

Abstract
A tetrazolium-based colorimetric selective assay (MTT-based CSA) was developed to assess the selectivity of antimalarial drugs. This in vitro assay, unlike all others, measures the ability of drugs to indirectly protect red blood cells (RBCs) from Plasmodium-falciparum-induced destruction. Optimum incubation time and number of cells needed were 5 days and 23 × 106 RBCs per well, respectively. A parasitemia range of 0.375% to 3% was found to be suitable for this assay. The MTT-based CSA determined anti-P. falciparum strain DD2 activity of chloroquine at a higher 50% effective concentration (EC50) value (21.0 μg/mL) than the isotopic microtest (10.0 μg/mL). Artesunate and oxytetracycline achieved 90% effect against DD2 with minimal or no toxicity to RBCs. Against chloroquine sensitive strain 3D7, chloroquine and Alchornea cordifolia had EC50 values of 0.025 μg/mL and 4.9 μg/mL respectively, and selective index (SI) values of >2,000 and >69.4 μg/mL, respectively.
1. Introduction
The threat of malaria has continued to plague millions of people living in most tropical countries despite numerous strategies to bring this disease under control. Chloroquine used to be the first drug of choice for the treatment of P. falciparum infections in Ghana and many African countries, but the emergence of drug-resistant strains has necessitated intensive search for new drugs [1–3]. Presently, Artesunate-Amodiaquine and Artesunate-Lumefantrine combinations are first-choice drugs in Ghana. The standard World Health Organization (WHO) method for testing new antimalarial drugs involves tedious microscopic parasite counting method (MPCM). Other in vitro methods are haem polymerization inhibition assay (HPIA) [4], lactate dehydrogenase assay (LDHA) [5, 6] and the isotopic microtest (IMT) [7]. All these in vitro tests measure some activity of the parasite and the endpoints are directly parasite related. Destruction of red blood cells (RBCs) is the end result of malarial infection. None of the available in vitro assays uses RBCs-related activity to determine endpoints. Furthermore, the available in vitro assays do not test for drug selectivity unless a separate assay on RBC toxicity is done. In this paper, we report on a tetrazolium-based colorimetric selective assay (MTT-based CSA) that uses indirect protection of RBCs from parasite destruction as an endpoint for measuring the effect of drugs on parasites and also determines the possible drug toxicity to the RBCs.
2. Materials and Methods
2.1. Drugs and Plant Extracts
Chloroquine diphosphate (Sigma CQ) was obtained from SIGMA, St Louis, Mo, USA. Also tested in comparison were formulated chloroquine (Troge Medical GmBH, Hamburg, Germany), formulated chloroguanide (proguanil) (Zeneca Pharmaceuticals, UK), formulated artesunate (Guilin Pharmaceutical Works, China), and formulated oxytetracycline (Bimeda, Ireland) purchased from a local pharmacy. Plant extracts GHX-26F and GHX-6L are aqueous extracts from the fruits of Alchornea cordifolia and the leaves of Ficus polita respectively. The aqueous extracts were freeze-dried. Both plants are effective against human immunodeficiency virus in vitro [8] and are suspected to have antiprotozoal activities. All drugs and plant extracts were dissolved in culture medium and filtered through sterile 0.45 μm filters.
2.2. Parasites
Strains 3D7 (chloroquine sensitive) and DD2 (chloroquine resistant) of P. falciparum were obtained from Dr. Jorgehn Kurtzhals, Center for Medical Parasitology, University of Copenhagan, Denmark.
2.3. Cell Cultures
P. falciparum infected and uninfected red blood cells (RBCs) were cultured in RPMI 1640 (Gibco, Parsley, UK) supplemented with 20 mmol/mL L-glutamine (Sigma-Aldrich Co. Ltd., UK), 20 μg/mL gentamicin and 2.5 μg/mL fungizone (Gibco, Parsley, UK), 10% albumax II (Gibco BRL, Grand Island, NY, USA), or 10% normal human AB serum (Sigma-Aldrich Co. Ltd., UK). The cultures were flushed with a gas mixture consisting of 3% O2, 6% CO2, and 91% N2, sealed and incubated at 37°C. For experiments in 96-well microtitre plates, the cultures were put in a desiccator with a lit candle and nearly completely covered. As soon as the candle went off, the desiccator was completely closed.
