

Abstract
Metastatic breast cancer is a life-threatening stage of cancer and is the leading cause of death in advanced breast cancer patients. Estrogen signaling and the estrogen receptor (ER) are implicated in breast cancer progression, and the majority of the human breast cancers start out as estrogen dependent. Accumulating evidence suggests that ER signaling is complex, involving coregulatory proteins and extranuclear actions. ER-coregualtory proteins are tightly regulated under normal conditions with miss expression primarily reported in cancer. Deregulation of ER coregualtors or ER extranuclear signaling has potential to promote metastasis in ER-positive breast cancer cells. This review summarizes the emerging role of ER signaling in promoting metastasis of breast cancer cells, discusses the molecular mechanisms by which ER signaling contributes to metastasis, and explores possible therapeutic targets to block ER-driven metastasis.
1. Introduction
The steroid hormone, estradiol, plays an important role in the progression of breast cancer, and a majority of the human breast cancers start out as estrogen dependent and express the estrogen receptor (ER). The biological effects of estrogen are mediated by its binding to one of the structurally and functionally distinct ERs (ERα and ERβ) [1]. Endocrine therapy using Tamoxifen, a selective estrogen receptor modulator [2], and aromatase inhibitors, which ablate peripheral estrogen synthesis, has been shown to substantially improve disease-free survival [3]. Endocrine therapy has also been shown to have a positive effect on the treatment of ER-positive breast cancer [4]. Despite these positive effects, initial or acquired resistance to endocrine therapies frequently occurs with tumors recurring as metastatic. Tumor metastasis comprises a series of discrete biological processes that moves tumor cells from the primary neoplasm to a distant location [5] and involves a multi-step cascade of coordinated cell adhesion and contractility as well as proteolytic remodeling of the extracellular matrix (ECM) [6, 7]. Even though substantial information is available on the process of metastasis, the molecular basis of breast cancer progression to metastasis and the role of ERα signaling in this process remain poorly understood. A few early studies suggested a negative effect of ERα signaling on motility and invasion of cells [8, 9], while several recent studies showed a positive effect of ER signaling on motility [10–14]. In this review, we summarized the emerging evidence for the role of ERα signaling in breast cancer progression to metastasis and discuss the possibility of targeting ERα signaling crosstalk with cytosolic kinases as a possible additional therapeutic target for treating/preventing ER-positive metastatic breast cancer.
2. ERα Signaling Mechanisms
ERα is the major ER subtype in the mammary epithelium and plays a critical role in mammary gland biology as well as in breast cancer progression [15, 16]. The ERα comprises an N-terminal AF1 domain, a DNA-binding domain, and a C-terminal ligand-binding region that contains an AF2 domain [17]. Upon the binding of estrogen to ERα, the ligand-activated ERα translocates to the nucleus, binds to the responsive element in the target gene promoter, and stimulates gene transcription (genomic/nuclear signaling) [18, 19]. Emerging evidence suggests that ER signaling is complex, involving coregulatory proteins and also genomic actions and extranuclear actions [20, 21].
Multiprotein complexes containing coregulators assemble in response to hormone binding and activate ER-mediated transcription [18]. The ERα transcriptional outcome is regulated by dynamic chromatin modifications of the histone tails, and the ligand-bound ERα facilitates these modifications via coregulator recruitment [22]. For example, coactivators like SRC-1, amplified in breast cancer (AIB1), and CBP have been shown to possess histone acetyltransferase activity, whereas corepressors, such as NCOR and MTA1, are associated with histone deacetylases [20, 23]. It is generally accepted that some of the diverse functions of E2 depend on differential recruitment of coregulators to the E2-ER complex [24]. Even though coregulators modulate ER functions, each coregulator protein appears to play an important but not overlapping function in vivo [25–27].
Emerging findings suggest that ER-coregulatory proteins have potential to be differentially expressed in malignant tumors and that their functions may be altered, leading to tumor progression [28]. In vivo studies using wild type (WT) and SRC3/AIB1−/− mice harboring the mouse mammary tumor virus-polyomavirus middle T (PyMT) transgene (Tg) revealed that AIB1 knock down significantly reduces lung metastasis but not mammary tumorigenesis. Compared with WT/PyMT mice, Tg SRC-1−/−/PyMT mice had intravasation of mammary tumor cells. In addition, the frequency and extent of lung metastasis were drastically lower in the Tg mice than in the WT mice [29]. Another study using Tg SRC-1−/− mice reported that deficiency of SRC-1 coregulator increases MMTV-neu-mediated tumor latency and differentiation-specific gene expression and decreases metastasis [30]. Collectively, these emerging findings implicate the role of the ERα-coregulator-associated activities/functions in breast cancer metastasis.
3. ERα Genomic Actions and Metastasis
Within the last decade, research has provided substantial data to suggest that alteration in cellular concentration or genetic dysfunction of coregulators can contribute to a pathologic outcome by modulating ER genomic actions and has potential to drive cancer cell proliferation and metastasis [31]. Loss of the epithelial adhesion molecule E-cadherin is implicated with a critical role in metastasis by disrupting intercellular contacts, an early step in metastatic dissemination [32]. Functional or transcriptional loss is commonly associated with an invasive and poorly differentiated phenotype [33]. Deregulation of ER-coregulator signaling can lead to aberrant expression of Snail, resulting in the loss of expression of E-cadherin and invasive growth. For example, MTA1, a commonly deregulated coregulator in breast cancer, promotes transcriptional repression of ER, leading to metastatic progression [34]. The ERα coregulator (AIB1) amplified in breast cancer has been shown to promote breast cancer metastasis by activation of PEA3-mediated matrix metalloproteinase 2 (MMP2) and MMP9 expression [35]. SRC-1, another ER coregulator, has also been shown to promote breast cancer invasiveness and metastasis by coactivating PEA3-mediated Twist expression [36]. Recent studies have found deregulation of the ER coregulator PELP1 in invasive and metastatic breast tumors [37, 38]. Recent studies using PELP1 overexpression and knockdown demonstrated that PELP1 plays an important role in ERα-positive metastasis [10]. Collectively, these studies indicate that ERα and ER coregulators modulate expression of genes involved in metastasis.
