| ≥4.7 | 38 | 24 (63.2) | |
| CEA (ng/mL)** | 0.907 | ||
| ≤5 | 61 | 37 (60.7) | |
| >5 | 37 | 22 (59.5) | |
| CA19-9 (U/mL)** | 0.829 | ||
| <37 | 74 | 45 (60.8) | |
| ≥37 | 24 | 14 (58.3) |
*mean age of 101 patients, 64.7 ys (range, 37.3–89.5). **available cases in tumor size, 99; in serum CEA and CA19-9 determinant, 98.
2.2. Total RNA Extraction and Reverse Transcription Reaction
The HCT116 cell line was cultured in Dulbecco's modified Eagle's medium with 5 mM of glutamine according to routine culture procedures. RNA from this cell line was extracted using the Easy Pure Total RNA Mini Kit (Bioman, Taiwan) and reverse transcribed for single-stranded cDNAs using an oligo(dT)12 primer with the ABI Reverse Transcriptase kit (ABI, USA), according to the manufacturer's protocols [25]. Fecal RNA was prepared and reverse transcribed as described in our previous reports [22]. In brief, synthesized cDNA could be used directly in the following quantitative PCR analyses.
2.3. Quantitative Real-Time PCR (qRT-PCR)
The quantitations of RPS19 (NM001022), BAX (NM138764), and 18s rRNA (X03205) in fecal cDNA were performed using a TaqMan probe (probe no. 87 for RPS19, no. 55 for BAX, and no. 77 for 18 s rRNA) from the Human Universal Probe Library (Roche Diagnostics, Germany). The 18 s rRNA served as a housekeeping gene. Generally, each run of fecal samples must include the human reference cDNA (Clontech, USA) as standard to avoid errors because of run-to-run differences in RNA quantity. The primer sequences for these quantitations are listed in Table 2.
Table 2.
Primers' sequences and universal probe numbers for qRT-PCR analysis.
| Gene name | Primer sequence* | Probe no. |
|---|---|---|
| RPS19 | F: 5′-TCAGGGACAAAGAGATCTGGA-3′ | 87 |
| R: 5′-CATGGTTTGTTCTAATGCTTCTTG-3′ | ||
| BAX | F: 5′-CAAGACCAGGGTGGTTGG-3′ | 55 |
| R: 5′-CACTCCCGCCACAAAGAT-3′ | ||
| 18 s RNA | F: 5′-CTCAACACGGGAAACCTCAC-3′ | 77 |
| R: 5′-CGCTCCACCAACTAAGAACG-3′ |
*F, forward; R, reverse; Probe no., from the “Human Universal Probe Library” of Roche Diagnostics, Mannheim, Germany.
2.4. Lentivirus-Mediated RNA Interference (RNAi) of RPS19
The lentiviral construct encoding the siRPS19 hairpin (pLKO.1-RPS19 : TRCN0000074915) for gene silencing (shRPS19) was obtained from the National RNAi Core Facility located at the Institute of Molecular Biology/Genomic Research Center, Academia Sinica, Taipei Taiwan. Additionally, the control (shLuc) for the lentivirus was pLKO.1-Luc (TRCN0000072246), and the infection of each lentivirus into colonic cells was performed according to our previous report [20]. The change in the expression of target RPS19 was quantified by qRT-PCR as previously described and immunodetected using western blotting as the routine procedure with minor modifications [25]. Briefly, 5 μg of protein was mixed with the reducing agent NuPAGE SDS sample buffer (Life Technologies, Carlsbad, CA, USA), denatured for 10 min at 95°C, separated by a 12% SDS-PAGE, blotted onto a polyvinylidene difluoride membrane (Millipore, Billerica, MA, USA), and probed with mouse anti-human RPS19 (1 : 1000; sc-100836; Santa Cruz Biotechnology, Santa Cruz, CA, USA) or rabbit anti-human actin (1 : 500; sc-1616-R; Santa Cruz Biotechnology) following standard procedures. The blots were then incubated with anti-mouse (for RPS19) or anti-rabbit (for actin) secondary antibodies (0.2 μg/mL) conjugated to horseradish peroxidase. All western blots were developed using the Western Blot Chemiluminescence Reagent (PerkinElmer Life and Analytical Sciences, Waltham, MA, USA) according to the manufacturer's instructions.
