

Abstract
Background
The knowledge of mosquito species diversity and the level of anthropophily exhibited by each species in a region are of great importance to the integrated vector control. Culicine species are the primary vectors of Japanese encephalitis (JE) virus and filariasis in China. Anopheles sinensis plays a major role in the maintenance of Plasmodium vivax malaria transmission in China. The goal of this study was to compare the abundance and host-seeking behavior of culicine species and An. sinensis in Yongcheng city, a representative region of P. vivax malaria. Specifically, we wished to determine the relative attractiveness of different animal baits versus human bait to culicine species and An. sinensis.
Results
Culex tritaeniorhynchus was the most prevalent mosquito species and An. sinensis was the sole potential vector of P. vivax malaria in Yongcheng city. There were significant differences (P < 0.01) in the abundance of both An. sinensis and Cx. tritaeniorhynchus collected in distinct baited traps. The relative attractiveness of animal versus human bait was similar towards both An. sinensis and Cx. tritaeniorhynchus. The ranking derived from the mean number of mosquitoes per bait indicated that pigs, goats and calves frequently attracted more mosquitoes than the other hosts tested (dogs, humans, and chickens). These trends were similar across all capture nights at three distinct villages. The human blood index (HBI) of female An. sinensis was 2.94% when computed with mixed meals while 3.70% computed with only the single meal. 19:00~21:00 was the primary peak of host-seeking female An. sinensis while 4:00~5:00 was the smaller peak at night. There was significant correlation between the density of female An. sinensis and the average relative humidity (P < 0.05) in Wangshanzhuang village.
Conclusions
Pigs, goats and calves were more attractive to An. sinensis and Cx. tritaeniorhynchus than dogs, humans, and chickens. Female An. sinensis host-seeking activity mainly occurred from 19:00 to 21:00. Thus, we propose that future vector control against An. sinensis and Cx. tritaeniorhynchus in the areas along the Huang-Huai River of central China should target the interface of human activity with domestic animals and adopt before human hosts go to bed at night.
Keywords: Host-seeking behavior, mosquito, culicine species, Anopheles vectors, ecology, malaria elimination
Background
Malaria remains a serious global public health threat and causes substantial morbidity and mortality in the world. The current estimate of human lives at risk from Plasmodium vivax malaria is 2.6 billion [1,2] and South and East Asia account for 52% of the total P. vivax malaria burden [3]. The estimated global cost of P. vivax malaria, including lost productivity, cost of health care, and transport to clinics, is between U.S. $1.4 and $4 billion per year [3]. The interruption of P. vivax malaria transmission worldwide is still one of the greatest challenges for international health and development communities [4]. Despite significant reductions in the overall burden of malaria in the 20th century [5-7], this parasitic disease still represents a major public health problem in China [6,8], with dramatic re-emergence in the Huang-Huai River region of central China in 2001 [5]. In Henan Province [9-13] and Anhui Province [14,15], large-scale epidemics in recent years have caused a major public health concern. This situation not only has an impact on the region's economic development and people's living standards, but also poses a challenge for the routine malaria control strategy. In response to the global initiative to eradicate malaria [16,17], an action plan for malaria elimination was proposed by the Chinese Ministry of Health in 2009 and a national elimination campaign was launched by the Chinese Government in 2010, to eliminate malaria in most endemic regions by 2015 and to achieve ultimate national elimination by 2020 [18].
Anopheles sinensis, Anopheles lesteri [19,20], Anopheles minimus and Anopheles dirus are considered to be four important vector species of malaria in China [21,22]. However, malaria outbreaks and re-emergences were only in areas with An. sinensis in recent years [5]. In studies conducted in the areas along the Huang and Huaihe River, An. sinensis plays an important role in the maintenance of P. vivax malaria transmission [23,24]. Yongcheng city, as a re-emergence region of P. vivax malaria in Henan Province, is one of the major malaria epidemic areas [25]. After extensive control efforts, malaria was nearly eliminated in the 1990s [9,25]; however, epidemics resurged and maintained in the area in the 2000s with the highest cases of 2,890 in 2006 [25,26]. According to surveillance data of Yongcheng Center for Disease Control and Prevention (Yongcheng CDC), a decline in the malaria incidence rate was observed [27]. Based on a study conducted by Zhou et al in 2010, An. sinensis was considered to be the sole potential vector of P. vivax malaria in Yongcheng city [5,25,27]. The estimated vectorial capacity of An. sinensis population in this area was 2.78 times higher in 2010 (0.4983) than in the 1990s [28]. The epidemiological consequences of this change are unknown, but this observed entomological change cannot be ignored when transitioning from malaria control to elimination in China. In recent years, several factors, such as the adjustment of regional agricultural structures and the reduction of biological barriers (calf, pig, etc.), are likely to increase human-vector contact and increase the transmission rate of P. vivax malaria [5].
Vector abundance, host-seeking behavior and preference [29] are all important components of disease-transmission cycles [29,30]. Host-seeking behavior has been operationally defined as the in-flight orientation of the avid female toward a potential blood meal host [31]. Host-seeking typically commenced shortly after sunset and usually peaks during the succeeding one to three hours, the hottest and driest period at night [32,33]. The host-seeking strategies used by arthropod vectors can, in part, affect the efficiency of disease transmission [30,34]. Mosquitoes which prefer to feed on animals are less important in transmitting human disease than those which prefer to feed on humans [35]. In particular, the knowledge of vector abundance and host-seeking activities is of critical importance for integrated vector control. Culex tritaeniorhynchus is the primary vector of the Japanese encephalitis (JE) virus [36,37], and Culex pipiens pallens is the primary vector of the JE virus and filariasis in China. The host-seeking behavior of culicine species is of great significance in the epidemiology of the JE virus and filariasis and the host-seeking behavior of Anopheline mosquitoes is of great significance in the epidemiology of malaria [38,39]. The host-seeking behavior can be studied by a variety of methods [40,41], such as human landing catch (HLC) [42], human bait indoors and outdoors [38,43], indoor residual spray (IRS), baited mosquito nets, baited resting boxes [44], light traps [45,46], etc. Several approaches including serologic techniques, enzyme-linked immunosorbent assay (ELISA) and DNA-based techniques, have been applied to study the blood-feeding behavior of mosquitoes, black flies, ticks and other blood-feeding arthropods as it relates to host-parasite interactions and pathogen transmission [47-58].
Since culicine species and An. sinensis play a distinct role in disease transmission, the abundance and host-seeking behavior of culicine species and An. sinensis are of fundamental epidemiological importance [38], however, little information is available regarding species composition, abundance, host-seeking behavior, and the level of anthropophily exhibited by each species in Yongcheng city. The goals of this study were to compare the abundance and host-seeking behavior of culicine species and An. sinensis in villages of Yongcheng city characterized by different levels of historical incidence of P. vivax malaria and determine the relative attractiveness of different animal baits versus human bait to culicine species and An. sinensis. Such information is required in order to understand the comprehensive vector biology of culicine species and An. sinensis, for the design of effective vector control strategies and for implementation of the ongoing malaria elimination campaign in China.
Methods
Study site
The current study was conducted in three villages of Yongcheng city characterized by different levels of historical incidence of P. vivax malaria. These included a high risk village Dingtang (Lizhai township, annual average incidence rate > 100/100,000), an intermediate risk village Renhu (Houling township, annual average incidence rate 10~100/100,000 and a low risk village Wangshanzhuang (Chenji township, annual average incidence rate < 10/100,000) [25], (Figure 1). The inhabitants of these villages live in houses which are made with bricks. These villages are located in an area between latitudes of 33°42' and 34°18', and longitudes of 115°58' and 116°39'. The Tuohe River is the main environmental feature in the area. The river breaks into small ponds during the hot and dry season, and ponds which are formed by the river become adequate breeding habitats of An. sinensis and culicine species. Most of the regions are plain at 33 meters altitude. The climate is warm temperate from May to October, and the annual average temperature is 14.3°C. The range of annual rainfall is between 556.2 mm and 1,648.9 mm, and mostly rainfall is concentrated to June, July, August, and September. The main crops of these villages are wheat, soybean, corn, and a small amount of cotton and potato. During summer, most of local residents tend to sleep outdoors [21].
Figure 1.
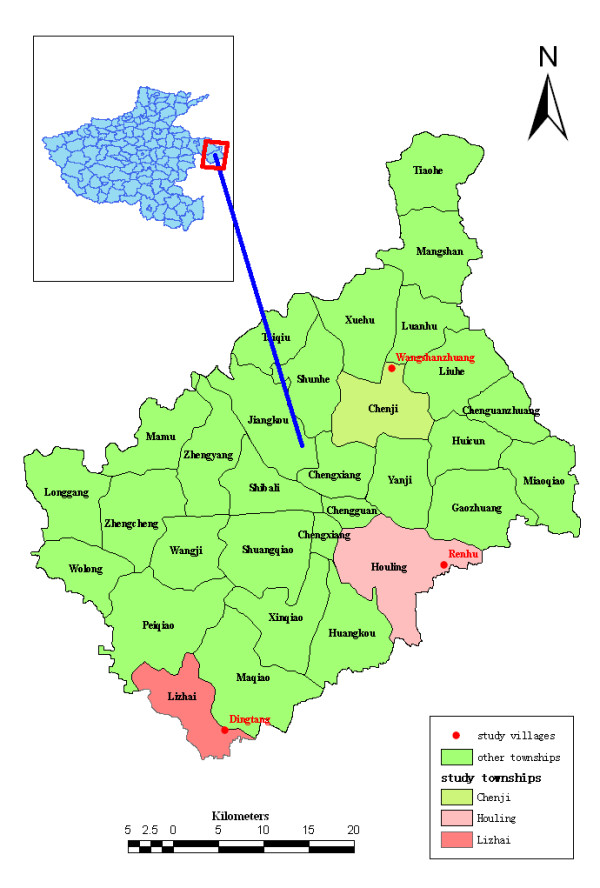
Map showing the study villages in Yongcheng city of Henan Province, People's Republic of China. Red dot represents the study village; Green square frame represents the boundary of Yongcheng city; Yellow square frame represents the low risk township, Chenji; A little pink square frame represents the intermediate risk township, Houling; Pink square frame represents the high risk township, Lizhai.