4. ERα Extranuclear Actions and Metastasis
Emerging evidence suggests that the ERα participates in extranuclear signaling [39]. ERα activation, by E2, induces key features of motile cells including rapid cytoskeletal reorganization and the development of specialized structures including fillopodia and ruffles [37]. To establish the role of E2-mediated extranuclear actions, researchers developed E2-Dendrimers (EDCs), which are nanoparticles coated with estrogen. These EDCs uniquely localize in the membrane and cytoplasm, preferably activating ERα extranuclear signaling. Using these EDCs, researchers have demonstrated that ERα extranuclear pathways have distinct biological outcomes [40]. Our laboratory using EDCs provided further evidence that ERα extranuclear signaling has the potential to contribute to the breast cancer cell motility (Figure 1) [10]. ERα extranuclear signaling promotes stimulation of the Src kinase, mitogen-activated protein kinase (MAPK), phosphatidylinositol 3-kinase (PI3K), and protein kinase C pathways in the cytosol (10, 11). Recent studies identified PELP1 as one of the components of the ERα signalosome in the cytoplasm, and estrogen-mediated extranuclear signaling promotes cytoskeleton reorganization via ER-Src-PELP1-PI3K-ILK1 pathway [10]. Many of the kinases activated by ERα extra-nuclear signaling are implicated in breast cancer metastasis. For example, ERK and protein kinase B (AKT) phosphorylation play important roles in breast cancer cell migration [14], and Src and ILK1 kinases play critical roles in invasion and metastasis of breast cancer cells [41, 42].
Figure 1.
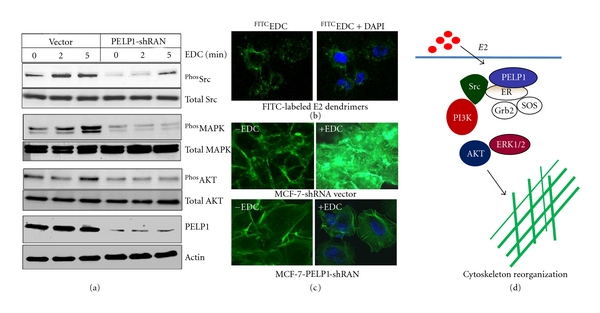
ER-extranuclear signaling promotes actin reorganization via ER coregulator PELP1. (a) MCF7 shRNA vector control and MCF7-PELP1-shRNA cells were cultured in 5% DCC serum containing medium treated with or without estrogen dendrimers (EDC). The activation of signaling pathways was analyzed by Western blotting of total protein lysates with phospho-specific antibodies. (b) MCF7 cells were treated with FITC-labeled EDC and localization of EDC was analyzed by confocal microscopy. Green; EDC; Blue, DAPI. (c) MCF7 or MCF7-PELP1-shRNA cells were treated either with E2 or EDC and the F-actin status was analyzed by phalloidin staining and visualized by confocal microscopy. (d) Schematic representation of estrogen-mediated extranuclear signaling. Adapted from [10].
In addition to ERα interactions with cytosolic kinases, few other mechanisms by which the ERα activates extranuclear signaling have been reported. Membrane-bound ERα has been reported to be associated with growth factor receptors such as IGF-1R, EGFR, and HER2; such interactions play a role in cytoskeleton reorganization [43]. Dysregulation of HER2 in breast cancer cells enhances the expression of an isoform of MTA1 (MTA1s), which promotes the cytoplasmic sequestration of ERα leading to constitutive activation of MAPK. These study findings implicate the regulation of the cellular localization of ERα by MTA1s as a mechanism for enhancing ERα extranuclear actions by nuclear exclusion [44]. Recent studies also found that the ERα was methylated via posttranslational modifications, and methylated ERα was predominantly present in the cytoplasm, suggesting that deregulation of arginine methylases may have consequences in activation of ERα extranuclear actions [45]. Collectively, these emerging results suggest that ER extranuclear signaling has the potential to promote breast cancer cell migration and metastasis.
5. ERα Regulation of Metastasis
Metastases spawned by malignant tumors that have acquired increased invasiveness are responsible for almost all breast-cancer-related morbidity and mortality. The majority of ERα-positive cells retain their ERα and respond positively to initial endocrine therapy for the treatment of advanced metastatic disease. Several recent studies have detected the presence of ERα expression in metastatic tumors [46–48]. A correlation between ERα-positive tumors and the development of bone metastasis has been observed clinically [49, 50]. Many metastatic tumors retain ERα. If primary tumors are ERα positive, greater than 80% of the lymph node metastases, and 65–70% of distant metastases retain ERα [46, 47]. A clinical correlation has also been reported between ERα-positive tumors and the development of bone metastasis [49, 50]. ERα signaling has also been shown to enhance lung metastasis [51]. In addition, ERα-mediated signaling has enhanced lung metastasis by promoting host-compartment response [51]. These emerging findings suggest that ERα signaling plays a role in metastasis.
6. ERβ Regulation of Cell Migration and Metastasis
ERβ, similar to ERα, also functions as a transcription factor that mediates different physiological responses to estrogen signaling. However, the physiological consequences of ERβ-mediated transcriptional regulation are distinct from those of ERα [1]. A number of recent studies suggest that an increase in ERβ expression decreases cell proliferation and that ERβ has antiproliferative (tumor suppressor) functions [52–54]. Reduced expression of ERβ was reported in invasive breast cancer [55], and ERβ expression is associated with less invasive and proliferating tumors [56]. Downregulation of ERβ is shown to promote epithelial-to-mesenchymal transition (EMT) in prostate cancer cells [57]. A recent study using breast cancer model cells provided evidence that ERβ expression was associated with less cell migration. Mechanistic studies indicated that ERβ affects integrin expression and clustering and consequently modulates adhesion and migration of breast cancer cells [58]. Collectively, the emerging evidence in various model cells (including ovary and prostrate) suggests that ERβ signaling may promote antimigratory and anti-invasive responses; however, future studies using breast models are needed to further validate these findings.
