

Abstract
Candida albicans, the single most frequently isolated human fungal pathogen, was thought to be asexual until the recent discovery of the mating-type-like locus (MTL). Homozygous MTL strains were constructed and shown to mate. Furthermore, it has been demonstrated that opaque-phase cells are more efficient in mating than white-phase cells. The similarity of the genes involved in the mating pathway in Saccharomyces cerevisiae and C. albicans includes at least one gene (KEX2) that is involved in the processing of the α mating pheromone in the two yeasts. Taking into account this similarity, we searched the C. albicans genome for sequences that would encode the α pheromone gene. Here we report the isolation and characterization of the gene MFα1, which codes for the precursor of the α mating pheromone in C. albicans. Two active α-peptides, 13 and 14 amino acids long, would be generated after the precursor molecule is processed in C. albicans. To examine the role of this gene in mating, we constructed an mfα1 null mutant of C. albicans. The mfα1 null mutant fails to mate as MTLα, while MTLa mfα1 cells are still mating competent. Experiments performed with the synthetic α-peptides show that they are capable of inducing growth arrest, as demonstrated by halo tests, and also induce shmooing in MTLa cells of C. albicans. These peptides are also able to complement the mating defect of an MTLα kex2 mutant strain when added exogenously, thereby confirming their roles as α mating pheromones.
Candida albicans is the most common opportunistic fungal pathogen of humans and mostly infects immunosuppressed patients (3). It inhabits diverse niches, which include the gastrointestinal tract and the vagina, and causes infection of skin, mucous membranes, and the bloodstream. Since each of these sites of infection presumably requires differences in gene expression, a great deal of effort has been spent in looking at how such adaptation occurs in Candida. Much of this effort was predicated on the characterization of C. albicans as an asexual obligate diploid.
That this fungus is asexual was disproved after the C. albicans genome sequencing project revealed the presence of sequences homologous to the Saccharomyces cerevisiae MAT (mating-type) loci. The C. albicans homologues of the MAT loci, the MTL (mating-type-like) loci, were found to be heterozygous in common laboratory strains tested (17). Strains homozygous for the MTL locus were generated either by a deletion strategy (18) or by loss, induced by growth on sorbose as the sole carbon source (19), of one homologue of chromosome 5, the site of the MTL loci (29). Strains thus constructed were able to mate both under laboratory conditions (29) and in the animal host (18). These studies set the stage for dissecting the mating pathway in C. albicans.
Mating in fungi has been examined in both yeasts and molds of various sorts. Certain general parts of the process seem to hold across genera and indeed across the kingdom. These include dissimilar regulatory genes (mating-type loci), soluble pheromones, and signal transduction pathways. There are significant homologies in the structures of the proteins involved in the mating process as well. For example, the mat A idiomorph in Neurospora crassa encodes a protein similar to the MATα1 gene product in S. cerevisiae (9, 12). The general structure of the pheromones is also highly conserved, although the amino acid sequence is usually highly divergent, since this determines the species specificity of the pheromone. Thus, one kind of pheromone (mating pheromone a) tends to be farnesylated at the C terminus, while the other (mating pheromone α) is synthesized as a precursor which must be cleaved by specific proteinases. Both have been found in fungi as evolutionarily separated as Schizosaccharomyces pombe and Magnoporthe grisea (7, 37).
Mating in C. albicans is relatively unstudied compared to that in other well-characterized fungi, such as S. cerevisiae, where the mating pathway has been explored in detail. Haploid cells of S. cerevisiae are of two mating types, MATa and MATα. Two opposite mating types fuse upon mixing to form a diploid cell. The first step that allows each mating type to sense the presence of the other is the secretion of mating-type-specific diffusible pheromones. MATa cells secrete pheromone a, and pheromone α is secreted from MATα cells. Binding of these pheromones to the opposite cell type triggers a signal transduction pathway which leads to the expression of several genes required for mating in this budding yeast. Cytoplasmic and nuclear fusion occur when the two mating cell types fuse with each other to form a zygote, which gives rise to diploid daughter cells (38).
Recent work on the characterization of the mating pathway in C. albicans has revealed that the opaque-phase cells of C. albicans are more efficient in mating than the white cell type (33). It has also been demonstrated that the genes required in the mitogen-activated protein kinase signal transduction pathway in mating in S. cerevisiae are also needed for mating in C. albicans, thereby indicating that the mitogen-activated protein kinase pathway is functionally conserved between the two yeasts (6, 28). Furthermore, mating figures or shmoos (cells with elongated, nonseptated projections) have been observed in mixes of MTLa and MTLα cells (27). Several genes homologous to those that are involved in the processing of the mating pheromones in S. cerevisiae are also present in C. albicans, including the serine proteinase gene KEX2 (35). KEX2 is involved in the generation of the mature active α-peptide in S. cerevisiae (22). The C. albicans kex2 mutant fails to mate if the mating type is MTLα, but it mates efficiently if the mating type is MTLa (28). Based on these observations, we searched the C. albicans genome assembly for potential α pheromone-encoding genes. In this report we describe the identification and detailed characterization of the α pheromone precursor gene MFα1 (mating factor α1) of C. albicans.
MATERIALS AND METHODS
Strains and culture conditions.
Table 1 lists the strains used in these experiments. The MFα1 null mutant was derived from strain RM1000. Strain 3861 (MTLα) was used as the parent for constructing the revertants. The revertant and pseudorevertant strains (3898 [MTLα] and 3902 [MTLα]) are derived from strain 3861; in these strains the intact MFα1 gene or a truncated copy has been reinserted by transformation with a plasmid as described below. All of the strains were grown in YEPD (2% Bacto Peptone, 1% yeast extract, and 20% dextrose) or in SD medium (6.7% yeast nitrogen base plus ammonium sulfate, without amino acids, with 2% dextrose, and with 2% Bacto agar for solid media) supplemented with required amino acids and nucleotides at 30°C. Opaque cells were grown in modified Lee's medium (1) at room temperature, and the opaque-phase cell phenotype was confirmed microscopically. Mating was carried out as described previously (29). Revertants were mated against strains 3685 (MTLα) and 3710 (MTLa), which are mycophenolic acid (MPA)-resistant mating tester strains (28), and the mating products were selected on minimal medium containing 10 μg of MPA per ml.
TABLE 1.
Strains used in this work
| Strain | Genotype | MTL | Reference or source |
|---|---|---|---|
| 3752 (RM1000) | his1/his1 ura3/ura3 | a/α | 34 |
| 3842 | MFα1/mfα1::HIS1 | a/α | This work |
| 3845 | mfα1::URA3-dp l200/mfα1::HIS1 | a/α | This work |
| 3852 | mfα1::dpl200/mfα1::HIS1 | a/α | This work |
| 3861 (3852α) | mfα1::dpl200/mfα1::HIS1 | α/α | This work |
| 3863 (3852a) | mfα1::dpl200/mfα1::HIS1 | a/a | This work |
| 3898 (3861) | mfα1/mfα1 pGEMURA3/MFα1 | α/α | This work |
| 3902 (3861) | mfα1/mfα1 pGEMURA3/mfα1Δ | α/α | This work |
| 3896 (CAI4 derivative)a | his3/his3::URA3 | a/a | Elitra Pharmaceuticals, Montreal, Canada |
| 3897 (CAI4 derivative)a | his3/his3::URA3 | α/α | Elitra Pharmaceuticals |
| 3142 | MTLa/mtlα1 mtlα2::hisG ura3/ura3 | a/a | 18 |
| 3716 (3142 derivative)a,b | MTLa1 ura3/ura3 | a/a | This study |
| 3144a | mtla1::URA3/MTLα ade2/ade2 | α/α | 18 |
| 3710 | ura3/ura3 his1/his1 arg5,6/arg5,6 (URA3 IMH3-1)c | a/a | 28 |
| 3685 | ura3/ura3 his1/his1 arg5,6/arg5,6 (URA3 IMH3-1)c | α/α | 28 |
| 3301 | kex2::hisG kex2::hisG ura3/ura3 | α/α | 35 |
| 3108 | kex2::hisG kex2::hisG ura3/ura3 | a/a | 35 |
| 3745(A505 derivative)a,b | trp1/trp1 lys2/lys2 | a/a | 23 |
The opaque cell type of this strain was used for this work. The strain was streaked on YEPD plates and incubated at room temperature to obtain the opaque cells.