Besides the difference in the levels of historical incidence of P. vivax malaria, other characteristic differences between the three studied villages are as follows: First, Dingtang village is adjacent to Guoyang County, and Renhu village is adjacent to Suixi County. Guoyang and Suixi County are unstable regions of P. vivax malaria in Anhui Province while Wangshanzhuang village is not adjacent to Anhui Province. Second, the water-body distributions and appropriate breeding habitats of culicine species and An. sinensis larvae in Dingtang village and Renhu village are more than those in Wangshanzhuang village. During the study period, there were many ponds and canals that contained endogenous lotus, and a large number of Anopheline mosquitoes and culicine species larvae were observed in Renhu village and Dingtang village. Third, the number of animal hosts in Renhu village and Dingtang village was larger than that of Wangshanzhuang village. Based on the investigative data of the village doctors, Dingtang village covers approximately 3,500 acres of arable land, with a population of 1,890 people, 1 calf, and 150 goats within 480 households, Renhu village covers approximately 4,500 acres of arable land, with a population of 1,875 people, 60 calves, 70 pigs, and 180 goats within 470 households, Wangshanzhuang village covers approximately 1,600 acres of arable land, with a population of 750 people, and 5 calves, 45 pigs, and 70 goats within 170 households, respectively.
Host-seeking behavior experiments
Host-seeking behavior experiments of culicine species and An. sinensis were conducted from August 28th to October 5th. The reason that these experiments were conducted during this period is that this period was considered to be the epidemic episode of P. vivax malaria along the Huaihe River region in China [59]. Three villages were sampled in different weeks because of limited man power and material resources. Outdoor bednet traps [34,60] were employed to study the host-seeking activity of culicine species and An. sinensis. The size of the bednet traps was 3.6 m × 2.4 m × 1.6 m. There was twenty centimeters between the floor and the bottom of the bednet traps. The locations of the bednet traps were close to ponds, ditches and drains, in a line or ellipse, at intervals of 50 m. The distance from the bednet traps to the nearest resident's house was about 60 m, and people living in the range of this distance near to the water-bodies had higher risk of malaria infection than those living beyond the distance [5]. In each sampled village, major local domestic animals, such as calves, pigs, goats, dogs, and chickens, were used as animal baits to attract mosquitoes to the bednet traps. Some members of staff at Yongcheng CDC and China CDC were selected as human baits. These members (under double bednet traps to avoid mosquitoes bites) were also used repeatedly throughout the entire duration of the study to avoid bias [35,45].
To control for the plausible confounding bias of locations of the bednet traps and collection nights, one 5 × 5 Latin square design and two 6 × 6 Latin square design experiments [34,46,61] were conducted. The Latin square design is used where researchers desire to control the variation in an experiment that is related to rows and columns in the field. Treatments (baits) were assigned at random within rows (nights) and columns (locations of the bednet trap), with each treatment once per row and once per column [34]. During this period, three independent experiments were conducted, with one experiment per village. A 5 × 5 Latin square design experiment was conducted in Dingtang village because five types of bait, namely calves, goats, dogs, chickens and humans, existed in this village while two 6 × 6 Latin square design experiments were conducted in Renhu village and Wangshanzhuang village because six types of bait, namely calves, pigs, goats, dogs, chickens, and humans existed in these two villages.
Before the experiment began, the bednet traps were fixed at previously selected locations. The locations of the bednet traps were alternated every night, each bednet trap being moved with the bait to avoid the effect of odor contamination. Calves, pigs, dogs, goats, and chickens were placed into the bednet traps. Dogs and pigs were put into metal cages under the bednet traps to prevent staff from being bitten by them (Figure 2). Metal cages were welded in Yongcheng city and the bednet traps were fixed before 19:00. The interval between 19:00 and 7:00 was decided upon by consulting the literature, which reported the host-seeking behavior of culicine species and An. sinensis [61-63]. Calves and pigs were chosen as young small animals and experiments were carried out with one calf, one pig, two goats, two dogs, and five chickens at a time under the bednet traps [34]. Every hour, all mosquitoes inside the bednet traps were collected by an electrical aspirator for 15 minutes per hour throughout the 12 hour period [64]. Collection time periods were subdivided into early evening (19:00~22:00), late evening (22:00~01:00), post-midnight (01:00~ 04:00), and pre-dawn (04:00~07:00) [64]. The staff was divided into four teams with three people in each team. The first team worked for the first three hours from 19:00 to 22:00, followed by the second team from 22:00 to 01:00. The third team worked from 01:00 to 04:00, and the last team worked from 04:00 to 07:00.
Figure 2.

Map showing the photos of bednet traps in the field in Yongcheng city of Henan Province, People's Republic of China. Photo A represents pig-baited bednet trap; Photo B represents human-baited bednet trap; Photo C represents dog-baited bednet trap; Photo D represents calf-baited bednet trap; Photo E represents goat-baited bednet trap; Photo F represents chicken-baited bednet trap.
Data was recorded for collection dates and sites, sequences of locations of the bednet traps, collection nights, and numbers of each mosquito species. Temperature (°C) and relative humidity (%) were the average value from a weather web in China http://www.weather.com.cn. Ambient outdoor air temperature and relative humidity of each collection hour was recorded using a WS-1 Thermo-Hygrometer device. Since some events such as heavy rains would prevent host-seeking activity, mosquito sampling occasions were scheduled based on the pattern of weather conditions. During the trapping days, however, rainfall values (mean minimum and maximum in mm) were obtained from a local meteorological station located in Yongcheng city, so that the effect of this environmental factor could be evaluated while controlling for between-night variation in mosquito abundance.
Blood meal identification
Collected mosquitoes were narcotized by ether and morphologically identified in the field using commonly accepted guidelines [65,66]. All collected Anopheline mosquitoes were put into 1.5 ml centrifuge tubes. All blood-fed female An. sinensis were put individually into tubes and then transported to the laboratory of The Department of Vector Biology and Control in China CDC. To reveal the species of Anopheline mosquitoes in Yongcheng city [5,66], a ribosomal DNA PCR assay was conducted [67-72]. Fifty percent of blood-fed female Anopheline mosquitoes which were selected as equal proportions from different bednet traps in three villages were further identified to species. A Qia Amp DNA Mini Kit (Qiagen Inc., CA) was used and DNA was extracted from the thorax of An. sinensis according to the manufacturer's instructions. The PCR conditions used were identical to a study of Ma et al in China [68]. A multiplex PCR assay was conducted to identify the origins of blood meal from the same Anopheline mosquitoes used for species identification. The whole blood of domestic animals and humans was obtained in Yongcheng city. Blood DNA of blood-fed female An. sinensis was extracted from the mosquitoes' abdomen and the multiplex PCR conditions and primers used in the current study were the same as that previously described by Kent & Norris in 2005 [52] (Table 1).
Table 1.
Primer sequences for the cytochrome b-based polymerase chain reaction blood meal identification assay
| 1Primers | 5'-3'sequences | Product size with Unrev1025 |
|---|---|---|
| Human741F | GGC TTA CTT CTC TTC ATT CTC TCC T | 334 |
| Cow121F | CAT CGG CAC AAA TTT AGT CG | 561 |
| Pig573F | CCT CGC AGC CGT ACA TCT C | 453 |
| Goat894F | CCT AAT CTT AGT ACT TGT ACC CTT CCT C | 132 |
| Dog368F | GGA ATT GTA CTA TTA TTC GCA ACC AT | 680 |
| Unrev1025 | GGT TGT CCT CCA ATT CAT GTT A | - |
1Primers of this multiplex PCR assay were designed based on the sequence differences in cytochrome b sequences.
Data analyses
The species composition of mosquitoes in the three villages was calculated. Abundance (mean catch per bait per night) of culicine species and An. sinensis were calculated for each site and bait. The number of culicine species and An. sinensis of each bait was log (x+1) transformed and subjected to variance analysis after a satisfactory check for normality of the distribution. Univariate analysis of variance (general linear models functions) was used to test the main effect and interaction effects of bait types, collection nights, and locations of the bednet traps. Probabilities of the Univariate analysis of variance were at α = 0.05 level. Pair wise comparison had been performed to rank host-seeking behavior of culicine species and An. sinensis. Normal distributions of the An. sinensis density, the average temperature, and the average relative humidity were tested by One-sample of Kolmogorove-Smirnov Test before using Pearson correlation analysis. The association between the density of An. sinensis and the average temperature, between the density of An. sinensis and the average relative humidity were examined by Pearson correlation analysis. Data was entered and analyzed by SPSS (Statistical Package for the Social Sciences) statistical software (version 11.5).
Ethical approval
Ethical approval for this study was obtained from the Ethical Committee of China CDC and permission was also obtained from the Municipal Government, the Municipal Health Bureau and CDC in Yongcheng city.
Results
Species composition
A total of 35,312 mosquitoes were captured in the three villages during the study period, of which 76.6% (n = 27,048) were Cx. tritaeniorhynchus, 12.2% (n = 4,313) were Cx. pipiens pallens, 10.6% (n = 3,755) were An. sinensis, 0.02% (n = 9) were Aedes albopictus, 0.5% (n = 165) were Armigeres subalbatus (Coquillett), and 0.1% (n = 22) were others. Cx. tritaeniorhynchus was the most prevalent mosquito species, and An. sinensis was the sole potential vector of P. vivax malaria in Yongcheng city. As far as sex proportions of captured An. sinensis is concerned, 1.25% (n = 47) out of 3, 755 An. sinensis were males, while 98.75% (n = 3,708) were females. Fifty percent (n = 102) of blood-fed female Anopheline mosquitoes, which were sampled, about the same proportions from the bednet traps in the three villages, were further identified to species by ribosomal DNA PCR assay. All collected adult female Anopheline mosquitoes examined belonged to An. sinensis, confirming that there was not any other species of Anopheline mosquitoes of Hyrcanus Complex in Yongcheng city [72,73].