7. Estrogen Regulation of EMT
EMT constitutes the loss of hallmark structures and physiologic properties associated with the epithelia and the gain of new properties, including migratory and invasive growth patterns [59]. Loss of E-cadherin is a key initial step in the transdifferentiation of epithelial cells to a mesenchymal phenotype, which occurs when tumor epithelial cells invade the surrounding tissues [60]. Evolving evidence suggests that estrogen signaling can influence EMT and ERα signaling crosstalk with several EMT regulators such as Snail and Slug. ERα directly binds to and regulates the promoter of metastasis tumor antigen (MTA) 3 that suppresses Snail, a gene implicated in EMT transition [61]. ERα downregulates Slug transcription by the formation of a corepressor complex involving HDAC1 (histone deacetylase 1) and N-CoR (nuclear receptor co-repressor) [62]. Estrogen promotes down-regulation of E-cadherin via transcriptional regulation by recruitment of corepressors such as scaffold attachment factor B [63]. Estrogen plays an important role in cytoskeletal rearrangements mediated by delocalization of E-cadherin [64]. Furthermore, a recent study found that E2 promotes reversible EMT-like transition as well as collective motility in ERα-positive cells [65]. Estrogen-regulated EMT is complex and is dependent on temporal expression patterns of MTA family members, cell-adhesion-essential regulators, and ER coregulators [66]. ERα signaling negatively regulates EMT by modulating MTA3 expression and thus promotes differentiation [61]. Collectively, these findings implicate that estrogen-mediated EMT depends on the cellular repertoire of ERα coregulators and EMT regulators and that their cross talk has potential to differentially affect breast cancer progression, leading to metastasis via EMT changes.
8. Tumor Microenvironment Regulation of ER Signaling
The metastasis signaling cascade is orchestrated through the activation of biochemical pathways that involve the tumor microenvironment. Stromal cells (fibroblasts, inflammatory cells, and endovascular cells) play important roles to create a supportive environment for tumor cell growth [67, 68]. Chemokines produced by stromal cells have potential to influence ERα-positive breast cancer progression to metastasis. The chemokine CXCL12/SDF-1 and its G-protein-coupled receptor CXCR4-mediated signaling pathways play important roles in the migration and invasion of breast cancer cells. Some evidence suggests that HER2-mediated breast tumor metastasis may involve HER2 and CXCR4 signaling pathway cross talk [69]. CXCR4 overexpression correlated with worse prognosis in patients and constitutive activation of CXCR4 in poorly metastatic ER-positive MCF7 cells led to enhanced tumor growth and metastasis. The results from this study showed that enhanced CXCR4 signaling is sufficient to drive ERα-positive breast cancers to a metastatic and endocrine-therapy-resistant phenotype via increases in MAPK signaling [70].
The intratumoral levels of estrogens and growth factors are regulated by the tumor-stromal interactions in the tumor microenvironment [71]. Cross talk between the tumor and stromal cells promote expression of aromatase, a key enzyme in E2 biosynthesis, resulting in intra-tumoral estrogen production in postmenopausal breast tumors [72]. Tumor-stromal cross talk regulates aromatase gene expression via the production of various factors such as COX2, tumor necrosis factor-α, interleukin-6, and interleukin-11 [71]. Tumor-stromal interactions also contribute to the expression of growth factors such as EGF and IGF-1, which activate the ERα through growth factor receptor cross talk, leading to ERα-positive breast cancer progression [73].
9. ER Signaling Components as Potential Biomarkers for Predicting Metastasis
ERα status is routinely used in the clinic for treatment selection; however, additional markers are urgently needed to predict metastasis. Considering the evolving significance of ERα coregulators (SRC family members such as SRC-3/AIB1) in mammary tumor invasion and metastasis [74], SRC-3 status could be used as a diagnostic biomarker. Similarly, expression of the ER coregulator PELP1 is deregulated in metastatic breast tumors [37], and PELP1 protein expression is an independent prognostic predictor of breast cancer-specific survival and disease-free survival [38]. Since PELP1 plays a critical role in estrogen-mediated extranuclear signaling, these findings suggest that PELP1 could be used as a potential biomarker for predicting ER-driven metastasis. Several studies using various Src kinase inhibitors and dominant-negative mutants demonstrated that inhibiting c-Src activity decreased the metastatic potential of breast cancer cells [75]. Given the role of Src kinase in ER signaling, phosphor-c-Src is an attractive biomarker for predicting breast cancer metastasis in conjunction with other prognostic factors. Few recent preclinical studies using Src inhibitors confirmed the downstream target of Phos-Src and -FAK and could be possible diagnostic markers [76]. Because AKT signaling is implicated in invasive ductal carcinoma of the breast and implicated in ERα-mediated extranuclear actions leading migration/invasion, Phospho AKT (pAKT) status could be a potential biomarker in the prediction of therapeutic response in invasive ductal carcinoma of the breast [74]. Even though these emerging findings suggest ERα-signaling molecules as potential biomarkers, additional studies using a large set of human tumor samples are needed to clearly establish them as prognostic markers.
10. Therapeutic Targeting of ERα Signaling for Blocking Metastasis
The emerging significance of the ERα in the metastatic cascade indicates novel possibilities for therapeutic targeting of specific ERα signaling components that mediate migration, invasion, and EMT. A large portion of metastases retain their ERα when the primary tumors are ERα positive. Several recent studies detected the presence of ERα and aromatase expression in metastatic tumors [46–48]. We envision that the therapies targeting ER signaling axis leading to metastasis are more suitable for early stage patients who have tumors that are amenable to biopsy and IHC analysis. Potential markers of ERα signaling that are implicated in metastasis (including kinases such as Src, AKT, and PI3K and coregulators such as PELP1, AIB1, and SRC-1) could be used in addition to traditional ERα status to identify this subset of patients.
Aromatase is recognized as a potent target in endocrine therapy for the treatment of postmenopausal breast cancers [73]. Because some metastases retain their ERα signaling, screening of patients with advanced breast cancer for expression of ERα, ER-coregulators, and aromatase may provide a rationale for the development of customized treatment of a subset of patients with ERα-positive and aromatase-positive cancer. These patients could be treated with an aromatase inhibitor (Letrozole) that ablates peripheral estrogen synthesis and ERα degraders/signaling blockers for their ERα-positive metastatic tumors.
Because ERα and ERβ have different physiological functions and have ligand-binding properties that differ enough to be selective in their ligand binding, opportunities now exist for testing of novel ER subtype-specific, selective ER modulators [77]. Several synthetic or novel natural compounds derived from plant materials have the potential to function as ERβ agonists [54, 78], and these compounds may have utility in augmenting ERβ tumor suppressive functions.