2.4. Determination of Optimum RBC Number and Incubation Time
P. falciparum-strain-3D7-infected (1% parasitemia) and uninfected cultures were incubated as described above and observed daily for color differences between the two cultures. The parasite-infected culture started becoming visually pale from day 3. The difference in color between the infected and uninfected cultures increased up to day 5. There was no change in the observed paleness of the infected culture from day 5 to day 6. Day 5 was therefore considered as the probable optimum incubation time for visually determined optimum parasite-induced damage to RBCs. The number of RBCs in culture that would give maximum difference between optical densities (ODs) of uninfected and P. falciparum-infected RBCs was then determined as follows. The numbers of healthy RBCs in uninfected and parasite-infected stock cultures were determined by the trypan blue exclusion method and adjusted to be the same per milliliter. Twofold serial dilutions of stock cell cultures (uninfected or 5% infected) were made and 100 μL was put in wells of 96-well microtitre plates that already contained 100 μL of growth medium bringing the total volume per well to 200 μL. Each cell dilution was put in six replicate wells. The cultures were incubated in a desiccator as described above and after 5 days of incubation, cell viability was determined by the MTT-based colorimetric method as previously described [9]. Twenty microliters (20 μL) of MTT (7.5 mg/mL) was added to each well and the plates incubated at 37°C for another 2 h. One hundred and fifty microliters (150 μL) of medium was then carefully taken from each well and discarded. Two hundred microliters (200 μL) of 10% Triton X-100 in acidified isopropanol was added to each well in order to dissolve any formed formazan. With the aid of a multichannel micropipettor, the contents of each well were mixed several times, and the plates kept at room temperature in the dark for 24 h. The optical densities of the wells were read by Emax Precision Microplate reader (Molecular Devices Corporation, Menlo Park, Calif, USA) at 565 nm and 690 nm. Readings at 690 nm were automatically subtracted from readings at 565 nm by the microplate reader. The experiment was repeated once. The averages of all readings were recorded for each dilution. A graph of cell number versus differences in OD readings of uninfected and infected wells was then plotted to determine the optimum cell number that will give the highest OD reading difference between uninfected and infected wells.
2.5. Determination of Optimum Parasitemia
Approximately, 23 × 106 cells per well were determined to give optimum difference in OD readings between uninfected and P. falciparum infected cells in the above experiment. Twenty-three million (23 × 106) cells infected at various parasitemia ranging from 0% to 6% were put into wells of 96-well microtitre plates and incubated at 37°C for 5 days. Each parasitemia was tested at 6 replicates. The cells were processed for MTT-based colorimetric assay as described above. The average OD readings at various levels of parasitemia were subtracted from the average OD reading of control uninfected wells and the values plotted against % parasitemia.
2.6. Tetrazolium-Based Colorimetric Selective Assay (MTT-Based CSA)
From the results of the foregoing experiments, optimum time of incubation, optimum number of RBCs, and optimum parasitemia were determined to be 5 days, 23 × 106 cells per well, and 3% parasitemia, respectively. The drug inhibition experiments therefore utilized these three values in the testing of chloroquine, chloroguanide (proguanil), artesunate, oxytetracycline, and two plant extracts against P. falciparum infection in RBC cultures. In the case of Sigma CQ, 1.5% parasitemia was used. Drug or plant extract was serially diluted, and a 100 μL of each dilution was put into six wells of 96-well microtitre plates. One hundred microliters (100 μL) of uninfected RBCs was added to three replicate drug dilutions. One hundred microliters (100 μL) of infected RBCs was added to three other replicate drug dilutions. Drug-medium controls in the top and bottom outer wells were included in the experiments. The cultures were incubated in a desiccator as described in the previous experiments. The MTT-based colorimetric test was then performed on the cultures. The percent protection of RBCs from malaria parasite destruction by different drugs or plant extracts concentrations were determined by the following formula:
| (1) |
where (ODT)Pf is the average OD measured for a given concentration of the test drug in P. falciparum-infected RBCs; (ODT)mock is the average OD measured for a given concentration of test drug in uninfected RBCs; (ODC)mock is the average OD measured for the control untreated uninfected RBCs; (ODC)Pf is the average OD measured for the control untreated P. falciparum-infected RBCs. The average percent RBC protection values for each three replicates were plotted against the concentrations of drugs and the concentrations needed to achieve 50% and 90% RBC protection, that is, effective concentrations (EC50 and EC90) determined by regression analysis. The percent RBC survival determined the extent of drug toxicity and was calculated by the following formula:
| (2) |
where (ODT)cf is the average OD measured for cell-free treated wells; and (ODC)cf is the average OD measured for cell-free untreated wells. These values were plotted against the concentrations of the drugs and the concentrations needed to reduce RBC survival by 50%, that is, 50% cytotoxic concentrations (CC50) determined by regression analysis. Anti-P. falciparum indices or selective indices (SI) were then calculated as CC50/EC50.
