

Abstract
The microenvironment at the site of tumor metastasis plays a key role in determining the fate of the metastasizing tumor cells. This ultimately has a direct impact on the progression of cancer. Bone is the preferred site of metastasis of breast cancer. Painful, debilitating osteolytic lesions are formed as a result of crosstalk between breast cancer cells and cells in the bone, predominantly the osteoblasts and osteoclasts. In this paper, we have discussed the temporal and spatial role of hedgehog (Hh) signaling in influencing the fate of metastatic breast cancer cells in bone. By virtue of its secreted ligands, the Hh pathway is capable of homotypic and heterotypic signaling and consequently altering the microenvironment in the bone. We also have put into perspective the therapeutic implications of using Hh inhibitors to prevent and/or treat bone metastases of breast cancer.
1. Introduction
The overwhelming numbers of cancer patients (≥90%) that die due to the dissemination of cancer cells rather than the primary tumor throw the process of metastasis to the centre stage of clinical management of cancer [1]. However, even as we embark on this review, the most poorly understood aspect of the pathogenesis and progression of cancer is the process of metastasis of the tumor.
Evolving literature supports that metastasis is a second disease imposed on the primary tumor. The outcome of metastasis is determined by the interplay between the subpopulation of metastatic cells and host homeostatic factors in the specific organ microenvironment [2]. The metastatic cascade can be conceptually organized and simplified into two major phases: (i) physical translocation of a cancer cell from the primary tumor to the microenvironment of a distant tissue (Figure 1) and (ii) colonization of secondary site (Figure 2) [3].
Figure 1.
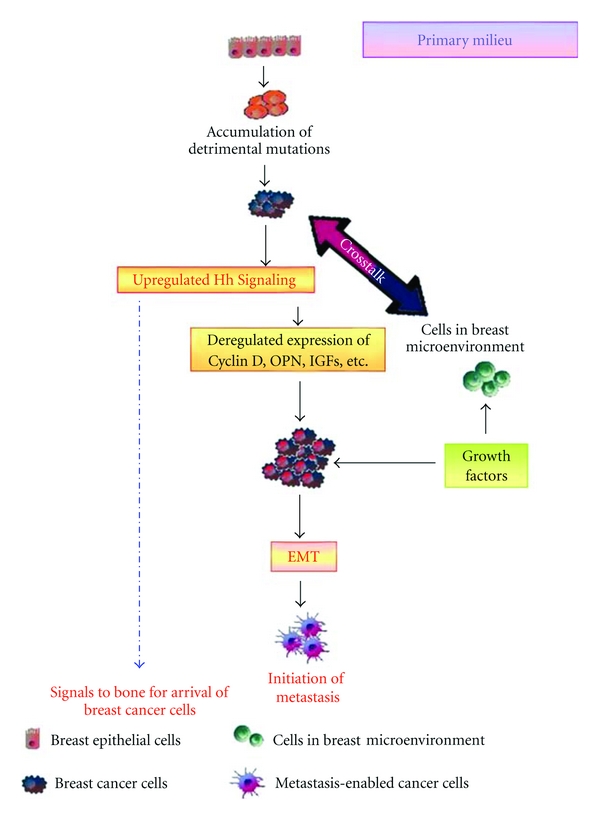
Hh signaling conditions the milieu to support metastasis of breast cancer cells to the bone. Depicted here is the first of the two microenvironments, the milieu of the primary tumor. Hh signaling in the tumor cells impacts the stromal cells in the environment, which in turn amplify paracrine Hh signaling by producing growth factors that propel epithelial-mesenchymal transition. Concomitantly, secreted, soluble proteins produced by the primary tumor contribute towards conditioning the secondary site for the arrival of the tumor cells.
Figure 2.
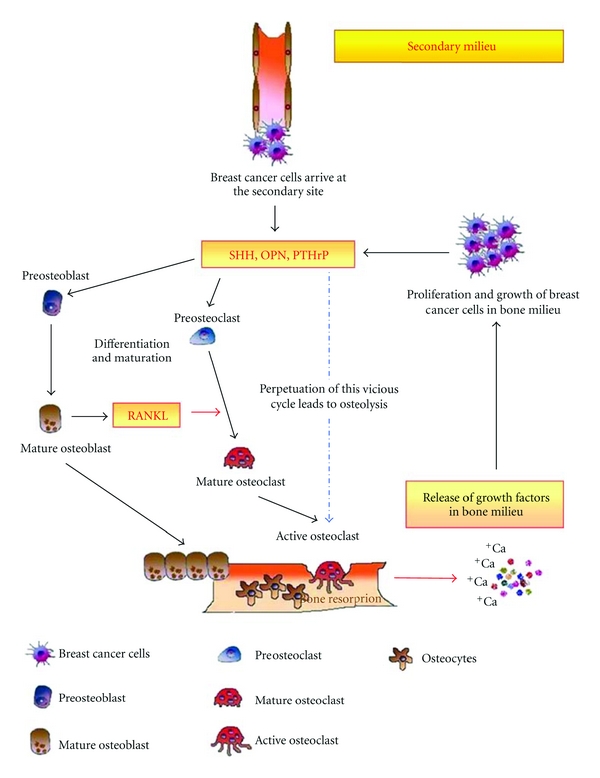
Breast cancer cells armed with Hh signaling disrupt the dynamic equilibrium in the bone to serve its purpose of self propagation and subsequent osteolysis. Breast cancer cells engane in a crosstalk with osteoblasts and osteoclasts. This cumulatively results in the differentiation and activation of osteoclasts and eventually leads to enhancing osteolysis and growth of breast tumor cells in the bone. Overall, this figure addresses the role of Hh signaling in the vicious cycle of osteolytic metastasis of breast cancer.
The metastasizing tumor cells hijack many of the pathways that play major roles during normal development. Many of the embryonic developmental signaling pathways, such as the Wnt, Hedgehog (Hh), and Notch pathways, affect the survival of tumor stem cells and orchestrate a complex microenvironment that promotes tumor survival and progression. In this review, we will highlight the significance of the Hh pathway in developmental biology and our present understanding of its role in regulating breast cancer metastasis to bone. We will elaborate how a pathway that is so critical in normal development of the embryo is usurped by the breast cancer cells to serve their own purpose of invading the tissue of its origin, extravasation, survival during translocation, and adaptation at the distant site to bring about proliferation and colonization.