If ERβ can hamper the regulation of ERα and inhibit the proliferation as well as affect the crosstalk with growth factors and their receptors, testing of ERβ agonist in combination with other endocrine therapies will provide a novel means to target ERα-driven metastasis. Recent studies found a therapeutic efficacy using ERβ agonists in combination with aromatase inhibitors, and this strategy may be useful in treating aromatase-inhibitor-(AI-) resistant metastatic breast cancer [79].
ERα-positive metastasis has been associated with chemokine signaling through SDF-1-CXCR4. Therefore, CXCR4 signaling is a rational therapeutic target for the treatment of ER-positive advanced breast carcinomas [70]. Integrin-linked kinase (ILK) is a nodal molecule in many molecular pathways that are implicated in cancer metastasis. Recent evidence suggests that ER extranuclear signaling utilizes the ILK axis [10]; therefore, ILK inhibitors such as QLT-0267 could be used to curb motility of breast cancer cells [80]. Since arginine methylation is implicated in ERα extranuclear signaling, blocking arginine methylases could be a possible therapeutic target. Compounds such as guanidine-nitrogen-substituted peptides or the thioglycolic amide RM65 may be useful to block this pathway [81, 82]. SRC3/AIB1 is frequently amplified or overexpressed in human breast cancer and is implicated in breast cancer progression to advanced ERα-positive tumors. Mechanistic studies showed AIB1 overexpression activates the mammalian target of rapamycin (mTOR), and activation of mTOR pathway is critical for AIB1-driven tumorigenesis [83]. Recent studies suggest that mTOR inhibition and ER-targeted endocrine therapy may improve the outcome of the subset of patients with ER-positive breast cancers overexpressing AIB1 [84].
Emerging evidence suggest that Src participates in ERα extranuclear actions and its wide deregulation in breast tumors suggests that it could be a potential candidate for treating ERα-positive metastasis [85]. The fact that Src can mediate interactions between the ERα and growth-factor-signaling pathways is of particular importance because cross talk between these pathways is implicated in activation of ERα extranuclear signaling leading to cell migration and invasion [10]. Further, the ability of the Src axis to promote local estrogen synthesis via aromatase activation has potential to form an autocrine loop of ERα signaling leading to tumor cell proliferation and metastasis [86]. Thus, blocking the Src axis could block ERα signaling at multiple fronts and thus reducing the ability of the ERα to promote metastasis. Recent studies found that inhibition of the Src family tyrosine kinases using inhibitors such as dasatinib can block ERα-mediated extranuclear actions leading to cell migration and invasion [10]. Therefore, it is tempting to speculate that combination of hormonal therapy with dasatinib, an orally available inhibitor of Src family tyrosine kinases that is currently approved for clinical trials to treat solid tumors [87–89], may be useful in curbing breast cancer metastases.
11. Conclusions/Significance
The most deadly aspect of breast cancer is its ability to spread or metastasize. Recent mechanistic studies have increased our understanding and highlight a role of estrogen-induced rapid ERα extranuclear signaling in facilitating the metastatic process. This signaling pathway thus provides new targets for therapeutic intervention. During progression from tumorigenesis to invasion, tumor cells trigger signals that activate ERα-extranuclear-signaling pathways, leading to enhanced cell migratory functions and metastasis, thus ER extranuclear signaling represents an important target for metastatic control of ERα-positive tumors (Figure 2). Since multiple signaling pathways in addition to estrogen are involved in activating ERs, combination therapies using both endocrine and nonendocrine agents that block different pathways may have better therapeutic effects and may delay the development of estrogen-driven metastasis. Future studies identifying the molecular mechanisms of ERα signaling contributing to ERα-driven metastasis as well as examining the prognostic/diagnostic significance of ERα signaling components using a larger sample size of tumors is warranted. Further, elucidation of the pathologic roles of ERα extranuclear signaling in metastasis will have important implications for development of novel breast cancer therapeutics and in the development of the next generation of selective ER modulators.
Figure 2.
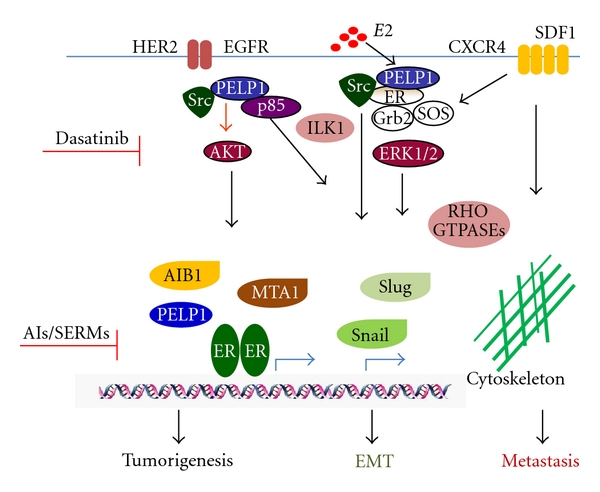
Schematic representation of hormonal regulation of metastasis. ERα-mediated signaling involves nuclear as well as extranuclear actions and growth factor signaling cross talk. Estrogen signaling has the potential to activate extranuclear signaling that activates several kinase cascades, which have potential to alter cytoskeleton, EMT and enhance cell migration. Deregulation of ERα-mediated signaling crosstalk will have implications in estrogen-mediated tumor progression to metastasis.
Acknowledgments
This work was supported by NIH-CA095681 (RKV), DOD-W81XWH-08-1-0604 (RKV) and NIH T32CA148724 (SSR) Grants.