The strain was derived from the indicated parent by growth on minimal medium with sorbose as the sole carbon source.
MPA-resistant strain.
The MTL series of primers listed in Table 2 were used for analysis of the MTL alleles in all of the experiments.
TABLE 2.
PCR primers used in this work
| Primera | Nucleotide sequence (5′→3′)b |
|---|---|
| mfαdelF | ATCACCAACAAACTACTAATCACTCTATAACATCAACTAATTAAATCAACAAAAATAACAGTTTTCCCAGTCACGCGTT |
| mfαdelR | TTCATCTAAACAAACATAAAAGTATGATTTCAGTATGCTTTCCATCTTCTTTACTTACTTTGTGGAATTGTGAGCGGATA |
| 5′-detect (1) | TTTTTCATTAAGACCATCATC |
| 3′-detect (2) | ATAATGGAGATAGAAACTTT |
| Ura3detect (4) | AGACCTATAGTGAGAGAGCA |
| ORFΔdetect (3) | AAGGAAAAAAGCGGCCGCGAAGCTAAGTCTAAAGGTGG |
| mfαNotIF | AAGGAAAAAAGCGGCCGCGGTTTTAGTTTATATTCAAATCAGG |
| mfαMluIR | CGACGCGTAAATATTGCATAACAATAACA |
| MTLaF | TTGAAGCGTGAGAGGCAGGAG |
| MTLaR | GTTTGGGTTCCTTCTTTCTCATTC |
| MTLαF | TTCGAGTACATTCTGGTCGCG |
| MTLαR | TGTAAACATCCTCAATTGTACCCGA |
| KEX2F | GTTGAAAGAGGAAAGAAGAGGCGG |
| KEX2R | CGTCATCAGCACGCCTAGTGTCGG |
Primers mfαdelF and mfαdelR were used to amplify both the HIS1 and the URA3 cassettes from the plasmids pGEMHIS1 and pDDB57, respectively. The numbers in parentheses are used in the text to describe the deletion strategy.
Restriction endonuclease recognition sites are in bold. NotI, 5′-GCGGCCGC-3′; MluI, 5′-ACGCGT-3′. Sequence homologous to the vectors are underlined.
Construction of the MFα1 null mutant and generation of MTLa and MTLα derivatives from the null mutant.
MFα1 gene-specific primers (60 bases) mfαdelF and mfαdelR (Table 2) were used to amplify HIS1 and the mini-URA3 blaster cassettes (41) from plasmids pGEMHIS1 and pDDB57, respectively. The amplified fragments were used to transform the strain RM1000 (his1/his1 ura3/ura3) by the lithium acetate transformation method as described previously (11). A 432-bp coding region of MFα1 was deleted by using the HIS1 marker for the first allele and the URA3 marker for the second allele, resulting in strains 3842 (MTLa/α) and 3845 (MTLa/α). Strain 3845 was plated on 5-fluoro-orotic acid (5-FOA) to select for intrachromosomal recombination between the homologous dpl200 sequences of the mini-URA3 blaster cassette, resulting in the loss of the URA3 selectable marker, thereby generating a ura3 strain, 3852 (MTLa/α).
The MFα1 null mutant, 3852 (MTLa/α), was plated on minimal medium with the required supplements and sorbose as the sole carbon source for isolation of strains 3861 (MTLα) and 3863 (MTLa), which are homozygous for either of the mating types. The plates were incubated at 37°C for 1 week, and the Sou+ colonies were analyzed by PCR for the MTL loci as described previously (29).
Reintroduction of MFα1 into its original locus was achieved in strain 3861 (MTLα) by transformation with plasmid pGEMURA3/MFα1. This plasmid consisted of a 2.3-kb DNA fragment containing the 432-bp coding region of MFα1 preceded by 1.5 kb of the promoter region and followed by a 348-bp region downstream of the open reading frame (ORF). The 2.3-kb fragment was PCR amplified with primers mfαNotIF and mfαMluIR, which contain the NotI and MluI restriction enzyme sites (Table 2). The amplified fragment was digested with NotI and MluI and cloned into NotI- and MluI-digested pGEMURA3. To rule out the positional effect of the URA3 gene, a 0.55-kb band containing the truncated ORF of MFα1 was also cloned into pGEMURA3 by using the same enzyme sites. Both of these clones were linearized with HindIII, which is present in the region downstream from the ORF, preceding the MluI restriction enzyme site. Integration of these clones into the deleted MFα1 locus thus occurred at the HindIII site.
Proper integration of the two cassettes during the deletion events and the reintroduction of the wild-type gene into the MFα1 null mutant was verified in each strain and at each step by PCR with primers localized outside the cassettes (primers 1 and 2), a coding-region-specific primer (primer 3), and a URA3-specific primer (primer 4) (Table 2; see Fig. 2A to C). All of the fragments were also sequenced for confirmation of their identity. Total genomic DNA was extracted as described previously (14) and used as the template for the PCRs. The PCR mixtures were typically heated to 94°C for 2 min, followed by 29 cycles of 94°C for 1 min, 55°C for 1 min, and 72°C for 2 min. After a final extension at 72°C for 10 min, reaction mixtures were stored at 4°C. The enzyme Taq DNA polymerase used for the PCRs was from Sigma-Aldrich, St. Louis, Mo.
FIG. 2.