2. The Hh Pathway in Normal Development
The Hh pathway plays a central role in embryonic development and maintenance of stem or progenitor cells in many adult tissues [4]. The Hh family of secreted proteins signal through both autocrine and paracrine mechanisms to control cell proliferation, differentiation, and morphology [5]. The ligands comprise desert hedgehog (DHH), Indian hedgehog (IHH), and Sonic hedgehog (SHH). Hh signaling in mammalian cells is mediated by the GLI family of zinc finger transcription factors comprising GLI1, GLI2, and GLI3. GLI1 is a strong transcriptional activator; GLI2 can function as an activator or a repressor in a context-dependent manner; GLI3 is mostly a repressor [6]. In its classical form, in the absence of the ligand, the Hh-signaling pathway is inactive, GLI1 is sequestered in the cytoplasm and repressed for its transcription activity. Binding of the Hh ligands to the receptor, a 12-pass transmembrane protein called patched-1 or patched-2 (PTCH1 or -2), releases the inhibitory affect of PTCH on a serpentine protein called Smoothened (SMO) [7]. SMO gets hyperphosphorylated and localizes to primary cilia where [8] GLI1 is activated by release from a large protein complex and translocates to the nucleus to function as a transcriptional activator [9] of several target genes, including PTCH, insulin-like growth factor-binding protein and cyclin D2 [10].
The involvement of the Hh pathway, in particular the ligand SHH, with the skeletal system begins with embryonic development, where SHH is expressed in the notochord, the floorplate of the neural tube, the brain, the zone of polarizing activity in the developing limbs, and the gut [11, 12]. SHH specifically functions in many different ways to contribute to the patterning of a developing embryo in a concentration-dependent manner along a target range [13]. A variety of embryonic defects and diseases result from mutations in the Hh pathway [14]. The long-range morphogenic properties of SHH signaling are also evident in the development of the CNS [15]. Thus, temporal and spatial regulation of SHH signaling is key to proper organogenesis. However, in the adults, this pathway is mainly inactive [16] and may play a role in the maintenance and renewal of normal stem cell population in the nervous system [17]. Moreover, Lavine et al. reported that the Hh signaling is essential for cardiac function at the level of the coronary vasculature [18].
3. The Hedgehog Pathway in Cancer
The Hh pathway is required for normal proliferation of human melanocytes in vitro and for proliferation and survival of human melanoma in vivo [19, 20]. In esophageal squamous cell carcinoma, GLI1 expression has been associated with lymphatic metastasis [21], while in breast cancer, strong nuclear GLI staining was observed [22]. Li et al. have recently reported that pancreatic cancer stem cells express high levels of SHH [23]. This is interesting given the implications for SHH in adult stem cell renewal, in pancreatic ductal progenitor cells, and also in adult hair follicle stem cells [24]. SHH is misregulated in pancreatic adenocarcinoma, prostate adenocarcinoma, esophageal and stomach cancer, and nonsmall cell carcinoma [14]. As such, Hh signaling has been shown to be active in multiple cancer types [22, 25–48] (Table 1).
Table 1.
Cancers with aberrant activation of Hh signaling.
| Milieu | Hh Signaling caused by | Molecule(s) involved | Type of cancer | Reference |
|---|---|---|---|---|
| I | Overexpression | GLI1 | Glioblastoma | [30] |
| Mutations | PTCH | Basal cell carcinoma (BCC) | [31, 32] | |
| SMO | Basal cell carcinoma | [31, 32] | ||
| PTCH | Medulloblastoma | [33] | ||
| PTCH | Rhabdomyosarcoma | [34] | ||
| PTCH1 | Gorlin syndrome BCC | [35, 36] | ||
| SMO & PTCH1 | Nonfamilial BCC | [37] | ||
|
| ||||
| II | Ligand-dependent autocrine | Breast | [22] | |
| Pancreatic | [38] | |||
| Lung cancer | [39] | |||
| Oesophagal | [40] | |||
| Prostate | [41] | |||
| Gastric adenocarcinoma | [42] | |||
| Colorectal | [43] | |||
| Hepatocellular adenocarcinoma | [44] | |||
| Ovarian carcinoma | [45, 49] | |||
| Ligand-dependent paracrine | Pancreatic | [46–48] | ||
Milieu I represents the microenvironment of the primary tumor; Milieu II represents the microenvironment at the metastatic site.
Active Hh signaling is also found to influence the tumor stromal microenvironment [27] and supports stem cells in the tumor in an undifferentiated, proliferative state [26, 50]. SHH is not only a mediator of angiogenesis but has also been shown to induce vessel formation in endothelial cells [51] and activate expression of angiopoietins I and II, and VEGF-signaling proteins from mesenchymal cells, highlighting the significance of tumor-associated fibroblasts in combination with canonical Hh signaling to mediate blood vessel formation [52]. Cancer cells utilize abnormal Hh signaling (both autocrine and paracrine) to influence proliferation and differentiation of their surrounding environment.
The role of Hh signaling in cancer has been revealed by studies that have manipulated the expression of the GLI transcription factors or the ligands or upon treatment with pharmacologic inhibitors that restrict Hh signaling. In pancreatic cancer cell lines, disruption of Hh signaling by the inhibitor cyclopamine, inhibited epithelial-mesenchymal-transition (EMT) [53, 54]. Tumor burden and metastasis in both prostate and pancreatic adenocarinomas were also reduced as a result of Hh signaling inhibition [53, 55]. In contrast, enforced expression of GLI1 induced the expression of Snail [56], an EMT marker. Conversely, we observed loss of mesenchymal markers upon abrogation of GLI1 expression [19]. Overall, GLI1 silencing had a pronounced effect on tumor malignancy in vivo by reducing metastasis. We also reported that signaling via the Hh pathway transcriptionally upregulates OPN [19]. OPN is a secreted protein that influences multiple downstream signaling events that allow cancer cells to resist apoptosis, invade through extracellular matrix, evade host immunity [57], and influence growth of indolent tumors [58, 59]. OPN constitutes a component of the secretome of several melanoma-derived cell lines [60, 61] and is also expressed in metastatic breast cancer cell lines [62]. It is highly probable that active Hh signaling in a subset of cancer cells can be propagated in a paracrine manner by OPN secreted into the tumor microenvironment. OPN, by virtue of its ability to signal through multiple receptors, can promote malignant behavior in neighboring cancer cells, regardless of the status of the Hh pathway, thereby propagating paracrine Hh signaling. Thus, at the site of origin, the breast tumor cells not only potentiate their own aggressiveness by influencing the neighboring cells, but also send signals to the secondary target organ to condition for relocalization [58, 63, 64].