2.7. Isotopic Microtest (IMT) Assay
The effect of Sigma CQ on P. falciparum strain DD2 was tested in the IMT assay concurrently as in the MTT-based CSA. In the IMT assay, the incubation was interrupted after 24 hours for the addition of 0.5 μCi of 3H-hypoxanthine, and incubation continued for additional 24 hours before measurement of 3H-hypoxanthine uptake as published before [7]. The percent inhibition was plotted against concentration of drug and the EC50 and EC90 values determined by regression analysis.
3. Results
3.1. Optimum Incubation Time and Cell Number
By visual examination, 5 days of incubation was found to give maximum lysis in infected RBCs with very little cell death in uninfected control cultures. Figure 1(a) shows the effect of number of RBCs on the difference in OD readings between infected and uninfected RBCs. The cell number that produced the highest difference in OD readings between infected and uninfected RBCs was about 23 × 106 per well.
Figure 1.
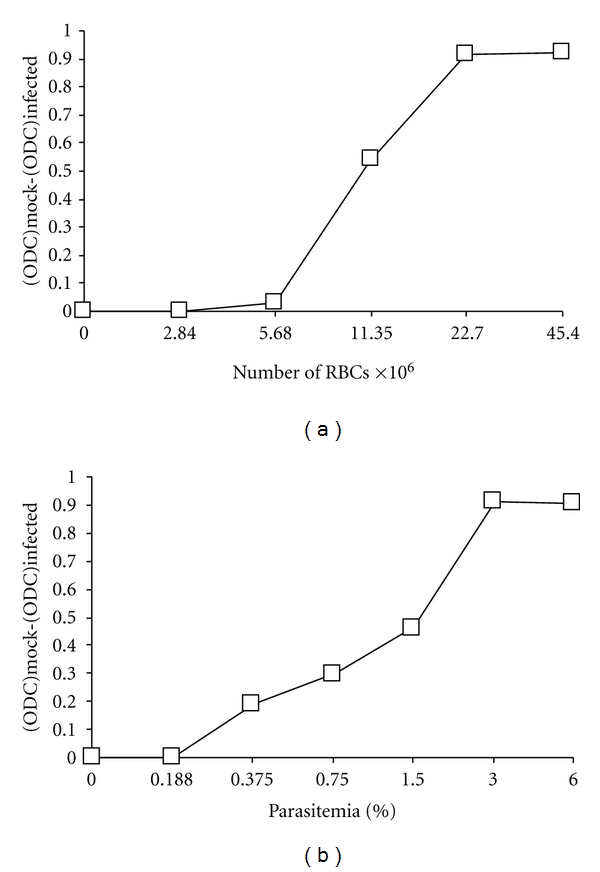
Effects of number of red blood cells (a) and parasitemia (b) on P. falciparum cytopathicity, that is, optical density of mock-infected RBCs minus optical density of parasite-infected RBCs [(ODC)mock − (ODC)infected]. Infection was done with strain 3D7.
3.2. Optimum Parasitemia
Figure 1(b) shows the effect of parasitemia on the ability of RBCs to metabolize tetrazolium to formazan. A parasitemia of 3% caused the optimum damage to RBCs leading to high OD reading difference between uninfected and infected RBCs of 0.92. The lowest parasitemia at which enough damage to RBCs could be detected was found to be 0.375% where the OD reading difference between uninfected and infected RBCs was 0.2.
