

Abstract
Some unique subclasses of Camelidae antibodies are devoid of light chain and the antigen binding site is comprised exclusively of the variable domain of the heavy chain (VHH). Although conventional antibodies dominate current assay development, recombinant VHHs have a high potential as alternative reagents for the next generation of immunoassay. We expressed VHHs from an immunized alpaca and developed a VHH-based immunoassay using 3-phenoxybenzoic acid (3-PBA), a major metabolite of pyrethroid insecticides as a model system. A phage VHH library was constructed and seven VHH clones were selected by competitive binding with 3-PBA. The best immunoassay developed with one of these VHHs showed an IC50 of 1.4 ng/mL (limit of detection (LOD) = 0.1 ng/mL). These parameters were further improved by using the phage borne VHH, IC50 = 0.1 ng/mL and LOD = 0.01 ng/mL. Both assays showed a similar tolerance to methanol and dimethylsulfoxide up to 50% in assay buffer. The assay was highly specific to 3-PBA and its 4-hydroxylated derivative, 4-hydroxy 3-PBA (150% cross reactivity) with negligible cross reactivity with other tested structural analogs and the recovery from spiked urine sample ranged from 80 to 112%. In conclusion, a highly specific and sensitive VHH for 3-PBA was developed using sequences from immunized alpaca and phage display technology for antibody selection.
INTRODUCTION
Since the first radioimmunoassay was reported1, countless immunoassays have been developed and proven to be invaluable analytical methods for in vitro diagnostics and environmental monitoring for wide array of substances such as viruses, bacteria, disease biomarkers, food toxins, and environmental pollutants including endocrine disruptors and pesticides and their metabolites2–6. For an immunoassay to be applied to a real sample, it should have high sensitivity and robustness in the matrix in which it is detected. These properties are largely dependent on the availability of antibodies with high affinity and specificity to their target analyte along with a high stability in the matrix. Monoclonal antibodies (MAbs) mostly derived from murine hybridoma cell lines, along with polyclonal antibodies (PAbs) from sera of rabbits, goats, sheep, and other species are traditional reagents used in immunoanalytical techniques.
Conventional antibodies (IgG subclass) have an average molecular weight of 150 kDa, and they are composed of two identical heavy and light chains connected by disulfide bonds. Each antibody contains two antigen binding pockets. Although PAbs can be easily obtained at a low production cost, they are finite, which requires subsequent antibody characterization and assay optimization because of animal to animal variation in immune response. This can be a limiting factor for the use of PAbs in development of an assay for large scale production or commercialization. MAbs obtained from hybridoma cell lines can overcome the reproducibility issues of PAbs. An established hybridoma can produce MAb indefinitely. However MAb technology is expensive, sometimes frozen hybridoma cell lines are hard to recover and generation of high quality MAbs to small molecules is difficult. Because of the size, the requirement for two chains, and sophisticated post-translational modification, functional conventional antibodies are difficult to express recombinantly. Many antibody-derived proteins that are derivatives of an intact antibody molecule have been developed, including monovalent fragments such as Fab, scFv and engineered variants including diabodies, triabodies, minibodies, and single-domain antibodies7–9. However, these constructs often have lower affinity and are less stable than the intact antibody. Production can be problematic because the limited solubility and aggregation of the expressed antibody fragments can reduce yields of expressed proteins10, 11. In addition, the diversity of phage antibody libraries from hybridoma cell lines should be very large to increase the possibility of selecting a desirable antibody fragment due to the high level of non-functional association of VHs and VLs12.
In 1993, Hamers-Casterman discovered a new subclass of antibodies in members of the Camelidae family (i.e. camels, llamas, and alpacas) that have an antigen binding pocket comprised solely of a variable region of the heavy chain and completely devoid of light chains13, 14. Recombinant expression of these heavy chain variable domains yields single domain antibodies (VHHs). The single domain nature of VHHs provides many advantages over the other recombinant antibody fragments; in particular, ease of expression in various expression systems, high thermal stability, excellent solubility, resistance to proteolysis, and ease of genetic manipulation15–17. Antigen-binding patterns of VHHs are very unique compared to conventional antibodies. Since the binding area of a conventional antibody is comprised of a heavy chain and a light chain, they tend to bind to proteins through a large surface area, and they encompass small molecules in a deep cleft formed at an interface of a heavy and a light chain. Because heavy chain antibodies need to compensate for the missing light chain, their complementary determining region 3 (CDR3) is normally longer, which allows the formation of a `finger-like' shape and subsequent binding in a groove of the protein target18. This allows for heavy chain antibodies to be used as enzyme inhibitors19. When binding to small molecules, their binding occurs through a lateral recognition involving the coordinated contribution of the three hyper variable loops and the framework regions (FRs)20–22. It has also been observed that the binding of a VHH against caffeine is accompanied by a hapten-induced dimerization of the VHHs23.