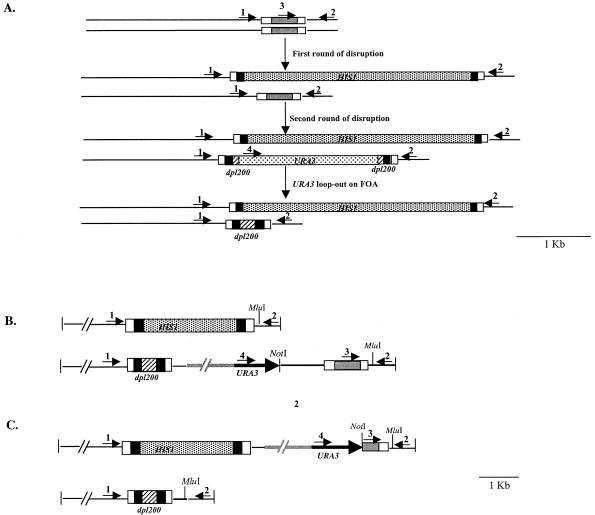
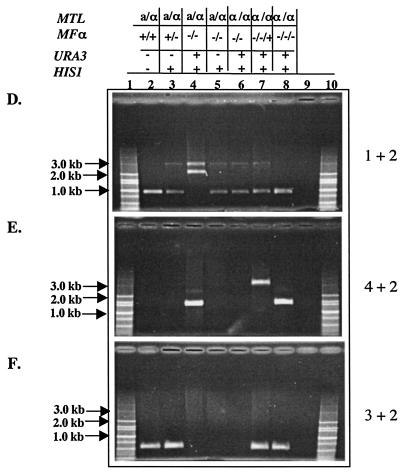
Deletion of the MFα1 gene from strain RM1000 and reintroduction of MFα1 in the null mutant. (A) Schematic representation of the deletion and gene return strategy. The black box is the sequence from the vectors pGEMHIS1 and pDDB57 used as DNA templates for amplifying the HIS1 gene and the mini-URA3 blaster cassette. The grey box represents the 432-bp ORF of the MFα1 gene, which is marked at the end by the upstream and downstream sequences (open boxes) of this gene. Primers used for deleting the MFα1 ORF were homologous to the region represented by these open boxes. The hatched boxes at the end of the URA3 blaster cassette are the dpl200 repeats. Arrowheads 1 to 4 represent primers used in the PCR confirmation of the constructs shown in panels D, E, and F. The 3.1-kb band in panel D represents the mfα1::HIS1 allele. In the first round of disruption, the first allele of the MFα1 ORF (432 bp) was deleted by using HIS1 as the marker, generating a 3.1-kb band and a 0.86-kb wild-type band of MFα1 with primers 1 and 2 (panel D, lane 3). The URA3 miniblaster cassette used for the deletion of the second allele generates bands of 3.1 and 2.3 kb (represents the mfα1::URA3-dpl200 allele) with primers 1 and 2 (panel D, lane 4). The 0.86-kb band observed in panel D, lane 5, is generated as a result of subjecting the mfα1 null mutant to 5-FOA for obtaining the ura3 derivative. The wild-type band and the dpl200 band left after the 5-FOA step are of the same size. (B) Return of the MFα1 gene to the deleted locus. The MFα1 gene was returned to the mutant by integration into the mfα1::dpl200 allele. NotI and MluI mark the ends of the 2.3-kb MFα1 fragment cloned into the vector pGEMURA3. The region in grey marks the vector pGEMURA3 integrated into the deleted MFα1 locus. The integration was confirmed by PCR with primers 1 and 2, generating a 3.1-kb band and a 0.86-kb band from the reintegrated wild-type MFα1 gene (panel D, lane 7). The presence of the ORF in this clone was confirmed by using primers 3 and 2, which generate a 0.55-kb band (panel F, lane 7). Primers 4 and 2 generate a 3.2-kbfragment (panel E, lane 7) for this integration event. (C) Return of a truncated MFα1 gene to the deleted locus. A clone containing the truncated MFα1 ORF was integrated into the mfα1::HIS1 allele. NotI and MluI mark the ends of the 0.55-kb fragment carrying the truncated MFα1 ORF (0.29 kb), cloned into the vector pGEMURA3. Primers 1 and 2 generate the 0.86-kb dpl200 band only (panel D, lane 8). The presence of URA3 (1.6 kb) and the truncated MFα1 gene (0.55 kb) is confirmed by using primers 4 and 2 and primers 3 and 2, respectively (panels E and F, lanes 8) The details of the deletion and replacement strategies are described in Materials and Methods. (D, E, and F) PCR results confirm the integration of the two cassettes in the deletion construct. Each set of reactions shows the results of PCR with different primer pairs (primers 1 and 2, 4 and 2, and 3 and 2, respectively) carried out on genomic DNA from the wild type (3752) (lanes 2), 3842 (lanes 3), 3845 (lanes 4), 3852 (lanes 5), 3861 (lanes 6), 3898 (lanes 7), and 3902 (lanes 8). Lanes 9, no-DNA controls for the PCR. Lanes 1 and 10, DNA molecular size markers.
Synthesis and biological activity of the α-peptide.
The 13-amino-acid (SP1, GFRLTNFGYFEPG) and 14-amino-acid (SP2, GFRLTNFGYFEPGK) active peptides were synthesized at the microchemical facility, University of Minnesota, Minneapolis, by solid-phase peptide synthesis with 9-fluorenylmethoxy carbonyl chemistry (Perkin-Elmer/ABI Pioneer peptide synthesizer). When experiments were performed with both peptides SP1 and SP2, the concentrations were always 10 μg of SP1 per ml and 5 μg of SP2 per ml to reflect the genomic ratio of 2SP1 to 1SP2. The shmooing experiments with the peptides were performed with the opaque cell type of a given strain. The peptides were dissolved in 2% dimethyl sulfoxide (DMSO) at a stock concentration of 1 mg/ml. Opaque cells from strains 3716 (MTLa) and 3144 (MTLα) were inoculated into modified Lee's medium and grown for 24 h at room temperature. Approximately 3 × 106 cells from each of the two strains were incubated at room temperature with both of the peptides in a final volume of 1.0 ml in a plastic 15-ml Falcon tube. The same experiment was also carried out in the presence of either SP1 (10 μg/ml) or SP2 (5 μg/ml) separately. The same volume of 2% DMSO was added to control samples. Aliquots of 200 μl were taken every 2 h, fixed with 3.7% formaldehyde for 30 min, and stained with Calcofluor at a final concentration of 0.1 μg/μl for another 30 min in the dark. The cells were washed twice with water before final resuspension in 200 μl of water. Three microliters of cells was examined on an E800 Nikon microscope. Images were recorded with a Nikon Cool camera, and Adobe Photoshop software was used for processing the images.
Experiments with the peptides were also carried out on plates. The two peptides were spotted together or separately (10 μg of SP1 per ml or 5 μg of SP2 per ml) onto the patched strains 3716 (MTLa) and 3144 (MTLα). The plates were incubated at room temperature and photographed after 3 days. Cells from the patches were scooped, and phase-contrast images were taken on an E800 Nikon microscope and recorded with a Nikon Cool camera.
Halo tests with the peptides.
The halo tests were performed with either the SP1 or SP2 peptide at a concentration of 5 μg/ml on the opaque cell type of strain 3745 (MTLa). Cells from a fresh single opaque colony were resuspended in 200 μl of water and spread onto an SD medium plate supplemented with the required amino acids. Five microliters of peptides SP1 and SP2 from a stock solution of 1 mg/ml was spotted onto the plate. The plates were monitored for the appearance of halos after 24 and 48 h.
Complementation assays.
The complementation assays were performed with the MTLα kex2 (3301) and MTLα mfα1 (3861) strains. The MTLa kex2 (3108) and MTLa mfα1 (3863) strains were used as controls for this experiment. Matings were performed as described earlier (29) except that the plates contained the purified peptides SP1 and SP2 added separately or together at an SP1/SP2 ratio of 2:1 to give final concentrations of 6 × 10−5, 6 × 10−6, 6 × 10−7, and 6 × 10−8 M. Since the mating experiments performed in our laboratory are nonquantitative assays (28), the number of mating products that appear on plates with SP1 and SP1 plus SP2 for the MTLα kex2 mutant strain varies on the two plates (see Fig. 7A).