3.3. Comparative Activities of Drugs and Plant Extracts against DD2 and 3D7 Strains of P. falciparum
The antimalarial activity of chloroquine diphosphate obtained from Sigma was tested against strain DD2 in both the isotopic IMT and the MTT-based CSA and the results are given in Figure 2 and Table 1. Sigma CQ was slightly more active in the IMT than in the MTT-based CSA. The IMT unlike the MTT-based CSA, does not include a toxicity test and is therefore not applicable to the determination of selective index of a drug. In the MTT-based CSA, formulated CQ (Figure 3(a) and Table 1) like Sigma CQ had moderate activity against this strain of P. falciparum. Proguanil (Figure 4(a)) and plant extract GHX-26F (Figure 3(b) and Table 1) were minimally effective against DD2. Formulated artesunate (Figure 4(b)) and oxytetracycline (Figure 4(c)) were very effective against DD2. Formulated CQ and plant extract GHX-26F were very effective in the MTT-based CSA against chloroquine sensitive 3D7 with SI (CC50/EC50) values of >2,000 and >69.4, respectively (Table 1). Figure 5 is from a separate experiment done for pictures to be taken to demonstrate that the effects of drugs can be observed visually in the MTT-based CSA. Clearly, formulated CQ and GHX-26F were very effective against strain 3D7, whereas GHX-6L had no activity.
Figure 2.
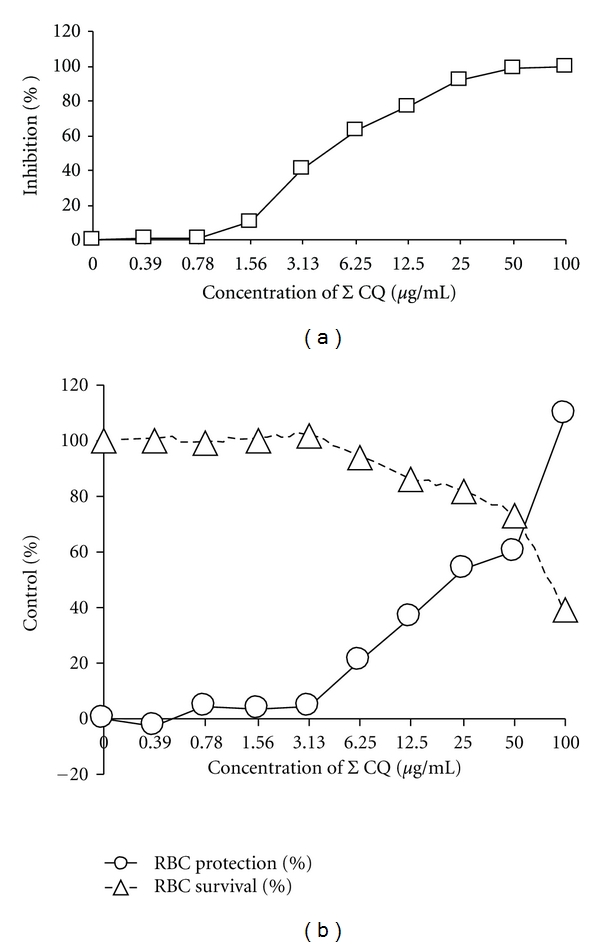
Anti-P. falciparum activity and toxicity of chloroquine (purchased from Sigma) tested in the IMT (a) and MTT-based CSA (b) using parasite strain DD2.
Table 1.
Effects of drugs and plant extracts against P. falciparum strains DD2 (A) and 3D7 (B) infection in RBCs.
| Drug | EC50 μg/mL | EC90 μg/mL | CC50 μg/mL | SI (CC50/EC50) | |
|---|---|---|---|---|---|
| (A) | Sigma CQa∗ | 10.0 | 20.4 | NA | NA |
| Sigma CQb∗ | 21.0 | 77.9 | 82.7 | 3.9 | |
| Formulated CQb | 32.6 | >50c | >50c | >1.5 | |
| GHX-6Lb | >445c | >445c | >445c | ND | |
| GHX-26Fb | 272.8 | >340c | >340c | >1.2 | |
| (B) | Formulated CQb | 0.025 | 0.05 | >50c | >2,000 |
| GHX-6Lb | >445c | >445c | >445c | ND | |
| GHX-26Fb | 4.9 | 16.2 | >340c | >69.4 |
aTested by IMT, bTested by MTT-based CSA, *Parasitemia used was 1.5%. For the other tests, parasitemia used was 3.0%, cHighest concentration tested, SI: selective index, NA: not applicable, ND: not determined.
Figure 3.