To date, most reports about biotechnological applications of VHHs are related to protein or other macromolecular antigens23, 24, while few reports refer to small-molecules. Some VHH-based assays have been developed for the detection of small haptens, but, with the exception of two VHHs against azo-dye Reactive Red20 and ochratoxin A26, the μM range sensitivities of the assays are somewhat unsatisfactory for practical application to real samples27–32. We recently demonstrated that the generation of VHHs to triclocarban (TCC) is not favored in the llama immune response, but that high-affinity anti-TCC VHHs can be isolated by competitive selection from phage libraries33. The VHH-based ELISA showed IC50 value of 3.5 ng/mL. To further test this hypothesis and evaluate the potential of VHHs for the development of practical applications, we present in this work the isolation of alpaca VHHs to 3-phenoxybenzoic acid (3-PBA) (MW=214), and their use in the immunodetection of this compound in human urine samples. 3-PBA is a human urinary metabolite of pyrethroid insecticides and can be used as a biomarker to monitor human exposure to these pesticides. We have previously developed a PAb-based enzyme-linked immuosorbent assay (ELISA) for 3-PBA with IC50 values of 120 ng/mL for a homologous ELISA and 1.6 ng/mL for a heterologous ELISA34. The same antigen was used in this study to immunize alpacas, and the VHHs were isolated from a library prepared from their peripheral blood lymphocytes, leading to the develop of immunoassays to 3-PBA with equal sensitivity than that obtained with conventional PAbs and excellent recoveries in real samples. Since VHHs can be produced easily and without the batch-to-batch variations of PAbs, the demonstration of their high profile performance in a practical application is an important contribution to the immunoassay technology for small molecules.
EXPERIMENTAL SECTION
A three-year old castrated male alpaca was immunized subcutaneously with 200 μg of 3-PBA hapten-thyroglobulin conjugate mixed with Freund's incomplete adjuvant. The immunization, homologous, and heterologous coating antigens were previously synthesized in this laboratory34 and their structures are presented in figure S-1, supporting information. Total RNA was extracted from peripheral blood lymphocytes isolated from alpaca blood and then used for first strand cDNA synthesis. The phage-VHH library was constructed by cloning PCR amplified VHH genes into the pComb3XSS phagemid vector. Phage anti-3-PBA VHHs were selected using gradually decreasing concentrations of 3-PBA hapten-BSA conjugate and free 3-PBA and more stringent washing. Phage VHH clones that show inhibitory binding to a coating antigen by free 3-PBA were sequenced and soluble VHH fragments was expressed from TOP10F' cells of E. coli transformed with phagemids extracted from clones with unique sequences. ELISAs with isolated phage-VHHs (PELISAs) and soluble VHHs (VELISAs) were used to determine assay sensitivity. The phage-VHH and VHH with the highest sensitivity were further used for the estimation of cross-reactivity and assay validation using human urine samples. For more detailed experimental description, see the supporting information.
RESULTS
Library construction and selection of 3-PBA specific VHHs
The total RNA was extracted from alpaca blood with the leukoLOCK total RNA isolation system (Applied Biosystems, Foster City, CA) following manufacturer's protocols. VHH genes were obtained by PCR using a set of sense primers specific for the 5' region of the heavy chain variable genes paired with a reverse primer specific for either long (IgG2) or short (IgG3) hinge regions. The amplified genes were cloned into the phagemid vector pComb3XSS, which was then electroporated into electrocompetent ER2738 cells from E. coli in order to construct the phage VHH library. The size of the phage VHH library was 3.4×107 colony forming units (c.f.u)/mL.