2.5. Statistical Analysis
The overall survival probabilities were estimated using the Kaplan-Meier method and compared with the log-rank test using SPSS 13.0 software (SPSS). The MedCalc software statistical package was used to generate a receiver operating characteristic (ROC) curve. Significance was set at P < 0.05.
3. Results
Over 40 ribosomal or ribosomal-associated genes were clustered due to their significantly differential expressions (P < 0.05) in the feces of CRC patients from our previous report [23]. Notably, six p53-associated RPs, including two large- and four small-ribosomal protein transcripts, were clustered by the average-linkage hierarchical clustering method (Figure 1). The 11 CRC patients and two distinct normal pools could be classified into two groups (E and L). First, 80% (4 of 5) of patients of the L group were in the late Dukes' stages (one in Dukes' stage C and three in stage D), and, with the exception of RPS6 (NM001010), their genes were downregulated. Conversely, RPS27L (NM015920), RPS7 (NM001011), and RPL26 (NM000987) were clustered together due to their increasing expressions in the feces of those in group E, which comprised mostly early-stage patients (67%, 4 of 6) and normal controls.
Figure 1.
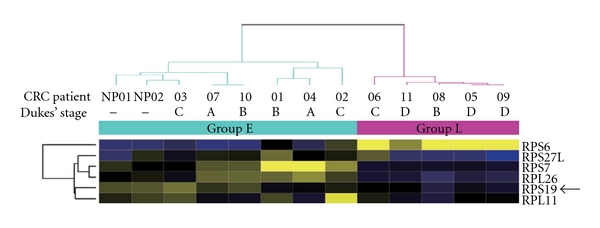
Six differentially expressed p53-associated ribosomal proteins in feces of CRC patients. Differentially expressed genes with statistic significance (P < 0.05) are grouped by average-linkage hierarchical clustering. Each row represents a gene and each column is a sample. Group L, five patients (one at Dukes' stage B, one at stage C, and three at stage D). Group E, six patients (two at Dukes' stage A, two at stage B, and two at stage C) and two normal control pools (NP01 and NP02). NP01, pooled by two healthy men; NP02, pooled by three healthy women. A region cluster depicts the genes based on the similarity between their expressions in cases. High expression is shown in deep yellow, low expression in blue. Arrow indicates RPS19.
The clinical significances of RPS19 were further analyzed. The mRNA levels of RPS19 in the feces of CRC patients were stratified into two groups, RPS19+ (≥2.76 × 10−5) and RPS19− (<2.76 × 10−5), using ROC curve analysis. The area under the ROC curve for fecal RPS27L was 0.657 (P = 0.012) with a 95% CI of 0.556 to 0.749 (Figure 2). As shown in Figure 3, the RPS19+ group (n = 62) had the better six-year overall survival rate (74.3 ± 12.2%) than the RPS19− group did (40.9 ± 14.2%; n = 39) (P = 0.008, log-rank test).
Figure 2.
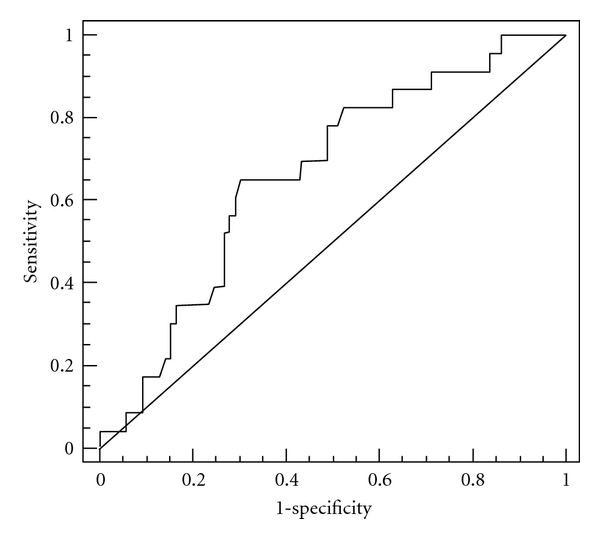
Receiver operating characteristic (ROC) curve for fecal RPS19. The points on the curve represent the relative mRNA levels of RPS19 in the feces and the sensitivity and (1-specificity) of the marker for overall survival.