References
- 1.Thomas C, Gustafsson J-A. The different roles of ER subtypes in cancer biology and therapy. Nature Reviews Cancer. 2011;11(8):597–608. doi: 10.1038/nrc3093. [DOI] [PubMed] [Google Scholar]
- 2.Lewis-Wambi JS, Jordan VC. Treatment of postmenopausal breast cancer with selective estrogen receptor modulators (SERMs) Breast Disease. 2005;24(1):93–105. doi: 10.3233/bd-2006-24108. [DOI] [PubMed] [Google Scholar]
- 3.Leary A, Dowsett M. Combination therapy with aromatase inhibitors: the next era of breast cancer treatment? British Journal of Cancer. 2006;95(6):661–666. doi: 10.1038/sj.bjc.6603316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Utsumi T, Kobayashi N, Hanada H. Recent perspectives of endocrine therapy for breast cancer. Breast Cancer. 2007;14(2):194–199. doi: 10.2325/jbcs.959. [DOI] [PubMed] [Google Scholar]
- 5.Steeg PS. Tumor metastasis: mechanistic insights and clinical challenges. Nature Medicine. 2006;12(8):895–904. doi: 10.1038/nm1469. [DOI] [PubMed] [Google Scholar]
- 6.Stetler-Stevenson WG, Liotta LA, Kleiner DE., Jr. Extracellular matrix 6: role of matrix metalloproteinases in tumor invasion and metastasis. FASEB Journal. 1993;7(15):1434–1441. doi: 10.1096/fasebj.7.15.8262328. [DOI] [PubMed] [Google Scholar]
- 7.Friedl P, Wolf K. Tumour-cell invasion and migration: diversity and escape mechanisms. Nature Reviews Cancer. 2003;3(5):362–374. doi: 10.1038/nrc1075. [DOI] [PubMed] [Google Scholar]
- 8.Rochefort H, Platet N, Hayashido Y, et al. Estrogen receptor mediated inhibition of cancer cell invasion and motility: an overview. Journal of Steroid Biochemistry and Molecular Biology. 1998;65(1–6):163–168. doi: 10.1016/s0960-0760(98)00010-7. [DOI] [PubMed] [Google Scholar]
- 9.Sisci D, Aquila S, Middea E, et al. Fibronectin and type IV collagen activate ERα AF-1 by c-Src pathway: effect on breast cancer cell motility. Oncogene. 2004;23(55):8920–8930. doi: 10.1038/sj.onc.1208098. [DOI] [PubMed] [Google Scholar]
- 10.Chakravarty D, Nair SS, Santhamma B, et al. Extranuclear functions of ER impact invasive migration and metastasis by breast cancer cells. Cancer Research. 2010;70(10):4092–4101. doi: 10.1158/0008-5472.CAN-09-3834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zheng S, Huang J, Zhou K, et al. 17β-Estradiol enhances breast cancer cell motility and invasion via extra-nuclear activation of actin-binding protein ezrin. PLoS ONE. 2011;6(7, article e22439) doi: 10.1371/journal.pone.0022439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Giretti MS, Fu XD, De Rosa G, et al. Extra-nuclear signalling of estrogen receptor to breast cancer cytoskeletal remodelling, migration and invasion. PLoS ONE. 2008;3(5) doi: 10.1371/journal.pone.0002238. Article ID e2238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Sanchez AM, Flamini MI, Baldacci C, Goglia L, Genazzani AR, Simoncini T. Estrogen receptor-α promotes breast cancer cell motility and invasion via focal adhesion kinase and N-WASP. Molecular Endocrinology. 2010;24(11):2114–2125. doi: 10.1210/me.2010-0252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Li Y, Wang JP, Santen RJ, et al. Estrogen stimulation of cell migration involves multiple signaling pathway interactions. Endocrinology. 2010;151(11):5146–5156. doi: 10.1210/en.2009-1506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Warner M, Nilsson S, Gustafsson JÅ. The estrogen receptor family. Current Opinion in Obstetrics and Gynecology. 1999;11(3):249–254. doi: 10.1097/00001703-199906000-00003. [DOI] [PubMed] [Google Scholar]
- 16.Hewitt SC, Couse JF, Korach KS. Estrogen receptor knockout mice: what their phenotypes reveal about mechanisms of estrogen action. Breast Cancer Research. 2000;2(5):345–352. doi: 10.1186/bcr79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kumar V, Green S, Stack G, Berry M, Jin JR, Chambon P. Functional domains of the human estrogen receptor. Cell. 1987;51(6):941–951. doi: 10.1016/0092-8674(87)90581-2. [DOI] [PubMed] [Google Scholar]
- 18.McKenna NJ, Lanz RB, O’Malley BW. Nuclear receptor coregulators: cellular and molecular biology. Endocrine Reviews. 1999;20(3):321–344. doi: 10.1210/edrv.20.3.0366. [DOI] [PubMed] [Google Scholar]
- 19.McDonnell DP, Norris JD. Connection and regulation of the human estrogen receptor. Science. 2002;296(5573):1642–1644. doi: 10.1126/science.1071884. [DOI] [PubMed] [Google Scholar]
- 20.Tsai MJ, O’Malley BW. Molecular mechanisms of action of steroid/thyroid receptor superfamily members. Annual Review of Biochemistry. 1994;63:451–486. doi: 10.1146/annurev.bi.63.070194.002315. [DOI] [PubMed] [Google Scholar]
- 21.Barnes CJ, Vadlamudi RK, Kumar R. Novel estrogen receptor coregulators and signaling molecules in human diseases. Cellular and Molecular Life Sciences. 2004;61(3):281–291. doi: 10.1007/s00018-003-3222-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Collingwood TN, Urnov FD, Wolffe AP. Nuclear receptors: coactivators, corepressors and chromatin remodeling in the control of transcription. Journal of Molecular Endocrinology. 1999;23(3):255–275. doi: 10.1677/jme.0.0230255. [DOI] [PubMed] [Google Scholar]
- 23.Kumar R, Gururaj AE, Vadlamudi RK, Rayala SK. The clinical relevance of steroid hormone receptor corepressors. Clinical Cancer Research. 2005;11(8):2822–2831. doi: 10.1158/1078-0432.CCR-04-1276. [DOI] [PubMed] [Google Scholar]
- 24.Hall JM, McDonnell DP. Coregulators in nuclear estrogen receptor action: from concept to therapeutic targeting. Molecular Interventions. 2005;5(6):343–357. doi: 10.1124/mi.5.6.7. [DOI] [PubMed] [Google Scholar]