Figure 1 presents the number of phage output in each panning. The number of phage output decreased after every round of panning except for the second. Over all there was a 10,000-fold decrease in the phage output over 5 panning cycles. The 4th and 5th phage outputs were used to identify positive phage VHH clones for analysis of 3-PBA. Although the number of phage output decreased with decreasing concentrations of the coating antigen and 3-PBA, we observed that 100% of tested clones showed different degrees of binding inhibition by free 3-PBA (data not shown). This result indicates that our panning scheme was very effective in isolating positive VHH clones. We devised a panning strategy that increased the selection pressure throughout the panning and minimized the carryover of nonspecific binders to a next round of panning; preventing the non specific binders from outnumbering the specific ones. We performed five cycles of panning with modulation of the concentration of the 3-PBA hapten-BSA conjugate for capturing phage-VHHs, the washing intensity, the concentration of 3-PBA for the competitive elution of the captured phage-VHHs, and the post absorption of phage eluate to BSA. To capture high affinity phage-VHHs to the coating antigens, the concentration of the coating antigen (0.5, 0.25, 0.1, 0.02, and 0.01 μg per well over the panning) was gradually decreased. We also decreased the concentration of 3-PBA by ten-fold for each subsequent round of panning, to elute phage-VHHs with high affinity to the 3-PBA. To remove non-specific and weakly binding phages we increased the washing intensity in each round of panning, the phage-VHH captured wells were washed five times for the 1st panning and washed ten times for the 2nd panning with PBS containing 0.05% Tween 20 before the competitive elution. For the rest of the pannings, the wells were washed ten times and then the plates were shaken for 30 min while the wells were filled with PBS containing 0.1% Tween 20, which was followed by ten more washings. Non-specific phages were minimized by increasingly stringent washings and post absorption of phage output to BSA.
Figure 1.

Number of phage output. Ten μl of the phage output were diluted in a sterile PBS from each panning cycle and were mixed with 1ml of ER2738 cells with absorbance of 0.4 at O.D600. The mixture was incubated for 25 min at 37 °C without shaking. The mixture was plated on a LB-plate with ampicillin. The number of colonies was counted the following day.
Sequence alignment of antibody fragments to 3-PBA
Sixteen clones after the 4th round of panning and 25 clones from the 5th round of panning that showed an inhibition of binding to the coating antigen by free 3-PBA were sequenced. Three and 4 unique sequences were found from 4th and 5th round of panning, respectively (Figure S-2, supporting information). The main features of heavy chain antibodies that distinguish them from the variable heavy chain domains of conventional IgGs are the occurrence of amino acids, F, E, R and F/G in the FR2, which stabilize the VHH fragments and the extended length of CDR3 regions (15–22 amino acids). The amino acid sequences of the alpaca VHH clones in the region are in agreement with the findings. The deduced amino acid sequences of the VHH clones show these amino acids in FR2. Clone 3P5ThC19 and 3P5ThC15 have G and P in the region instead of E and G. All clones show 18 amino acids in the CDR3 region with no significant amino acid variation.
Competitive ELISA for 3-PBA
The sensitivities of assays were estimated by performing an indirect competitive ELISA with the soluble VHH fragments (Figure 2). The VHHs were expressed from TOP10F' cells transformed with each phagemid vector isolated from positive clones. The TOP10F' is a non-suppressor strain, allowing expression of VHHs without the phage pIII protein. The VHH fragments with 6xHis and HA tags were obtained by purifying cell lysates on the Ni-NTA affinity columns. The size of purified VHHs was determined on a 12% SDS PAGE gel as 17 kDa major bands (data not shown). Prior to the indirect ELISA, the optimization of the coating antigen (3-PBA hapten-BSA) and VHH concentrations were determined by a checker board titration. As Figure 2 shows, the lowest IC50 value (1.4 ng/mL, 0.007 μM) was observed from clone 3P5ThC12 which was isolated after the 5th round of panning. The clone 3P5ThC1 showed a higher IC50 value of 22.8 ng/mL (0.1 μM). Other clones showed similar IC50 values ranged from 8.2 to 15.7 ng/mL (0.04–0.07 μM). Comparing the sensitivity of the VHH-based ELISA (VELISA) with the clone 3P5ThC12 to that of our previously developed PAb-based 3-PBA ELISA, the VELISA is 100-fold more sensitive than the homologous coating antigen-based ELISA (IC50 of 120 ng/mL) and almost identical to the best heterologous coating antigen-based ELISA (IC50 of 1.6 ng/mL). Previously, we increased the sensitivity of the PAb 3-PBA ELISA by use of 3-PBA-BSA heterologous coating antigen by conjugation using the carboxylic acid moiety. We tested if this approach would also increase the sensitivity of the VELISA; however, the VHH showed no recognition to the heterologous coating antigen, indicating that the acid moiety plays a critical role in the binding of the VHH to the 3-PBA. The ELISA using the VHH from 3P5ThC12 clone has at least a several hundred-fold better sensitivities than the most other camelid VHH-based ELISAs reported in literature. Therefore, the VHHs and phage-VHHs derived from 3P5ThC12 clone were used throughout the study.
Figure 2.

Competition ELISA with soluble VHHs. Representative curves showing the inhibition of VHH clones binding to 3-PBA hapten-BSA with increasing amount of the free 3-PBA. 3-PBA hapten-BSA was immobilized and competition mixture was added. After washing, bound VHH was detected with goat anti-HA tag PAb-HRP. Data are represented as an average +/− standard deviation of four replicates.
