

Abstract
Cucurbitacin E, a tetracyclic triterpenes compound extracted from cucurbitaceous plants, has been shown to exhibit anticancer and anti-inflammatory activities. The purpose of this study was to elucidate whether cucurbitacin E promotes cell cycle arrest and induces apoptosis in T24 cells and further to explore the underlying molecular mechanisms. The effects of cucurbitacin E on T24 cell's growth and accompanied morphological changes were examined by MTT assay and a phase-contrast microscope. DNA content, mitochondrial membrane potential (ΔΨm) and annexin V/PI staining were determined by flow cytometry. The protein levels were measured by Western blotting. Our results demonstrated that cucurbitacin E-induced G2/M arrest was associated with a marked increase in the levels of p53, p21 and a decrease in phospho-signal transducer and activator of transcription 3 (STAT3), cyclin-dependent kinase 1 (CDK1) and cyclin B. Cucurbitacin E-triggered apoptosis was accompanied with up-regulation of Fas/CD95, truncated BID (t-BID) and a loss of ΔΨm, resulting in the releases of cytochrome c, apoptotic protease activating factor 1 (Apaf-1) and apoptosis-inducing factor (AIF), and sequential activation of caspase-8, caspase-9, and caspase-3. Our findings provided the first evidence that STAT3/p53/p21 signaling, Fas/CD95 and mitochondria-dependent pathways play critical roles in cucurbitacin E-induced G2/M phase arrest and apoptosis of T24 cells.
1. Introduction
In 2008, approximately 37500 new bladder cancer cases were diagnosed worldwide [1]. In the United States, approximately 68,810 people and 14,100 deaths from bladder cancer were reported in 2008 alone [1]. According to the Taiwan Department of Health annual reports 2009, bladder cancer is 14th leading cause of cancer death in Taiwanese men and about three persons per one hundred thousand die annually from that. Today, bladder cancer is usually treated with surgery, chemotherapy, and combination of chemotherapy and radiotherapy, but they are often intolerable because of the strong systemic toxicity and local irritation [2]. Therefore, the finding for a novel adjuvant agent to treat bladder cancer that can reduce the recurrence rate, decrease side effects, and increase overall survival is urgent. In the past ten years, investigators have focused on plant food-derived phytochemicals to find potential anticancer drugs [3, 4]. Approximately 70% of the currently approved anticancer drugs are derived from or based on natural products [5]. Cucurbitacin E (molecular formula: C32H44O8) is a natural flavonoid found in Cucurbitaceae but also presents in other plants [6–8] and traditional Chinese herbal medicine such as Cucurbita pepo cv dayangua [9]. Cucurbitacin E has been reported to exhibit biological activities, including antioxidant, anti-inflammatory, and anticancer effects [6, 9, 10]. Cucurbitacin E induced cytotoxic effects, including suppression of the proliferation and induction of apoptosis in ovarian [11, 12], leukemia [13, 14], and pancreatic [15] cancer cell lines. Cucurbitacin E also suppressed tumor angiogenesis through interacting vascular-endothelial-growth-factor-receptor-2-(VEGFR2-) mediated Janus kinase 2 (Jak2)-signal transducer and activator of transcription 3 (STAT3) signals [9].
It is well documented that apoptosis plays an important role in the maintenance of tissue homeostasis for the elimination of excessive cells [16]. However, it is also well known that the induction of apoptosis of cancer cells by anticancer drugs such as etoposide, cisplatin, and paclitaxel has been used for treatment of cancer in target cells [17–20]. Many reports have shown that numerous cytotoxic and DNA damaging agents could arrest the cell cycle at the G1, S, or G2/M phase and induce apoptotic cell death [21–23] which is involved in downregulation of phosphorylated STAT3, an oncogene that has a vital role at all stages of tumorigenesis [15]. Therefore, the present study investigated the induction of apoptosis by cucurbitacin E in human bladder cancer cells, and we also attempted to clarify the possible signaling pathways involved in cucurbitacin E-induced apoptosis. Our results indicated cucurbitacin E induced G2/M phase arrest and apoptosis in human bladder cancer T24 cells through STAT3/p53/p21, Fas/CD95, and mitochondria-dependent pathways.
2. Material and Methods
2.1. Chemicals and Reagents
Cucurbitacin E, dimethyl sulfoxide (DMSO), propidium iodide (PI), 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), RNase A, and Triton X-100 were purchased from Sigma-Aldrich Corp. (St. Louis, MO, USA). All primary and secondary antibodies were obtained from Santa Cruz Biotechnology Inc. (Santa Cruz, CA, USA). The fluorescent probe DiOC6 and all culture media and reagents were purchased from Invitrogen Life Technologies (Carlsbad, CA, USA).
2.2. Cell Culture
The human bladder cancer cell line (T24) (transitional cell carcinoma) was purchased from the Food Industry Research and Development Institute (Hsinchu, Taiwan) and was cultured with McCoy's 5a medium supplemented with 10% FBS, 100 Units/mL penicillin, 100 μg/mL streptomycin, and 2 mM L-glutamine at 37°C under a humidified 5% CO2 and 95% air at one atmosphere. The medium was changed every 2 days [24].
2.3. Determinations of Cell Morphology and Viability
About 2 × 105 T24 cells/well were seeded into 12-well plates for cell adherence with three wells for each concentration level. Cucurbitacin E was individually added to the final concentrations of 0, 250, 500, 1000, and 2000 nM, respectively. In control group, equal amount of 1% DMSO was added, while in blank control group, only culture medium was added to each well. In the end of incubation for 24 or 48 h, cells from each well were examined and photographed under a phase-contrast microscope at 200× magnification and then cell viability was determined using the MTT method, as described previously [25, 26].
2.4. Cell Cycle Distribution and Phase Determination
The cell cycle distribution and sub-G1 phase were analyzed by flow cytometry, as described previously [27, 28]. A total of 2 × 105 T24 cells/well on 12-well plates were incubated with 1000 nM cucurbitacin E or only with vehicle (DMSO, 1% in culture media) for 0, 12, 24, and 48 h. At the end of incubation, the cells on each well were trypsinized, washed with PBS, and fixed with 70% ethanol overnight at −20°C. At the next day, all fixed cells were from experimental and control groups washed twice with PBS and then were stained with PI-working solution (40 μg/mL PI and 100 μg/mL RNase A and 0.1% Triton X-100) for cellular staining at room temperature for 1 h in the dark. Then all cells were analyzed for cell cycle distribution and sub-G1 phase (apoptosis) by flow cytometry, as described previously [27, 28]. The fractions of the cells in G0/G1, S, and G2/M phase were analyzed using a FACSCalibur flow cytometer (Becton-Dickinson, Franklin Lakes, NJ, USA), and data were analyzed with a ModFit LT program (Verity Software House Inc., Topsham, ME, USA).