FIG. 7.
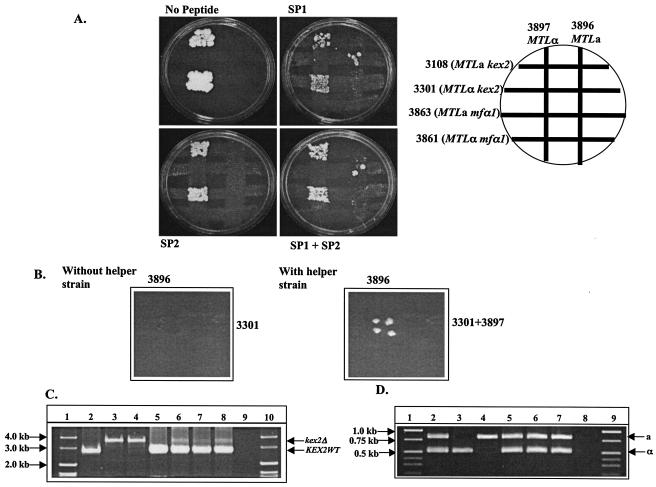
Mating of the kex2 and mfα1 null mutants in the presence of exogenously added α-peptides. (A) Synthetic peptide can complement the mating defect in an MTLα kex2 strain but not in an MTLα mfα1 strain. A diagram of the mating plate is shown on the upper right. The strains used were 3108 (MTLa kex2), 3301 (MTLα kex2), 3863 (MTLa mfα1), 3861(MTLα mfα1), 3896 (MTLa his3−), and 3897 (MTLα his3−). SP1 and SP2 concentrations of 6 × 10−5 M of were used in the plates. (B) An MTLα helper strain can complement the mating defect in an MTLα kex2 strain but not in an MTLα mfα1 strain. The strains used were 3896 (MTLa his3−), 3301(MTLα kex2), and the α helper strain 3897 (MTLα his3−). (C) PCR analysis of the KEX2 locus in parents and mating products. The position of the wild-type band (KEX2WT) and the deletion band (kex2Δ) of KEX2 are marked by arrows. Lane 2, SC5314; lane 3, 3301; lane 4, 3108; lane 5, 3896; lane 6, mating product picked from right plate in panel B; lanes 7 and 8, mating products from plates with SP1 and SP1 plus SP2, respectively (A); lane 9, no-DNA control for the PCR; lanes 1 and 10, the DNA molecular size markers. (D) PCR analysis of the MTL alleles in parents and mating products. The positions of the MTLa- and MTLα-specific bands are marked by arrows. Lane 2, SC5314; lane 3, 3301; lane 4, 3108; lane 5, mating product picked from right plate in panel B; lanes 6 and 7, mating products from plates with SP1 and SP1 plus SP2, respectively (A); lane 8, no-DNA control for the PCR; lanes 1 and 9, DNA molecular size markers.
Matings in the presence of an MTLα helper strain (3897) as a source for the α pheromone were performed by mixing strain 3301 (MTLα kex2) with strain 3897 (MTLα) in one set and 3861 (MTLα mfα1) with 3897 (MTLα) in the other set and mating each pair with strain 3896 (MTLa). Strains 3896 (MTLa) and 3897 (MTLα) have the same auxotrophic marker and were chosen for this experiment in order to avoid selecting for the tetraploid products that would arise from mating between these two strains. For the mating experiment, one strip contained cells from strain 3301 (MTLα kex2) or 3861 (MTLα mfα1) costreaked with cells from strain 3897 (MTLα) and the second strip had cells from strain 3896 (MTLa) on a YEPD plate at 30°C. The rest of the mating experiment was carried as described previously (29). The mating products arising from this experiment were selected on minimal medium (see Fig. 7B). The mating products obtained from the 3301 (MTLα kex2) × 3896 (MTLa) cross were subjected to PCR analysis of the MTL alleles and also to PCR analysis for the KEX2 gene (for sequences of the primers, see Table 2).
RESULTS
Identification and sequence analysis of the gene encoding the α mating pheromone (MFα1).
In a number of fungi, generation of one of the mating pheromones involves cleavage of the precursor polypeptide specifically at the C-terminal side of a Lys-Arg sequence by a serine proteinase; in S. cerevisiae, this proteinase is encoded by KEX2. In order to screen for the C. albicans MFα1-encoding gene, a genome-wide search for ORFs with KEX2 proteinase cleavage sites flanking small repeated sequences was carried out. This search led to the identification of a putative gene sequence in C. albicans with structural similarities to the α mating pheromone of S. cerevisiae (Orf 6-4306 at http://www-sequence.Stanford.edu/group/candida/) (Fig. 1). This gene was designated MFα1 for mating factor α. Another group independently reported this gene sequence as a pheromone candidate but carried out no further analysis (36).
FIG. 1.

Nucleotide sequence of the MFα 1 gene and deduced amino acid sequence of its product. The sequence of a 1.5-kb C. albicans genomic fragment carrying the MFα1 gene is shown. One-letter codes for amino acids are shown beneath the nucleotide sequence. Regions encoding the mature α mating pheromones are in boldface and underlined. Possible cleavage signals (Lys-Arg) recognized by the KEX2 proteinase are highlighted in grey. A stretch of 19 amino acids which could serve as the possible signal sequence for translocation of the precursor polypeptide across the membrane of the endoplasmic reticulum is in boldface and italic. A putative TATA box in the region upstream of the ORF is in boldface. Putative PREs are in boldface and italic.
Analysis of the gene sequence revealed a 432-bp ORF that codes for three repeats separated by KEX2 proteinase cleavage sites, unlike the S. cerevisiae MFα1 gene, which contains four repeats (26). The three putative α pheromone peptides are separated by spacer sequences, each containing a pair of Lys-Arg residues, the KEX2 proteinase cleavage sites. Two of the three α-peptides (SP1) are identical in sequence; the third contains an extra Lys residue at the C terminus (SP2) (Fig. 1). The N termini of two of the peptides have the -x-A spacer dipeptides, which in S. cerevisiae are known to be removed by the action of dipeptidyl peptidase encoded by the STE13 gene. The presence of a homologue of STE13 in the Candida genome suggests a similar role for this gene. The first repeat has a different N-terminal sequence, KG.
Analysis of the sequence upstream of the ATG start codon revealed the presence of a putative TATA box and sequences that are important for regulating the expression of this gene in S. cerevisiae. An example is the presence of pheromone response elements (PREs) in the upstream region of MFα1 (10, 20). The hydrophobicity profile of the 143-amino-acid precursor peptide revealed the presence of a unique hydrophobic region (19 amino acid residues) at the N terminus; this region may serve as a signal sequence during translocation of the precursor across the membrane of the endoplasmic reticulum (Fig. 1). Thus, the overall structural similarity of the MFα1 gene of C. albicans to the S. cerevisiae MFα1 gene indicates that the mechanism for maturation of the C. albicans MFα1 gene product may be similar to that of the S. cerevisiae pheromone (26, 38).
The MFα1 null mutant fails to mate as MTLα.