Cross-reactivity
The specificity of the assay was tested for the clone 3P5ThC12, which exhibited the best IC50 value, using four known pyrethroid metabolites and five pyrethroid parent compounds (Figure 3). The cross-reactivity was determined using the formula, IC50 of 3-PBA ÷ IC50s of cross-reacting compounds × 100. Overall, negligible cross reactivity was observed, except for 4'-hydroxy 3-PBA which showed 150% cross reactivity. Since the immunizing hapten was prepared with the linker attached to –OH group of the 4'-hydroxy-3-PBA, this indicates that the VHH has some interaction with this oxygen atom.
Figure 3.

Cross reactivity. The 96 well plates were coated with 3-PBA hapten-BSA conjugate at 5 ng/well. Serial dilutions of 3-PBA or tested compounds in PBS were mixed with an equal amount of 1/1000 dilution of the VHH in PBS. One hundred μl of the mixture was added into wells. The bound VHH was detected with 1/10000 dilution of goat anti-HA tag PAb-HRP.
Phage-VHH ELISA (PELISA)
The M13 bacteriophage is a filamentous virus with a diameter of 6 nm and length of 0.9 μm containing single strained DNA packed with around 2700 major and several minor coat proteins. When a VHH fragment possesses a good binding affinity, an assay using the phage displayed-VHH can provide advantages over a monomeric soluble expressed VHH fragment. First, the phage can improve assay sensitivity by using it as a multiple binding element, secondly, the phage particle is also adaptable to various detection methods such as immuno-PCR36, 37, and numerous types of biosensors38. Another advantage of the PELISA is that phage-VHH can be easily and inexpensively prepared in high concentrations. Phage-VHH expressed in a culture medium can be readily isolated by simple incubation and centrifugation, eliminating laborious procedures such as cell lysis and Ni-NTA purification necessary for the expression of soluble VHH fragments. Goldman et al.39, 40 evaluated the utility of phage-VHHs derived from llama for ricin and botulinum A employing a sandwich ELISA and Luminex technology. They observed one to two orders of magnitude of sensitivity improvement compared to a standard ELISA with soluble VHHs.
We also evaluated the performance of the PELISA using the phage-displayed VHH of the clone 3P5ThC12 in an indirect competitive ELISA format (Figure 4). The checkerboard titration was used to determine the optimal concentrations of the coating antigen and the phage-VHH using two blocking reagents (1% BSA and 3% skim milk in PBS). As shown in the figure, when the plate was blocked with 1% BSA, the IC50 value was 0.6 ng/mL, showing slightly enhanced sensitivity compared to the VELISA. However, we observed a 6-fold improvement of the assay sensitivity when 3% of skim milk was used for blocking with an IC50 of 0.1 ng/mL which is 10-fold better than the VELISA. To generate a similar maximal signal to the VELISA, the PELISA required the use of coating antigen at 0.4 μg/mL which is 8-fold higher than the concentration for the VELISA. We tested the VELISA using a plate blocked with 3% skim milk, the assay sensitivity was similarly enhanced, but signal variations among replicates were significantly higher (data not shown). Thus, we used 1% BSA blocking for the VELISA.
Figure 4.

Competitive binding of the phage-VHH. A 96-well plate was coated with 3-PBA hapten-BSA conjugate at 0.4 μg/mL. Dilutions of 3-PBA in PBS was mixed with a 1/2500 dilution of the phage-VHH in PBS and 100 μl of the mixture was added into wells. The bound phage-VHHs were detected by adding 100 μl of mouse anti-M13 phage MAb-HRP diluted in PBS at 1/5000.
Solvent effect
High tolerance to solvents is beneficial when developing immunoassays to be used for analysis of lipophilic analytes, because water miscible solvents are commonly used for extraction of analyte prior to analysis. When the matrix effects for solvent extracts are negligible less dilution of the extracts is needed, which results in enhanced assay sensitivity. Although the camelid VHHs have been known to be highly thermally stable, no evaluation has been done for the tolerance of VHHs or phage-displayed VHHs to such water miscible solvents. Hence, we estimated the effect of methanol (MeOH) and dimethylsulfoxide (DMSO) on the performance of the VELISA and PELISA and compared to that of the PAb-based 3-PBA ELISA. Each solvent was added to the assay buffer at concentrations of 0, 10, 25 and 50% (v/v), while maintaining constant ionic strength. Figure 5 presents the MeOH effects on the assay performance. Typically, increasing the concentration of MeOH in ELISAs causes an increase in both the maximal signal and the IC50 values. All assays showed the increase of the maximal signals likewise when the MeOH concentrations increased with more significant increase in the PELISA. Interestingly, the assay sensitivity also was improved for the VELISA and PELISA, which indicates that the addition of MeOH facilitates the binding interaction with minimal effect on the protein function. However, the MeOH showed a more significant effect on the sensitivity of the 3-PBA ELISA showing a small but gradual increase of IC50 values. A four-fold increase in IC50 value was observed at 50% (v/v) of MeOH. This result demonstrated that both the VHH and phage-VHH were highly tolerant to MeOH.