2.5. Assessment of Apoptosis
Apoptotic cells were measured with an Annexin V-FITC/PI detection kit (Invitrogen Life Technologies). Approximately 2 × 105 cells in 12-well plates per treatment or control samples (1000 nM cucurbitacin E for 12 and 24 h) were harvested and analyzed by flow cytometry, as described previously [26, 29]. Annexin V-FITC is a sensitive probe for identifying cells undergoing apoptosis as phosphatidylserine (PS) exposure occurs early in the apoptotic process. PI is excluded from live cells with intact plasma membranes, but it is incorporated into nonviable cells.
2.6. CDK1 Kinase Activity
T24 cells were seeded onto 75 cm2 tissue culture flask and then treated with 0, 250, 500, and 1000 nM of cucurbitacin E for 24 h. Cells were suspended in a final volume of 0.2 mL buffer containing 50 mM Tris-HC1 (pH 7.5), 1 mM phenylmethylsulfonyl fluoride, 50 pg/mL leupeptin, 10 mM 2-mercaptoethanol, 1 mM MgCl2, 2 mM EGTA, 0.5 mM dithiothreitol, 0.01% Brij35, 25 mM β-glycerophosphate, and 0.5 M NaCl. Cell suspensions were sonicated and centrifuged at 10,000 ×g for 30 min. CDK1 kinase activity condition was determined by using MV Peptide (CDK1 kinase assay kit, Medical & Biological Laboratories Co., Ltd., Nagoya, Japan) and measuring OD492, as described previously [26, 30].
2.7. Assay of Mitochondrial Membrane Potential (ΔΨm)
A density of 2 × 105 T24 cells/well on 12-well plates was maintained with 1000 nM cucurbitacin E for 0, 4, 8, 12, 24, and 48 h, to determine the level of ΔΨm by flow cytometry. At the end of incubation, cells were harvested, washed twice by PBS, and then resuspended in 500 μL of DiOC6 (1 μmol/l) for the level of ΔΨm. Subsequently, cells were maintained at dark room for 30 min at 37°C and then all samples were analyzed immediately by flow cytometry, as described previously [31, 32].
2.8. Detection of Cell Viability after Pretreatment with a General Caspase Inhibitor
T24 cells at a density of 1 × 104 cells/100 μL seeded in 96-well plates were preincubated with or without 10 μM of the general caspase inhibitor (Z-VAD-FMK, R&D Systems, Minneapolis, MN, USA) for 2 h before cells were exposed to 1000 nM cucurbitacin E for a 48-h exposure. Thereafter, cells were determined for the viability by using a MTT assay, as previously described [25, 26].
2.9. Western Blot Analysis for Protein Levels
T24 cells (1 × 106/well) seeded into 6-well plates were treated with 1000 nM cucurbitacin E or solvent control. Cells were harvested from each treatment then were washed with cold PBS and then scraped and washed twice by centrifugation at 1,000xg for 5 min at 4°C. All pellets were individually resuspended in the PRO-PREP protein extraction solution (iNtRON Biotechnology, Seongnam, Gyeonggi-Do, Korea) for 3 h at −20°C, as described previously [33, 34]. The lysate from each sample was collected by centrifugation at 12,000 ×g for 30 min at 4°C, and the supernatant was stored at −20°C. Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) gels were used to separate the protein before each sample was incubated with the primary antibodies (Santa Cruz Biotechnology Inc.) followed by secondary antibodies. These blots were then detected by ECL kit (Millipore, Bedford, MA, USA) and autoradiography using X-ray film [27, 35]. Each PVDF membrane (Immobilon-P; Millipore) was stripped and reported with anti-β-actin antibody as the loading control for ensuring that equal proteins were loaded [32, 36].
2.10. Statistical Analysis
All values are expressed as mean ± standard deviation (S.D.) and performed in triplicate. One-way ANOVA was used to compare the difference between control and concentration treatments in each group followed by Bonferroni's multiple comparison tests. A P value of less than 0.05 was considered to be statistically significant.
3. Results
3.1. The Effects of Cucurbitacin E on Cell Growth and Viability of T24 Cells
The examined concentrations of cucurbitacin E inhibited cell growth and induced cytotoxicity of T24 cells. Cells were treated with various concentrations (0, 250, 500, 1000, and 2000 nM) of cucurbitacin E or 1%, DMSO as a control for 24 and 48 h. Percentage of the live cells in each group was determined by MTT method. Our data indicated that cucurbitacin E at the concentrations of 250–2000 nM inhibited cell growth in a concentration- and time-dependent manner (Figures 1(a) and 1(b)) with the half maximal inhibitory concentration (IC50) of 1012.32 ± 10.6 nM after a 48 h treatment.
Figure 1.
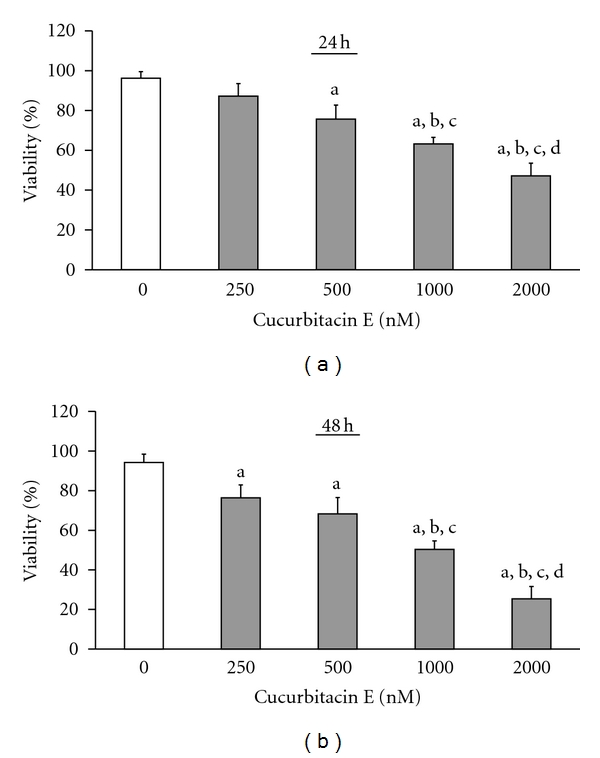
The chemical structure of cucurbitacin E and its effects of cell viability on human bladder cancer T24 cells. Cells were treated with different concentrations of cucurbitacin E for 24 h (a) and 48 h (b) and then cells were harvested for calculated the percentage of viable cells as described in the Materials and Methods. Data are presented as mean ± S.D. in triplicate. a, P ≤ 0.05, is significant different compared with the DMSO-treated control; b, c and d, P ≤ 0.05, are significant different with 250, 500 and 1000 nM of cucurbitacin E treatment, respectively, by one-way ANOVA followed by Bonferroni's test for multiple comparisons.