The abundance of female culicine species and An. sinensis per baited trap at each study site
The abundance of female culicine species and An. sinensis per baited trap at each study site was calculated (Tables 2 &3). There were significant differences in the abundance of female An. sinensis for each bait [F (5, 1153) = 53.722, P < 0.01]. There were significant differences in the abundance of female Cx. tritaeniorhynchus for each bait [F (5, 1153) = 28.261, P < 0.01]. There were no differences in the abundance of female Cx. pipiens pallens for each bait [F (5, 1153) = 1.111, P = 0.353]. Pair wise comparison showed that the rank of abundance of host-seeking female An. sinensis for each bait from high to low was pigs (7.27 ± 11.146 catch per bait per night), goats (5.14 ± 8.614 catch per bait per night), calves (4.81 ± 7.553 catch per bait per night), dogs (1.62 ± 4.802 catch per bait per night), humans (0.90 ± 2.364 catch per bait per night), and chickens (0.65 ± 1.422 catch per bait per night). The rank of abundance of host-seeking female Cx. tritaeniorhynchus for each bait from high to low was pigs (49.83 ± 115.660 catch per bait per night), goats (31.03 ± 55.729 catch per bait per night), calves (19.84 ± 45.657 catch per bait per night), dogs (16.93 ± 34.208 catch per bait per night), humans (6.19 ± 11.381 catch per bait per night), and chickens (6.18 ± 17.569 catch per bait per night). These findings suggested that there was a consistent trend of the rank of abundance of host-seeking female An. sinensis and Cx. tritaeniorhynchus.
Table 2.
1The abundance of female culicine species and An. sinensis in the study sites of Yongcheng city, Henan Province, China
| 2The study sites | The species of mosquitoes | 3N | Mean | Std. Deviation | Std. Error | 95% Confidence Interval for Mean | |
|---|---|---|---|---|---|---|---|
| Lower Bound | Upper Bound | ||||||
| Dingtang village | An. sinensis | 407 | 1.38 | 2.72 | 0.16 | 1.07 | 1.69 |
| Cx. tritaeniorhynchus | 7,631 | 25.87 | 48.79 | 2.84 | 20.28 | 31.46 | |
| Cx. pipiens pallens | 131 | 0.44 | 2.22 | 0.13 | 0.19 | 0.70 | |
| Renhu village | An. sinensis | 2,960 | 6.85 | 10.10 | 0.49 | 5.90 | 7.81 |
| Cx. tritaeniorhynchus | 14,019 | 32.45 | 77.32 | 3.72 | 25.14 | 39.76 | |
| Cx. pipiens pallens | 2,782 | 6.44 | 20.95 | 1.01 | 4.46 | 8.42 | |
| Wangshan- zhuang village |
An. sinensis | 341 | 0.79 | 1.96 | 0.09 | 0.60 | 0.97 |
| Cx. tritaeniorhynchus | 1,801 | 4.17 | 11.43 | 0.55 | 3.09 | 5.25 | |
| Cx. pipiens pallens | 292 | 0.68 | 1.29 | 0.06 | 0.55 | 0.80 | |
1 The abundance refers to mean catch per bait per night.
2 Dingtang village in Lizhai township belongs to high levels of historical incidence of P. vivax malaria; Renhu village in Houling township belongs to intermediate levels of historical incidence of P. vivax malaria; Wangshanzhuang village in Chenji township belongs to low levels of historical incidence of P. vivax malaria.
3 N represents the total number of female mosquitoes in the study sites.
Table 3.
1The abundance of female culicine species and An. sinensis in bednet traps baited with different animal and human hosts in Yongcheng city, Henan Province, China
| Bait types | The species of mosquitoes | 2N | Mean | Std. Deviation | Std. Error | 95% Confidence Interval for Mean | |
|---|---|---|---|---|---|---|---|
| Lower Bound | Upper Bound | ||||||
| Human | An. sinensis | 182 | 0.90 | 2.364 | 0.166 | 0.57 | 1.22 |
| Cx.tritaeniorhynchus | 1,256 | 6.19 | 11.381 | 0.799 | 4.61 | 7.76 | |
| Cx. pipiens pallens | 315 | 1.55 | 3.315 | 0.233 | 1.09 | 2.01 | |
| Calf | An. sinensis | 977 | 4.81 | 7.553 | 0.530 | 3.77 | 5.86 |
| Cx.tritaeniorhynchus | 4,028 | 19.84 | 45.657 | 3.205 | 13.52 | 26.16 | |
| Cx. pipiens pallens | 318 | 1.57 | 4.732 | 0.332 | 0.91 | 2.22 | |
| Goat | An. sinensis | 1,043 | 5.14 | 8.614 | 0.605 | 3.95 | 6.33 |
| Cx.tritaeniorhynchus | 6,300 | 31.03 | 55.729 | 3.911 | 23.32 | 38.75 | |
| Cx. pipiens pallens | 576 | 2.84 | 12.787 | 0.897 | 1.07 | 4.61 | |
| Pig | An. sinensis | 1,047 | 7.27 | 11.146 | 0.929 | 5.43 | 9.11 |
| Cx.tritaeniorhynchus | 7,175 | 49.83 | 115.660 | 9.638 | 30.77 | 68.88 | |
| Cx. pipiens pallens | 919 | 6.38 | 29.694 | 2.475 | 1.49 | 11.27 | |
| Dog | An. sinensis | 328 | 1.62 | 4.802 | 0.337 | 0.95 | 2.28 |
| Cx.tritaeniorhynchus | 3,437 | 16.93 | 34.208 | 2.401 | 12.20 | 21.67 | |
| Cx. pipiens pallens | 622 | 3.06 | 10.302 | 0.723 | 1.64 | 4.49 | |
| Chicken | An. sinensis | 131 | 0.65 | 1.422 | 0.100 | 0.45 | 0.84 |
| Cx.tritaeniorhynchus | 1,255 | 6.18 | 17.569 | 1.233 | 3.75 | 8.61 | |
| Cx. pipiens pallens | 455 | 2.24 | 7.279 | 0.511 | 1.23 | 3.25 | |
1 The abundance refers to mean catch per bait per night.
2 N represents the total number of female mosquitoes of distinct types of bait.
Host-seeking activity of female culicine species and An. sinensis
As to the host-seeking activity of female Cx. tritaeniorhynchus, the results of Univariate analysis of variance demonstrated that there were significant differences among the number of female Cx. tritaeniorhynchus captured at each bait [F (5,1105) = 8.117, P < 0.01], and at each bednet location [F(5,1105) = 3.413, P = 0.005], but no differences at each night [F(5,1105) = 1.648, P = 0.145]. There were no interactions between nights and locations of the bednet traps [F (3, 1105) = 0.031, P = 0.993], locations of the bednet traps and baits [F (5, 1105) = 1.782, P = 0.114], and there were interactions between nights and baits [F (3, 1105) = 2.233, P = 0.08]. As far as the host-seeking activity of female Cx. pipiens pallens is concerned, there were significant differences among the number of female Cx. pipiens pallens captured at each bait [F(5,1105) = 6.989, P < 0.01], at each night [F(5,1105) = 7.118, P < 0.01], and at each bednet location [F(5,1105) = 3.257, P = 0.006]. There were interactions between nights and locations of the bednet traps [F (3, 1105) = 4.170, P = 0.006], nights and baits [F (3, 1105) = 3.903, P = 0.009], and locations of the bednet traps and baits [F (5, 1105) = 3.015, P = 0.01]. There were significant differences among the number of female An. sinensis captured at each bait [F (5, 1105) = 13.161, P < 0.01], at each night [F (5, 1105) = 4.460, P < 0.01], but no differences of each bednet location [F (5, 1105) = 0.259, P = 0.90]. There were no interactions between nights and locations of the bednet traps [F (3, 1105) = 0.912, P = 0.434], nights and baits [F (3, 1105) = 1.961, P = 0.118], and locations of the bednet traps and baits [F (5, 1105) = 1.094, P = 0.362].
Multiple blood meals of female engorged An. sinensis highlighted by blood meal analysis
We used a host-specific multiplex PCR assay based on the mitochondrial cytochrome b gene to identify the mammalian blood meals of field-collected blood-fed female An. sinensis. This PCR assay correctly identified the blood meals of control groups of blood of different domestic animals 100% of the time and yielded no amplification of unfed An. sinensis template DNA. Ethidium bromide-stained agarose gel showed host-specific cytochrome b PCR products amplified from whole blood DNA extractions and collected female blood-fed An. sinensis in the field (Figure 3). No blood-fed female An. sinensis were collected in chicken-baited mosquito nets during this period. Therefore, only four types of the most common domestic animals in the study sites and humans were included in this multiplex PCR assay. A total of 102 samples of blood-fed female An. sinensis were sampled from three villages in Yongcheng city. The result showed that 81 blood meals belonged to a single origin, including pigs (n = 24), calves (n = 23), goats (n = 16), dogs (n = 4), humans (n = 3), and others (n = 11) while 21 blood meals belonged to mixed origin, including calf/goat (n = 5), calf/pig (n = 9), pig/dog (n = 1), calf/pig/goat (n = 2), calf/pig/dog (n = 2), and others (n = 2). The HBI calculated from identification of blood meals of blood-fed An. sinensis including mixed meals was 2.94% and 3.70% when computed with only the single meals (Table 4). High proportions of mixed meals were encountered (25.9%).
Figure 3.

Ethidium bromide-stained agarose gel showing host-specific cytochrome b polymerase chain reaction products amplified from whole blood DNA extractions and engorged female An. sinensis in the field. Control products amplified from whole blood are shown in lane 1 (calf, 561 bp), lane 2 (pig, 453 bp), lane 3 (goat, 132 bp), lane 4 (dog, 680 bp), and lane 5 (human, 334 bp), respectively. Engorged female An. sinensis in three villages in Yongcheng city are shown in lane 6-22, respectively. Lane 23, negative control. Outside lanes are 100 bp DNA ladders. bp = basepairs.