Figure 5.
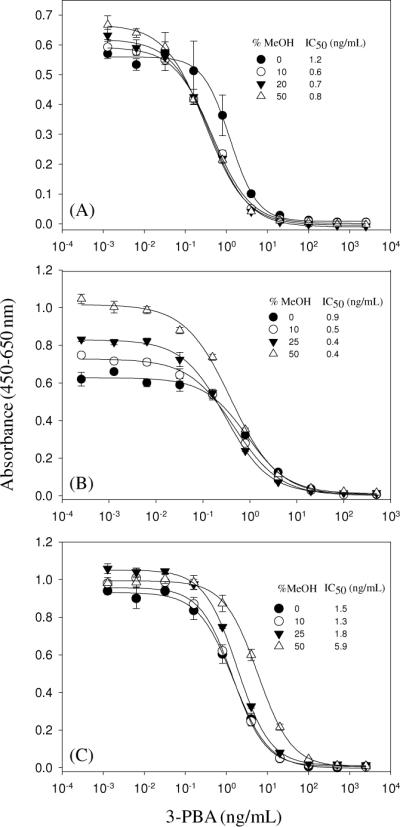
Effects of MeOH on the performance of the VELISA (A), PELISA (B), and 3-PBA ELISA (C). PBS buffers containing MeOH at different concentrations were used to make serial dilutions of 3-PBA. The serial dilutions were mixed with an equal volume of VHH, phage-VHH, or 3-PBA PAb and then 100 μL of the mixture was added into wells. The bound immunoreagents were detected by adding 100 μL of goat anti-HA tag PAb, mouse anti-M13 phage MAb, and goat anti-rabbit PAb conjugated with HRP, respectively. Values are the mean +/− standard deviation of 4 well replicates.
DMSO did not affect the binding of the VHH and the phage-VHH, because IC50 values changed insignificantly in the presence of up to 50% (v/v) of DMSO (Figure 6). In contrast, DMSO had a significant effect in the PAb 3-PBA ELISA causing 50% reduction in maximal signal at 50% (v/v) DMSO with slight increase of IC50s. The above results demonstrate that the VHH and phage-VHH are more tolerant to solvent effect than the PAb ELISA.
Figure 6.
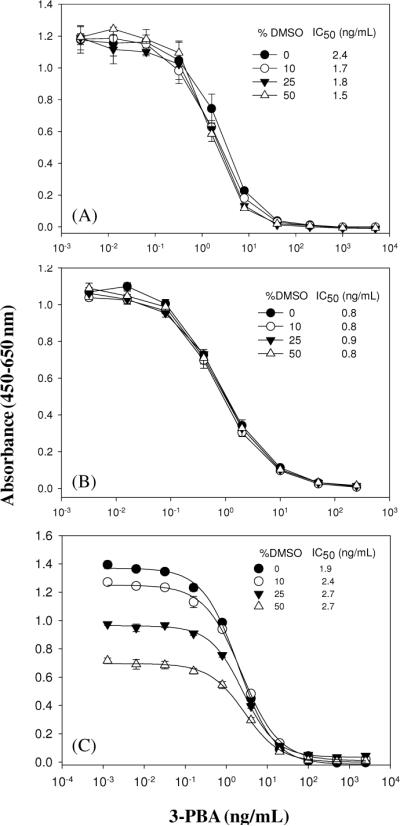
Effects of DMSO on the performance of the VELISA (A), PELISA (B), and 3-PBA ELISA (C). PBS buffers containing DMSO at different concentrations were used to make serial dilutions of 3-PBA. The serial dilutions were mixed with an equal volume of VHH, phage-VHH, or 3-PBA PAb and then 100 μl of the mixture was added into wells. The bound immunoreagents were detected by adding 100 μl of goat anti-HA tag PAb, mouse anti-M13 phage MAb, and goat anti-rabbit PAb conjugated with HRP, respectively. Values are the mean +/− standard deviation of 4 well replicates.