- 25.Han SJ, DeMayo FJ, Xu J, Tsai SY, Tsai MJ, O’Malley BW. Steroid receptor coactivator (SRC)-1 and SRC-3 differentially modulate tissue-specific activation functions of the progesterone receptor. Molecular Endocrinology. 2006;20(1):45–55. doi: 10.1210/me.2005-0310. [DOI] [PubMed] [Google Scholar]
- 26.Xu J, Qiu Y, DeMayo FJ, Tsai SY, Tsai MJ, O’Malley BW. Partial hormone resistance in mice with disruption of the steroid receptor coactivator-1 (SRC-1) gene. Science. 1998;279(5358):1922–1925. doi: 10.1126/science.279.5358.1922. [DOI] [PubMed] [Google Scholar]
- 27.Xu J, Liao L, Ning G, Yoshida-Komiya H, Deng C, O’Malley BW. The steroid receptor coactivator SRC-3 (P/CIP/RAC3/AIB1/ACTR/TRAM-1) is required for normal growth, puberty, female reproductive function, and mammary gland development. Proceedings of the National Academy of Sciences of the United States of America. 2000;97(12):6379–6384. doi: 10.1073/pnas.120166297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.O’Malley BW. Little molecules with big goals. Science. 2006;313(5794):1749–1750. doi: 10.1126/science.1132509. [DOI] [PubMed] [Google Scholar]
- 29.Wang S, Yuan Y, Liao L, et al. Disruption of the SRC-1 gene in mice suppresses breast cancer metastasis without affecting primary tumor formation. Proceedings of the National Academy of Sciences of the United States of America. 2009;106(1):151–156. doi: 10.1073/pnas.0808703105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Han JS, Crowe DL. Steroid receptor coactivator 1 deficiency increases MMTV-neu mediated tumor latency and differentiation specific gene expression, decreases metastasis, and inhibits response to PPAR ligands. BMC Cancer. 2010;10, article 629 doi: 10.1186/1471-2407-10-629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.O’Malley BW, Kumar R. Nuclear receptor coregulators in cancer biology. Cancer Research. 2009;69(21):8217–8222. doi: 10.1158/0008-5472.CAN-09-2223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Onder TT, Gupta PB, Mani SA, Yang J, Lander ES, Weinberg RA. Loss of E-cadherin promotes metastasis via multiple downstream transcriptional pathways. Cancer Research. 2008;68(10):3645–3654. doi: 10.1158/0008-5472.CAN-07-2938. [DOI] [PubMed] [Google Scholar]
- 33.Beavon IRG. The E-cadherin-catenin complex in tumour metastasisstructure, function and regulation. European Journal of Cancer. 2000;36(13):1607–1620. doi: 10.1016/s0959-8049(00)00158-1. [DOI] [PubMed] [Google Scholar]
- 34.Mazumdar A, Wang RA, Mishra SK, et al. Transcriptional repression of oestrogen receptor by metastasis-associated protein 1 corepressor. Nature Cell Biology. 2001;3(1):30–37. doi: 10.1038/35050532. [DOI] [PubMed] [Google Scholar]
- 35.Qin L, Liao L, Redmond A, et al. The AIB1 oncogene promotes breast cancer metastasis by activation of PEA3-mediated matrix metalloproteinase 2 (MMP2) and MMP9 expression. Molecular and Cellular Biology. 2008;28(19):5937–5950. doi: 10.1128/MCB.00579-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Qin L, Liu Z, Chen H, Xu J. The steroid receptor coactivator-1 regulates Twist expression and promotes breast cancer metastasis. Cancer Research. 2009;69(9):3819–3827. doi: 10.1158/0008-5472.CAN-08-4389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Rajhans R, Nair S, Holden AH, Kumar R, Tekmal RR, Vadlamudi RK. Oncogenic potential of the nuclear receptor coregulator proline-, glutamic acid-, leucine-rich protein 1/modulator of the nongenomic actions of the estrogen receptor. Cancer Research. 2007;67(11):5505–5512. doi: 10.1158/0008-5472.CAN-06-3647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Habashy HO, Powe DG, Rakha EA, et al. The prognostic significance of PELP1 expression in invasive breast cancer with emphasis on the ER-positive luminal-like subtype. Breast Cancer Research and Treatment. 2010;120(3):603–612. doi: 10.1007/s10549-009-0419-9. [DOI] [PubMed] [Google Scholar]
- 39.Levin ER. Integration of the extranuclear and nuclear actions of estrogen. Molecular Endocrinology. 2005;19(8):1951–1959. doi: 10.1210/me.2004-0390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Harrington WR, Kim SH, Funk CC, et al. Estrogen dendrimer conjugates that preferentially activate extranuclear, nongenomic versus genomic pathways of estrogen action. Molecular Endocrinology. 2006;20(3):491–502. doi: 10.1210/me.2005-0186. [DOI] [PubMed] [Google Scholar]
- 41.Finn RS. Targeting Src in breast cancer. Annals of Oncology. 2008;19(8):1379–1386. doi: 10.1093/annonc/mdn291. [DOI] [PubMed] [Google Scholar]
- 42.Persad S, Dedhar S. The role of integrin-linked kinase (ILK) in cancer progression. Cancer and Metastasis Reviews. 2003;22(4):375–384. doi: 10.1023/a:1023777013659. [DOI] [PubMed] [Google Scholar]
- 43.Björnström L, Sjöberg M. Mechanisms of estrogen receptor signaling: convergence of genomic and nongenomic actions on target genes. Molecular Endocrinology. 2005;19(4):833–842. doi: 10.1210/me.2004-0486. [DOI] [PubMed] [Google Scholar]
- 44.Kumar R, Wang RA, Mazumdar A, et al. A naturally occurring MTA1 variant sequesters oestrogen receptor-α in the cytoplasm. Nature. 2002;418(6898):654–657. doi: 10.1038/nature00889. [DOI] [PubMed] [Google Scholar]
- 45.Teyssier C, Le Romancer M, Sentis S, Jalaguier S, Corbo L, Cavaillès V. Protein arginine methylation in estrogen signaling and estrogen-related cancers. Trends in Endocrinology and Metabolism. 2010;21(3):181–189. doi: 10.1016/j.tem.2009.11.002. [DOI] [PubMed] [Google Scholar]