Table 4.
Origin of blood meal of engorged female An. sinensis in the study sites during the work period
|
1Origin of blood meal of engorged female An.sinensis |
The research sites | Total | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dingtang village | Renhu village | Wangshanzhuang village | ||||||||||||||
| Human bednet trap | Calf bednet trap |
Goat bednet trap |
2Pig bednet trap |
Dog bednet trap |
Human bednet trap | Calf bednet trap |
Goat bednet trap |
Pig bednet trap |
Dog bednet trap |
Human bednet trap | Calf bednet trap |
Goat bednet trap |
Pig bednet trap |
3Dog bednet trap |
||
| Single host | 0 | |||||||||||||||
| Human | 3 | 3 | ||||||||||||||
| Goat | 2 | 9 | 1 | 4 | 16 | |||||||||||
| Dog | 2 | 2 | 4 | |||||||||||||
| Pig | 2 | 1 | 12 | 1 | 8 | 24 | ||||||||||
| Calf | 1 | 7 | 1 | 7 | 1 | 2 | 3 | 1 | 23 | |||||||
| Other | 3 | 1 | 1 | 2 | 1 | 3 | 11 | |||||||||
| Mixed host | 0 | |||||||||||||||
| Calf/goat | 1 | 4 | 5 | |||||||||||||
| Calf/pig | 2 | 6 | 1 | 9 | ||||||||||||
| Pig/dog | 1 | 1 | ||||||||||||||
| Calf/pig/dog | 2 | 2 | ||||||||||||||
| Calf/pig/goat | 1 | 1 | 2 | |||||||||||||
| Other | 1 | 1 | 2 | |||||||||||||
| Total | 15 | 8 | 10 | 0 | 5 | 2 | 9 | 9 | 14 | 5 | 3 | 12 | 2 | 8 | 0 | 102 |
1Numbers of single and mixed blood meals in Yongcheng city. 2No pig was kept by the residents of Dingtang village. 3No blood-fed An. sinensis was captured from dog-baited bednet trap in Wangshanzhuang village. No blood-fed An. sinensis were captured from chicken-baited bednet trap.
Hourly host-seeking activity of female An. sinensis
There were significant differences among the average number of captured female An. sinensis at different hours at night [F (11, 3697) = 32.542, P < 0.01] (Table 5). Host-seeking activity of female An. sinensis showed a similar bimodal pattern at all study sites. The primary peak of female An. sinensis was from 19:00 to 21:00 while the smaller peak was from 4:00 to 5:00.
Table 5.
Variations in the hourly catches of host-seeking female An. sinensis in bednet traps baited with different animal and human hosts during 17 nights in Yongcheng city, Henan Province, China
| Hour interval | 1N | 2Mean | Std. Deviation | Std. Error | 95% Confidence Interval for Mean |
|
|---|---|---|---|---|---|---|
| Lower Bound | Upper Bound | |||||
| 19:00~20:00 | 426 | 23.58 | 21.074 | 1.021 | 21.58 | 25.59 |
| 20:00~21:00 | 494 | 26.98 | 20.735 | 0.933 | 25.14 | 28.81 |
| 21:00~22:00 | 307 | 18.50 | 14.275 | 0.815 | 16.90 | 20.11 |
| 22:00~23:00 | 286 | 19.55 | 19.555 | 1.156 | 17.27 | 21.82 |
| 23:00~0:00 | 255 | 11.68 | 10.689 | 0.669 | 10.36 | 13.00 |
| 0:00~1:00 | 218 | 9.61 | 6.513 | 0.441 | 8.74 | 10.47 |
| 1:00~2:00 | 296 | 16.79 | 11.468 | 0.667 | 15.48 | 18.10 |
| 2:00~3:00 | 311 | 16.38 | 11.034 | 0.626 | 15.15 | 17.61 |
| 3:00~4:00 | 255 | 15.77 | 13.583 | 0.851 | 14.09 | 17.44 |
| 4:00~5:00 | 315 | 20.20 | 13.693 | 0.772 | 18.68 | 21.72 |
| 5:00~6:00 | 232 | 14.86 | 12.238 | 0.803 | 13.28 | 16.45 |
| 6:00~7:00 | 313 | 16.97 | 12.395 | 0.701 | 15.60 | 18.35 |
| Total | 3,708 | 18.59 | 16.091 | 0.264 | 18.07 | 19.11 |
1N represents the total number of female mosquitoes of each hour. 2The mean of captured mosquitoes refers to the average number of An. sinensis per net per hour.
Variation in the density of female An. sinensis with environment variables
Table 6 represents the density of female An. sinensis relative to the average temperature and the average relative humidity in the study sites. Each day's collection (12 hours capture) represents the total number of female An. sinensis captured in three sites and showed the variation in the density of female An. sinensis according to the study period. Pearson's correlation analysis showed that there were no correlations between the density of female An. sinensis and the average temperature (r = -0.038, P = 0.951) or the average relative humidity (r = -0.041, P = 0.947) in Dintang village. There were no correlations between the density of female An. sinensis and the average temperature (r = -0.658, P = 0.155) or the average relative humidity (r = -0.598, P = 0.210) in Renhu village. There were no correlations between the density of female An. sinensis and the average temperature (r = 0.610, P = 0.199), but significant correlations between the density of female An. sinensis and the average relative humidity (r = 0. 859, P = 0.029) in Wangshanzhuang village.
Table 6.
The distribution of the An. sinensis density, the average temperature, and the average relative humidity in the study sites
| 1The study sites | Parameters | 3N | Mean | Std.Deviation | Minimum | Maximum | One-Sample Kolmogorov-Simirnov Test | |
|---|---|---|---|---|---|---|---|---|
| Kolmogorov-Simirnov Z | 4Sig. | |||||||
| Dingtang village | 2The An. sinensis density(mosquito/net/night) | 5 | 16.28 | 7.25 | 7.00 | 24.40 | 0.482 | 0.974 |
| The average temperature(°C) | 5 | 26.62 | 3.68 | 23.00 | 31.50 | 0.531 | 0.940 | |
| The average relative humidity (%) | 5 | 84.20 | 9.70 | 71.00 | 95.00 | 0.422 | 0.994 | |
| Renhu village | The An. sinensis density(mosquito/net/night) | 6 | 82.22 | 27.13 | 34.67 | 111.67 | 0.490 | 0.970 |
| The average temperature(°C) | 6 | 23.63 | 3.42 | 19.50 | 28.50 | 0.475 | 0.978 | |
| The average relative humidity (%) | 6 | 78.83 | 11.80 | 60.00 | 90.00 | 0.488 | 0.971 | |
| Wangshan-zhuang village | The An. sinensis density(mosquito/net/night) | 6 | 9.47 | 9.67 | 2.50 | 28.50 | 0.932 | 0.351 |
| The average temperature(°C) | 6 | 17.88 | 2.77 | 13.80 | 21.00 | 0.449 | 0.988 | |
| The average relative humidity (%) | 6 | 84.17 | 6.77 | 74.00 | 95.00 | 0.555 | 0.918 | |
1Dingtang village in Lizhai township belongs to high levels of historical incidence of P. vivax malaria; Renhu village in Houling township belongs to intermediate levels of historical incidence of P. vivax malaria; Wangshanzhuang village in Chenji township belongs to low levels of historical incidence of P. vivax malaria. 2The density of An. sinensis refers to a total of female An. sinensis per net per night. 3N refers to the total of night. 4Sig. = significance.
Discussion
The results obtained from the current research demonstrated that the species composition of mosquitoes in Yongcheng city includes Cx. tritaeniorhynchus, Cx. pipiens pallens, An. sinensis, Ae. albopictus, and Ar. subalbatus(Coquillett). This was the first time that the species composition of mosquitoes was reported in Yongcheng city. Though Cx. tritaeniorhynchus, Cx. pipiens pallens, Ae. albopictus, and Ar. subalbatus(Coquillett) are not vectors of P. vivax malaria, these species could be considered as the major nuisance on the basis of their high proportions. The influence of culicine species, especially Cx. tritaeniorhynchus and Cx. pipiens pallens should not be ignored, since Cx. tritaeniorhynchus is the primary vector of JE virus, and Cx. pipiens pallens is the primary vector of JE virus and filariasis in China [36]. The ribosomal DNA PCR assay revealed that An. sinensis was the sole species of Hyrcanus Complex in identified female Anopheline mosquitoes in the study sites. The results of this study were consistent with those reported earlier in China [5,25,27]. Based on previous researches and results of the current PCR assay, it was probable that there weren't any other species of Anopheline mosquitoes of Hyrcanus Complex except for An. sinensis present in Yongcheng city.
Mosquito host-seeking behavior can be studied by different methods [30,33,74-77]. In the northeastern USA, host-seeking activities and avian host preferences of mosquitoes associated with West Nile virus (WNV) transmission were studied by a custom-designed trap baited with dry ice [77]. To study potential WNV vectors, horse- and bird-baited traps and HLC methods were carried out weekly from May to October 2004 at two Camargue sites [45]. In the Toledo District, Belize, Central America, host-feeding preferences of Anopheline species were collected by manual aspiration, mechanical aspiration, and a vehicle-mounted trap [78]. In the present study, baited bednet traps [34,79] were adopted to study the host-seeking behavior of culicine species and An. sinensis. The current study was similar to a study in Bernalillo County, New Mexico. Hosts used in their study were calves, chickens, dogs, and horses [80]. The rationale for using baited bednet traps in the current study was that they would separate the zoophilic and the anthropophilic species while other methods of collection, such as HLC and IRS may not be suitable to use for this purpose.
Latin square design (LSD) was employed in the present study because the experimental set-up consisted of several baits tested on 5-6 experimental nights. Previous studies had established that the LSD would greatly reduce potential bias related to the location and alternated position of the baited-bednet samplers, allowing comparison of the attractiveness of different hosts to the mosquitoes in the field [34,46].