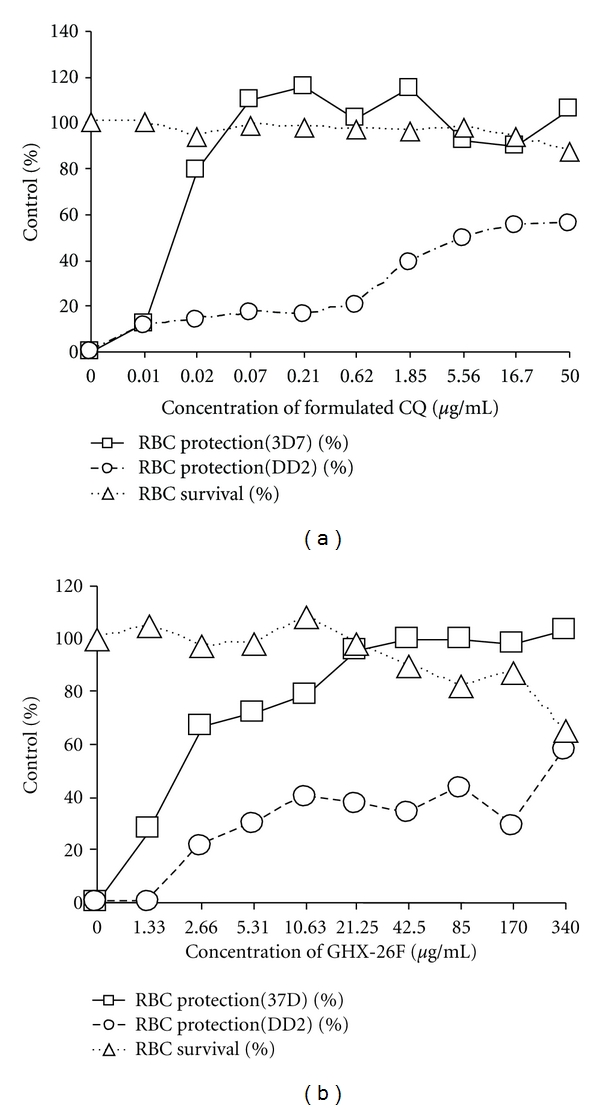
Anti-P. falciparum activity and toxicity of formulated chloroquine (a) and plant extract GHX-26F (b) tested in the MTT-based CSA using parasite strains 3D7 and DD2.
Figure 4.
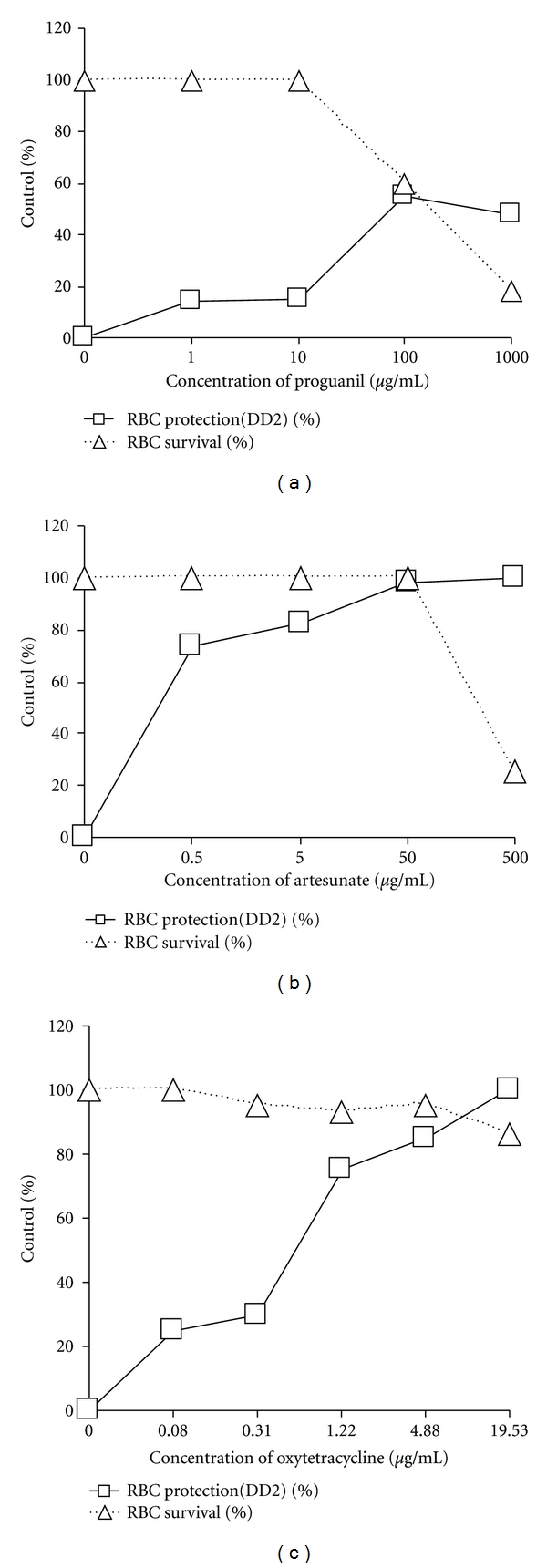
Anti-P. falciparum activity and toxicity of formulated proguanil (a), artesunate (b), and oxytetracycline (c) tested in the MTT-based CSA using parasite strain DD2.