- 46.Harrell JC, Dye WW, Allred DC, et al. Estrogen receptor positive breast cancer metastasis: altered hormonal sensitivity and tumor aggressiveness in lymphatic vessels and lymph nodes. Cancer Research. 2006;66(18):9308–9315. doi: 10.1158/0008-5472.CAN-06-1769. [DOI] [PubMed] [Google Scholar]
- 47.Zheng WQ, Lu J, Zheng JM, Hu FX, Ni CR. Variation of ER status between primary and metastatic breast cancer and relationship to p53 expression. Steroids. 2001;66(12):905–910. doi: 10.1016/s0039-128x(01)00121-0. [DOI] [PubMed] [Google Scholar]
- 48.Lu J, Li H, Cao D, et al. Clinical significance of aromatase protein expression in axillary node negative breast cancer. Journal of Cancer Research and Clinical Oncology. 2007;133(6):401–409. doi: 10.1007/s00432-006-0186-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Koenders PG, Beex LVAM, Langens R, Kloppenborg PWC, Smals AGH, Benraad TJ. Steroid hormone receptor activity of primary human breast cancer and pattern of first metastasis. Breast Cancer Research and Treatment. 1991;18(1):27–32. doi: 10.1007/BF01975440. [DOI] [PubMed] [Google Scholar]
- 50.Wang J, Jarrett J, Huang CC, Satcher RL, Levenson AS. Identification of estrogen-responsive genes involved in breast cancer metastases to the bone. Clinical and Experimental Metastasis. 2007;24(6):411–422. doi: 10.1007/s10585-007-9078-6. [DOI] [PubMed] [Google Scholar]
- 51.Banka CL, Lund CV, Nguyen MTN, Pakchoian AJ, Mueller BM, Eliceiri BP. Estrogen induces lung metastasis through a host compartment-specific response. Cancer Research. 2006;66(7):3667–3672. doi: 10.1158/0008-5472.CAN-05-4416. [DOI] [PubMed] [Google Scholar]
- 52.Speirs V, Carder PJ, Lane S, Dodwell D, Lansdown MRJ, Hanby AM. Oestrogen receptor β: what it means for patients with breast cancer. Lancet Oncology. 2004;5(3):174–181. doi: 10.1016/S1470-2045(04)01413-5. [DOI] [PubMed] [Google Scholar]
- 53.Ström A, Hartman J, Foster JS, Kietz S, Wimalasena J, Gustafsson JÅ. Estrogen receptor β inhibits 17β-estradiol-stimulated proliferation of the breast cancer cell line T47D. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(6):1566–1571. doi: 10.1073/pnas.0308319100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Nilsson S, Gustafsson JÅ. Estrogen receptors: therapies targeted to receptor subtypes. Clinical Pharmacology and Therapeutics. 2011;89(1):44–55. doi: 10.1038/clpt.2010.226. [DOI] [PubMed] [Google Scholar]
- 55.Skliris GP, Munot K, Bell SM, et al. Reduced expression of oestrogen receptor β in invasive breast cancer and its re-expression using DNA methyl transferase inhibitors in a cell line model. Journal of Pathology. 2003;201(2):213–220. doi: 10.1002/path.1436. [DOI] [PubMed] [Google Scholar]
- 56.Jarvinen TAH, Pelto-Huikko M, Holli K, Isola J. Estrogen receptor β is coexpressed with ERα and PR and associated with nodal status, grade, and proliferation rate in breast cancer. American Journal of Pathology. 2000;156(1):29–35. doi: 10.1016/s0002-9440(10)64702-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Mak P, Leav I, Pursell B, et al. ERβ impedes prostate cancer EMT by destabilizing HIF-1α and inhibiting VEGF-mediated snail nuclear localization: implications for gleason grading. Cancer Cell. 2010;17(4):319–332. doi: 10.1016/j.ccr.2010.02.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Lindberg K, Ström A, Lock JG, Gustafsson J-A, Haldosén L-A, Helguero LA. Expression of estrogen receptor β increases integrin α1 and integrin β1 levels and enhances adhesion of breast cancer cells. Journal of Cellular Physiology. 2010;222(1):156–167. doi: 10.1002/jcp.21932. [DOI] [PubMed] [Google Scholar]
- 59.Nieto MA. The ins and outs of the epithelial to mesenchymal transition in health and disease. Annual Review of Cell and Developmental Biology. 2011;10(27):347–376. doi: 10.1146/annurev-cellbio-092910-154036. [DOI] [PubMed] [Google Scholar]
- 60.Schmalhofer O, Brabletz S, Brabletz T. E-cadherin, β-catenin, and ZEB1 in malignant progression of cancer. Cancer and Metastasis Reviews. 2009;28(1-2):151–166. doi: 10.1007/s10555-008-9179-y. [DOI] [PubMed] [Google Scholar]
- 61.Fujita N, Jaye DL, Kajita M, Geigerman C, Moreno CS, Wade PA. MTA3, a Mi-2/NuRD complex subunit, regulates an invasive growth pathway in breast cancer. Cell. 2003;113(2):207–219. doi: 10.1016/s0092-8674(03)00234-4. [DOI] [PubMed] [Google Scholar]
- 62.Ye Y, Xiao Y, Wang W, Yearsley K, Gao JX, Barsky SH. ERα suppresses slug expression directly by transcriptional repression. Biochemical Journal. 2008;416(2):179–187. doi: 10.1042/BJ20080328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Oesterreich S, Deng W, Jiang S, et al. Estrogen-mediated down-regulation of E-cadherin in breast cancer cells. Cancer Research. 2003;63(17):5203–5208. [PubMed] [Google Scholar]
- 64.DePasquale JA. Rearrangement of the F-actin cytoskeleton in estradiol-treated MCF-7 breast carcinoma cells. Histochemistry and Cell Biology. 1999;112(5):341–350. doi: 10.1007/s004180050415. [DOI] [PubMed] [Google Scholar]
- 65.Planas-Silva MD, Waltz PK. Estrogen promotes reversible epithelial-to-mesenchymal-like transition and collective motility in MCF-7 breast cancer cells. Journal of Steroid Biochemistry and Molecular Biology. 2007;104(1-2):11–21. doi: 10.1016/j.jsbmb.2006.09.039. [DOI] [PubMed] [Google Scholar]
- 66.Zhang H, Stephens LC, Kumar R. Metastasis tumor antigen family proteins during breast cancer progression and metastasis in a reliable mouse model for human breast cancer. Clinical Cancer Research. 2006;12(5):1479–1486. doi: 10.1158/1078-0432.CCR-05-1519. [DOI] [PubMed] [Google Scholar]