The results of variance analysis demonstrated that there were significant differences among the abundance of Cx. tritaeniorhynchus for each bait, the abundance of An. sinensis for each bait, but no differences of female Cx. pipiens pallens for each bait. There were some major characteristics, which might influence the abundance of female Cx. tritaeniorhynchus and An. sinensis in these villages. First, during the study period, the locations of the bednet traps of Renhu village were near to a wide canal and many ponds (harboring endogenous lotus). High density of culicine and Anopheline mosquito larvae were observed from these breeding sites. In another experiment concerning breeding sites of Anopheline mosquitoes in 2010, Renhu village had more breeding sites of Anopheline mosquitoes than the other two villages. Second, the number of animal baits from high to low was consistent with the abundance of collected female culicine species and An. sinensis in the study sites. Differences in the availability of blood meals for female culicine species and female An. sinensis might exist within the study villages. Third, the locations selected for our study were sampled at different periods. This could possible explain the observed differences in the number of both Culicine and Anopheline mosquitoes caught between the villages. Fourth, the local selective pressures within which mosquitoes thrive might be added to the variability of the results.
By far, field studies have revealed that each mosquito species had its own host preference [81,82]. Pair wise comparison showed that the rank of abundance of host-seeking female Cx. tritaeniorhynchus and An. sinensis for each bait from high to low was pigs, goats, calves, dogs, humans, and chickens, respectively. There was a consistent trend of the rank of abundance of host-seeking activity between female Cx. tritaeniorhynchus and An. sinensis. In Thailand, An. sinensis was almost entirely zoophilic in comparative biting tests involving man and cow; almost none of them were attracted to man [83]. This phenomenon could be explained by an opportunistic feeding behavior present in these species. The anti-vector behavior at the community level, namely practices often used by local people for protection from mosquitoes, also had some influence on the host-seeking behavior of these species. Olfaction may be the major sensory modality involved in the resource searching behavior of insects [84]. In mosquitoes, it is mainly exploited in host-seeking and finding a suitable place for oviposition. Differences in host-preference in the current study were likely to be reflected in their response to different host odors offered [85-88]. However, the focus on the current research was only one aspect of behavior in mosquitoes and cannot fully explain the dramatic re-emergence of malaria in the Huang-Huai River region of central China. Zhou et al [5] suggested that the spatial distribution between malaria cases and water-body, the changing of meteorological factors, and the increasing vectorial capacity and basic reproductive rate of An. sinensis were the possible determinants of malaria re-emergence in these areas.
In the current study, successful amplification was obtained by template DNA from the whole blood and female blood-fed An. sinensis in the research villages. All bednet traps sampled approximately the same proportions of female blood-fed An. sinensis. The results of multiplex PCR assay showed that only 3 blood meals were human in origin, while most of blood meals originated from domestic animals, such as pigs, calves, and goats. The overall HBI calculated from this study including mixed blood meals was 2.94% and 3.70% when compared with only the single blood meal. Similar studies were reported in Japan in 1951 [89]; in north Kyonggi-do in 1962 (1.7%), in 1999 (0.7%), and in 2000 (0.8%) [90]. Though the HBI of these tests was very low, they readily fed on humans in high numbers where domestic animals were not nearby for feeding [91]. The HBI of the current study was higher than that of these findings mentioned above. Gonotrophic discordance, or taking multiple blood meals during the gonotrophic cycle, has been reported in many different mosquito taxa. In these studies, approximately 10% to 40% of blood-fed field specimens contained multiple blood meals [92-99]. Amplification of template derived from mosquito abdomens containing mixed blood meals confirmed that multiple blood meals from different mammals could be detected in a single mosquito. Our studies demonstrated that high proportions of mixed blood meals were encountered (21/102), and cryptic blood meals were likely to be more numerous. Two to three sources of blood meal were detected from the abdomen of female blood-fed An. sinensis, and this phenomenon could be explained by a modification of the traditional view of the gonotrophic cycle [34]. This host-feeding behavior can influence pathogen transmission through increased frequency of vector-human contact, or possibly reduce vector-human contact if some blood meals are taken from alternative mammalian hosts [52]. In the current study, we failed to identify the origin of the blood meal in 11 out 102 field specimens examined. Insufficiency in host DNA concentrations due to the low blood meal volume in some mosquito specimens is presumed to be the cause of this failure. In addition, the process of blood digestion may have denatured host DNA, or the mosquito may have fed on an animal not included in the diagnostic assay [52].
The current study demonstrated that the host-seeking activity of female An. sinensis showed two peaks. The primary host-seeking activity occurred between 19:00 and 21:00 and a smaller peak between 4:00 and 5:00. It reported that An. sinensis bites man and animals soon after dark and throughout the night, with a peak generally in the first quarter of the night [21]. In east China, its peak was from 19:00 to 21:00 [100]. Chow (1949) found that the peak of An. sinensis biting buffalo in Chongqing was from 20:00 to 22:00. Zhou et al [5] reported that the biting time of An. sinensis was from 19:30 to 24:00 locally, with a peak time from 21:00 to 24:00 in Yongcheng city. The primary peak time in the present study was earlier than that of Zhou' s report [5] and consistent with a previous study in east China [100]. This pattern suggested that vector abatement directed at adult An. sinensis would be most effective if initiated from 19:00 to 21:00, namely before human host go to bed at night.
Previous studies have shown that meteorological factors, such as the average temperature, rainfall, and relative humidity have a certain correlation to mosquito density in the corresponding period [101-105]. Temperature was found to be the most important environmental factor, followed by rainfall and relative humidity in the Delphi evaluation. However, relative humidity was found to be more important than rainfall and temperature in the ranking list according to the three single environmental factor regression models [106]. Paaijmans et al [107] believed that the incubation period for malaria parasites within the mosquito is exquisitely temperature-sensitive, and temperature is a major determinant of malaria risk. Yang et al [98] found that the distribution of the mosquito vector was mainly related to relative humidity, which determined the extent of malaria transmission. In the current study, there were no correlations between the density of female An. sinensis and the average temperature in the study sites, while significant correlations between the density of female An. sinensis and the average relative humidity (r = 0.859, P = 0.029) were observed in Wangshanzhuang village. It could be explained due to the average relative humidity affecting the distribution and breeding of An. sinensis [106].
Care needs to be taken in interpreting the results of this study. Several factors may have affected the variability in the abundance and host-seeking behaviour of the culicine mosquitoes and An. sinensis in our study. First, biases inherent in the trapping method may have affected the observed number and species composition of mosquitoes within baited traps. Some mosquitoes may fly out from the gaps under the baited bednet traps more easily than others, resulting in some species being underrepresented. Second, the physiological status and infected proportions of mosquitoes were not recorded in the field; therefore, the importance of gravid female An. sinensis is not emphasized in the current report. Third, the abundance of Ae. albopictus, Ar. subalbatus (Coquillett), and other mosquito species in each site were not calculated because they were poorly represented in our samples. Further research should be undertaken using this multiplex PCR protocol for the identification of the origin of blood meals of female blood-fed An. sinensis, in addition, further studies are needed to better characterize the malaria vector and its respective role in malaria transmission. An olfactometry assay could be conducted in the future to clarify if behavior of An. sinensis is not directional and possibly provide an indication of how this species will behave in different circumstances. The origin of blood meals of Cx. tritaeniorhynchus could also be studied in the future because of its high proportions and epidemiologic implications [108].
Conclusions
Given the observed data in the studied research sites, the epidemiologic implications of culicine species, especially Cx. tritaeniorhynchus, could not be ignored on the basis of their high proportions. An. sinensis was the sole potential vector of P. vivax malaria in Yongcheng city. Pigs, goats, and calves had a greater tendency to attract both female Cx. tritaeniorhynchus and female An. sinensis than dogs, humans, and chickens at night. High proportions of mixed meals of female engorged An. sinensis were encountered, and cryptic meals were likely to be more numerous. When it comes to the host-seeking activity of female An. sinensis, 19:00~21:00 was the primary peak while 4:00~5:00 was the relatively smaller peak at night. These aspects mentioned above are of great importance to the understanding of vector biology of culicine species and An. sinensis, and the design of effective vector control strategies. These findings should be considered in the implementation of malaria elimination campaigns in China, especially in malaria re-emergence areas.
List of abbreviations used
PCR: polymerase chain reaction; JE: Japanese encephalitis; HLC: human landing catch; IRS: indoor residual spray; HBI: human blood index.
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
LXB and LQY designed and carried out statistical analysis and wrote the paper. GYH, RDS, ZY, LZF and LJL performed data management. JJY, ZGC and LHS were involved in the execution of the research. LQY, LXB, LM, GYH, GWD drafted and wrote up the manuscript. LQY supervised data collection. LQY, GYH, RDS, ZCJ, LHZ and LQ reviewed this paper for publication. All authors approved the final version of the manuscript.
Contributor Information
Xiao-Bo Liu, Email: liuxiaobo19831016@yahoo.com.cn.
Qi-Yong Liu, Email: liuqiyong@icdc.cn.
Yu-Hong Guo, Email: guoyuhong@icdc.cn.
Jing-Yi Jiang, Email: HNYCCDC@163.com.
Dong-Sheng Ren, Email: rendongsheng@icdc.cn.
Guang-Chao Zhou, Email: yczgc@126.com.
Can-Jun Zheng, Email: zcj007007@hotmail.com.
Yan Zhang, Email: Jessica.yanzhang@gmail.com.
Jing-Li Liu, Email: liujingli9931@163.com.
Zhi-Fang Li, Email: lzf080122@126.com.
Yun Chen, Email: chenyun0727@126.com.
Hong-Sheng Li, Email: lihongshengyccdc@163.com.
Lindsay C Morton, Email: lmorton@health.usf.edu.
Hua-Zhong Li, Email: lhzcdc@126.com.
Qun Li, Email: fyklq@vip.sina.com.
Wei-Dong Gu, Email: weidonggu@cdc.gov.