3.2. The Effects of Cucurbitacin E on DNA Content of T24 Cells
Cell-cycle distribution of T24 cells after treatment with 1000 nM cucurbitacin E for 12, 24, and 48 h was measured by flow cytometry. Results were shown that the profile from flow cytometry and BD CellQuest Pro software in the number of cells in G2/M phase was increased after treatment with 1000 nM of cucurbitacin E for 24 h (Figure 2(a)). The number of cells in G2/M phase was increased from 19.4% to 24.6%, 48.66% and 65.89% after 12, 24 and 48 h treatments, respectively and these effects are time-dependent responses (Figure 2(b)). These data suggest that the induction of G2/M phase arrest accounts for the growth inhibitory effects of cucurbitacin E-treated T24 cells.
Figure 2.
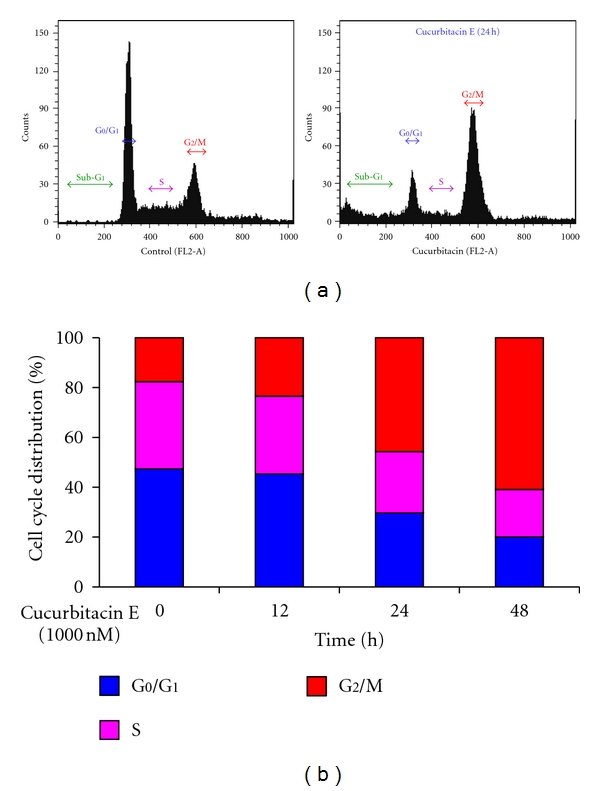
The effects of cucurbitacin E on the G2/M phase arrest in T24 cells. Cells at a density of 2 × 105 cells per well were placed in 12-well plates and then were treated with 1000 nM cucurbitacin E for different time periods (12, 24, or 48 h). Cells were harvested for evaluating the cell cycle distribution as described in the Materials and Methods. (a) The representative of profiles of DNA content; (b) the percentage of cells in G0/G1, S, and G2/M phase in T24 cells. Data revealed a representative experiment in triplicate with similar results.
3.3. The Effects of Cucurbitacin E on the Levels of STAT3, p53, and p21 as well as G2/M Phase-Associated Protein Levels in T24 Cells
To investigate the molecular mechanisms of cucurbitacin E-induced G2/M phase arrest in T24 cells, cucurbitacin E-treated cells determined the G2/M phase-modulated relative protein levels. Results from western blotting and CDK1 activity were shown in Figure 3. Our data indicated that the level of p53 was increased, and the level of p-STAT3 was decreased, but the level of STAT3 was not altered (Figure 3(a)) in cucurbitacin E-treated T24 cells. The level of p21 was upregulated, but the levels of CDK1 and cyclin B were downregulated, leading to induce G2/M phase arrest. The CDK1 activity also was inhibited by cucurbitacin E and this effect is a dose-dependent manner (Figure 3(c)). Based on these results, we suggest that cucurbitacin E-increased G2/M phase arrest in T24 cells might be involved in STAT3/p53/p21 signaling.
Figure 3.
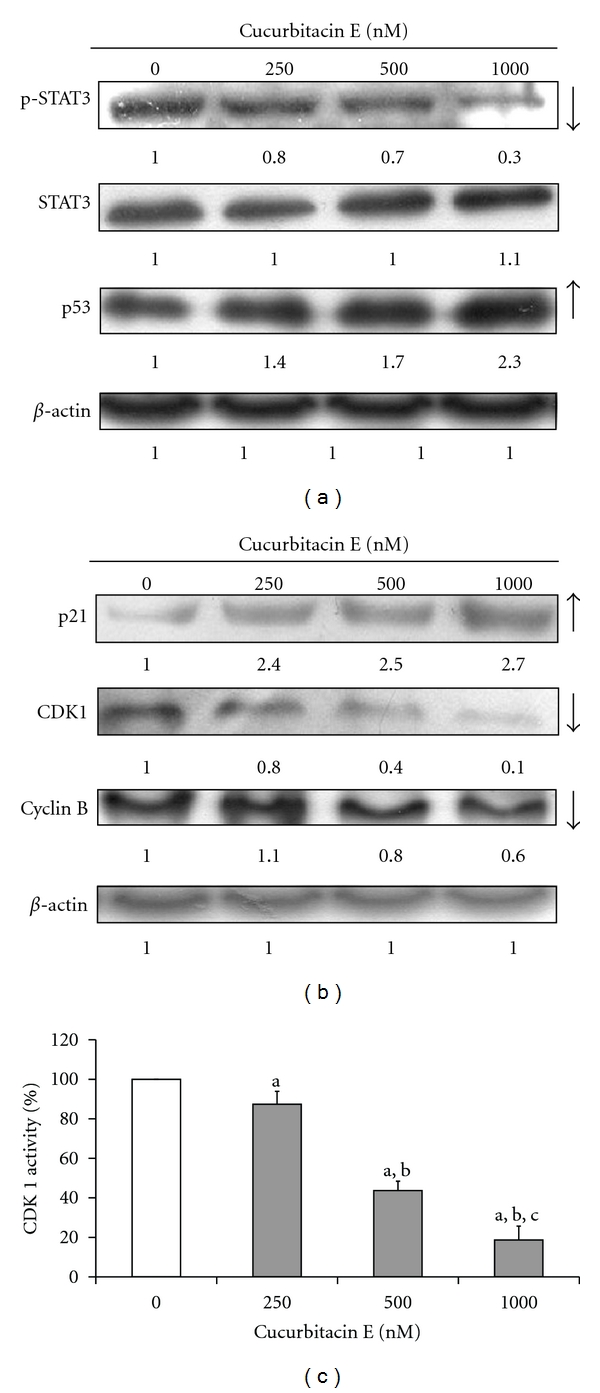
The effects of cucurbitacin E on the STAT3 phosphorylation, cell cycle-regulated associated proteins, and CDK1 activity in T24 cells. Cells at 1 × 106 (cells/well) were placed in 6-well plates and then treated without and with 250, 500, and 1000 nM of cucurbitacin E for 24 h. Thereafter, cells were harvested and total proteins were collected for western blot analysis to examine the changes of protein levels of p-STAT, STAT, and p53 (a), p21, CDK1, and cyclin B1 (b) as well as CDK1 activity (c) as described in Materials and Methods. Each value is mean ± S.D. of three experiments; a, P ≤ 0.05, is significantly different compared with control (0 nM); b and c, P ≤ 0.05, reveal significantly difference compared with 250 and 500 nM of cucurbitacin E, respectively (one-way ANOVA followed by Bonferroni's test for multiple comparisons).