- 67.Kalluri R, Zeisberg M. Fibroblasts in cancer. Nature Reviews Cancer. 2006;6(5):392–401. doi: 10.1038/nrc1877. [DOI] [PubMed] [Google Scholar]
- 68.Allinen M, Beroukhim R, Cai L, et al. Molecular characterization of the tumor microenvironment in breast cancer. Cancer Cell. 2004;6(1):17–32. doi: 10.1016/j.ccr.2004.06.010. [DOI] [PubMed] [Google Scholar]
- 69.Li YM, Pan Y, Wei Y, et al. Upregulation of CXCR4 is essential for HER2-mediated tumor metastasis. Cancer Cell. 2004;6(5):459–469. doi: 10.1016/j.ccr.2004.09.027. [DOI] [PubMed] [Google Scholar]
- 70.Rhodes LV, Short SP, Neel NF, et al. Cytokine receptor CXCR4 mediates estrogen-independent tumorigenesis, metastasis, and resistance to endocrine therapy in human breast cancer. Cancer Research. 2011;71(2):603–613. doi: 10.1158/0008-5472.CAN-10-3185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Simpson ER, Davis SR. Minireview: aromatase and the regulation of estrogen biosynthesis—Some new perspectives. Endocrinology. 2001;142(11):4589–4594. doi: 10.1210/endo.142.11.8547. [DOI] [PubMed] [Google Scholar]
- 72.Santen RJ, Santner SJ, Pauley RJ, et al. Estrogen production via the aromatase enzyme in breast carcinoma: which cell type is responsible? Journal of Steroid Biochemistry and Molecular Biology. 1997;61(3–6):267–271. [PubMed] [Google Scholar]
- 73.Yamaguchi Y. Microenvironmental regulation of estrogen signals in breast cancer. Breast Cancer. 2007;14(2):175–181. doi: 10.2325/jbcs.975. [DOI] [PubMed] [Google Scholar]
- 74.Park SS, Kim SW. Activated Akt signaling pathway in invasive ductal carcinoma of the breast: correlation with HER2 overexpression. Oncology Reports. 2007;18(1):139–143. [PubMed] [Google Scholar]
- 75.Sánchez-Muñoz A, Pérez-Ruiz E, Jiménez B, et al. Targeted therapy of metastatic breast cancer. Clinical & Translational Oncology. 2009;11(10):643–650. doi: 10.1007/s12094-009-0419-6. [DOI] [PubMed] [Google Scholar]
- 76.Jones RJ, Young O, Renshaw L, et al. Src inhibitors in early breast cancer: a methodology, feasibility and variability study. Breast Cancer Research and Treatment. 2009;114(2):211–221. doi: 10.1007/s10549-008-9997-1. [DOI] [PubMed] [Google Scholar]
- 77.Lo R, Matthews J. A new class of estrogen receptor beta-selective activators. Molecular Interventions. 2010;10(3):133–136. doi: 10.1124/mi.10.3.3. [DOI] [PubMed] [Google Scholar]
- 78.Mersereau JE, Levy N, Staub RE, et al. Liquiritigenin is a plant-derived highly selective estrogen receptor beta agonist. Molecular and Cellular Endocrinology. 2008;283(1-2):49–57. doi: 10.1016/j.mce.2007.11.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Nair HB, Kirma NB, Ganapathy M, Vadlamudi RK, Tekmal RR. Estrogen receptor-β activation in combination with letrozole blocks the growth of breast cancer tumors resistant to letrozole therapy. Steroids. 2011;76(8):792–796. doi: 10.1016/j.steroids.2011.02.038. [DOI] [PubMed] [Google Scholar]
- 80.Kalra J, Warburton C, Fang K, et al. QLT0267, a small molecule inhibitor targeting integrin-linked kinase (ILK), and docetaxel can combine to produce synergistic interactions linked to enhanced cytotoxicity, reductions in P-AKT levels, altered F-actin architecture and improved treatment outcomes in an orthotopic breast cancer model. Breast Cancer Research. 2009;11(3, article R25) doi: 10.1186/bcr2252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Spannhoff A, Machmur R, Heinke R, et al. A novel arginine methyltransferase inhibitor with cellular activity. Bioorganic and Medicinal Chemistry Letters. 2007;17(15):4150–4153. doi: 10.1016/j.bmcl.2007.05.088. [DOI] [PubMed] [Google Scholar]
- 82.Lakowski TM, T Hart P, Ahern CA, Martin NI, Frankel A. N η-substituted arginyl peptide inhibitors of protein arginine N-methyltransferases. ACS Chemical Biology. 2010;5(11):1053–1063. doi: 10.1021/cb100161u. [DOI] [PubMed] [Google Scholar]
- 83.Torres-Arzayus MI, De Mora JF, Yuan J, et al. High tumor incidence and activation of the PI3K/AKT pathway in transgenic mice define AIB1 as an oncogene. Cancer Cell. 2004;6(3):263–274. doi: 10.1016/j.ccr.2004.06.027. [DOI] [PubMed] [Google Scholar]
- 84.Torres-Arzayus MI, Yuan J, DellaGatta JL, Lane H, Kung AL, Brown M. Targeting the AIB1 oncogene through mammalian target of rapamycin inhibition in the mammary gland. Cancer Research. 2006;66(23):11381–11388. doi: 10.1158/0008-5472.CAN-06-2316. [DOI] [PubMed] [Google Scholar]
- 85.Russello SV, Shore SK. SRC in human carcinogenesis. Frontiers in Bioscience. 2004;9:139–144. doi: 10.2741/1138. [DOI] [PubMed] [Google Scholar]
- 86.Rajhans R, Nair HB, Nair SS, et al. Modulation of in situ estrogen synthesis by proline-, glutamic acid-, and leucine-rich protein-1: potential estrogen receptor autocrine signaling loop in breast cancer cells. Molecular Endocrinology. 2008;22(3):649–664. doi: 10.1210/me.2007-0350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Huang F, Reeves K, Han X, et al. Identification of candidate molecular markers predicting sensitivity in solid tumors to dasatinib: rationale for patient selection. Cancer Research. 2007;67(5):2226–2238. doi: 10.1158/0008-5472.CAN-06-3633. [DOI] [PubMed] [Google Scholar]
- 88.Summy JM, Gallick GE. Treatment for advanced tumors: Src reclaims center stage. Clinical Cancer Research. 2006;12(5):1398–1401. doi: 10.1158/1078-0432.CCR-05-2692. [DOI] [PubMed] [Google Scholar]
- 89.Araujo J, Logothetis C. Dasatinib: a potent SRC inhibitor in clinical development for the treatment of solid tumors. Cancer Treatment Reviews. 2010;36(6):492–500. doi: 10.1016/j.ctrv.2010.02.015. [DOI] [PMC free article] [PubMed] [Google Scholar]