Acknowledgements
This work is financially supported by the Major National Science and Technology Projects of China awarded #2008ZX10004-010 and the National Basic Research Program of China (973 Program) awarded #2012CB955500. The funder had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript, and the content is solely the responsibility of the authors and does not necessarily represent the official views of the Major National Science and Technology Projects of China and the National Basic Research Program of China (973 Program). We sincerely thank the anonymous referees and Prof. Chris Arme for their series of excellent comments and suggestions. We sincerely thank Yu-Jing Liu, Ph.D. from Northwest A&F University for her enthusiastic offering to help embellish the language of the revised version. We also sincerely thank Robert J. Novak, Ph.D. from University of South Florida who gave us guidance in the field in Yongcheng city. The authors would like to thank Wei-Zhong Yang from China CDC who helped in the guidance of the experiment. The authors would like to thank Feng-Xia Meng, Liang Lu, Hai-Xia Wu, Gui-Chang Li, Dong-Mei Li, Jun Wang and Xiu-Ping Song at Department of Vector Biology and Control, ICDC, China CDC who helped in conducting of the experiments. The authors would like to thank Zhen-Rong Cui, Yang Shen, Yun-Jun Su, Ming-Jie Zhu, Hai-Shan Wang and Guang-Xu Zhou from Yongcheng CDC who participated in mosquito collections, Ke-Hua Peng, Qing-Lin Zhu, and Heng-Chao Xie from township hospital who helped in organizing the experiments and capturing mosquitoes at night. And lastly, we are again grateful to the village doctors of the research sites who participated completely in this project.
References
- Guerra CA, Snow RW, Hay SI. Mapping the global extent of malaria in 2005. Trends Parasitol. 2006;22(8):353–358. doi: 10.1016/j.pt.2006.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guerra CA, Snow RW, Hay SI. Defining the global spatial limits of malaria transmission in 2005. Adv Parasitol. 2006;62:157–179. doi: 10.1016/S0065-308X(05)62005-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Price RN, Tjitra E, Guerra CA, Yeung S, White NJ, Anstey NM. Vivax malaria: neglected and not benign. Am J Trop Med Hyg. 2007;77(6 Suppl):79–87. [PMC free article] [PubMed] [Google Scholar]
- Alonso PL, Brown G, Arevalo-Herrera M, Binka F, Chitnis C, Collins F, Doumbo OK, Greenwood B, Hall BF, Levine MM, A research agenda to underpin malaria eradication. PLoS Med. p. e1000406. [DOI] [PMC free article] [PubMed]
- Zhou SS, Huang F, Wang JJ, Zhang SS, Su YP, Tang LH. Geographical, meteorological and vectorial factors related to malaria re-emergence in Huang-Huai River of central China. Malar J. 2010;9:337. doi: 10.1186/1475-2875-9-337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tang LH. Progress in malaria control in China. Chin Med J (Engl) 2000;113(1):89–92. [PubMed] [Google Scholar]
- Zhou ZJ. The malaria situation in the People's Republic of China. Bull World Health Organ. 1981;59(6):931–936. [PMC free article] [PubMed] [Google Scholar]
- Lin H, Lu L, Tian L, Zhou S, Wu H, Bi Y, Ho SC, Liu Q. Spatial and temporal distribution of falciparum malaria in China. Malar J. 2009;8:130. doi: 10.1186/1475-2875-8-130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sleigh AC, Liu XL, Jackson S, Li P, Shang LY. Resurgence of vivax malaria in Henan Province, China. Bull World Health Organ. 1998;76(3):265–270. [PMC free article] [PubMed] [Google Scholar]
- Xu BL, Su YP, Shang LY, Zhang HW. Malaria control in Henan Province, People's Republic of China. Am J Trop Med Hyg. 2006;74(4):564–567. [PubMed] [Google Scholar]
- Liu XZ, Xu BL. Malaria situation and evaluation on the control effect in Henan Province during 1990-2005. Chinese Journal of Parasitology and Parasitic Diseases. 2006;24(3):226–229. [PubMed] [Google Scholar]
- Zizhao L, Luoyuan S, Lian Z, Dongfang L, Yunpu S. Control strategies of malaria in Henan Province, China. Southeast Asian J Trop Med Public Health. 1999;30(2):240–242. [PubMed] [Google Scholar]
- Liu X, Jackson S, Song J, Sleigh AC. Malaria control and fever management in Henan Province, China, 1992. Trop Med Int Health. 1996;1(1):112–116. doi: 10.1046/j.1365-3156.1996.d01-5.x. [DOI] [PubMed] [Google Scholar]
- Wang LP, Fang LQ, Xu X, Wang JJ, Ma JQ, Cao WC, Jin SG. Study on the determinants regarding malaria epidemics in Anhui province during 2004-2006. Chinese Journal of Epidemiology. 2009;30(1):38–41. (in Chinese) [PubMed] [Google Scholar]
- Zhang W, Wang L, Fang L, Ma J, Xu Y, Jiang J, Hui F, Wang J, Liang S, Yang H. et al. Spatial analysis of malaria in Anhui province, China. Malar J. 2008;7:206. doi: 10.1186/1475-2875-7-206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lines J Whitty CJ Hanson K Prospects for eradication and elimination of malaria: A technical briefing for DFID London School of Hygiene & Tropical Medicine 2007. 22118883
- Tanner M, de Savigny D. Malaria eradication back on the table. Bull World Health Organ. 2008;86(2):82. doi: 10.2471/BLT.07.050633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu QY, Liu XB. Prevention and control of vector Anopheles: a key approach for malaria elimination in China. Chin J Vector Biol & Control. 2010;21(5):409–413. (in Chinese) [Google Scholar]
- Qu FY. Historical review on the classification and rectification of Anopheles anthropophagus to An. lesteri in China. Chinese Journal of Parasitology and Parasitic Diseases. 2008;26(3):234–235. [PubMed] [Google Scholar]
- Gu ZC, Shang LY, Chen JS, Zheng X, Su YJ, Li AM, Liu H, Luo MZ, Qian HL, Tang LH. The role of Anopheles anthropophagus in malaria transmission in in Xinyang City of Henan Province. Chinese Journal of Parasitology and Parasitic Diseases. 2001;19(4):221–224. [PubMed] [Google Scholar]
- Chow CY. Malaria vectors in China. Chinese Journal of Entomology. 1991;Special Publ(6):67–79. (in Chinese) [Google Scholar]
- Cox FE. History of the discovery of the malaria parasites and their vectors. Parasit Vectors. 2010;3(1):5. doi: 10.1186/1756-3305-3-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gou GX, Li DF, Shang LY, Guo XS, Wang WX, Sui QL, Shen YD, Hao JL, Hu ZT, Liang DP. et al. The study on ecological habits of Anopheles sinensis in Guantang, Luyi county from 1971 to 1996. Chin J Vector Biol & Control. 1998;9:133–134. (in Chinese) [Google Scholar]
- Qu CZ, Su TZ. Vectorial capacity of malaria transmission of Anopheles sinensis in Zhengzhou in nature. Joumal of Henan Medieal University. 2000;35:394–396. (in Chinese) [Google Scholar]
- Zhang HW, Su YP, Zhou GC. et al. Re-emerging malaria in Yongcheng city of Henan province. Chin J Vector Biol & Control. 2007;18(1):42–44. (in Chinese) [Google Scholar]
- Zhou GC, Zhang HW, Su YP. et al. Evaluation of therapeutic measures of radical treatment for malaria in pre-transmission season in Yongcheng, Henan, China in 2007. Journal of Pathogen Biology. 2009;4(2):112–114. (in Chinese) [Google Scholar]
- Zhou GC, Zhang HW, Su YP. et al. Investigation into the results of control of malaria outbreak in Yongcheng County by biological control of mosquito larvae. China Tropical Medicine. 2009;9(2):228–229+259. (in Chinese) [Google Scholar]
- Hu YX, Miao YG, Fan TB. et al. The further study on ecological habits of Anopheles sinensis in the area along Huang River and Huai River. Chin J Parasit Parasitic Dis. 1988;S1:135. (in Chinese) [Google Scholar]
- Dia I, Ba H, Mohamed SA, Diallo D, Lo B, Diallo M. Distribution, host preference and infection rates of malaria vectors in Mauritania. Parasit Vectors. 2009;2(1):61. doi: 10.1186/1756-3305-2-61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jonathan FD. Host-seeking strategies of mosquito disease vectors. J Am Mosq Control Assoc. 2005;21(sp1):17–22. doi: 10.2987/8756-971X(2005)21[17:HSOMDV]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- Bowen MF. The sensory physiology of host-seeking behavior in mosquitoes. Annu Rev Entomol. 1991;36:139–158. doi: 10.1146/annurev.en.36.010191.001035. [DOI] [PubMed] [Google Scholar]
- Reisen WK, Lothrop HD, Meyer RP. Time of host-seeking by Culex tarsalis (Diptera: Culicidae) in California. J Med Entomol. 1997;34(4):430–437. doi: 10.1093/jmedent/34.4.430. (438) [DOI] [PubMed] [Google Scholar]
- Laarman JJ. The host-seeking behaviour of anopheline mosquitoes. Trop Geogr Med. 1958;10(4):293–305. [PubMed] [Google Scholar]