For the purpose of this review, we have focused the remainder of the article on discussing the role of Hh signaling in impacting breast cancer metastasis to the bone. This complication of breast cancer continues to present a challenge to oncologists and reduces the chances of survival for breast cancer patients. Among breast cancers that become aggressive, metastasis to bone marrow is common. Detection of bone metastasis often signals the onset of the life-threatening phase of breast cancer. The 5-year survival rate is 98% for breast cancer when detected early; this precipitously drops to 83% for patients initially diagnosed with regional spread and to 26% for those with distant metastases. In the following sections, we will discuss the role of Hh signaling in mediating a crosstalk between breast cancer cells and cells in the bone and the overall impact on the ability of breast cancer cells to sculpt the bone microenvironment and cause osteolysis (Figures 1 and 2).
4. The Bone Microenvironment
The bone microenvironment comprises osteoblasts, osteoclasts, mineralized bone matrix, and other cell types, such as the osteocytes embedded within bone. Of these, the most important ones (from the perspective of this article) are the bone-resorbing osteoclasts and bone-forming osteoblasts.
Osteoblasts are derived from mesenchymal stem cells, which can also give rise to chondrocytes, fibroblasts, myocytes, or adipocytes [65]. Formation of new bone and the regulation of osteoclastogenesis through expression of RANKL and OPG are two main functions of the osteoblasts. Various growth factors and hormones like BMPs, PTHrP, TGF0β, and so forth are known to take part in the differentiation of preosteoblasts into mature osteoblasts. Eventually, mature, mineralizing osteoblasts become embedded in the newly secreted bone matrix and undergo terminal differentiation to form osteocytes. Although the osteocytes have much reduced activity as compared to osteoblasts, their long processes allow them to connect the entire matrix via a series of canaliculi. It is understood that the osteocytes ensure communication between sites deep in the bone and the extraosseous world; they create an enormous increase in mineral surface exposed to extracellular fluid and cellular activity and function as mechanosensory cells of bone, involved in the transduction of mechanical loads into biochemical signals [66].
Osteoclasts, on the other hand, are large multinucleated terminally differentiated cells with a unique ability for bone resorption [67]. They are derived from hematopoietic stem cells. The cells undergo proliferation in response to M-CSF. The precursor cells flaunt receptor activator of nuclear factor κB (RANK) on the surface, while the ligand RANKL is expressed by the bone marrow stromal cells and osteoblasts. Binding of the ligand to the receptor commits the precursor cells to the osteoclast lineage. The same interaction is also critical for osteoclast formation and can also promote osteoclast activity, since RANK is also present on the surface of terminally differentiated osteoclasts. The fusion of osteoclast precursor cells results in the formation of large multinucleated active osteoclasts.
Osteoprotegerin (OPG) is a soluble decoy receptor and a competitor of RANKL in its binding with RANK and thus can inhibit osteoclastogenesis. Therefore, the balance of RANKL and OPG is critical for osteoclast formation and activity. Osteoclasts attach to the bone surface via actin-rich podosomes enabling them to form sealed zones with ruffled borders. Proteolytic enzymes such as CTSK (Cathepsin K) and MMPs are secreted into this isolated environment, resulting in degradation of the bone matrix, dissolution of the bone mineral, and resorption of the bone [68]. Evidently behind its outward rigidity, bone is a highly dynamic organ where homeostasis is tightly controlled and largely dependent upon cellular communication between osteoclasts and osteoblasts. This tight coupling between bone resorption and bone formation is essential for the correct function and maintenance of the skeletal system, repairing microscopic skeletal damage, and replacing aged bone. Any deviation from this homeostasis results in a range of pathologic diseases, including osteoporosis and cancer-induced bone disease.
5. The Metastasis of Breast Cancer Cells to the Bone
The vertebral venous system is the most common mode of transport of breast cancer cells from the breast to bone [69]. This allows breast cancer cells to come into contact with the axial skeleton, including the ribs, spine, pelvis, and proximal humerus and femur, which is the main distribution of bone metastases in breast cancer patients [70]. Tumor cells, even at their site of origin, send signals to their preferred secondary site [64] of metastasis. This modulates the micro-environment of that region. It is likely that the Hh ligands and secreted factors such as IGFs and OPN may impact this “homing” mechanism. It can be speculated that the factors secreted by breast cancer cells create a “premetastatic niche” as termed by Lyden and colleagues [64, 71]. The role of chemokines and cytokines as well as the homing mechanism has also been elaborately discussed in a review by Bussard et al. [72]. Our findings show that expression and secretion of Hh ligands by the breast cancer cells augments these processes (Figure 1). Once malignant cells have migrated to the bone, their ability to colonize is facilitated by the bone microenvironment. MMPs, chemokine receptor 4 (CXCR4), VEGF, and connective tissue growth factors supposedly target metastatic tumor cells to bone and facilitate their survival within the bone microenvironment [73, 74]. Physical factors within the bone microenvironment, including hypoxia, acidic pH, and extracellular calcium, and bone-derived growth factors, such as TGF-β and insulin-like growth factors activate tumor expression of VEGF, PDGF, and endothelin (ET-1) [75]. Factors such as PTHrP, TGF-β, and IL-11 produced by breast cancer cells favor osteoclast maturation and osteolysis, leading to the release of growth factors that stimulate malignant tumor growth [76]. In fact, expression of IL-11 and OPN by breast cancer cells has been found to be critical for the osteolytic activity of breast cancer cells [74]. Thus, signals from the breast cancer cells at their primary site might trigger a cascade of events involving the osteoblast-mediated initiation of osteoclastogenesis which releases a plethora of growth factors in the bone milieu which not may only act as chemoattractants for the “metastasis-enabled” breast cancer cells but also favor the latter's establishment and further proliferation once they have migrated to the bone. This would in turn tilt the balance in favor of osteoclastogenesis as more favorable factors are then readily available to the osteoclasts in the bone milieu itself and thus would lead to a self-perpetuating vicious cycle of events (Figure 2).