Figure 3.
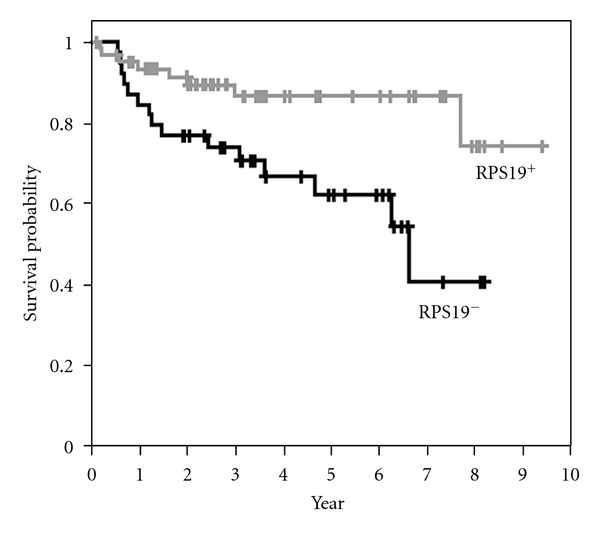
The Kaplan-Meier overall survival curves in patients with colorectal cancer according to fecal RPS19. The relative mRNA levels of RPS19 in the feces are stratified into two groups: RPS19− (<2.76 × 10−5) and RPS19+ (≥2.76 × 10−5). The six-year overall survival rate of the RPS19+ group (n = 62) is better than that of the RPS19− group (P = 0.008, log-rank test).
To correlate the cellular function of RPS19, we infected lentiviruses into a colonic cell line with wild-type p53 (HCT116 p53+/+) to knockdown RPS19 (shRPS19) and control shRNA (shLuc). In the lentivirus-mediated RNAi experiment, shRPS19 achieved efficient knockdown at both the levels of mRNA and RPS19 protein (Figure 4). These RPS19-silent cells expressed only 9.2% BAX mRNA from RPS19-expressing cells compared to that from RPS19-expressing cells (Figure 5).
Figure 4.
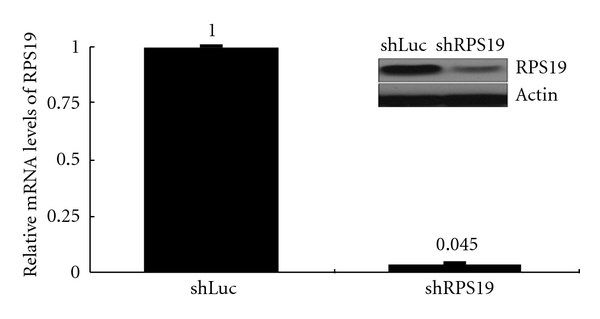
Efficiency of RPS19 silence in colonic cells by RNA interference. RPS19 silence is achieved by the lentivirus-mediated RNAi experiment. Relative mRNA levels of RPS19 are quantified by qRT-PCR with TaqMan probes and normalized by individual level of 18 s rRNA. The relative expression level of shLuc-infected cells is considered as 1. Results are representative of those obtained in two-to-three separate experiments with error bars showing standard error. Changes of protein levels are immunoblotted with antibodies against RPS19 and β-actin (in black square).
Figure 5.