Figure 5.
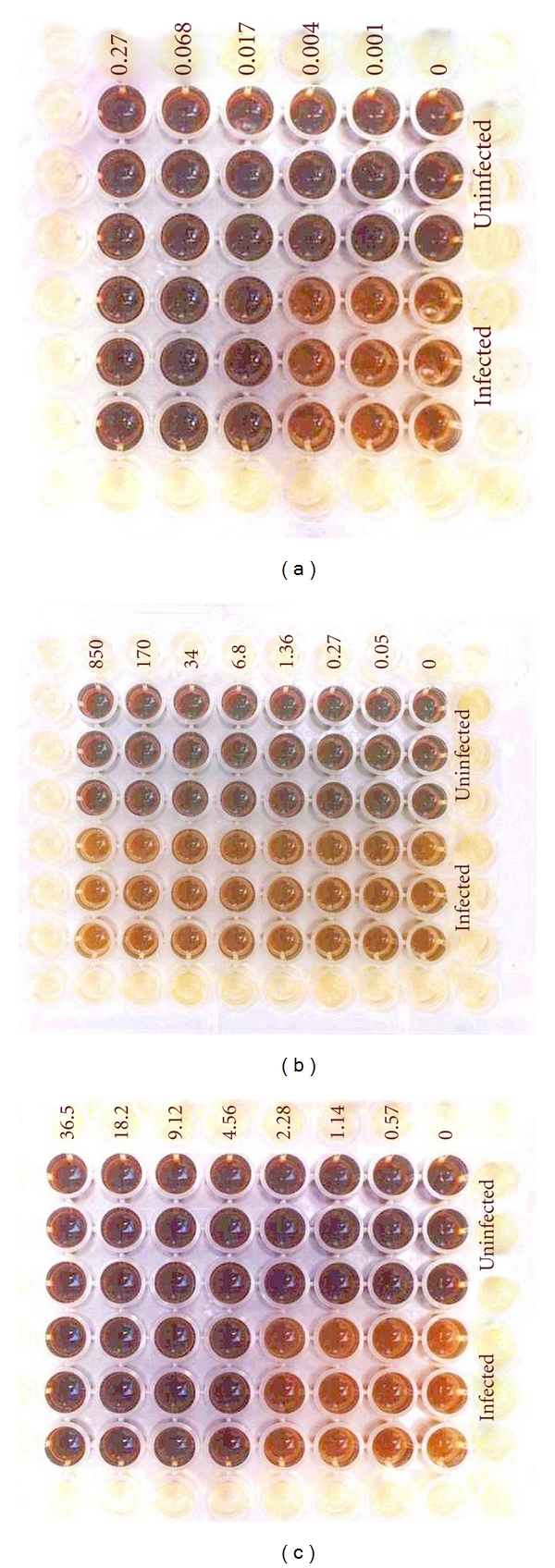
Photographs of parasite strain 3D7-infected (bottom three replicates) and -uninfected (top three replicates) RBCs treated with formulated chloroquine (a), plant extract GHX-6L (b), and plant extract GHX-26F (c) after the tetrazolium reaction. Concentrations are given as μg/mL.