6. Hh Signaling in the Bone Microenvironment
Hh-signaling-activated GLI2 transcription mediates osteoblast differentiation [77]. This is likely due to the regulated expression of bone morphogenetic protein-2, BMP-2, that is involved in osteogenic differentiation by promoting commitment of mesenchymal stem cells to the osteoblast lineage. GLI2 transcriptionally activates BMP-2 expression and also synergizes with BMP-2 in osteoblasts [78]. These contentions are contradicted by Plaisant et al. who have reported that Hh signaling causes a decrease in the expression of Runx2, a key transcription factor that regulates osteoblast differentiation [79]. It is proposed that Hh signaling may be regulating different aspects of bone formation in rodent and human systems.
OPN is one of the abundant noncollagenous proteins in bone. It promotes osteoclast function and is consistently overexpressed in highly metastatic cells. OPN accumulates at cement lines in remodeling bone [80] and is localized to cell-matrix and matrix-matrix interfaces in mineralized tissue, where it is deposited by actively resorbing osteoclasts. OPN positively impacts osteoclast formation, migration, and resorptive activity [81, 82]. We recently reported that OPN is regulated, in part, by the Hh pathway [19]. We have also shown that breast cancer cells express Hh ligands and engage in a crosstalk with osteoblasts and osteoclasts [83]. Our recent studies (communicated to Breast Cancer Research) have shown that the Hh pathway plays a role in initial osteoblasts maturation, especially in the presence of breast cancer cells (Figure 2). Following an initial accelerated differentiation process, characterized by the expression of alkaline phosphatase and expression of collagenous and noncollagenous matrix proteins such as BSP and OPN and osteoclast-maturation proteins including RANKL and PTHrP, the osteoblasts appear to undergo apoptosis.
The Hh ligands also mediate a direct dialogue between breast cancer cells and preosteoclasts and induce changes in preosteoclasts that influence the production of OPN and essential bone-resorbing proteases, CTSK, and MMP9 by osteoclasts [83]. Thus, Hh ligands produced by the metastasizing breast cancer cells are instrumental in initiating a crosstalk directly with osteoclasts and promote osteoclast differentiation and resorption activity (Figure 2). Breast cancer cells also express PTHrP as a result of Hh signaling, further amplify paracrine Hh signaling in the bone microenvironment, and add to the overall osteolytic conditions [84].
Thus, the vicious cycle of bone metastasis involves a complex crosstalk between the metastasizing breast tumor cells and the bone microenvironment through multiple extracellular factors and signaling pathways with the Hh pathway playing an essential role. Based on our findings, we would like to propose that the newly arrived breast tumor cells induce initial osteoblast differentiation which stimulates osteoclast differentiation. Soon, the situation is overwhelmed by osteoclast differentiation followed by intense bone resorption leading to the local release of generous amounts of growth factors that not only encourage their growth but also alter their phenotype, making them (cancer cells) resistant to standard cytotoxic antitumor treatments see the appendix [85, 86].
7. Conclusion
The bone microenvironment with ongoing bone resorption almost resembles sites of wound healing. The bone stroma is almost guaranteed to provide hospitable sites for disseminating colonization-competent breast cancer cells [61]. This ensures the successful proliferation and ultimate colonization of the bone by metastasizing breast tumor cells. The crosstalk between the metastasizing breast cancer cells and the bone cells, namely, the osteoblasts and the osteoclasts occurs in a fashion that not only favors proliferation of the newly arrived tumor cells in the bone milieu but also ultimately the complete subjugation of the resident (bone) pathways to serve the purpose of establishment and well-being of the tumor cells with concurrent destruction of the host environment. Therefore, it is essential to understand the interactions between tumor and bone and identify microenvironment-selective agents to halt tumor growth and bone metastasis thereby reducing the morbidity of skeletal related events [62]. Thus, given the fact that breast cancer cells express Hh ligands and that Hh signaling propels breast cancer progression, it is likely that administration of pharmacological Hh inhibitors can inhibit Hh signaling in both breast cancer cells and osteoclasts and may reduce breast-cancer-mediated bone loss in metastatic disease. This strategy targets the tumor cells as well as the bone and its microenvironment and can reduce tumor burden and tumor-derived bone lesions.
Acknowledgments
The author acknowledge support from the NIH (CA138850 to L. A. Shevde & CA140472 to R. S. Samant), Department of Defense (IDEA Award BC061257 to L. A. Shevde), Mayer Mitchell Award (to L. A. Shevde), and the USA Mitchell Cancer Institute.
Abbreviations
- BMP:
Bone morphogenetic protein
- CTSK:
Cathepsin K
- CXCR4:
Chemokine receptor 1
- DHH:
Desert hedgehog
- EMT:
Epithelial-Mesenchymal transition
- ET-1:
Endothelin-1
- GLI:
Glioma-associated oncogene
- Hh:
Hh pathway
- IHH:
Indian Hedgehog
- IL-11:
Interleukin-11
- M-CSF:
Macrophage colony-stimulating factor
- MMP9:
Matrix metalloprotease 9
- OPG:
Osteoprotegerin
- OPN:
Osteopontin
- PTCH:
Patched
- PDGF:
Platelet-derived growth factor
- PTHrP:
Parathyroid Hormone-related protein
- RANK:
Receptor activator of NF-κB
- RANKL:
Receptor activator of NF-κB ligand
- SHH:
Sonic hedgehog
- SMO:
Smoothened
- TGF-β:
Transforming growth factor-β
- VEGF:
Vascular endothelial growth factor.
Appendix
Some of the Key Players in Osteolytic Metastasis of Breast Cancer
-
BMP:
bone morphogenetic protein, a group of cytokines responsible for the tissue architecture throughout the body.
-
IGF:
insulin-like growth factors are responsible for cell proliferation and form the IGF axis.
-
PDGF:
platelet derived growth factor, a secreted molecule that regulates growth and cell division.
-
PTHrP:
parathyroid hormone-related protein is a hormone that regulates endochondral bone development and also regulates epithelial mesenchymal interactions in mammary gland formation. It is secreted by several cancer cells.
-
MMPs:
matrix metalloproteases are zinc-dependent endopeptidases, capable of degrading all kinds of extracellular matrix proteins and processing a number of bioactive molecules. They play a major role on cell proliferation, migration (adhesion/dispersion), differentiation, angiogenesis, apoptosis, and host defense.