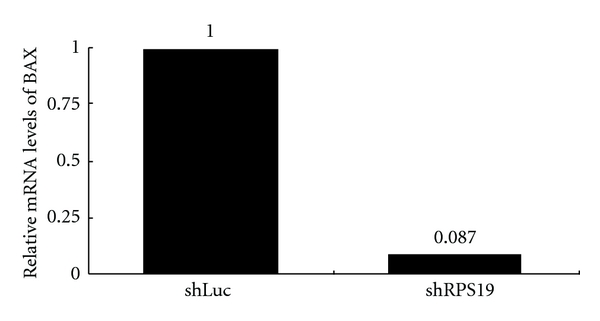
Changes of BAX expression in RPS19-silent colonic cells. Relative mRNA levels of BAX are quantified by qRT-PCR with TaqMan probes and normalized by individual level of 18 s rRNA. The relative expression level of shLuc-infected cells is considered as 1. Results are representative of those obtained in one experiment.
4. Discussion
Our previous results revealed that numerous fecal molecules were differentially expressed in the feces of CRC patients [26]. Among the fecal molecules, a number of RP genes were listed with statistic significance [23]. RPs are involved not only in the cellular process of translation [27] but also in the growth and maintenance of all cell types [28]. Additionally, numerous reports emphasized that a strong correlation was found between RPs and p53 protein in cellular functions [17, 18, 29]. For example, RPL26 can regulate the translation and induction of p53 after DNA damage [30]. RPS6 participated in the activation of a p53-dependent cell cycle checkpoint [31]. RPS27L was proven to be a p53-inducible modulator of cell fate in response to genotoxic stress [32], and mutant p53 seemed to cause aberrant RPS27L expression, which can lead to the accumulation of tumorigenic CRC cells and a poor prognosis [19].
Besides the p53-associated RPs, other RPs were also revealed to contribute to the onset of cancer or multidrug resistance [33–35]. For example, RPS3 is involved in the onset of cancer [33], RPS13 and RPL23 promote the multidrug resistance of gastric cancer cells [35], and RPL19 is involved in the prognosis of prostate cancer and CRC [23, 36]. Recently, the direct relationship between feces and colonic cells was proven [37, 38]. When combined, detecting fecal RPs, specifically p53-related RPs, might be valuable for evaluating the molecular pathogenesis of CRC [14, 15, 39].
In this study, the overall survival data revealed that CRC patients with higher fecal expressions of RPS19 had a better prognosis. When RPS19 was knocked down in colonic cells, an apoptotic gene (BAX) extremely reduced the expressions in RPS19-silent cells. Nevertheless, most studies of RPS19 examined mutations in the RPS19 gene in patients with Diamond-Blackfan anemia [40, 41]. Cells with an RPS19 deficiency correlate with p53 dysregulation, which may cause developmental anomalies [24]. To our knowledge, we are the first to reveal the differentially expressed RPS19 in CRC with clinical significance. From the results of consistent expressions of RPS19 and BAX, we suggest that downregulated RPS19 might impair the apoptotic function of colonic cells. This argument supports the clinical data that CRC patients with lower fecal expressions of RPS19 had a poor prognosis.
5. Conclusions
We succeeded in quantifying the mRNA level of RPS19 in feces. The potential mechanism of RPS19 in CRC possibly involves cellular apoptosis through the BAX/p53 pathway [42]. Our results provide some evidence that the levels of fecal RPS19 may function as a prognostic predictor for CRC patients. Finally, clinical use of feces in translational research is promising for the future of CRC diagnosis [43].
Authors' Contribution
C.-C. Chien and T.-C. Tu contributed equally to this work.
Acknowledgments
The authors would like to thank all of the patients involved in this study. This study was partially supported by grant from the National Science Council, Taiwan (NSC100-2320-B-281-001 to C.-J. Huang).