4. Discussion
Determination of in vitro selectivity is crucial in identifying new compounds with potentials as true antimalarial drugs. There is no available in vitro antimalarial assay system that determines drug selectivity. The term selectivity of action in this paper refers to the extent to which the effect of drug is due to parasite events alone and not an indirect effect resulting from effects on uninfected RBCs. This is very important because a compound may destroy RBCs and therefore prevent malaria parasite replication which will lead to false-positive results. All available assays measure parasite-related activity. Our MTT-based CSA measures RBC-related activity of metabolism of tetrazolium into formazan. The end result is a measure of the ability of a compound to indirectly protect RBCs from P. falciparum-mediated lysis. The formula for calculating the percent RBC protection is the same as published for antihuman immunodeficiency drugs [9]. The formula for calculating the percent RBC survival (toxicity) has however been modified to account for plant extracts and other drugs whose color may increase the background OD reading. The maximum difference in OD values between uninfected and infected RBCs may vary depending on the source of RBCs used as a result of differences in certain genetic factors involved in drug metabolism.
We used a specific number of RBCs instead of a specific hematocrit because the formation of formazan is dependent on healthy and metabolically active RBCs. Even though approximately 23 × 106 RBCs/well was determined to give the maximum difference between OD readings of mock-infected and parasite-infected RBCs, lower numbers of RBCs that give OD reading difference of at least 0.2 can be considered as a minimum requirement (Figure 1(a)). A similar case can be made for the parasitemia where reliable determinations can be made using at least 0.375% parasitemia (Figure 1(b)). Chloroquine diphosphate (SIGMA CQ) was tested in the IMT and MTT-based CSA using 1.5% parasitemia similar to other tests. For the other drugs, 3% parasitemia was used in order to be able to clearly visualize the effect of compounds before and after the addition of tetrazolium without OD reading by microplate reader. The choice of not changing the medium during the 5 days of incubation was to allow for the death of the parasites so that only the metabolic activity of the RBCs would be determined by the MTT-based assay.
As expected, chloroquine was not very effective against P. falciparum strain DD2 when tested in either the IMT or MTT-based CSA. There was very little difference between unformulated chloroquine bought from SIGMA and formulated chloroquine bought from a local pharmacy even though the latter was tested at a higher parasitemia. Proguanil, artesunate, and oxytetracycline were tested just to determine their effectiveness in inhibiting chloroquine-resistant DD2 induced RBC lysis. No attempt was made to find EC50 concentrations of these dugs since only four concentrations of each drug were tested. Proguanil needs to be activated to a triazine derivative in vivo to have any significant effect on P. falciparum infection [10]. The minimal activity of proguanil observed against chloroquine resistant strain DD2 may be due to the little intrinsic activity of the drug. Artesunate and oxytetracycline were very effective against DD2 at concentrations with very little or no toxicity to RBCs. Artesunate is a peroxide antimalarial drug which is activated by haem and through a series of processes, it is suspected to yield reactive species capable of alkylating key cellular components and that cause parasite death [11, 12]. Oxytetracycline inhibits protein synthesis and was expected to be effective against DD2 strain as confirmed in our results.
The use of RBC-related activity to determine endpoint drug effect makes it inappropriate to compare our assay with other available in vitro assays. Nevertheless, it would suffice to say that our assay has the advantage of determining the selectivity of drug action. We have used the MTT-based CSA to show that one of two plants suspected to have antiprotozoal activity, that is, Alchornea cordifolia, is indeed very effective against chloroquine-sensitive 3D7 with a high-selective index. The development of an in vitro assay that determines the selectivity of action would be even more important for malarial parasite infection in RBCs from people with glucose-6-phosphate dehydrogenase (G-6-PD) deficiency or those with congenital deficiency of nicotinamide adenine dinucleotide (NADH) methemoglobin reductase in whom antimalarial drug toxicity is of paramount concern. One drawback that is common for all in vitro testing is that the potential new drug may need liver enzyme metabolism to the active form. An additional step of incubating the compound with liver extract before testing will remedy this drawback. Whereas existing in vitro antimalarial tests may be faster and reliable for drugs with known toxicity profiles, our MTT-based CSA stands out as providing results on the selectivity of action of potential new antimalarial drugs thereby allowing for early decision making for further development. Finally, using the extent of RBCs destruction to measure the endpoint gives us a much clearer assessment of what is likely to occur in vivo.