3.4. The Effects of Cucurbitacin E on Morphological Observation and Apoptosis in T24 Cells
Cells were exposed to 0 (control), 500, and 1000 nM of cucurbitacin E for 48 h, and then examined and photographed by a phase-contrast microscope. Moreover, cells were treated with 1000 nM cucurbitacin E for 0, 12, 24, or 48 h and then assayed the percentage of sub-G1 phase by flow cytometry. The results shown in Figure 4(a) indicated that cucurbitacin E induced morphological changes such as membrane blebbing, reduction in cell volume, and these effects are dose-dependent manners (Figure 4(a)). Figure 4(b) also shows that cucurbitacin E induced sub-G1 phase (apoptosis) in cell cycle distribution that indicated that cucurbitacin E induced apoptosis, and this effect is a time-dependent manner. Apoptotic cells were also confirmed by Annexin V/PI staining assay in cucurbitacin E-treated T24 cells as seen in Figure 4(c).
Figure 4.
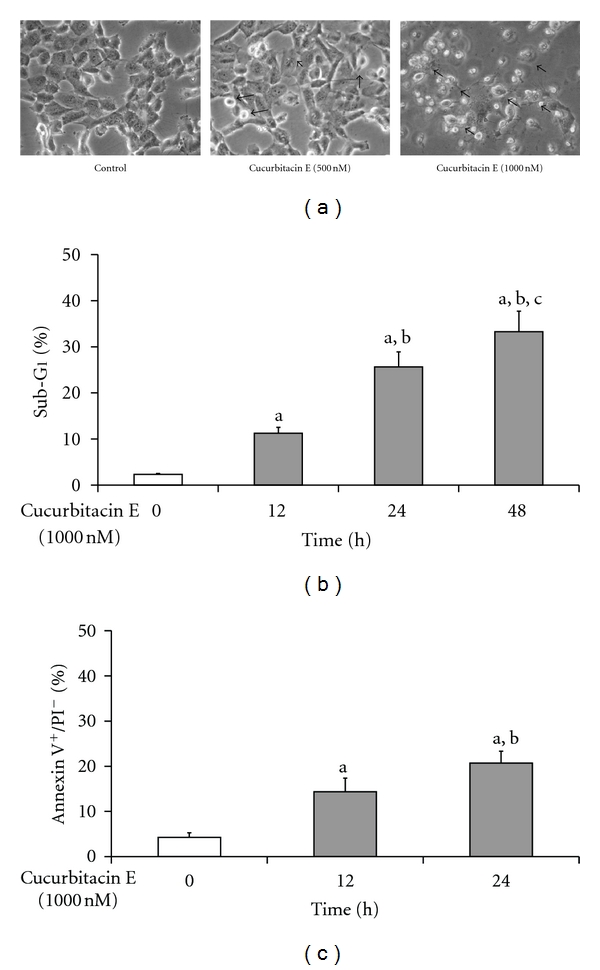
The effects of cucurbitacin E on the apoptotic death of T24 cells. Cells at a density of 2 × 105 cells/well were placed in 12-well plates then were treated with 500 and 1000 nM of cucurbitacin E for examining the morphological changes (a). Cells were incubated with 1000 nM cucurbitacin E for different time periods (12, 24, and 48 h), and then cells were harvested for evaluating the sub-G1 phase (apoptosis) by flow cytometry (b). Cells were exposed to 1000 nM cucurbitacin E for 12 and 24 h, and then cells were harvested for determining the annexin V positive/PI negative (apoptotic cells) as described in Materials and Methods. The results are shown as a mean ± SD (n = 3); a is significantly different (P ≤ 0.05) compared to control; b and c represent significant difference (P ≤ 0.05) compared with 1000 nM cucurbitacin E for 12 and 24 h, respectively, by one-way ANOVA followed by Bonferroni's test for multiple comparisons.
3.5. The Effects of Cucurbitacin E on Protein Levels of Caspase-3, -8, and -9 in T24 Cells
To investigate if cucurbitacin E induces apoptosis in T24 cells through caspases-dependent pathway, cells were pretreated with a general caspase inhibitor (Z-VAD-FMK) and then exposed to cucurbitacin E for 48 h treatment. Results indicate that Z-VAD-FMK is able to protect against cucurbitacin E-reduced cell viability in T24 cells. Also, our results from western blotting indicated that the levels of active form of caspase-3, -8, and -9 were stimulated in cucurbitacin E-treated T24 cells (Figure 5(b)). This study suggests that cucurbitacin E-induced apoptosis of T24 cells is mediated via caspase cascade-dependent pathway.
Figure 5.
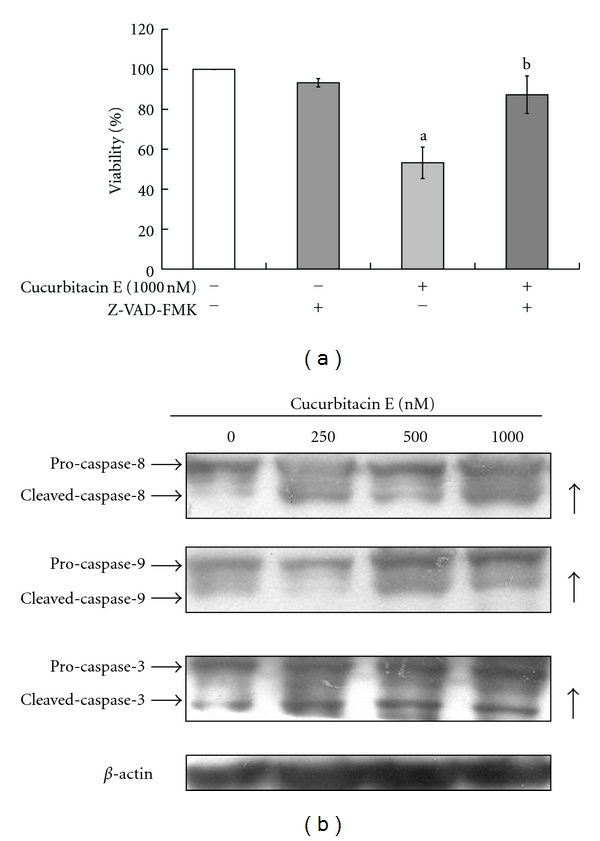
The effects of a general caspase inhibitor (Z-VAD-FMK) on cucurbitacin E-induced cell viability and caspase cascades protein levels on T24 cells. Cells were pretreated with 10 μM Z-VAD-FMK and then exposed to 1000 nM cucurbitacin E for 48 h. Cells were harvested and determined for percentage of viable cells (a) and for evaluating the protein levels of caspase-8, -9, and -3 (b) as described in Materials and Methods. Columns: mean (n = 3); bars: SD. a, P ≤ 0.05, is significantly different compared with untreated control; b, P ≤ 0.05, shows significant difference compared with 1000 nM cucurbitacin E treatment group by one-way ANOVA followed by Bonferroni's multiple comparison test.