Assay validation
The VELISA and PELISA were validated using a urine sample collected from a person with no known exposure to pyrethroid insecticides. The urine was spiked with 3-PBA at 5 different concentrations. The spiked urine samples were diluted with PBS buffer to a final concentration of urine of 4%. An aliquot of the diluted urine samples was mixed with an equal volume of the VHH or phage-VHH diluted in PBS for competition (Figure 7). The matrix effect test revealed that the urine content in the assay buffer should be less than 2% to remove the matrix interference on the assay (data not shown). In general, both assays showed good correlations between the spiked and recovered concentrations with an average recovery of 80–110%. We also observed minor false positive detection from urine with no 3-PBA. It should be noted that although the dilution of urine samples can be used for the analysis of urine containing concentrations of 3-PBA higher than 100 ng/mL, sample preparation by solid-phase extraction (SPE) or a mixed-mode SPE method is necessary to detect low concentrations of 3-PBA and for more accurate determination in urine samples41. For this, the urine sample was spiked with four low concentrations of 3-PBA (1, 5, 20, and 50 ng/mL) and the analytes were extracted by the mixed-mode SPE method. The recoveries by the VELISA were compared to the recoveries by a gas-chromatography mass spectrometry (GC-MS). We observed an excellent correlation between two methods with VELISA recoveries of 82–110% (Table S-1, supporting information).
Figure 7.

The validation of the VELISA and PELISA in a urine matrix. 3-PBA standard compound was spiked into the urine sample before it was diluted 25-fold with PBS buffer. The dilutions of 3-PBA spiked urine were mixed with an equal volume of VHH or phage-VHH diluted in PBS before analysis.
CONCLUSION
In this case we were able to obtain a homologous VELISA and PELISA that were of equal or greater sensitivity to our best PAb-based heterologous assay for the development of a sensitive assay for 3-PBA using alpaca VHH and phage-VHH fragments. We demonstrated that the alpaca is a good source for the VHH recombinant antibodies leading to small molecular weight compounds. The assays using the VHH and phage-VHH derived from clone 3P5ThC12 showed several hundred times better sensitivity than other assays that have been developed using camelid VHHs for small molecules. This observation indicates that a good panning strategy can be very effective in isolating a high affinity VHH for small haptens starting with alpaca RNA. The sensitivity of the VELISA and PELISA with the homologous coating antigen was approximately 80- and 1200-fold better than the homologous 3-PBA ELISA, respectively. The VELISA showed almost identical sensitivity to our best heterologous ELISA, but 10-fold better sensitivity was observed from the PELISA. Although monoclonal type VHH showed a similar sensitivity to the 3-PBA heterologous assay, the use of heterologous antigens with linkers attached to –OH moiety of the 3-PBA hapten may further improve assay sensitivity. The VHH and phage-VHH showed higher tolerance to MeOH and DMSO than the PAb. The size of a VHH is one tenth of conventional antibodies, and it is much more robust than MAbs and PAbs with high resistance to heat and solvent effects. The size and robustness of the VHH is a significant advantage when developing assays or biosensors, because the stability and the small size allow them to be immobilized on a miniaturized sensor chip in higher density and leads to a longer shelf life of the assay. This feature is particularly significant for field use diagnostics where refrigeration is often limited, solvent-based sample preparation conditions are required, and sample cleanup procedures must be simple. Since the phage-VHH retain the stability and affinity of the VHH, the phage particles are very robust and serve as a multi-binding scaffold for signal enhancement, which increased the sensitivity as we have demonstrated. The phage-VHH was easily obtained by simple procedures. The VELISA and PELISA both showed good recovery of spiked samples in a urine matrix with an average recovery of 80–110%. The dilution of urine sample can be used for urine containing relatively high concentrations of 3-PBA, if more sensitivity is needed samples would have to be extracted prior to analysis. Sample pretreatment by the simple mixed-mode SPE allowed the detection of 3-PBA at very low concentrations. As we have demonstrated here, camelid VHHs will be a useful tool in the future of small molecule immunoassays.
Supplementary Material
ACKNOWLEDGEMENTS
This work was supported in part by the following: The National Institute of Environmental Health Sciences (NIEHS) Superfund Basic Research Program [P42 ES004699]; the NIEHS Children's Environmental Health Center [P01ES011269]; and the Fogarty International Center Grant [TW05718]. The support of the Western Center for Agricultural Health and Safety at the University of California Davis [PHS OH07550] is also acknowledged. BDH is a George and Judy Marcus Senior Follow of the American Asthma Foundation. We thank Bonnie Potter for use of alpacas.