References
- 1.Jemal A, Siegel R, Ward E, Hao Y, Xu J, Thun MJ. Cancer statistics, 2009. CA Cancer Journal for Clinicians. 2009;59(4):225–249. doi: 10.3322/caac.20006. [DOI] [PubMed] [Google Scholar]
- 2.Bardelli A, Velculescu VE. Mutational analysis of gene families in human cancer. Current Opinion in Genetics and Development. 2005;15(1):5–12. doi: 10.1016/j.gde.2004.12.009. [DOI] [PubMed] [Google Scholar]
- 3.Rozen P. Cancer of the gastrointestinal tract: early detection or early prevention? European Journal of Cancer Prevention. 2004;13(1):71–75. doi: 10.1097/00008469-200402000-00011. [DOI] [PubMed] [Google Scholar]
- 4.Vogelstein B, Fearon ER, Hamilton SR, et al. Genetic alterations during colorectal-tumor development. New England Journal of Medicine. 1988;319(9):525–532. doi: 10.1056/NEJM198809013190901. [DOI] [PubMed] [Google Scholar]
- 5.Beerenwinkel N, Antal T, Dingli D, et al. Genetic progression and the waiting time to cancer. PLoS Computational Biology. 2007;3, article e225 doi: 10.1371/journal.pcbi.0030225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Center MM, Jemal A, Smith RA, Ward E. Worldwide variations in colorectal cancer. CA Cancer Journal for Clinicians. 2009;59(6):366–378. doi: 10.3322/caac.20038. [DOI] [PubMed] [Google Scholar]
- 7.Kimura Y, Sumiyoshi M, Baba K. Antitumor activities of synthetic and natural stilbenes through antiangiogenic action. Cancer Science. 2008;99(10):2083–2096. doi: 10.1111/j.1349-7006.2008.00948.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Watine JC, Bunting PS. Mass colorectal cancer screening: methodological quality of practice guidelines is not related to their content validity. Clinical Biochemistry. 2008;41(7-8):459–466. doi: 10.1016/j.clinbiochem.2007.12.020. [DOI] [PubMed] [Google Scholar]
- 9.Kikuchi-Yanoshita R, Konishi M, Ito S, et al. Genetic changes of both p53 alleles associated with the conversion from colorectal adenoma to early carcinoma in familial adenomatous polyposis and non-familial adenomatous polyposis patients. Cancer Research. 1992;52(14):3965–3971. [PubMed] [Google Scholar]
- 10.Lane DP, Benchimol S. p53: oncogene or anti-oncogene? Genes and Development. 1990;4(1):1–8. doi: 10.1101/gad.4.1.1. [DOI] [PubMed] [Google Scholar]
- 11.Iacopetta B. TP53 mutation in colorectal cancer. Human Mutation. 2003;21(3):271–276. doi: 10.1002/humu.10175. [DOI] [PubMed] [Google Scholar]
- 12.Hague A, Paraskeva C. Apoptosis and disease: a matter of cell fate. Cell Death and Differentiation. 2004;11(12):1366–1372. doi: 10.1038/sj.cdd.4401497. [DOI] [PubMed] [Google Scholar]
- 13.Hofseth LJ, Hussain SP, Harris CC. p53: 25 years after its discovery. Trends in Pharmacological Sciences. 2004;25(4):177–181. doi: 10.1016/j.tips.2004.02.009. [DOI] [PubMed] [Google Scholar]
- 14.Kobayashi T, Sasaki Y, Oshima Y, et al. Activation of the ribosomal protein L13 gene in human gastrointestinal cancer. International Journal of Molecular Medicine. 2006;18(1):161–170. [PubMed] [Google Scholar]
- 15.Wang H, Zhao LN, Li KZ, Ling R, Li XJ, Wang L. Overexpression of ribosomal protein L15 is associated with cell proliferation in gastric cancer. BMC Cancer. 2006;6, article no. 91 doi: 10.1186/1471-2407-6-91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Anderson SJ, Lauritsen JPH, Hartman MG, et al. Ablation of ribosomal protein L22 selectively impairs αβ T cell development by activation of a p53-dependent checkpoint. Immunity. 2007;26(6):759–772. doi: 10.1016/j.immuni.2007.04.012. [DOI] [PubMed] [Google Scholar]