3.6. The Effects of Cucurbitacin E on Mitochondrial Membrane Potential (ΔΨm) of T24 Cells
To investigate whether mitochondrial dysfunction is involved in cell apoptosis induced by cucurbitacin E in T24 cells, cells were exposed to cucurbitacin E for 0, 4, 8, 12, 24, and 48 h and subsequently harvested for determination of the level of ΔΨm by flow cytometry. Results shown in Figures 6(a) and 6(b) indicated that cucurbitacin E decreased the level of ΔΨm and this effect is a time-dependent manner.
Figure 6.
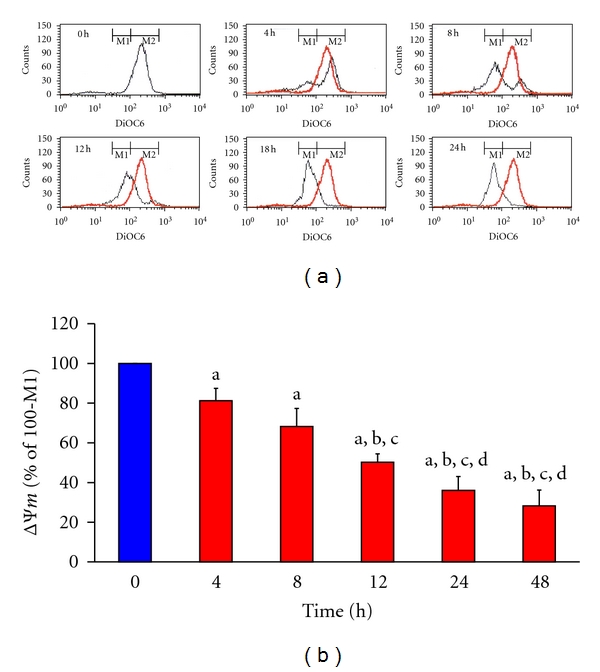
The effects of cucurbitacin E on mitochondrial membrane potential (ΔΨm) in T24 cells. Cells at a density of 2 × 105 cells/well placed in 12-well plates were treated with 1000 nM cucurbitacin E for 0, 4, 8, 12, 24, and 48 h and then harvested for measuring the level of ΔΨm as described in Materials and Methods. (a) The representative of profile of ΔΨm, and (b) the levels of ΔΨm quantified form BD CellQuest Pro software. Each experiment was done with triple sets: a, P ≤ 0.05, shows significant difference compared with 0 h treatment; b, c, and d, P ≤ 0.05, are significantly different compared to cucurbitacin E treatment for 4, 8, and 12 h, respectively, by one-way ANOVA followed by Bonferroni's multiple comparison test.
3.7. The Effects of Cucurbitacin E on Apoptosis-Associated Protein Levels in T24 Cells
It is well known that mitochondria-mediated activation of caspase-9 and caspase-3 involved the releases of cytochrome c and AIF [37, 38]. The treatment with cucurbitacin E increased the levels of cytochrome c, Apaf-1, AIF, truncated BID (Figure 7(a)), and Fas (Figure 7(b)) in cucurbitacin E-treated T24 cells. These data suggest that the increased levels of Fas/CD95, cytochrome c, Apaf-1, and truncated BID led to the activation of caspase-8, caspase-9, and caspase-3 (Figure 5(b)). Hence, cucurbitacin E-provoked apoptosis in T24 cells is mediated through the Fas/CD95 and mitochondria-dependent pathways.
Figure 7.

The effects of cucurbitacin E on the apoptosis-associated proteins in T24 cells. Cells (1 × 106/well) seeded into 6-well plates were treated with 1000 nM cucurbitacin and were treated with different concentrations of cucurbitacin E for 48 h and then harvested for western blotting to examine the protein levels of cytochrome c, Apaf-1, BID, and truncated BID (a), Fas and FasL (b) as described in Materials and Methods.
4. Discussion
It is well known that the resistant to current therapies is one of the poor prognostic outcomes of human bladder cancer and it is the leading cause of cancer-related death [39, 40]. Currently, the successful treatment with chemotherapeutic agents is based on these agents to trigger cell death in tumor cells [41–43]. Until now, investigators have been focusing on the novel inducers of apoptosis to provide a new therapeutic approach for cancer patients and some of the compounds for treatment of cancer are from natural products [44–46].
Results showed that cucurbitacin E induced G2/M phase arrest (Figure 2) in T24 cells. The protein levels of phospho-STAT3, CDK1, and cyclin B were decreased, and the levels of p53 and p21 were increased in cucurbitacin E-treated T24 cells in a time-dependent manner (Figures 3(a) and 3(b)). Furthermore, curcumin E also inhibited the CDK1 activity in T24 cells (Figure 3(c)). It is reported that activated STAT3 is formed through downregulating p53 level and affects the p53 promoter in in vitro and in vivo studies [47]. Moreover, CDKs regulate the cell-cycle progression in mammalian cells [48, 49]. In the G2/M phase progression which is regulated with CDK1 and CDK2 kinases that are activated primarily in association with cyclins A and B [50]; it is reported that distributing cell-cycle progression by alterations in cell cycle-related protein expression plays important roles in the proliferation of cancer cells [50, 51]. Therefore, we suggest that alterations in STAT3/p21/p53 signaling and G2/M phase-associated protein levels occurred in cucurbitacin E-treated T24 cells. This is also in agreement with other reports addressing that inhibitory effect of STAT3 level on cucurbitacin E and triterpene-derived natural products treated human tumor cells and umbilical vascular endothelial cells in vitro [9, 11, 51].
Our results also showed that cucurbitacin E induced apoptotic death of T24 cells and this effect is a dose- and time-dependent response (Figure 4). Apoptosis is a programmed cell death which is a multiple regulated process which can be divided into the caspases and mitochondrial signaling pathways [52, 53]. Cucurbitacin E treatment in T24 cells promoted the activations of caspase-8, -9, and -3 in a dose-dependent manner (Figure 5(b)). Otherwise, cells were pretreated with a general caspase inhibitor (Z-VAD-FMK) and exposed to cucurbitacin E, leading to increase the percentage of viable cells when compared to the cucurbitacin E-treated only cells (Figure 5(a)). The data indicated that the activation of caspase cascade is involved in cucurbitacin E-induced cell death
Result from Figures 6(a) and 6(b) indicated that cucurbitacin E decreased the level of ΔΨm in a time-dependent response. Bcl-2 family proteins (antiapoptotic and proapoptotic proteins) have been reported to regulate cytochrome c release from mitochondria [53, 54]. Our results also showed that cucurbitacin E promoted the level of cytochrome c (released from mitochondria) (Figure 7(a)). Cucurbitacin increased the levels of truncated BID which is related to the dysfunction of mitochondria (Figure 7(a)). Figure 7(a) also shows an increase in the level of AIF which is released from mitochondria. Based on these observations, we may suggest that cucurbitacin-induced apoptosis of T24 cell is carried out through the Fas/CD95 and mitochondrial signaling pathways.