5. Conclusions
A tetrazolium-based selective colorimetric assay was found to be suitable in identifying potential new antimalarial drugs. With the inclusion of toxicity studies in this assay, antimalarial indices can be calculated to make sure that new compounds are truly effective against the parasite. The plant extract Alchornea cordifolia was shown to be very effective against P. falciparum strain 3D7.
Acknowledgments
The authors thank Drs. Michael Aidoo (Centers for Disease Control, Atlanta, USA), Vincent Tsiagbe (Department of Pathology, New York University Medical School, USA), Isabella Quakye (School of Public Health, University of Ghana, Ghana), and Bill Rogers (Epidemiology Department, Noguchi Memorial Institute for Medical Research, University of Ghana, Ghana) for reading through this paper and making valuable suggestions. This project was supported by the Government of Ghana.
References
- 1.White NJ. Antimalarial drug resistance: the pace quickens. Journal of Antimicrobial Chemotherapy. 1992;30(5):571–585. doi: 10.1093/jac/30.5.571. [DOI] [PubMed] [Google Scholar]
- 2.van Es HHG, Skamene E, Schurr E. Chemotherapy of malaria: a battle against all odds? Clinical and Investigative Medicine. 1993;16(4):285–293. [PubMed] [Google Scholar]
- 3.Warhurst DC. Drug-resistant malaria: laboratory and field investigations. Journal of Pharmacy and Pharmacology. 1997;49(supplement 2):3–7. [Google Scholar]
- 4.Slater AFG, Cerami A. Inhibition of chloroquine of a novel haem polymerase enzyme activity in malaria trophozoites. Nature. 1992;355(6356):167–169. doi: 10.1038/355167a0. [DOI] [PubMed] [Google Scholar]
- 5.Makler MT, Ries JM, Williams JA, et al. Parasite lactate dehydrogenase as an assay for Plasmodium falciparum drug sensitivity. American Journal of Tropical Medicine and Hygiene. 1993;48(6):739–741. doi: 10.4269/ajtmh.1993.48.739. [DOI] [PubMed] [Google Scholar]
- 6.Delhaes L, Lazaro JE, Gay F, Thellier M, Danis M. The microculture tetrazolium assay (MTA): another colorimetric method of testing Plasmodium falciparum chemosensitivity. Annals of Tropical Medicine and Parasitology. 1999;93(1):31–40. doi: 10.1080/00034989958771. [DOI] [PubMed] [Google Scholar]
- 7.Desjardins RE, Canfield CJ, Haynes JD, Chulay JD. Quantitative assessment of antimalarial activity in vitro by a semiautomated microdilution technique. Antimicrobial Agents and Chemotherapy. 1979;16(6):710–718. doi: 10.1128/aac.16.6.710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ayisi NK, Nyadedzor C. Comparative in vitro effects of AZT and extracts of Ocimum gratissimum, Ficus polita, Clausena anisata, Alchornea cordifolia, and Elaeophorbia drupifera against HIV-1 and HIV-2 infections. Antiviral Research. 2003;58(1):25–33. doi: 10.1016/s0166-3542(02)00166-3. [DOI] [PubMed] [Google Scholar]
- 9.Ayisi NK, Gupta SV, Qualtiere LF. Modified tetrazolium-based colorimetric method for determining the activities of anti-HIV compounds. Journal of Virological Methods. 1991;33(3):335–344. doi: 10.1016/0166-0934(91)90033-v. [DOI] [PubMed] [Google Scholar]
- 10.Schmidt LH, Loo TL, Fradkin R, Hughes HB. Antimalarial activities of triazine metabolites of chlorguanide and dichlorguanide. Proceedings of the Society for Experimental Biology and Medicine. 1952;80(2):367–370. doi: 10.3181/00379727-80-19625. [DOI] [PubMed] [Google Scholar]
- 11.Meshnick SR, Taylor TE, Kamchonwongpaisan S. Artemisinin and the antimalarial endoperoxides: from herbal remedy to targeted chemotherapy. Microbiological Reviews. 1996;60(2):301–315. doi: 10.1128/mr.60.2.301-315.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Cumming JN, Ploipradith P, Posner GH. Antimalarial activity of artemisinin (quinghaosu) and related triazines: mechanism(s) of action. Advances in Pharmacology. 1996;37:253–296. doi: 10.1016/s1054-3589(08)60952-7. [DOI] [PubMed] [Google Scholar]