- Lardeux F, Loayza P, Bouchite B, Chavez T. Host choice and human blood index of Anopheles pseudopunctipennis in a village of the Andean valleys of Bolivia. Malar J. 2007;6:8. doi: 10.1186/1475-2875-6-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmidtmann ET, Jones CJ, Gollands B. et al. Comparative host-seeking activity of culicdides(Diptera:Ceratopogonidae) attracted to pastured livestock in central New York state, USA. J Med Entomol. 1980;17(3):221–231. (211) [Google Scholar]
- Masuoka P, Klein TA, Kim HC, Claborn DM, Achee N, Andre R, Chamberlin J, Small J, Anyamba A, Lee DK. et al. Modeling the distribution of Culex tritaeniorhynchus to predict Japanese encephalitis distribution in the Republic of Korea. Geospat Health. 2010;5(1):45–57. doi: 10.4081/gh.2010.186. [DOI] [PubMed] [Google Scholar]
- Rosen L, Shroyer DA, Lien JC. Transovarial transmission of Japanese encephalitis virus by Culex tritaeniorhynchus mosquitoes. Am J Trop Med Hyg. 1980;29(4):711–712. doi: 10.4269/ajtmh.1980.29.711. [DOI] [PubMed] [Google Scholar]
- Rwegoshora RT, Sharpe RG, Baisley KJ, Kittayapong P. Biting behavior and seasonal variation in the abundance of Anopheles minimus species A and C in Thailand. Southeast Asian J Trop Med Public Health. 2002;33(4):694–701. [PubMed] [Google Scholar]
- Djenontin A, Bio-Bangana S, Moiroux N, Henry MC, Bousari O, Chabi J, Osse R, Koudenoukpo S, Corbel V, Akogbeto M. et al. Culicidae diversity, malaria transmission and insecticide resistance alleles in malaria vectors in Ouidah-Kpomasse-Tori district from Benin (West Africa): A pre-intervention study. Parasit Vectors. 2010;3:83. doi: 10.1186/1756-3305-3-83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okumu FO, Madumla EP, John AN, Lwetoijera DW, Sumaye RD. Attracting, trapping and killing disease-transmitting mosquitoes using odor-baited stations - The Ifakara Odor-Baited Stations. Parasit Vectors. 2010;3:12. doi: 10.1186/1756-3305-3-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Owino EA. Trapping mosquitoes using milk products as odour baits in western Kenya. Parasit Vectors. 2010;3:55. doi: 10.1186/1756-3305-3-55. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Reddy MR, Overgaard HJ, Abaga S, Reddy VP, Caccone A, Kiszewski AE, Slotman MA. Outdoor host seeking behaviour of Anopheles gambiae mosquitoes following initiation of malaria vector control on Bioko Island, Equatorial Guinea. Malar J. 2011;10(1):184. doi: 10.1186/1475-2875-10-184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forattini OP, Kakitani I, Massad E, Marucci D. Studies on mosquitoes (Diptera: Culicidae) and anthropic environment. 11--Biting activity and blood-seeking parity of Anopheles (Kerteszia) in south-eastern Brazil. Rev Saude Publica. 1996;30(2):107–114. doi: 10.1590/S0034-89101996000200001. [DOI] [PubMed] [Google Scholar]
- Kweka EJ, Mwang'onde BJ, Mahande AM. Optimization of odour-baited resting boxes for sampling malaria vector, Anopheles arabiensis Patton, in arid and highland areas of Africa. Parasit Vectors. 2010;3:75. doi: 10.1186/1756-3305-3-75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balenghien T, Fouque F, Sabatier P, Bicout DJ. Horse-, bird-, and human-seeking behavior and seasonal abundance of mosquitoes in a West Nile Virus focus of southern France. J Med Entomol. 2006;43(5):936–946. doi: 10.1603/0022-2585(2006)43[936:HBAHBA]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- Burkett DA, Lee WJ, Lee KW. et al. Light, carbon dioxide, and octenol-baited mosquito trap and host-seeking activity evaluations for mosquitoes in a malarious area of the Republic of Korea. J Am Mosq Control Assoc. 2001;17(3):196–205. [PubMed] [Google Scholar]
- Kent RJ. Molecular methods for arthropod bloodmeal identification and applications to ecological and vector-borne disease studies. Mol Ecol Resour. 2009;9(1):4–18. doi: 10.1111/j.1755-0998.2008.02469.x. [DOI] [PubMed] [Google Scholar]
- Beier JC, Perkins PV, Wirtz RA, Koros J, Diggs D, Gargan TP, Koech DK. Bloodmeal identification by direct enzyme-linked immunosorbent assay (ELISA), tested on Anopheles (Diptera: Culicidae) in Kenya. J Med Entomol. 1988;25(1):9–16. doi: 10.1093/jmedent/25.1.9. [DOI] [PubMed] [Google Scholar]
- Washino RK, Tempelis CH. Mosquito host bloodmeal identification: methodology and data analysis. Annu Rev Entomol. 1983;28:179–201. doi: 10.1146/annurev.en.28.010183.001143. [DOI] [PubMed] [Google Scholar]
- Lombardi S, Esposito F. Enzyme-linked immunosorbent assay (ELISA) for the identification of mosquito bloodmeals. Parassitologia. 1983;25(1):49–56. [PubMed] [Google Scholar]
- Tempelis CH, Lofy MF. A modified precipitin method for identification of mosquito bloodmeals. Am J Trop Med Hyg. 1963;12:825–831. doi: 10.4269/ajtmh.1963.12.825. [DOI] [PubMed] [Google Scholar]
- Kent RJ, Norris DE. Identification of mammalian blood meals in mosquitoes by a multiplexed polymerase chain reaction targeting cytochrome B. Am J Trop Med Hyg. 2005;73(2):336–342. [PMC free article] [PubMed] [Google Scholar]
- Hatefi Y. The mitochondrial electron transport and oxidative phosphorylation system. Annu Rev Biochem. 1985;54:1015–1069. doi: 10.1146/annurev.bi.54.070185.005055. [DOI] [PubMed] [Google Scholar]
- Ngo KA, Kramer LD. Identification of mosquito bloodmeals using polymerase chain reaction (PCR) with order-specific primers. J Med Entomol. 2003;40(2):215–222. doi: 10.1603/0022-2585-40.2.215. [DOI] [PubMed] [Google Scholar]
- Townzen JS, Brower AV, Judd DD. Identification of mosquito bloodmeals using mitochondrial cytochrome oxidase subunit I and cytochrome b gene sequences. Med Vet Entomol. 2008;22(4):386–393. doi: 10.1111/j.1365-2915.2008.00760.x. [DOI] [PubMed] [Google Scholar]
- Boakye DA, Tang J, Truc P, Merriweather A, Unnasch TR. Identification of bloodmeals in haematophagous Diptera by cytochrome B heteroduplex analysis. Med Vet Entomol. 1999;13(3):282–287. doi: 10.1046/j.1365-2915.1999.00193.x. [DOI] [PubMed] [Google Scholar]
- Lee JH, Hassan H, Hill G, Cupp EW, Higazi TB, Mitchell CJ, Godsey MS, Unnasch TR Jr. Identification of mosquito avian-derived blood meals by polymerase chain reaction-heteroduplex analysis. Am J Trop Med Hyg. 2002;66(5):599–604. doi: 10.4269/ajtmh.2002.66.599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fall AG, Diaite A, Lancelot R, Tran A, Soti V, Etter E, Konate L, Faye O, Bouyer J. Feeding behaviour of potential vectors of West Nile virus in Senegal. Parasit Vectors. 2011;4:99. doi: 10.1186/1756-3305-4-99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou SS, Wang Y, Fang W, Tang LH. Malaria situation in the People's Republic of China in 2008. Chin J Parasit Parasitic Dis. 2009;27(6):455–457. (in Chinese) [PubMed] [Google Scholar]
- Diatta M, Spiegel A, Lochouarn L, Fontenille D. Similar feeding preferences of Anopheles gambiae and A. arabiensis in Senegal. Trans R Soc Trop Med Hyg. 1998;92(3):270–272. doi: 10.1016/S0035-9203(98)91005-7. [DOI] [PubMed] [Google Scholar]
- Burkett DA, Lee WJ, Lee KW, Kim HC, Lee HI, Lee JS, Shin EH, Wirtz RA, Cho HW, Claborn DM. et al. Late season commercial mosquito trap and host seeking activity evaluation against mosquitoes in a malarious area of the Republic of Korea. Korean J Parasitol. 2002;40(1):45–54. doi: 10.3347/kjp.2002.40.1.45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Braack LE, Coetzee M, Hunt RH, Biggs H, Cornel A, Gericke A. Biting pattern and host-seeking behavior of Anopheles arabiensis (Diptera: Culicidae) in northeastern South Africa. J Med Entomol. 1994;31(3):333–339. doi: 10.1093/jmedent/31.3.333. [DOI] [PubMed] [Google Scholar]
- Miller JE, Gibson G. Behavioral response of host-seeking mosquitoes (Diptera: Culicidae) to insecticide-impregnated bed netting: a new approach to insecticide bioassays. J Med Entomol. 1994;31(1):114–122. doi: 10.1093/jmedent/31.1.114. [DOI] [PubMed] [Google Scholar]
- Muenworn V, Sungvornyothin S, Kongmee M, Polsomboon S, Bangs MJ, Akrathanakul P, Tanasinchayakul S, Prabaripai A, Chareonviriyaphap T. Biting activity and host preference of the malaria vectors Anopheles maculatus and Anopheles sawadwongporni (Diptera: Culicidae) in Thailand. J Vector Ecol. 2009;34(1):62–69. doi: 10.1111/j.1948-7134.2009.00008.x. [DOI] [PubMed] [Google Scholar]
- Lu BL, Li PS, Ji SH, Fauna sinica insecta(Diptera: Culicidae) Beijing, China: Science Press; 1997. (in Chinese) [Google Scholar]