In conclusion, cucurbitacin E decreased the percentage of viable T24 cells through the cell cycle arrest and induction of apoptosis in human bladder T24 cancer cells in vitro. Cucurbitacin E-induced G2/M phase arrest was associated with the inhibitions of phosphorylation STAT3, promotion of p53 and p21, and reduction of CDK1 and cyclin B. Cucurbitacin E-induced apoptosis was accompanied with upregulation of Fas/CD95, decrease the level of ΔΨm and then led to cytochrome c release from mitochondria and promoted the activations caspase-8, caspase-9, and caspase-3, leading to apoptosis of T24 cells. The proposed signaling pathways can be seen in Figure 8.
Figure 8.
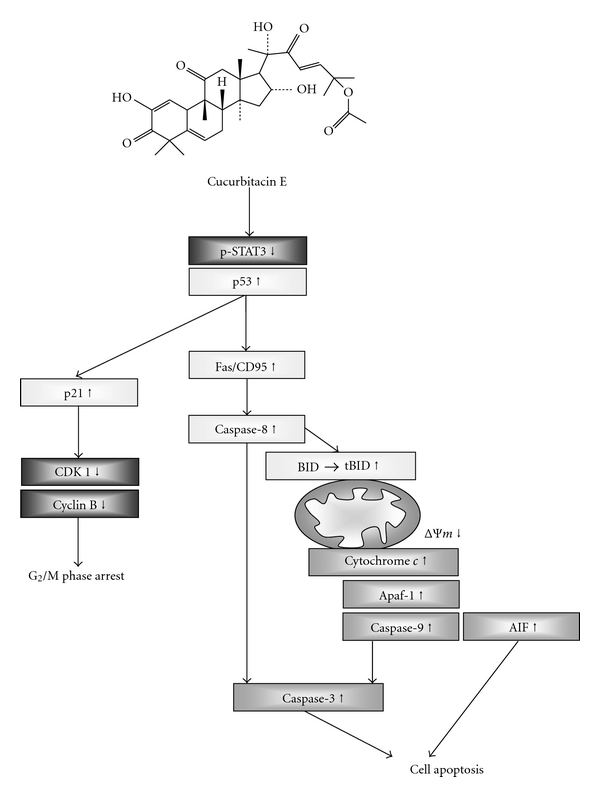
The possible signaling pathways for cucurbitacin E-induced G2/M phase arrest through STAT3/p53/p21 signaling and apoptosis via Fas/CD95 and mitochondria-dependent pathways in human bladder cancer T24 cells.
Acknowledgments
This study was supported by the Grants CMU99-COL-27-1 and CMU99-COL-27-2 from China Medical University, Taichung, Taiwan. The authors also thank the National Science Council of the Republic of China for financial support (NSC 97–2320-B-039-004-MY3).
References
- 1.Kumar B, Koul S, Petersen J, et al. p38 Mitogen-activated protein kinase-driven MAPKAPK2 regulates invasion of bladder cancer by modulation of MMP-2 and MMP-9 activity. Cancer Research. 2010;70(2):832–841. doi: 10.1158/0008-5472.CAN-09-2918. [DOI] [PubMed] [Google Scholar]
- 2.Zapatero A, Martin de Vidales C, Arellano R, Bocardo G, Pérez M, Ríos P. Updated results of bladder-sparing trimodality approach for invasive bladder cancer. Urologic Oncology. 2010;28(4):368–374. doi: 10.1016/j.urolonc.2009.01.031. [DOI] [PubMed] [Google Scholar]
- 3.Matés JM, Segura JA, Alonso FJ, Márquez J. Anticancer antioxidant regulatory functions of phytochemicals. Current Medicinal Chemistry. 2011;18(15):2315–2338. doi: 10.2174/092986711795656036. [DOI] [PubMed] [Google Scholar]
- 4.Neto CC, Amoroso JW, Liberty AM. Anticancer activities of cranberry phytochemicals: an update. Molecular Nutrition and Food Research. 2008;52(1):S18–S27. doi: 10.1002/mnfr.200700433. [DOI] [PubMed] [Google Scholar]
- 5.Newman DJ, Cragg GM, Holbeck S, Sausville EA. Natural products and derivatives as leads to cell cycle pathway targets in cancer chemotherapy. Current Cancer Drug Targets. 2002;2(4):279–308. doi: 10.2174/1568009023333791. [DOI] [PubMed] [Google Scholar]
- 6.Attard E, Brincat MP, Cuschieri A. Immunomodulatory activity of cucurbitacin E isolated from Ecballium elaterium. Fitoterapia. 2005;76(5):439–441. doi: 10.1016/j.fitote.2005.02.007. [DOI] [PubMed] [Google Scholar]
- 7.Gilbert JN, Mathieson DW. The functional groupings of cucurbitacin E (alpha-elaterin) The Journal of Pharmacy and Pharmacology. 1958;10(supplement):252T–255T. doi: 10.1111/j.2042-7158.1958.tb10408.x. [DOI] [PubMed] [Google Scholar]
- 8.Momma K, Masuzawa Y, Nakai N, et al. Direct interaction of Cucurbitacin E isolated from Alsomitra macrocarpa to actin filament. Cytotechnology. 2008;56(1):33–39. doi: 10.1007/s10616-007-9100-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Dong Y, Lu B, Zhang X, et al. Cucurbitacin E, a tetracyclic triterpenes compound from Chinese medicine, inhibits tumor angiogenesis through VEGFR2-mediated Jak2-STAT3 signaling pathway. Carcinogenesis. 2010;31(12):2097–2104. doi: 10.1093/carcin/bgq167. [DOI] [PubMed] [Google Scholar]
- 10.Tannin-Spitz T, Bergman M, Grossman S. Cucurbitacin glucosides: antioxidant and free-radical scavenging activities. Biochemical and Biophysical Research Communications. 2007;364(1):181–186. doi: 10.1016/j.bbrc.2007.09.075. [DOI] [PubMed] [Google Scholar]
- 11.Shan X-L, Zhou X-Y, Yang J, Wang Y-L, Deng Y-H, Zhang M-X. Inhibitory effect and mechanism of cucurbitacin E on the proliferation of ovarian cancer cells. Chinese Journal of Cancer. 2010;29(1):20–23. doi: 10.5732/cjc.009.10223. [DOI] [PubMed] [Google Scholar]
- 12.Sadzuka Y, Hatakeyama H, Sonobe T. Enhancement of doxorubicin concentration in the M5076 ovarian sarcoma cells by cucurbitacin E co-treatment. International Journal of Pharmaceutics. 2010;383(1-2):186–191. doi: 10.1016/j.ijpharm.2009.08.040. [DOI] [PubMed] [Google Scholar]
- 13.Nakashima S, Matsuda H, Kurume A, et al. Cucurbitacin E as a new inhibitor of cofilin phosphorylation in human leukemia U937 cells. Bioorganic and Medicinal Chemistry Letters. 2010;20(9):2994–2997. doi: 10.1016/j.bmcl.2010.02.062. [DOI] [PubMed] [Google Scholar]
- 14.Li Y, Wang R, Ma E, et al. The induction of G2/M cell-cycle arrest and apoptosis by cucurbitacin e is associated with increased phosphorylation of eIF2α in leukemia cells. Anti-Cancer Drugs. 2010;21(4):389–400. doi: 10.1097/CAD.0b013e328336b383. [DOI] [PubMed] [Google Scholar]
- 15.Sun C, Zhang M, Shan X, et al. Inhibitory effect of cucurbitacin e on pancreatic cancer cells growth via STAT3 signaling. Journal of Cancer Research and Clinical Oncology. 2010;136(4):603–610. doi: 10.1007/s00432-009-0698-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kidd VJ. Proteolytic activities that mediate apoptosis. Annual Review of Physiology. 1998;60:533–573. doi: 10.1146/annurev.physiol.60.1.533. [DOI] [PubMed] [Google Scholar]