In order to ascertain the role of MFα1 in mating in C. albicans, both copies of this gene were deleted in the strain RM1000 (3752). The MFα1 null mutant was generated by a complete deletion of the 432-bp ORF as described in Materials and Methods (Fig. 2A, D, E, and F). The null mutant (3852) was then grown on sorbose-containing medium to obtain MTLa (3863) and MTLα (3861) strains and tested for mating (Table 1). The MTLα mfα1−/− cells were unable to mate, but the MTLa mfα1−/− cells were as fertile as the parent (Fig. 3A). Reintroduction of the MFα1 gene in the null mutant restored mating in the MTLα cells (3898) (Fig. 2B to F and 3A), indicating that the MFα1 gene is cell type specific. Reintroduction of a construct containing the truncated mfα1 gene in the null mutant (3902) did not restore mating, indicating that restoration of the mating phenotype in the revertant strain (3898) was solely due to the MFα1 gene and was not due to problems with the expression of the URA3 gene (Fig. 2C to F and 3A). PCR analysis of the mating products confirmed the mating phenotype by demonstrating the presence of both the MTLa and MTLα alleles (Fig. 3B).
FIG. 3.

Mating of the mfα1 null strains and the revertants. (A) Strains were grown and cross streaked, and mating products were selected on minimal MPA plates as described in Materials and Methods. The strains used were 3861 (MTLα mfα1−/−), 3863 (MTLa mfα1−/−), 3898 (MTLα mfα1 pGEMURA3/MFα1), 3902 (MTLα mfα1 pGEMURA3/mfα1Δ), 3685 (MTLα MPAR his− arg−), and 3710 (MTLa MPAR his− arg−). MTLα mfα1R and MTLα mfα1RΔ are the same as strains 3898 and 3902, respectively. (B) PCR analysis of the MTL alleles in the parents and the mating products. Mating products were picked from the mating plates and restreaked on a minimal MPA (10 μg/ml) plate. Genomic DNAs from strains 3861 (lane 2), 3863 (lane 3), 3898 (lane 4), 3902 (lane 5), 3710 (lane 6), 3685 (lane 7), the 3863 × 3685 mating product (lanes 8 and 9), the 3898 × 3710 mating product (lanes 10 and 11), and SC5314 (lane 12) were used as templates for performing PCR with the MTLa- and MTLα-specific primers. Lane 13, no-DNA control for the PCR. Lanes 1 and 14, DNA molecular size markers. The positions of the MTLa-specific and MTLα-specific bands are marked by the arrows on the right.
Biological activity of the two α pheromone peptides and their effect on MTLa and MTLα cell types.
In S. cerevisiae the α-factor is known to induce morphological changes in MATa cells that are characteristic of the mating process in this budding yeast. The appearance of morphologically abnormal cells allows a sensitive method for the assay of α-factor biological activity (reviewed in reference 8). Similar morphological changes have been observed in mixtures of C. albicans MTLa and MTLα cells (27).
When the α-peptides SP1 and SP2 were spotted onto patches of MTLa (3716) and MTLα (3144) cells as described in Materials and Methods, only the MTLa patch became wrinkled in response to the SP1 peptide after 3 days of incubation at 25°C, while there was no change observed in either the MTLα cells (Fig. 4A and B) or the MTLa white cells (data not shown) of the same strain. Since Miller and Johnson showed that opaque-phase cells are several orders of magnitude more efficient at mating (33), it is not surprising that they are more sensitive to the pheromone than are the cells in the white phase. When cells were sampled from the wrinkled streak, a high number of cells with evaginations or shmooing were observed (Fig. 4E and G). Every single cell seemed to filament in response to SP1. In contrast to SP1, peptide SP2 did not induce wrinkling in the MTLa cells, but cells that were scooped from the patch did show some shmooing (Fig. 4B and F). Fewer cells shmoo in response to SP2 than do so in response to SP1. The MTLα opaque-phase cells from strain 3144 did not respond to either of the two peptides (Fig. 4A to C). Peptide SP2 (GFRLTNFGYFEPGK) differs from SP1 (GFRLTNFGYFEPG) in having an extra lysine residue at the C terminus.
FIG. 4.

Colony phenotypes induced by α-peptides. Strains were patched, and peptides SP1 and SP2 were spotted onto them as described in Materials and Methods. (A) MTLa (3716) and MTLα (3144) opaque-phase cells spotted with 2% DMSO (upper panel) and with 10 μg of SP1 per ml plus 5 μg of SP2 per ml (lower panel). (B) Cells spotted with 10 μg of SP1 per ml (upper panel) and 10 μg of SP2 per ml (lower panel). (C to G) Phase-contrast images of cells scooped from plates A and B. Bar, 5 μm.
In order to assay directly for the activity of the MFα1-encoded peptide, opaque-phase cells from homozygous MTLa (3716) and MTLα (3144) strains of C. albicans were incubated with the α-peptides (SP1 plus SP2) in modified Lee's medium at 25°C and monitored for changes in the morphology at different time points as described in Materials and Methods. The MTLa cells started to evaginate 2 h after incubation with the peptides and formed tube-like structures (called shmooing) with time, similar to the mating figures observed in previous studies (27), while the MTLa cells without the peptides and MTLα cells with peptides did not shmoo even after 24 h of incubation (Fig. 5). Calcofluor, which stains chitin and thus the septa of yeast cells, was used to examine these cells. The shmooing figures formed in response to the peptide were continuous, with no visible septation even after 24 h of incubation, thereby authenticating the absence of hyphae and presence of true shmoos (Fig. 5, right panels). To demonstrate that this effect of the α-peptide is not strain specific, different strains homozygous at the MTLa locus were tested for shmooing (data not shown). In all of the cases shmooing was limited to the MTLa cells, while the MTLα cells did not shmoo at all.
FIG. 5.

Shmooing of opaque-phase cells in response to the α-peptides occurs exclusively in the MTLa cell type. The response of the cells in modified Lee's medium is shown. The left and right columns in each panel show phase-contrast and fluorescent images of Calcofluor-stained cells, respectively. (A) 3716 (MTLa) opaque-phase cells without peptide; (B) 3716 (MTLa) opaque-phase cells with peptides SP1 and SP2 (10 and 5 μg/ml, respectively); (C) 3144 (MTLα) opaque-phase cells with peptides SP1 and SP2 (10 and 5 μg/ml, respectively). The arrows point to the septum (A and C), while the arrowhead points to the region where a septum should have been present (B). Bar, 10 μm.
SP1 and SP2 generate halos when placed on MTLa opaque-phase cells.
The a and α pheromones in S. cerevisiae are known to act on the opposite mating types by inducing a reversible arrest in the G1 phase of the cell cycle. This arrest, which occurs prior to shmooing, is important for synchronization of the cell cycles in the two mating types (2, 32, 38). One of the qualitative assays for the mating pheromones is based on their ability to inhibit the growth of cells of the opposite mating type (in this case MTLa cells in response to the α-factor) in the surrounding area, resulting in a clear zone or halo. In order to examine the role of the MFα1-encoded peptides in growth inhibition, we carried out halo tests with peptides SP1 and SP2. We found that peptide SP1 is more capable of inhibiting the growth of the MTLa opaque-phase cells, while SP2 has a lesser but observable effect on these cells at the concentrations used in this experiment (Fig. 6). The halos produced in response to SP1 or SP2 are not as clear as the halos observed with the S. cerevisiae α pheromone (30). This might be due either to some of the cells switching back to the white cell type or to the presence of a barrier activity similar to the endopeptidase observed in S. cerevisiae (13). As discussed below, C. albicans has candidates for such an activity.
FIG. 6.