-
OPG:
osteoprotegerin (OPG), also known as osteoclastogenesis inhibitory factor (OCIF), or tumor necrosis factor receptor superfamily member 11B (TNFRSF11B), is a basic glycoprotein that is a decoy receptor for the receptor activator of nuclear factor kappa B ligand (RANKL) and can inhibit osteoclastogenesis.
-
RANK:
receptor activator of nuclear factor κB (RANK), also known as TRANCE Receptor, is a type I membrane protein expressed on the surface of osteoclasts and is involved in their activation upon ligand binding.
-
RANKL:
receptor activator of nuclear factor kappa B ligand, also known as tumor necrosis factor ligand superfamily member 11 (TNFSF11), TNF-related activation-induced cytokine (TRANCE), osteoprotegerin ligand (OPGL), and osteoclast differentiation factor (ODF). It functions as a key factor for osteoclast differentiation and activation.
-
TGF-β:
transforming growth factor beta is an antiproliferative factor protein that controls proliferation, cellular differentiation, and other functions in most cells.
-
VEGF:
vascular endothelial growth factor is a signal protein produced by cells that stimulates vasculogenesis and angiogenesis.
References
- 1.Weigelt B, Peterse JL, Van’t Veer LJ. Breast cancer metastasis: markers and models. Nature Reviews Cancer. 2005;5(8):591–602. doi: 10.1038/nrc1670. [DOI] [PubMed] [Google Scholar]
- 2.Kraljevic Pavelic S, Sedic M, Bosnjak H, Spaventi S, Pavelic K. Metastasis: new perspectives on an old problem. Molecular Cancer. 2011;10, article 22 doi: 10.1186/1476-4598-10-22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Chaffer CL, Weinberg RA. A perspective on cancer cell metastasis. Science. 2011;331(6024):1559–1564. doi: 10.1126/science.1203543. [DOI] [PubMed] [Google Scholar]
- 4.Beachy PA, Karhadkar SS, Berman DM. Mending and malignancy. Nature. 2004;431(7007):p. 402. doi: 10.1038/431402a. [DOI] [PubMed] [Google Scholar]
- 5.Ingham PW, McMahon AP. Hedgehog signaling in animal development: paradigms and principles. Genes and Development. 2001;15(23):3059–3087. doi: 10.1101/gad.938601. [DOI] [PubMed] [Google Scholar]
- 6.Ruiz i Altaba A, Mas C, Stecca B. The Gli code: an information nexus regulating cell fate, stemness and cancer. Trends in Cell Biology. 2007;17(9):438–447. doi: 10.1016/j.tcb.2007.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Murone M, Rosenthal A, De Sauvage FJ. Sonic hedgehog signaling by the patched-smoothened receptor complex. Current Biology. 1999;9(2):76–84. doi: 10.1016/s0960-9822(99)80018-9. [DOI] [PubMed] [Google Scholar]
- 8.Corbit KC, Aanstad P, Singla V, Norman AR, Stainier DYR, Reiter JF. Vertebrate Smoothened functions at the primary cilium. Nature. 2005;437(7061):1018–1021. doi: 10.1038/nature04117. [DOI] [PubMed] [Google Scholar]
- 9.Lipinski RJ, Gipp JJ, Zhang J, Doles JD, Bushman W. Unique and complimentary activities of the Gli transcription factors in Hedgehog signaling. Experimental Cell Research. 2006;312(11):1925–1938. doi: 10.1016/j.yexcr.2006.02.019. [DOI] [PubMed] [Google Scholar]
- 10.Yoon JW, Kita Y, Frank DJ, et al. Gene expression profiling leads to identification of GLI1-binding elements in target genes and a role for multiple downstream pathways in GLI1-induced cell transformation. Journal of Biological Chemistry. 2002;277(7):5548–5555. doi: 10.1074/jbc.M105708200. [DOI] [PubMed] [Google Scholar]
- 11.Odenthal J, Van Eeden FJM, Haffter P, Ingham PW, Nüsslein-Volhard C. Two distinct cell populations in the floor plate of the Zebrafish are induced by different pathways. Developmental Biology. 2000;219(2):350–363. doi: 10.1006/dbio.1999.9589. [DOI] [PubMed] [Google Scholar]
- 12.Roelink H, Augsburger A, Heemskerk J, et al. Floor plate and motor neuron induction by vhh-1, a vertebrate homolog of hedgehog expressed by the notochord. Cell. 1994;76(4):761–775. doi: 10.1016/0092-8674(94)90514-2. [DOI] [PubMed] [Google Scholar]
- 13.Ingham PW. Transducing Hedgehog: the story so far. EMBO Journal. 1998;17(13):3505–3511. doi: 10.1093/emboj/17.13.3505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Di Magliano MP, Hebrok M. Hedgehog signalling in cancer formation and maintenance. Nature Reviews Cancer. 2003;3(12):903–911. doi: 10.1038/nrc1229. [DOI] [PubMed] [Google Scholar]
- 15.Cayuso J, Ulloa F, Cox B, Briscoe J, Marti E. The Sonic hedgehog pathway independently controls the patterning, proliferation and survival of neuroepithelial cells by regulating Gli activity. Development. 2006;133(3):517–528. doi: 10.1242/dev.02228. [DOI] [PubMed] [Google Scholar]
- 16.Scales SJ, de Sauvage FJ. Mechanisms of Hedgehog pathway activation in cancer and implications for therapy. Trends in Pharmacological Sciences. 2009;30(6):303–312. doi: 10.1016/j.tips.2009.03.007. [DOI] [PubMed] [Google Scholar]
- 17.Ahn S, Joyner AL. In vivo analysis of quiescent adult neural stem cells responding to Sonic hedgehog. Nature. 2005;437(7060):894–897. doi: 10.1038/nature03994. [DOI] [PubMed] [Google Scholar]
- 18.Lavine KJ, Kovacs A, Ornitz DM. Hedgehog signaling is critical for maintenance of the adult coronary vasculature in mice. Journal of Clinical Investigation. 2008;118(7):2404–2414. doi: 10.1172/JCI34561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Das S, Harris LG, Metge BJ, et al. The hedgehog pathway transcription factor GLI1 promotes malignant behavior of cancer cells by up-regulating osteopontin. Journal of Biological Chemistry. 2009;284(34):22888–22897. doi: 10.1074/jbc.M109.021949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Stecca B, Mas C, Clement V, et al. Melanomas require HEDGEHOG-GLI signaling regulated by interactions between GLI1 and the RAS-MEK/AKT pathways. Proceedings of the National Academy of Sciences of the United States of America. 2007;104(14):5895–5900. doi: 10.1073/pnas.0700776104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kawahira H, Scheel DW, Smith SB, German MS, Hebrok M. Hedgehog signaling regulates expansion of pancreatic epithelial cells. Developmental Biology. 2005;280(1):111–121. doi: 10.1016/j.ydbio.2005.01.008. [DOI] [PubMed] [Google Scholar]