- 17.Kaufmann SH. Induction of endonucleolytic DNA cleavage in human acute myelogenous leukemia cells by etoposide, camptothecin, and other cytotoxic anticancer drugs: a cautionary note. Cancer Research. 1989;49(21):5870–5878. [PubMed] [Google Scholar]
- 18.Friesen C, Herr I, Krammer PH, Debatin KM. Involvement of the CD95 (APO-1/Fas) receptor/ligand system in drug-induced apoptosis in leukemia cells. Nature Medicine. 1996;2(5):574–577. doi: 10.1038/nm0596-574. [DOI] [PubMed] [Google Scholar]
- 19.Fulda S, Debatin KM. Death receptor signaling in cancer therapy. Current Medicinal Chemistry. 2003;3(4):253–262. doi: 10.2174/1568011033482404. [DOI] [PubMed] [Google Scholar]
- 20.Ramnath N, Khushalani N, Toth K, et al. S-phase modulation by irinotecan: pilot studies in advanced solid tumors. Cancer Chemotherapy and Pharmacology. 2005;56(5):447–454. doi: 10.1007/s00280-004-0951-6. [DOI] [PubMed] [Google Scholar]
- 21.Orren DK, Petersen LN, Bohr VA. Persistent DNA damage inhibits S-phase and G2 progression, and results in apoptosis. Molecular Biology of the Cell. 1997;8(6):1129–1142. doi: 10.1091/mbc.8.6.1129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Fujimoto K, Hosotani R, Doi R, et al. Induction of cell-cycle arrest and apoptosis by a novel retinobenzoic- acid derivative, TAC-101, in human pancreatic-cancer cells. International Journal of Cancer. 1999;81(4):637–644. doi: 10.1002/(sici)1097-0215(19990517)81:4<637::aid-ijc21>3.0.co;2-4. [DOI] [PubMed] [Google Scholar]
- 23.Gamet-Payrastre L, Li P, Lumeau S, et al. Sulforaphane, a naturally occurring isothiocyanate, induces cell cycle arrest and apoptosis in HT29 human colon cancer cells. Cancer Research. 2000;60(5):1426–1433. [PubMed] [Google Scholar]
- 24.Lin JG, Chen GW, Li TM, Chouh ST, Tan TW, Chung JG. Aloe-Emodin induces apoptosis in T24 human bladder cancer cells through the p53 dependent apoptotic pathway. Journal of Urology. 2006;175(1):343–347. doi: 10.1016/S0022-5347(05)00005-4. [DOI] [PubMed] [Google Scholar]
- 25.Wu P-P, Liu K-C, Huang W-W, et al. Triptolide induces apoptosis in human adrenal cancer NCI-H295 cells through a mitochondrial-dependent pathway. Oncology Reports. 2011;25(2):551–557. doi: 10.3892/or.2010.1080. [DOI] [PubMed] [Google Scholar]
- 26.Huang W-W, Ko S-W, Tsai H-Y, et al. Cantharidin induces G2/M phase arrest and apoptosis in human colorectal cancer colo 205 cells through inhibition of CDK1 activity and caspase-dependent signaling pathways. International Journal of Oncology. 2011;38(4):1067–1073. doi: 10.3892/ijo.2011.922. [DOI] [PubMed] [Google Scholar]
- 27.Wu SH, Hang LW, Yang JS, et al. Curcumin induces apoptosis in human non-small cell lung cancer NCI-H460 cells through ER stress and caspase cascade- and mitochondria-dependent pathways. Anticancer Research. 2010;30(6):2125–2133. [PubMed] [Google Scholar]
- 28.Yang JS, Hour MJ, Huang WW, Lin KL, Kuo SC, Chung JG. MJ-29 inhibits tubulin polymerization, induces mitotic arrest, and triggers apoptosis via cyclin-dependent kinase 1-mediated Bcl-2 phosphorylation in human leukemia U937 cells. Journal of Pharmacology and Experimental Therapeutics. 2010;334(2):477–488. doi: 10.1124/jpet.109.165415. [DOI] [PubMed] [Google Scholar]
- 29.Lu CC, Yang JS, Huang AC, et al. Chrysophanol induces necrosis through the production of ROS and alteration of ATP levels in J5 human liver cancer cells. Molecular Nutrition and Food Research. 2010;54(7):967–976. doi: 10.1002/mnfr.200900265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Chou LC, Yang JS, Huang LJ, et al. The synthesized 2-(2-fluorophenyl)-6,7-methylenedioxyquinolin-4-one (CHM-1) promoted G2/M arrest through inhibition of CDK1 and induced apoptosis through the mitochondrial-dependent pathway in CT-26 murine colorectal adenocarcinoma cells. Journal of Gastroenterology. 2009;44(10):1055–1063. doi: 10.1007/s00535-009-0111-1. [DOI] [PubMed] [Google Scholar]
- 31.Lin CC, Yang JS, Chen JT, et al. Berberine induces apoptosis in human HSC-3 oral cancer cells via simultaneous activation of the death receptor-mediated and mitochondrial pathway. Anticancer Research. 2007;27(5 A):3371–3378. [PubMed] [Google Scholar]
- 32.Chiang J-H, Yang J-S, Ma C-Y, et al. Danthron, an anthraquinone derivative, induces DNA damage and caspase cascades-mediated apoptosis in SNU-1 human gastric cancer cells through mitochondrial permeability transition pores and Bax-triggered pathways. Chemical Research in Toxicology. 2011;24(1):20–29. doi: 10.1021/tx100248s. [DOI] [PubMed] [Google Scholar]
- 33.Liu K-C, Huang A-C, Wu P-P, et al. Gallic acid suppresses the migration and invasion of PC-3 human prostate cancer cells via inhibition of matrix metalloproteinase-2 and -9 signaling pathways. Oncology Reports. 2011;26(1):177–184. doi: 10.3892/or.2011.1264. [DOI] [PubMed] [Google Scholar]