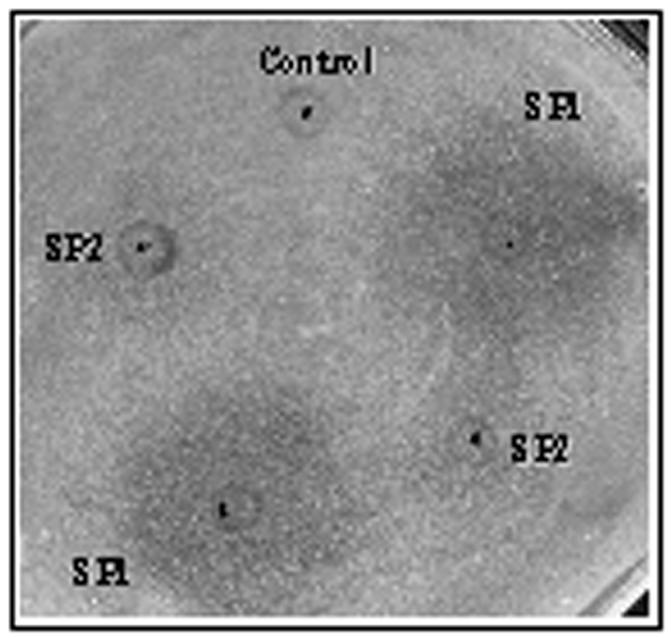
MTLa opaque-phase cells generate a halo in response to the α-peptides. The dots mark the areas where the peptides were spotted. Methanol (50%) was spotted as the control on a lawn of MTLa opaque-phase cells of strain 3745. Five microliters from a 1-mg/ml stock solution of the peptides SP1 and SP2 was spotted onto this lawn of cells in duplicates. The ring seen around the spotting point is an artifact generated due to the solvent. Halos represent the zone of growth inhibition of MTLa cells in response to peptides SP1 and SP2, as observed after 48 h.
Exogenously added synthetic α-peptide renders the MTLα kex2 mutant mating efficient.
We know from our previous work that the kex2 mutant does not mate as MTLα, while the MTLa kex2 mutant is as fertile as the parent strain (28). If the sole reason for the inability of the kex2 mutant to mate as MTLα is due to the absence of mature α pheromone, then the addition of the MFα1-encoded peptides, viz., SP1 and SP2, should be able to complement this mating defect. To test this possibility, the MTLα kex2 mutant was mated against an MTLa strain on plates supplemented with various concentrations (as mentioned in Materials and Methods) of peptides SP1 and SP2 added separately and in combination. The MTLα kex2 strain was able to mate in the presence of SP1 and SP1 plus SP2 at final concentrations of 6 × 10−5 and 6 × 10−6 M (Fig. 7A), but it failed to do so at concentrations of 6 × 10−7 and 6 × 10−8 M. This mutant strain was not able to mate in presence of SP2 alone even at the highest concentration (6 × 10−5 M), thereby suggesting that although SP2 alone can induce shmooing and a small halo in a cells, it is not potent enough to render kex2 cells mating competent at the concentrations used in this study (Fig. 7A, 5F, and 6). In order to achieve a physiological level of the α pheromone in the mating reaction, the MTLα kex2 mutant was mixed with strain 3896 (MTLa) in the presence and absence of strain 3897 (MTLα). This strain, which is his3− and cannot complement 3896, was added to serve as a source of α-factor in this mixture (Table 1). The mating defect of the MTLα kex2 mutant was also complemented in this experiment (Fig. 7B).
If the colonies arising from these experiments are true mating products, they should harbor a wild-type copy of the KEX2 gene along with the deleted copy, in addition to containing both of the MTL alleles. Therefore, two sets of primers were used for the PCR analysis of these mating products. PCR analysis with the KEX2-specific primers indicated the presence of both the wild-type and the deletion copies of this gene (Fig. 7C, lanes 6 to 8). Similarly PCR with the MTL-specific primers demonstrated the presence of both of the MTL alleles (Fig. 7D, lanes 5 to 7). These experiments confirmed the authenticity of these mating products and demonstrate that the MFα1-encoded peptide SP1 indeed is capable of supplementing the α pheromone defect in the MTLα kex2 mutant strains, thus confirming the role of this gene in mating in C. albicans. Experiments similar to those with the kex2 deletion strain were performed with the MTLα mfα1 mutant strain, but no complementation was observed (Fig. 7A).
DISCUSSION
Pheromones are ubiquitous in fungal mating systems, and even the general structure is conserved over the fungal kingdom, from S. pombe (7) and N. crassa (9, 12) to basidiomycetes such as Cryptococcus neoformans (5). The discovery of mating in C. albicans led to the search for the pheromones involved. The search for the α pheromone was guided by the similarity of pheromone structure across the fungal kingdom and by the observation that the KEX2 gene is essential for mating (28). Based on the KEX2 site specificity, we identified a sequence which encodes a putative protein of 143 amino acids containing a hydrophobic leader sequence, three identical repeats preceded by the sequence KR (the site of cleavage by the KEX2 proteinase), and sequences similar to the PREs of S. cerevisiae in the 5′ untranslated region. The pheromone-stimulated transcriptional induction of genes is conferred by the presence of PREs, which are cis-acting DNA sequences present in the upstream regions of genes (24, 40). The presence of these sequences suggests that the regulation of the α pheromone might be similar in the two yeasts. Further confirmation of the importance of these sequences will come from a detailed mutational analysis of the upstream region. Although there are three identical repeats in the gene, processing by the KEX2 proteinase would leave one of the three with a C-terminal lysine. We synthesized both the lysine-containing (SP2) and the lysine-free (SP1) peptides to determine their biological effects. Purification of the α-factor and subsequent determination of its peptide sequence will lead to the identification of the exact cleavage sites in this protein and should also determine whether the peptide containing the extra Lys residue is secreted. A homologue of KEX1, whose product cleaves C-terminal lysine and arginine, and a homologue of STE13, encoding the dipeptidyl aminopeptidase in S. cerevisiae which removes the amino acids at the N terminus, exist in the Candida genome database (39). However, since the first peptide repeat of MFα1 is preceded by a KG sequence, not the DA and EA of the other two repeats, the enzyme from C. albicans must have some differences in specificity compared to Ste13p from S. cerevisiae, since the S. cerevisiae Ste13p is known to cleave at the x-A dipeptides (21). Thus, although the mechanisms of precursor processing in the two yeasts are similar on the surface, some of the details must be different.
The biological significance of the MFα1 sequence was demonstrated by the fact that deletion of both alleles of this gene led to sterility in MTLα but not MTLa strains, and return of the wild-type MFα1 gene in the mfα1 null mutant restored its mating ability. The synthetic peptides (SP1 and SP2) induced morphological changes (shmooing) in MTLa but not MTLα strains. Furthermore, these peptides could also complement the mating defect of the kex2 null mutant in the mating reaction. All of these findings suggest that the gene we have isolated is the biologically active MFα1 gene and that, unlike Saccharomyces but similar to Neurospora (11) and S. pombe (7), C. albicans seems to carry only one copy of this pheromone gene. SP1, the 13-amino-acid (lysine-free) peptide seems to be more active than the 14-amino-acid SP2. In most experiments in which they were tried together and singly, the effect of the two together was not significantly different from that of SP1 alone, and the effect of SP2 was usually much less. SP2 alone also did not complement the mating defect of the kex2 mutant.