REFERENCES
- (1).Yalow RS, Berson SA. J. Clin. Invest. 1960;39:1157–1175. doi: 10.1172/JCI104130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (2).Iqbal SS, Mayo MW, Bruno JG, Bronk BV, Batt CA, Chambers JP. Biosens. Bioelectron. 2000;15:549–578. doi: 10.1016/s0956-5663(00)00108-1. [DOI] [PubMed] [Google Scholar]
- (3).Wu J, Fu Z, Yan F, Ju H. TrAC, Trends Anal. Chem. 2007;26:679–688. [Google Scholar]
- (4).Li P, Zhang Q, Zhang W. TrAC,Trends Anal. Chem. 2009;28:1115–1126. [Google Scholar]
- (5).Farré M, Kantiani L, Barceló D. TrAC, Trends Anal. Chem. 2007;26:1100–1112. [Google Scholar]
- (6).Morozova VS, Levashova AI, Eremin SA. J. Anal. Chem. 2005;60:202–217. [Google Scholar]
- (7).Kipriyanov SM, Gall FL. Mol. Biotechnol. 2004;26:39–60. doi: 10.1385/MB:26:1:39. [DOI] [PubMed] [Google Scholar]
- (8).Hudson PJ, Souriau C. Nat. Med. 2003;9:129–134. doi: 10.1038/nm0103-129. [DOI] [PubMed] [Google Scholar]
- (9).Yau KYF, Lee H, Hall JC. Biotechnol. Adv. 2003;21:599–637. doi: 10.1016/s0734-9750(03)00104-6. [DOI] [PubMed] [Google Scholar]
- (10).Whitlow M, Filpula D. Methods. 1991;2:97–105. [Google Scholar]
- (11).Kipriyanov S, Dübel. S.; Breitling F, Kontermann RE, Little M. Mol. Immunol. 1994;31:1047–1058. doi: 10.1016/0161-5890(94)90100-7. [DOI] [PubMed] [Google Scholar]
- (12).Vidarsson G, van de Winkel JGJ, van Dijk MA. J. Immunol. Methods. 2001;249:245–252. doi: 10.1016/s0022-1759(00)00337-9. [DOI] [PubMed] [Google Scholar]
- (13).Hamers-Casterman C, Atarhouch T, Muyldermans S, Robinson G, Hammers C, Songa EB, Bendahman N, Hammers R. Nature. 1993;363:446–448. doi: 10.1038/363446a0. [DOI] [PubMed] [Google Scholar]
- (14).Muyldermans S, Baral TN, Cortez Retamozzo V, De Baetselier P, De Genst E, Kinne J, Leonhardt H, Magez S, Nguyen VK, Revets H, Rothbauer U, Stijlemans B, Tillib S, Wernery U, Wyns L, Hassanzadeh-Ghassabeh G, Saerens D. Vet. Immunol. Immunop. 2009;128:178–183. doi: 10.1016/j.vetimm.2008.10.299. [DOI] [PubMed] [Google Scholar]
- (15).Harmsen MM, De Haard HJ. Appl. Microbiol. Biot. 2007;77:13–22. doi: 10.1007/s00253-007-1142-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (16).Arbabi Ghahroudi M, Desmyter A, Wyns L, Hamers R, Muyldermans S. FEBS Letters. 1997;414:521–526. doi: 10.1016/s0014-5793(97)01062-4. [DOI] [PubMed] [Google Scholar]
- (17).Beekwilder J, van Houwelingen A, van Beckhoven J, Speksnijder A. Eur. J. Plant Pathol. 2008;121:477–485. [Google Scholar]
- (18).De Genst E, Silence K, Decanniere K, Conrath K, Loris R, Kinne J, Muyldermans S, Wyns L. Proc. Natl. Acad. Sci. USA. 2006;103:4586–4591. doi: 10.1073/pnas.0505379103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (19).Buelens K, Hassanzadeh-Ghassabeh G, Muyldermans S, Gils A, Declerck PJ. J. Thromb. Haemost. 2010;8:1302–1312. doi: 10.1111/j.1538-7836.2010.03816.x. [DOI] [PubMed] [Google Scholar]
- (20).Spinelli S, Frenken LGJ, Hermans P, Verrips T, Brown K, Tegoni M, Cambillau C. Biochemistry. 2000;39:1217–1222. doi: 10.1021/bi991830w. [DOI] [PubMed] [Google Scholar]
- (21).Spinelli S, Tegoni M, Frenken L, van Vliet C, Cambillau C. J. Mol. Biol. 2001;311:123–129. doi: 10.1006/jmbi.2001.4856. [DOI] [PubMed] [Google Scholar]