- 34.Lan YU-H, Wu Y-C, Wu K-W, et al. Death receptor 5-mediated TNFR family signaling pathways modulate γ-humulene-induced apoptosis in human colorectal cancer HT29 cells. Oncology Reports. 2011;25(2):419–424. doi: 10.3892/or.2010.1087. [DOI] [PubMed] [Google Scholar]
- 35.Lai WW, Yang JS, Lai KC, et al. Rhein induced apoptosis through the endoplasmic reticulum stress, caspase- and mitochondria-dependent pathways in SCC-4 human tongue squamous cancer cells. In Vivo. 2009;23(2):309–316. [PubMed] [Google Scholar]
- 36.Huang WW, Chiu YJ, Fan MJ, et al. Kaempferol induced apoptosis via endoplasmic reticulum stress and mitochondria-dependent pathway in human osteosarcoma U-2 OS cells. Molecular Nutrition and Food Research. 2010;54(11):1585–1595. doi: 10.1002/mnfr.201000005. [DOI] [PubMed] [Google Scholar]
- 37.Antonsson B. Mitochondria and the Bcl-2 proteins in apoptosis signaling pathways. Molecular and Cellular Biochemistry. 2004;256-257(1-2):141–155. doi: 10.1023/b:mcbi.0000009865.70898.36. [DOI] [PubMed] [Google Scholar]
- 38.Kuwana T, Newmeyer DD. Bcl-2-family proteins and the role of mitochondria in apoptosis. Current Opinion in Cell Biology. 2003;15(6):691–699. doi: 10.1016/j.ceb.2003.10.004. [DOI] [PubMed] [Google Scholar]
- 39.Forster JA, Paul AB, Harnden P, Knowles MA. Expression of NRG1 and its receptors in human bladder cancer. British Journal of Cancer. 2011;104(7):1135–1143. doi: 10.1038/bjc.2011.39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Gopalkrishna V, Srivastava AN, Hedau S, Sharma JK, Das BC. Detection of human papillomavirus DNA sequences in cancer of the urinary bladder by in situ hybridisation and polymerase chain reaction. Genitourinary Medicine. 1995;71(4):231–233. doi: 10.1136/sti.71.4.231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Filion MC, Lépicier P, Morales A, Phillips NC. Mycobacterium phlei cell wall complex directly induces apoptosis in human bladder cancer cells. British Journal of Cancer. 1999;79(2):229–235. doi: 10.1038/sj.bjc.6690038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Konstantakou EG, Voutsinas GE, Karkoulis PK, Aravantinos G, Margaritis LH, Stravopodis DJ. Human bladder cancer cells undergo cisplatin-induced apoptosis that is associated with p53-dependent and p53-independent responses. International Journal of Oncology. 2009;35(2):401–416. [PubMed] [Google Scholar]
- 43.Li X, Su B, Liu R, Wu D, He D. Tetrandrine induces apoptosis and triggers caspase cascade in human bladder cancer cells. Journal of Surgical Research. 2011;166(1):e45–e51. doi: 10.1016/j.jss.2010.10.034. [DOI] [PubMed] [Google Scholar]
- 44.Shrader M, Pino MS, Lashinger L, et al. Gefitinib reverses TRAIL resistance in human bladder cancer cell lines via inhibition of AKT-mediated X-linked inhibitor of apoptosis protein expression. Cancer Research. 2007;67(4):1430–1435. doi: 10.1158/0008-5472.CAN-06-1224. [DOI] [PubMed] [Google Scholar]
- 45.Kelly K. The role of targeted agents in adjuvant therapy for non-small cell lung cancer. Clinical Cancer Research. 2005;11(13):5027s–5029s. doi: 10.1158/1078-0432.CCR-05-9002. [DOI] [PubMed] [Google Scholar]
- 46.Gitter S, Gallily R, Shohat B, Lavie D. Studies on the antitumor effect of cucurbitacins. Cancer research. 1961;21:516–521. [PubMed] [Google Scholar]
- 47.Niu G, Wright KL, Ma Y, et al. Role of Stat3 in regulating p53 expression and function. Molecular and Cellular Biology. 2005;25(17):7432–7440. doi: 10.1128/MCB.25.17.7432-7440.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Hartwell LH, Weinert TA. Checkpoints: controls that ensure the order of cell cycle events. Science. 1989;246(4930):629–634. doi: 10.1126/science.2683079. [DOI] [PubMed] [Google Scholar]
- 49.Molinari M. Cell cycle checkpoints and their inactivation in human cancer. Cell Proliferation. 2000;33(5):261–274. doi: 10.1046/j.1365-2184.2000.00191.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.O’Connor PM, Ferris DK, Pagano M, et al. G2 delay induced by nitrogen mustard in human cells affects cyclin A/cdk2 and cyclin B1/cdc2-kinase complexes differently. Journal of Biological Chemistry. 1993;268(11):8298–8308. [PubMed] [Google Scholar]
- 51.Boykin C, Zhang G, Chen Y-H, et al. Cucurbitacin IIa: a novel class of anti-cancer drug inducing non-reversible actin aggregation and inhibiting survivin independent of JAK2/STAT3 phosphorylation. British Journal of Cancer. 2011;104(5):781–789. doi: 10.1038/bjc.2011.10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Salvesen GS, Dixit VM. Caspases: intracellular signaling by proteolysis. Cell. 1997;91(4):443–446. doi: 10.1016/s0092-8674(00)80430-4. [DOI] [PubMed] [Google Scholar]
- 53.Gross A, McDonnell JM, Korsmeyer SJ. BCL-2 family members and the mitochondria in apoptosis. Genes and Development. 1999;13(15):1899–1911. doi: 10.1101/gad.13.15.1899. [DOI] [PubMed] [Google Scholar]
- 54.Kuo JH, Chu YL, Yang JS, et al. Cantharidin induces apoptosis in human bladder cancer TSGH 8301 cells through mitochondria-dependent signal pathways. International Journal of Oncology. 2010;37(5):1243–1250. doi: 10.3892/ijo_00000775. [DOI] [PubMed] [Google Scholar]