- 22.Kubo M, Nakamura M, Tasaki A, et al. Hedgehog signaling pathway is a new therapeutic target for patients with breast cancer. Cancer Research. 2004;64(17):6071–6074. doi: 10.1158/0008-5472.CAN-04-0416. [DOI] [PubMed] [Google Scholar]
- 23.Li C, Heidt DG, Dalerba P, et al. Identification of pancreatic cancer stem cells. Cancer Research. 2007;67(3):1030–1037. doi: 10.1158/0008-5472.CAN-06-2030. [DOI] [PubMed] [Google Scholar]
- 24.Katoh Y, Katoh M. Hedgehog signaling pathway and gastrointestinal stem cell signaling network (Review) International Journal of Molecular Medicine. 2006;18(6):1019–1023. [PubMed] [Google Scholar]
- 25.Evangelista M, Tian H, De Sauvage FJ. The Hedgehog signaling pathway in cancer. Clinical Cancer Research. 2006;12(20):5924–5928. doi: 10.1158/1078-0432.CCR-06-1736. [DOI] [PubMed] [Google Scholar]
- 26.Jiang J, Leong NL, Mung JC, Hidaka C, Lu HH. Interaction between zonal populations of articular chondrocytes suppresses chondrocyte mineralization and this process is mediated by PTHrP. Osteoarthritis and Cartilage. 2008;16(1):70–82. doi: 10.1016/j.joca.2007.05.014. [DOI] [PubMed] [Google Scholar]
- 27.Mukherjee S, Frolova N, Sadlonova A, et al. Hedgehog signaling and response to cyclopamine differ in epithelial and stromal cells in benign breast and breast cancer. Cancer Biology and Therapy. 2006;5(6):674–683. doi: 10.4161/cbt.5.6.2906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Xuan Y, Lin Z. Expression of Indian Hedgehog signaling molecules in breast cancer. Journal of Cancer Research and Clinical Oncology. 2009;135(2):235–240. doi: 10.1007/s00432-008-0451-x. [DOI] [PubMed] [Google Scholar]
- 29.Zhang X, Harrington N, Moraes RC, Wu MF, Hilsenbeck SG, Lewis MT. Cyclopamine inhibition of human breast cancer cell growth independent of Smoothened (Smo) Breast Cancer Research and Treatment. 2009;115(3):505–521. doi: 10.1007/s10549-008-0093-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kinzler KW, Bigner SH, Bigner DD. Identification of an amplified, highly expressed gene in a human glioma. Science. 1987;236(4797):70–73. doi: 10.1126/science.3563490. [DOI] [PubMed] [Google Scholar]
- 31.Gailani MR, Bale AE. Developmental genes and cancer: role of patched in basal cell carcinoma of the skin. Journal of the National Cancer Institute. 1997;89(15):1103–1109. doi: 10.1093/jnci/89.15.1103. [DOI] [PubMed] [Google Scholar]
- 32.Xie J, Murone M, Luoh SM, et al. Activating Smoothened mutations in sporadic basal-cell carcinoma. Nature. 1998;391(6662):90–92. doi: 10.1038/34201. [DOI] [PubMed] [Google Scholar]
- 33.Zurawel RH, Allen C, Chiappa S, et al. Analysis of PTCH/SMO/SHH pathway genes in medulloblastoma. Genes Chromosomes and Cancer. 2000;27(1):44–51. doi: 10.1002/(sici)1098-2264(200001)27:1<44::aid-gcc6>3.0.co;2-v. [DOI] [PubMed] [Google Scholar]
- 34.Tostar U, Malm CJ, Meis-Kindblom JM, Kindblom L-G, Toftgård R, Undén AB. Deregulation of the hedgehog signalling pathway: a possible role for the PTCH and SUFU genes in human rhabdomyoma and rhabdomyosarcoma development. Journal of Pathology. 2006;208(1):17–25. doi: 10.1002/path.1882. [DOI] [PubMed] [Google Scholar]
- 35.Hahn H, Wicking C, Zaphiropoulos PG, et al. Mutations of the human homolog of drosophila patched in the nevoid basal cell carcinoma syndrome. Cell. 1996;85(6):841–851. doi: 10.1016/s0092-8674(00)81268-4. [DOI] [PubMed] [Google Scholar]
- 36.Johnson RL, Rothman AL, Xie J, et al. Human homolog of patched, a candidate gene for the basal cell nevus syndrome. Science. 1996;272(5268):1668–1671. doi: 10.1126/science.272.5268.1668. [DOI] [PubMed] [Google Scholar]
- 37.Lam CW, Xie J, To KF, et al. A frequent activated smoothened mutation in sporadic basal cell carcinomas. Oncogene. 1999;18(3):833–836. doi: 10.1038/sj.onc.1202360. [DOI] [PubMed] [Google Scholar]
- 38.Thayer SP, Di Magliano MP, Heiser PW, et al. Hedgehog is an early and late mediator of pancreatic cancer tumorigenesis. Nature. 2003;425(6960):851–856. doi: 10.1038/nature02009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Watkins DN, Berman DM, Burkholder SG, Wang B, Beachy PA, Baylin SB. Hedgehog signalling within airway epithelial progenitors and in small-cell lung cancer. Nature. 2003;422(6929):313–317. doi: 10.1038/nature01493. [DOI] [PubMed] [Google Scholar]
- 40.Berman DM, Karhadkar SS, Maitra A, et al. Widespread requirement for Hedgehog ligand stimulation in growth of digestive tract tumours. Nature. 2003;425(6960):846–851. doi: 10.1038/nature01972. [DOI] [PubMed] [Google Scholar]
- 41.Fan L, Pepicelli CV, Dibble CC, et al. Hedgehog signaling promotes prostate xenograft tumor growth. Endocrinology. 2004;145(8):3961–3970. doi: 10.1210/en.2004-0079. [DOI] [PubMed] [Google Scholar]
- 42.Ma X, Chen K, Huang S, et al. Frequent activation of the hedgehog pathway in advanced gastric adenocarcinomas. Carcinogenesis. 2005;26(10):1698–1705. doi: 10.1093/carcin/bgi130. [DOI] [PubMed] [Google Scholar]