- (22).Fanning SW, Horn JR. Protein sci. 2011;20:1196–1207. doi: 10.1002/pro.648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (23).Sonneson GJ, Horn JR. Biochemistry. 2009;48:6693–6695. doi: 10.1021/bi900862r. [DOI] [PubMed] [Google Scholar]
- (24).Deffar K, Shi H, Li L, Wang X, Zhu X. Afr. J. Biotechnol. 2009;8:2645–2652. [Google Scholar]
- (25).Saerens D, Hassanzadeh-Ghassabeh G, Muyldermans S. Curr. Opin. Pharmacol. 2008;8:600–608. doi: 10.1016/j.coph.2008.07.006. [DOI] [PubMed] [Google Scholar]
- (26).Van Houwelingen A, Saeger SD, Rusanova T, Waalwijk C, Beekwilder J. World Mycotoxin J. 2008;1:407–417. [Google Scholar]
- (27).Ladenson RC, Crimmins DL, Landt Y, Ladenson JH. Anal. Chem. 2006;78:4501–4508. doi: 10.1021/ac058044j. [DOI] [PubMed] [Google Scholar]
- (28).Sheedy C, Yau KYF, Hirama T, MacKenzie CR, Hall JC. J. Agric. Food Chem. 2006;54:3668–3678. doi: 10.1021/jf060219i. [DOI] [PubMed] [Google Scholar]
- (29).Yau KYF, Groves MAT, Li S, Sheedy C, Lee H, Tanha J, MacKenzie CR, Jermutus L, Hall JC. J. Immunol. Methods. 2003;281:161–175. doi: 10.1016/j.jim.2003.07.011. [DOI] [PubMed] [Google Scholar]
- (30).Stewart CS, MacKenzie CR, Hall JC. Toxicon. 2007;49:699–709. doi: 10.1016/j.toxicon.2006.11.023. [DOI] [PubMed] [Google Scholar]
- (31).Alvarez-Rueda N, Behar G, Ferré V, Pugnière M, Roquet F, Gastinel L, Jacquot C, Aubry J, Baty D, Barbet J, Birklé S. Mol. Immunol. 2007;44:1680–1690. doi: 10.1016/j.molimm.2006.08.007. [DOI] [PubMed] [Google Scholar]
- (32).Doyle PJ, Arbabi-Ghahroudi M, Gaudette N, Furzera G, Savard ME, Gleddie S, McLean MD, Mackenzie CR, Hall JC. Mol. Immunol. 2008;45:3703–3713. doi: 10.1016/j.molimm.2008.06.005. [DOI] [PubMed] [Google Scholar]
- (33).Tabares-da Rosa S, Rossotti M, Carleiza C, Carriόn F, Pritsch O, Ahn KC, Last JA, Hammock BD, González-Sapienza G. Anal. Chem. 2011;83:7213–7220. doi: 10.1021/ac201824z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (34).Shan G, Huang H, Stoutamire DW, Gee SJ, Leng G, Hammock BD. Chem. Res. Toxicol. 2004;17:218–225. doi: 10.1021/tx034220c. [DOI] [PubMed] [Google Scholar]
- (35).Maass DR, Sepulveda J, Pernthaner A, Shoemaker CB. J. Immunol. Methods. 2007;324:13–25. doi: 10.1016/j.jim.2007.04.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (36).Guo Y-C, Zhou Y-F, Zhang X-E, Zhang Z-P, Qiao Y-M, Bi L-J, Wen J-K, Liang MF, Zhang JB. Nucleic Acids Res. 2006;34:e62. doi: 10.1093/nar/gkl260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (37).Kim H-J, McCoy M, Gee SJ, Gonzalez-Sapienza G, Hammock BD. Anal. Chem. 2011;83:246–253. doi: 10.1021/ac102353z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (38).Mao C, Liu A, Cao B. Angew. Chem. Int. Ed. 2009;48:6790–6810. doi: 10.1002/anie.200900231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (39).Goldman ER, Anderson GP, Bernstein RD, Swain MD. J. Immunol. Methods. 2010;352:182–185. doi: 10.1016/j.jim.2009.10.014. [DOI] [PubMed] [Google Scholar]
- (40).Goldman ER, Liu JL, Bernstein RD, Swain MD, Mitchell SQ, Anderson GP. Sensors. 2009;9:542–555. doi: 10.3390/s90100542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (41).Ahn KC, Lohstroh P, Gee SJ, Gee NA, Lasley B, Hammock BD. Anal. Chem. 2007;79:8883–8890. doi: 10.1021/ac070675l. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.