- 17.Chen D, Zhang Z, Li M, et al. Ribosomal protein S7 as a novel modulator of p53-MDM2 interaction: Binding to MDM2, stabilization of p53 protein, and activation of p53 function. Oncogene. 2007;26(35):5029–5037. doi: 10.1038/sj.onc.1210327. [DOI] [PubMed] [Google Scholar]
- 18.Ofir-Rosenfeld Y, Boggs K, Michael D, Kastan MB, Oren M. Mdm2 regulates p53 mRNA translation through inhibitory interactions with ribosomal protein L26. Molecular Cell. 2008;32(2):180–189. doi: 10.1016/j.molcel.2008.08.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Tien LT, Chien CC, Yang SH, Lin CM, Wu YY, Huang CJ. p53-dependent expression of ribosomal protein S27-like in colorectal cancer. Fu-Jen Journal of Medicine. 2010;8(1):11–19. [Google Scholar]
- 20.Huang C-J, Yang S-H, Huang S-M, et al. A predicted protein, KIAA0247, is a cell cycle modulator in colorectal cancer cells under 5-FU treatment. Journal of Translational Medicine. 2011;9, article 82 doi: 10.1186/1479-5876-9-82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kanaoka S, Yoshida KI, Miura N, Sugimura H, Kajimura M. Potential usefulness of detecting cyclooxygenase 2 messenger RNA in feces for colorectal cancer screening. Gastroenterology. 2004;127(2):422–427. doi: 10.1053/j.gastro.2004.05.022. [DOI] [PubMed] [Google Scholar]
- 22.Yang RN, Yang SH, Chang CC, Chien CC, Pan S, Huang CJ. Upregulation of fecal cytokeratin 19 is associated with prognosis in older colorectal cancer patients. Genetic Testing and Molecular Biomarkers. 2010;14(5):703–708. doi: 10.1089/gtmb.2010.0047. [DOI] [PubMed] [Google Scholar]
- 23.Huang CJ, Chien CC, Yang SH, et al. Faecal ribosomal protein L19 is a genetic prognostic factor for survival in colorectal cancer. Journal of Cellular and Molecular Medicine. 2008;12(5B):1936–1943. doi: 10.1111/j.1582-4934.2008.00253.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Danilova N, Sakamoto KM, Lin S. Ribosomal protein S19 deficiency in zebrafish leads to developmental abnormalities and defective erythropoiesis through activation of p53 protein family. Blood. 2008;112(13):5228–5237. doi: 10.1182/blood-2008-01-132290. [DOI] [PubMed] [Google Scholar]
- 25.Chang CC, Yang SH, Chien CC, et al. Clinical meaning of age-related expression of fecal cytokeratin 19 in colorectal malignancy. BMC Cancer. 2009;9, article no. 376 doi: 10.1186/1471-2407-9-376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Chien CC, Chang CC, Yang SH, Chen SH, Huang CJ. A homologue of the Drosophila headcase protein is a novel tumor marker for early-stage colorectal cancer. Oncology reports. 2006;15(4):919–926. [PubMed] [Google Scholar]
- 27.Ramakrishnan V, Moore PB. Atomic structures at last: the ribosome in 2000. Current Opinion in Structural Biology. 2001;11(2):144–154. doi: 10.1016/s0959-440x(00)00184-6. [DOI] [PubMed] [Google Scholar]
- 28.Wool IG. The structure and function of eukaryotic ribosomes. Annual Review of Biochemistry. 1979;48:719–754. doi: 10.1146/annurev.bi.48.070179.003443. [DOI] [PubMed] [Google Scholar]
- 29.MacInnes AW, Amsterdam A, Whittaker CA, Hopkins N, Lees JA. Loss of p53 synthesis in zebrafish tumors with ribosomal protein gene mutations. Proceedings of the National Academy of Sciences of the United States of America. 2008;105(30):10408–10413. doi: 10.1073/pnas.0805036105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Takagi M, Absalon MJ, McLure KG, Kastan MB. Regulation of p53 translation and induction after DNA damage by ribosomal protein L26 and nucleolin. Cell. 2005;123(1):49–63. doi: 10.1016/j.cell.2005.07.034. [DOI] [PubMed] [Google Scholar]