We examined the activity of the MFα1-encoded peptides by taking advantage of the ability of these peptides to diffuse through agar and cause halos in a lawn of MTLa opaque-phase cells. The growth inhibition is probably due to cell cycle arrest, but we have not shown this microscopically. Despite the fact that no homologue of the BAR1 (SST1) gene has been found in the Candida database, the halos were diffuse. Bar1p is an aspartyl proteinase, and C. albicans has a large gene family encoding secreted aspartyl proteinases (the SAP gene family) with widely differing patterns of expression. Some of these previously identified proteinases may function to degrade the pheromone; for example, the SAP1 and SAP3 genes have been shown to be differentially expressed in the opaque (mating) phase (15). Since mutations in each of these genes are available, their role in mating could be easily tested (16).
The ability of the exogenously added α-peptide (SP1) to alleviate the mating defect of the MTLα kex2 mutant is consistent with the results obtained with the MATα kex2 and the MATα ste13 mutant strains of S. cerevisiae, wherein the addition of exogenous α-factor has been shown to partially complement the mating defects of these two strains (4). It has been shown for S. cerevisiae that the MATα kex2 mutant secretes very small amounts of the MFα peptides, which are undetectable by bioassay and just barely detectable by a sensitive radioimmunoassay (21). This might be one reason why the MFα-encoded peptides were able to restore the mating defect in the MTLα kex2 mutant strains of both C. albicans and S. cerevisiae.
On the other hand, neither the addition of exogenous α-factor at various concentrations nor the presence of a helper MTLα strain could complement the mating defect of the mfα1 null mutant. This result was also obtained for MATα mfα1 mfα2 double mutants of S. cerevisiae (25). If the precursor peptides have a role in the mating reaction in MTLα cells, the failure of the added peptide to complement the MFα1 deletion would be explained, since the MTLα mfα1 strains of C. albicans and the MATα mfα1 mfα2 strains of S. cerevisiae totally lack the endogenous precursor peptide. One possible role of the precursor peptide could be to trigger a mating signal, distinct from the signals that lead to growth arrest or shmooing, in MTLa cells. A second possibility that could account for the failure of the α-peptides to complement the mating defect of the MTLα mfα1 null mutant would be the concentration of the peptides used for this experiment. In S. cerevisiae, high levels of exogenous α pheromone can complement the mating defect of the MATα mfα1 mfα2 strain in crosses to MATa strains that contain mutations that result in supersensitivity to α-factor (sst1-2 and sst2-1) (25). The supersensitivity of these mutants is due to a defect in the degradation of α-factor (30). If C. albicans has a highly active degradation process, the pheromones might not have reached the necessary concentration in our experiments.
In keeping with previous observations that the mating reaction in C. albicans resembles that in S. cerevisiae in a great many ways, the α pheromone involved is also similar, in that it is a small peptide, it requires processing by the product of the KEX2 gene, and it causes morphological changes in cells of the MTLa mating type. The amino acid sequence of the C. albicans α pheromone, however, is unrelated to that of S. cerevisiae, forming a sterility barrier between these two yeasts. The lack of any other MFα1 genes in the C. albicans genome assembly distinguishes this yeast's pheromone gene organization from that of S. cerevisiae, which has two MFα1 genes. Although an a pheromone gene has not yet been reported, it seems very likely that one must exist, and we are presently engaged in an attempt to identify one. This report further supports both the general properties of pheromones in fungal mating and the close homology in the mating processes of two distantly related yeasts, C. albicans and S. cerevisiae. It is somewhat surprising that the process has been so closely conserved over close to two hundred million years (31); this finding would suggest that mating in Candida has an important function, despite the reported absence of meiosis. One possible function of mating, of course, is generation of the diversity required for its unique life style, including both commensality and pathogenesis.
ADDENDUM IN PROOF
After this article was accepted for publication, two reports describing experiments on MFα1 and its products in C. albicans were published: S. R. Lockhart et al., Eukaryot. Cell 2:847-855, 2003; and R. J. Bennett, Mol. Cell. Biol. 23:8189-8201, 2003.
Acknowledgments
We thank Dana Davis and B. B Magee for their comments on the manuscript. We appreciate the assistance of Mark Sanders, Kelly Josephson, and Taryn O'Hall in generating the images on the microscope at the imaging center, University of Minnesota, St. Paul. We deeply appreciate Maryam-Gerami Nejad for helpful suggestions during the preparation of the manuscript.
This work was supported by grant AI16567 awarded to P.T.M.
Footnotes
NRC publication number 46156.
REFERENCES
- 1.Bedell, G. W., and D. R. Soll. 1979. Effects of low concentrations of zinc on the growth and dimorphism of Candida albicans: evidence for zinc-resistant and -sensitive pathways for mycelium formation. Infect. Immun. 26:348-354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bucking-Throm, E., W. Duntze, L. H. Hartwell, and T. R. Manney. 1973. Reversible arrest of haploid yeast cells in the initiation of DNA synthesis by a diffusible sex factor. Exp. Cell Res. 76:99-110. [DOI] [PubMed] [Google Scholar]
- 3.Calderone, R. 2002. Candida and candidiasis. American Society for Microbiology, Washington, D.C.