- 43.Qualtrough D, Buda A, Gaffield W, Williams AC, Paraskeva C. Hedgehog signalling in colorectal tumour cells: induction of apoptosis with cyclopamine treatment. International Journal of Cancer. 2004;110(6):831–837. doi: 10.1002/ijc.20227. [DOI] [PubMed] [Google Scholar]
- 44.Cheng WT, Xu K, Tian DY, Zhang ZG, Liu LJ, Chen Y. Role of Hedgehog signaling pathway in proliferation and invasiveness of hepatocellular carcinoma cells. International Journal of Oncology. 2009;34(3):829–836. doi: 10.3892/ijo_00000209. [DOI] [PubMed] [Google Scholar]
- 45.Chen X, Horiuchi A, Kikuchi N, et al. Hedgehog signal pathway is activated in ovarian carcinomas, correlating with cell proliferation: it’s inhibition leads to growth suppression and apoptosis. Cancer Science. 2007;98(1):68–76. doi: 10.1111/j.1349-7006.2006.00353.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Tian H, Callahan CA, Dupree KJ, et al. Hedgehog signaling is restricted to the stromal compartment during pancreatic carcinogenesis. Proceedings of the National Academy of Sciences of the United States of America. 2009;106(11):4254–4259. doi: 10.1073/pnas.0813203106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Yauch RL, Gould SE, Scales SJ, et al. A paracrine requirement for hedgehog signalling in cancer. Nature. 2008;455(7211):406–410. doi: 10.1038/nature07275. [DOI] [PubMed] [Google Scholar]
- 48.Yamasaki A, Kameda C, Xu R, et al. Nuclear factor kappaB-activated monocytes contribute to pancreatic cancer progression through the production of Shh. Cancer Immunology, Immunotherapy. 2010;59(5):675–686. doi: 10.1007/s00262-009-0783-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Ray A, Meng E, Reed E, Shevde LA, Rocconi RP. Hedgehog signaling pathway regulates the growth of ovarian cancer spheroid forming cells. International Journal of Oncology. 2011;39(4):797–804. doi: 10.3892/ijo.2011.1093. [DOI] [PubMed] [Google Scholar]
- 50.Jiang J, Hui CC. Hedgehog signaling in development and cancer. Developmental Cell. 2008;15(6):801–812. doi: 10.1016/j.devcel.2008.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Pola R, Ling LE, Silver M, et al. The morphogen Sonic hedgehog is an indirect angiogenic agent upregulating two families of angiogenic growth factors. Nature Medicine. 2001;7(6):706–711. doi: 10.1038/89083. [DOI] [PubMed] [Google Scholar]
- 52.Kanda S, Mochizuki Y, Suematsu T, Miyata Y, Nomata K, Kanetake H. Sonic hedgehog induces capillary morphogenesis by endothelial cells through phosphoinositide 3-kinase. Journal of Biological Chemistry. 2003;278(10):8244–8249. doi: 10.1074/jbc.M210635200. [DOI] [PubMed] [Google Scholar]
- 53.Feldmann G, Dhara S, Fendrich V, et al. Blockade of hedgehog signaling inhibits pancreatic cancer invasion and metastases: a new paradigm for combination therapy in solid cancers. Cancer Research. 2007;67(5):2187–2196. doi: 10.1158/0008-5472.CAN-06-3281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Hay ED. An overview of epithelio-mesenchymal transformation. Acta Anatomica. 1995;154(1):8–20. doi: 10.1159/000147748. [DOI] [PubMed] [Google Scholar]
- 55.Sanchez P, Hernández AM, Stecca B, et al. Inhibition of prostate cancer proliferation by interference with SONIC HEDGEHOG-GLI1 signaling. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(34):12561–12566. doi: 10.1073/pnas.0404956101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Zhu G, Ke X, Liu Q, et al. Recurrence of the D100N mutation in a Chinese family with brachydactyly type A1: evidence for a mutational hot spot in the Indian hedgehog gene. American Journal of Medical Genetics, Part A. 2007;143(11):1246–1248. doi: 10.1002/ajmg.a.31728. [DOI] [PubMed] [Google Scholar]
- 57.Bellahcène A, Castronovo V, Ogbureke KUE, Fisher LW, Fedarko NS. Small integrin-binding ligand N-linked glycoproteins (SIBLINGs): multifunctional proteins in cancer. Nature Reviews Cancer. 2008;8(3):212–226. doi: 10.1038/nrc2345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.McAllister SS, Gifford AM, Greiner AL, et al. Systemic endocrine instigation of indolent tumor growth requires osteopontin. Cell. 2008;133(6):994–1005. doi: 10.1016/j.cell.2008.04.045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Shevde LA, Das S, Clark DW, Samant RS. Osteopontin: an effector and an effect of tumor metastasis. Current Molecular Medicine. 2010;10(1):71–81. doi: 10.2174/156652410791065381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Riker AI, Enkemann SA, Fodstad O, et al. The gene expression profiles of primary and metastatic melanoma yields a transition point of tumor progression and metastasis. BMC Medical Genomics. 2008;1, article 13 doi: 10.1186/1755-8794-1-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Metge BJ, Liu S, Riker AI, Fodstad O, Samant RS, Shevde LA. Elevated osteopontin levels in metastatic melanoma correlate with epigenetic silencing of breast cancer metastasis suppressor 1. Oncology. 2010;78(1):75–86. doi: 10.1159/000292363. [DOI] [PubMed] [Google Scholar]
- 62.Shevde LA, Samant RS, Paik JC, et al. Osteopontin knockdown suppresses tumorigenicity of human metastatic breast carcinoma, MDA-MB-435. Clinical and Experimental Metastasis. 2006;23(2):123–133. doi: 10.1007/s10585-006-9013-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Psailaa B, Kaplana RN, Port ER, Lydena D. Priming the “soil” for breast cancer metastasis: the pre-metastatic niche. Breast Disease. 2006;26(1):65–74. doi: 10.3233/bd-2007-26106. [DOI] [PubMed] [Google Scholar]