- 31.Panić L, Tamarut S, Sticker-Jantscheff M, et al. Ribosomal protein S6 gene haploinsufficiency is associated with activation of a p53-dependent checkpoint during gastrulation. Molecular and Cellular Biology. 2006;26(23):8880–8891. doi: 10.1128/MCB.00751-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Li J, Tan J, Zhuang L, et al. Ribosomal protein S27-like, a p53-inducible modulator of cell fate in response to genotoxic stress. Cancer Research. 2007;67(23):11317–11326. doi: 10.1158/0008-5472.CAN-07-1088. [DOI] [PubMed] [Google Scholar]
- 33.Hegde V, Yadavilli S, Deutsch WA. Knockdown of ribosomal protein S3 protects human cells from genotoxic stress. DNA Repair. 2007;6(1):94–99. doi: 10.1016/j.dnarep.2006.09.004. [DOI] [PubMed] [Google Scholar]
- 34.Shen DW, Liang XJ, Suzuki T, Gottesman MM. Identification by functional cloning from a retroviral cDNA library of cDNAs for ribosomal protein L36 and the 10-kDa heat shock protein that confer cisplatin resistance. Molecular Pharmacology. 2006;69(4):1383–1388. doi: 10.1124/mol.105.017525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Shi Y, Zhai H, Wang X, et al. Ribosomal proteins S13 and L23 promote multidrug resistance in gastric cancer cells by suppressing drug-induced apoptosis. Experimental Cell Research. 2004;296(2):337–346. doi: 10.1016/j.yexcr.2004.02.009. [DOI] [PubMed] [Google Scholar]
- 36.Bee A, Ke Y, Forootan S, et al. Ribosomal protein L19 is a prognostic marker for human prostate cancer. Clinical Cancer Research. 2006;12(7):2061–2065. doi: 10.1158/1078-0432.CCR-05-2445. [DOI] [PubMed] [Google Scholar]
- 37.Chung DC. Stool DNA testing and colon cancer prevention: another step forward. Annals of Internal Medicine. 2008;149(7):509–510. doi: 10.7326/0003-4819-149-7-200810070-00013. [DOI] [PubMed] [Google Scholar]
- 38.Yajima S, Ishii M, Matsushita H, et al. Expression profiling of fecal colonocytes for RNA-based screening of colorectal cancer. International Journal of Oncology. 2007;31(5):1029–1037. [PubMed] [Google Scholar]
- 39.Clemens MJ. Targets and mechanisms for the regulation of translation in malignant transformation. Oncogene. 2004;23(18):3180–3188. doi: 10.1038/sj.onc.1207544. [DOI] [PubMed] [Google Scholar]
- 40.Badhai J, Schuster J, Gidlöf O, Dahl N. 5′UTR variants of ribosomal protein S19 transcript determine translational efficiency: Implications for Diamond-Blackfan anemia and tissue variability. PLoS One. 2011;6(3) doi: 10.1371/journal.pone.0017672. Article ID e17672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Flygare J, Karlsson S. Diamond-Blackfan anemia: erythropoiesis lost in translation. Blood. 2007;109(8):3152–3154. doi: 10.1182/blood-2006-09-001222. [DOI] [PubMed] [Google Scholar]
- 42.Sturm I, Köhne CH, Wolff G, et al. Analysis of the p53/BAX pathway in colorectal cancer: low BAX is a negative prognostic factor in patients with resected liver metastases. Journal of Clinical Oncology. 1999;17(5):1364–1374. doi: 10.1200/JCO.1999.17.5.1364. [DOI] [PubMed] [Google Scholar]
- 43.Koga Y, Yasunaga M, Katayose S, et al. Improved recovery of exfoliated colonocytes from feces using newly developed immunomagnetic beads. Gastroenterology Research Practice. 2008;2008:7 pages. doi: 10.1155/2008/605273. Article ID 605273. [DOI] [PMC free article] [PubMed] [Google Scholar]