- 4.Chan, R. K., L. M. Melnick, L. C. Blair, and J. Thorner. 1983. Extracellular suppression allows mating by pheromone-deficient sterile mutants of Saccharomyces cerevisiae. J. Bacteriol. 155:903-906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Chaturvedi, V., J. Fan, B. Stein, M. J. Behr, W. A. Samsonoff, B. L. Wickes, and S. Chaturvedi. 2002. Molecular genetic analyses of mating pheromones reveal intervariety mating or hybridization in Cryptococcus neoformans. Infect. Immun. 70:5225-5235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Chen, J., J. Chen, S. Lane, and H. Liu. 2002. A conserved mitogen-activated protein kinase pathway is required for mating in Candida albicans. Mol. Microbiol. 46:1335-1344. [DOI] [PubMed] [Google Scholar]
- 7.Davey, J. 1998. Fusion of a fission yeast. Yeast 14:1529-1566. [DOI] [PubMed] [Google Scholar]
- 8.Elion, E. A. 2000. Pheromone response, mating and cell biology. Curr. Opin. Microbiol. 3:573-581. [DOI] [PubMed] [Google Scholar]
- 9.Ferreira, A. V., Z. An, R. L. Metzenberg, and N. L. Glass. 1998. Characterization of mat A-2, mat A-3 and deltamatA mating-type mutants of Neurospora crassa. Genetics 148:1069-1079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Flessel, M. C., A. J. Brake, and J. Thorner. 1989. The MFα1 gene of Saccharomyces cerevisiae: genetic mapping and mutational analysis of promoter elements. Genetics 121:223-236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Gietz, R. D., R. H. Schiestl, A. R. Willems, and R. A. Woods. 1995. Studies on the transformation of intact yeast cells by the LiAc/SS-DNA/PEG procedure. Yeast 11:355-360. [DOI] [PubMed] [Google Scholar]
- 12.Glass, N. L., J. Groteleuschen, and R. L. Metzenberg. 1990. Neurospora crassa A mating type region. Proc. Natl. Acad. Sci. USA 87:4912-4916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hicks, J. B., and I. Herskowitz. 1976. Evidence for a new diffusible element of mating pheromones in yeast. Nature 260:246-248. [DOI] [PubMed] [Google Scholar]
- 14.Hoffman, C. S., and F. Winston. 1987. A ten minute DNA preparation from yeast efficiently releases autonomous plasmids for transformation of Escherichia coli. Gene 57:267-272. [DOI] [PubMed] [Google Scholar]
- 15.Hube, B., M. Monod, D. A. Schofield, A. J. Brown, and N. A. Gow. 1994. Expression of seven members of the gene family encoding secretory aspartyl proteinases in Candida albicans. Mol. Microbiol. 14:87-99. [DOI] [PubMed] [Google Scholar]
- 16.Hube, B., D. Sanglard, F. C. Odds, D. Hess, M. Monod, W. Schafer, A. J. Brown, and N. A. Gow. 1997. Disruption of each of the secreted aspartyl proteinase genes SAP1, SAP2, and SAP3 of Candida albicans attenuates virulence. Infect. Immun. 65:3529-3538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hull, C. M., and A. D. Johnson. 1999. Identification of a mating type-like locus in the asexual pathogenic yeast Candida albicans. Science 285:1271-1275. [DOI] [PubMed] [Google Scholar]
- 18.Hull, C. M., R. M. Raisner, and A. D. Johnson. 2000. Evidence for mating of the “asexual” yeast Candida albicans in a mammalian host. Science 289:307-310. [DOI] [PubMed] [Google Scholar]
- 19.Janbon, G., F. Sherman, and E. Rustchenko. 1998. Monosomy of a specific chromosome determines l-sorbose utilization: a novel regulatory mechanism in Candida albicans. Proc. Natl. Acad. Sci. USA 95:5150-5155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Jarvis, E. E., D. C. Hagen, and G. F. Sprague, Jr. 1988. Identification of a DNA segment that is necessary and sufficient for α-specific gene control in Saccharomyces cerevisiae: implications for regulation of α-specific and a-specific genes. Mol. Cell. Biol. 8:309-320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Julius, D., L. Blair, A. Brake, G. Sprague, and J. Thorner. 1983. Yeast α-factor is processed from a larger precursor polypeptide: the essential role of a membrane-bound dipeptidyl aminopeptidase. Cell 32:839-852. [DOI] [PubMed] [Google Scholar]
- 22.Julius, D., A. Brake, L. Blair, R. Kunisawa, and J. Thorner. 1984. Isolation of the putative structural gene for the lysine-arginine-cleaving endopeptidase required for processing of yeast prepro-alpha-factor. Cell 37:1075-1089. [DOI] [PubMed] [Google Scholar]
- 23.Kakar, S. N., R. M. Partridge, and P. T. Magee. 1983. A genetic analysis of Candida albicans: isolation of a wide variety of auxotrophs and demonstration of linkage and complementation. Genetics 104:241-255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kronstad, J. W., J. A. Holly, and V. L. MacKay. 1987. A yeast operator overlaps an upstream activation site. Cell 50:369-377. [DOI] [PubMed] [Google Scholar]
- 25.Kurjan, J. 1985. α-factor structural gene mutations in Saccharomyces cerevisiae: effects on α-factor production and mating. Mol. Cell. Biol. 5:787-796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kurjan, J., and I. Herskowitz. 1982. Structure of a yeast pheromone gene (MFα): a putative α-factor precursor contains four tandem copies of mature α-factor. Cell 30:933-943. [DOI] [PubMed] [Google Scholar]
- 27.Lockhart, S. R., K. J. Daniels, R. Zhao, D. Wessels, and D. R. Soll. 2003. Cell biology of mating in Candida albicans. Eukaryot. Cell 2:49-61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Magee, B. B., M. Legrand, A. M. Alarco, M. Raymond, and P. T. Magee. 2002. Many of the genes required for mating in Saccharomyces cerevisiae are also required for mating in Candida albicans. Mol. Microbiol. 46:1345-1351. [DOI] [PubMed] [Google Scholar]
- 29.Magee, B. B., and P. T. Magee. 2000. Induction of mating in Candida albicans by construction of MTLa and MTLα strains. Science 289:310-313. [DOI] [PubMed] [Google Scholar]
- 30.Manney, T. R. 1983. Expression of the BAR1 gene in Saccharomyces cerevisiae: induction by the α mating pheromone of an activity associated with a secreted protein. J. Bacteriol. 155:291-301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Massey, S. E., G. Moura, P. Beltrao, R. Almeida, J. R. Garey, M. F. Tuite, and M. A. Santos. 2003. Comparative evolutionary genomics unveils the molecular mechanism of reassignment of the CTG codon in Candida spp. Genome Res. 13:544-557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Michaelis, S., and I. Herskowitz. 1988. The a-factor pheromone of Saccharomyces cerevisiae is essential for mating. Mol. Cell. Biol. 8:1309-1318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Miller, M. G., and A. D. Johnson. 2002. White-opaque switching in Candida albicans is controlled by mating-type locus homeodomain proteins and allows efficient mating. Cell 110:293-302. [DOI] [PubMed] [Google Scholar]
- 34.Negredo, A., L. Monteoliva, C. Gil, J. Pla, and C. Nombela. 1997. Cloning, analysis and one-step disruption of the ARG5,6 gene of Candida albicans. Microbiology 143:297-302. [DOI] [PubMed] [Google Scholar]
- 35.Newport, G., and N. Agabian. 1997. KEX2 influences Candida albicans proteinase secretion and hyphal formation. J. Biol. Chem. 272:28954-28961. [DOI] [PubMed] [Google Scholar]
- 36.Newport, G., A. Kuo, A. Flattery, C. Gill, J. J. Blake, M. B. Kurtz, G. K. Abruzzo, and N. Agabian. 2003. Inactivation of Kex2p diminishes the virulence of Candida albicans. J. Biol. Chem. 278:1713-1720. [DOI] [PubMed] [Google Scholar]
- 37.Shen, W. C., P. Bobrowicz, and D. J. Ebbole. 1999. Isolation of pheromone precursor genes of Magnaporthe grisea. Fungal Genet. Biol. 27:253-263. [DOI] [PubMed] [Google Scholar]
- 38.Sprague, G., and J. Thorner. 1992. Pheromone response and signal transduction during the mating process of Saccharomyces cerevisiae, p. 657-674. In E. W. Jones, J. R. Pringle, and J. R. Broach (ed.), The molecular and cellular biology of the yeast Saccharomyces cerevisiae, vol. 2. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
- 39.Tzung, K. W., R. M. Williams, S. Scherer, N. Federspiel, T. Jones, N. Hansen, V. Bivolarevic, L. Huizar, C. Komp, R. Surzycki, R. Tamse, R. W. Davis, and N. Agabian. 2001. Genomic evidence for a complete sexual cycle in Candida albicans. Proc. Natl. Acad. Sci. USA 98:3249-3253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Van Arsdell, S. W., and J. Thorner. 1987. Hormonal regulation of gene expression in yeast, p. 325. In D. Granner (ed.), Transcriptional control mechanisms. A. R. Liss, New York, N.Y.
- 41.Wilson, R. B., D. Davis, B. M. Enloe, and A. P. Mitchell. 2000. A recyclable Candida albicans URA3 cassette for PCR product-directed gene disruptions. Yeast 16:65-70. [DOI] [PubMed] [Google Scholar]