- 64.Kaplan RN, Rafii S, Lyden D. Preparing the "soil": the premetastatic niche. Cancer Research. 2006;66(23):11089–11093. doi: 10.1158/0008-5472.CAN-06-2407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Harada SI, Rodan GA. Control of osteoblast function and regulation of bone mass. Nature. 2003;423(6937):349–355. doi: 10.1038/nature01660. [DOI] [PubMed] [Google Scholar]
- 66.Burger EH, Klein-Nulend J, Van der Plas A, Nijweide PJ. Function of osteocytes in bone-their role in mechanotransduction. Journal of Nutrition. 1995;125(7) doi: 10.1093/jn/125.suppl_7.2020S. [DOI] [PubMed] [Google Scholar]
- 67.Teitelbaum SL, Ross FP. Genetic regulation of osteoclast development and function. Nature Reviews Genetics. 2003;4(8):638–649. doi: 10.1038/nrg1122. [DOI] [PubMed] [Google Scholar]
- 68.Edwards CM, Mundy GR. Eph receptors and ephrin signaling pathways: a role in bone homeostasis. International Journal of Medical Sciences. 2008;5(5):263–272. doi: 10.7150/ijms.5.263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Harada M, Shimizu A, Nakamura Y, Nemoto R. Role of the vertebral venous system in metastatic spread of cancer cells to the bone. Advances in Experimental Medicine and Biology. 1992;324:83–92. doi: 10.1007/978-1-4615-3398-6_8. [DOI] [PubMed] [Google Scholar]
- 70.Abeloff AJ, Lichter AS, Niederhuber JE. Clinical Oncology. Elsevier Science; 2000. [Google Scholar]
- 71.Psaila B, Lyden D. The metastatic niche: adapting the foreign soil. Nature Reviews Cancer. 2009;9(4):285–293. doi: 10.1038/nrc2621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Bussard KM, Gay CV, Mastro AM. The bone microenvironment in metastasis; what is special about bone? Cancer and Metastasis Reviews. 2008;27(1):41–55. doi: 10.1007/s10555-007-9109-4. [DOI] [PubMed] [Google Scholar]
- 73.Guise TA, Mohammad KS, Clines G, et al. Basic mechanisms responsible for osteolytic and osteoblastic bone metastases. Clinical Cancer Research. 2006;12(20) doi: 10.1158/1078-0432.CCR-06-1007. [DOI] [PubMed] [Google Scholar]
- 74.Kang Y, Siegel PM, Shu W, et al. A multigenic program mediating breast cancer metastasis to bone. Cancer Cell. 2003;3(6):537–549. doi: 10.1016/s1535-6108(03)00132-6. [DOI] [PubMed] [Google Scholar]
- 75.Yin JJ, Mohammad KS, Käkönen SM, et al. A causal role for endothelin-1 in the pathogenesis of osteoblastic bone metastases. Proceedings of the National Academy of Sciences of the United States of America. 2003;100(19):10954–10959. doi: 10.1073/pnas.1830978100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Mundy GR. Metastasis to bone: causes, consequences and therapeutic opportunities. Nature Reviews Cancer. 2002;2(8):584–593. doi: 10.1038/nrc867. [DOI] [PubMed] [Google Scholar]
- 77.Kesper DA, Didt-Koziel L, Vortkamp A. Gli2 activator function in preosteoblasts is sufficient to mediate Ihh-dependent osteoblast differentiation, whereas the repressor function of Gli2 is dispensable for endochondral ossification. Developmental Dynamics. 2010;239(6):1818–1826. doi: 10.1002/dvdy.22301. [DOI] [PubMed] [Google Scholar]
- 78.Zhao M, Qiao M, Harris SE, Chen D, Oyajobi BO, Mundy GR. The zinc finger transcription factor Gli2 mediates bone morphogenetic protein 2 expression in osteoblasts in response to hedgehog signaling. Molecular and Cellular Biology. 2006;26(16):6197–6208. doi: 10.1128/MCB.02214-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Plaisant M, Fontaine C, Cousin W, Rochet N, Dani C, Peraldi P. Activation of hedgehog signaling inhibits osteoblast differentiation of human mesenchymal stem cells. Stem Cells. 2009;27(3):703–713. doi: 10.1634/stemcells.2008-0888. [DOI] [PubMed] [Google Scholar]
- 80.McKee MD, Nanci A. Osteopontin at mineralized tissue interfaces in bone, teeth, and osseointegrated implants: ultrastructural distribution and implications for mineralized tissue formation, turnover, and repair. Microscopy Research and Technique. 1996;33(2):141–164. doi: 10.1002/(SICI)1097-0029(19960201)33:2<141::AID-JEMT5>3.0.CO;2-W. [DOI] [PubMed] [Google Scholar]
- 81.Blair HC, Robinson LJ, Zaidi M. Osteoclast signalling pathways. Biochemical and Biophysical Research Communications. 2005;328(3):728–738. doi: 10.1016/j.bbrc.2004.11.077. [DOI] [PubMed] [Google Scholar]
- 82.Yoneda T, Sasaki A, Mundy GR. Osteolytic bone metastasis in breast cancer. Breast Cancer Research and Treatment. 1994;32(1):73–84. doi: 10.1007/BF00666208. [DOI] [PubMed] [Google Scholar]
- 83.Das S, Samant RS, Shevde LA. Hedgehog signaling induced by breast cancer cells promotes osteoclastogenesis and osteolysis. Journal of Biological Chemistry. 2011;286(11):9612–9622. doi: 10.1074/jbc.M110.174920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Sterling JA, Oyajobi BO, Grubbs B, et al. The Hedgehog signaling molecule Gli2 induces parathyroid hormone-related peptide expression and osteolysis in metastatic human breast cancer cells. Cancer Research. 2006;66(15):7548–7553. doi: 10.1158/0008-5472.CAN-06-0452. [DOI] [PubMed] [Google Scholar]
- 85.Brubaker KD, Brown LG, Vessella RL, Corey E. Administration of zoledronic acid enhances the effects of docetaxel on growth of prostate cancer in the bone environment. BMC Cancer. 2006;6 doi: 10.1186/1471-2407-6-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Miller RE, Roudier M, Jones J, Armstrong A, Canon J, Dougall WC. RANK ligand inhibition plus docetaxel improves survival and reduces tumor burden in a murine model of prostate cancer bone metastasis. Molecular Cancer Therapeutics. 2008;7(7):2160–2169. doi: 10.1158/1535-7163.MCT-08-0046. [DOI] [PubMed] [Google Scholar]