- Lee WJ, Klein TA, Kim HC, Choi YM, Yoon SH, Chang KS, Chong ST, Lee IY, Jones JW, Jacobs JS. et al. Anopheles kleini, Anopheles pullus, and Anopheles sinensis: potential vectors of Plasmodium vivax in the Republic of Korea. J Med Entomol. 2007;44(6):1086–1090. doi: 10.1603/0022-2585(2007)44[1086:AKAPAA]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- Collins FH, Paskewitz SM. A review of the use of ribosomal DNA (rDNA) to differentiate among cryptic Anopheles species. Insect Molecular Biology. 1996;5(1):1–9. doi: 10.1111/j.1365-2583.1996.tb00034.x. [DOI] [PubMed] [Google Scholar]
- Ma YJ, Qu FY, Xu JN, Zheng ZM. Differentiation of Anopheles sinensis and Anopheles anthropophagus using a ribosomal DNA PCR assay. Acad J Sec Mil Med Univ. 1998;19(3):237–239. (in Chinese) [Google Scholar]
- Porter CH, Collins FH. Species-diagnostic differences in a ribosomal DNA internal transcribed spacer from the sibling species Anopheles freeborni and Anopheles hermsi (Diptera:Culicidae) Am J Trop Med Hyg. 1991;45(2):271–279. doi: 10.4269/ajtmh.1991.45.271. [DOI] [PubMed] [Google Scholar]
- Rutledge CR, Cornel AJ, Meek CL, Collins FH. Validation of a ribosomal DNA-polymerase chain reaction species diagnostic assay for the common malaria mosquito (Diptera:Culicidae) sibling species complex. J Med Entomol. 1996;33(6):952–954. doi: 10.1093/jmedent/33.6.952. [DOI] [PubMed] [Google Scholar]
- Kengne P, Antonio-Nkondjio C, Awono-Ambene HP, Simard F, Awolola TS, Fontenille D. Molecular differentiation of three closely related members of the mosquito species complex, Anopheles moucheti, by mitochondrial and ribosomal DNA polymorphism. Med Vet Entomol. 2007;21(2):177–182. doi: 10.1111/j.1365-2915.2007.00681.x. [DOI] [PubMed] [Google Scholar]
- Ho C, Chou TC, Ch'En TH, Hsueh AT. The Anopheles hyrcanus group and its relation to malaria in east China. Chin Med J. 1962;81:71–78. [PubMed] [Google Scholar]
- Rueda LM, Pecor JE, Harrison BA. Updated distribution records for Anopheles vagus (Diptera: Culicidae) in the Republic of Philippines, and considerations regarding its secondary vector roles in Southeast Asia. Trop Biomed. 2011;28(1):181–187. [PubMed] [Google Scholar]
- Lillie TH, Kline DL, Hall DW. Host-seeking activity of Culicoides spp. (Diptera: Ceratopogonidae) near Yankeetown, Florida. J Am Mosq Control Assoc. 1988;4(4):485–493. [PubMed] [Google Scholar]
- Laarman JJ. The host-seeking behaviour of the malaria mosquito Anopheles maculipennis atroparvus. Acta Leiden. 1955;25:1–144. [PubMed] [Google Scholar]
- Ghosh KK, Das S, Chakraborty S, Bhattacharya S, Hati AK. Host preference of Anopheles annularis in different biotopes. Indian J Malariol. 1986;23(2):119–121. [PubMed] [Google Scholar]
- Suom C, Ginsberg HS, Bernick A, Klein C, Buckley PA, Salvatore C, Lebrun RA. Host-seeking activity and avian host preferences of mosquitoes associated with West Nile virus transmission in the northeastern U.S.A. J Vector Ecol. 2010;35(1):69–74. doi: 10.1111/j.1948-7134.2010.00060.x. [DOI] [PubMed] [Google Scholar]
- Grieco JP, Achee NL, Andre RG, Roberts DR. Host feeding preferences of Anopheles species collected by manual aspiration, mechanical aspiration, and from a vehicle-mounted trap in the Toledo District, Belize, Central America. J Am Mosq Control Assoc. 2002;18(4):307–315. [PubMed] [Google Scholar]
- Savage HM, Anderson M, Gordon E, McMillen L, Colton L, Delorey M, Sutherland G, Aspen S, Charnetzky D, Burkhalter K. et al. Host-seeking heights, host-seeking activity patterns, and West Nile virus infection rates for members of the Culex pipiens complex at different habitat types within the hybrid zone, Shelby County, TN, 2002 (Diptera: Culicidae) J Med Entomol. 2008;45(2):276–288. doi: 10.1603/0022-2585(2008)45[276:HHHAPA]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- Loftin KM, Byford RL, Loftin MJ, Craig ME, Steiner RL. Host preference of mosquitoes in Bernalillo County, New Mexico. J Am Mosq Control Assoc. 1997;13(1):71–75. [PubMed] [Google Scholar]
- Costantini C, Sagnon NF, della Torre A, Diallo M, Brady J, Gibson G, Coluzzi M. Odor-mediated host preferences of West African mosquitoes, with particular reference to malaria vectors. Am J Trop Med Hyg. 1998;58(1):56–63. doi: 10.4269/ajtmh.1998.58.56. [DOI] [PubMed] [Google Scholar]
- Mwandawiro C, Boots M, Tuno N, Suwonkerd W, Tsuda Y, Takagi M. Heterogeneity in the host preference of Japanese encephalitis vectors in Chiang Mai, northern Thailand. Trans R Soc Trop Med Hyg. 2000;94(3):238–242. doi: 10.1016/S0035-9203(00)90303-1. [DOI] [PubMed] [Google Scholar]
- Harrison BA, Scanlon JE. The subgenus Anopheles in Thailand. Contrib Am Entomol Inst. 1975;12:1–307. [Google Scholar]
- Takken W. The role of olfaction in host-seeking of mosquitoes:a review. Insect Sci Appl. 1991;12:287–295. [Google Scholar]
- Dekker T, Takken W, Braks MA. Innate preference for host-odor blends modulates degree of anthropophagy of Anopheles gambiae sensu lato (Diptera: Culicidae) J Med Entomol. 2001;38(6):868–871. doi: 10.1603/0022-2585-38.6.868. [DOI] [PubMed] [Google Scholar]
- Pureswaran DS, Poland TM. Host selection and feeding preference of Agrilus planipennis (Coleoptera: Buprestidae) on ash (Fraxinus spp.) Environ Entomol. 2009;38(3):757–765. doi: 10.1603/022.038.0328. [DOI] [PubMed] [Google Scholar]
- Costantini C, Gibson G, Brady J, Merzagora L, Coluzzi M. A new odour-baited trap to collect host-seeking mosquitoes. Parassitologia. 1993;35(1-3):5–9. [PubMed] [Google Scholar]
- Okumu FO, Govella NJ, Moore SJ, Chitnis N, Killeen GF. Potential benefits, limitations and target product-profiles of odor-baited mosquito traps for malaria control in Africa. PLoS One. 2010;5(7):e11573. doi: 10.1371/journal.pone.0011573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sasa M. Two years' observation on the seasonal activities and zoophilism of mosquitoes in Tokyo by animal trap method. Jap J Exp Med. 1951;20:509–517. [Google Scholar]
- Ree HI, Hwang UW, Lee IY, Kim TU. Daily survival and human blood index of Anopheles sinensis, the vector species of malaria in Korea. J Am Mosq Control Assoc. 2001;17:67–72. [PubMed] [Google Scholar]
- Ree HI. Studies on Anopheles sinensis, the vector species of vivax malaria in Korea. Korean J Parasitol. 2005;43(3):75–92. doi: 10.3347/kjp.2005.43.3.75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boreham PF, Lenahan JK, Boulzaguet R, Storey J, Ashkar TS, Nambiar R, Matsushima T. Studies on multiple feeding by Anopheles gambiae s.l. in a Sudan savanna area of north Nigeria. Trans R Soc Trop Med Hyg. 1979;73(4):418–423. doi: 10.1016/0035-9203(79)90167-6. [DOI] [PubMed] [Google Scholar]
- Scott TW, Chow E, Strickman D, Kittayapong P, Wirtz RA, Lorenz LH, Edman JD. Blood-feeding patterns of Aedes aegypti (Diptera: Culicidae) collected in a rural Thai village. J Med Entomol. 1993;30(5):922–927. doi: 10.1093/jmedent/30.5.922. [DOI] [PubMed] [Google Scholar]
- Klowden MJ, Briegel H. Mosquito gonotrophic cycle and multiple feeding potential: contrasts between Anopheles and Aedes (Diptera: Culicidae) J Med Entomol. 1994;31:618–622. doi: 10.1093/jmedent/31.4.618. [DOI] [PubMed] [Google Scholar]
- Wekesa JW, Yuval B, Washino RK. Multiple blood feeding in Anopheles freeborni (Diptera: Culicidae) Am J Trop Med Hyg. 1995;52:508–511. doi: 10.4269/ajtmh.1995.52.508. [DOI] [PubMed] [Google Scholar]
- Anderson RA, Brust RA. Field evidence for multiple host contacts during blood feeding by Culex tarsalis, Cx. restuans, and Cx. nigripalpus (Diptera: Culicidae) J Med Entomol. 1995;32(5):705–710. doi: 10.1093/jmedent/32.5.705. [DOI] [PubMed] [Google Scholar]
- Koella JC, Sorensen FL, Anderson RA. The malaria parasite, Plasmodium falciparum, increases the frequency of multiple feeding of its mosquito vector, Anopheles gambiae. Proc R Soc Lond B Biol Sci. 1998;265:763–768. doi: 10.1098/rspb.1998.0358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amerasinghe PH, Amerasinghe FP. Multiple host feeding in field populations of Anopheles culicifacies and An. subpictus in Sri Lanka. Med Vet Entomol. 1999;13(2):124–131. doi: 10.1046/j.1365-2915.1999.00160.x. [DOI] [PubMed] [Google Scholar]
- Scott TW, Amerasinghe PH, Morrison AC, Lorenz LH, Clark GG, Strickman D, Kittayapong P, Edman JD. Longitudinal studies of Aedes aegypti (Diptera: Culicidae) in Thailand and Puerto Rico: blood feeding frequency. J Med Entomol. 2000;37:89–101. doi: 10.1603/0022-2585-37.1.89. [DOI] [PubMed] [Google Scholar]
- Chow CY. Note on the time of feeding of Anopheles hyrcanus sinensis and Anopheles mininus in the vicinity of Chungking. Chin Med J. 1949;67:489–490. (in Chinese) [Google Scholar]
- Fredeen FJ, Mason PG. Meteorological factors influencing host-seeking activity of female Simulium luggeri (Diptera: Simuliidae) J Med Entomol. 1991;28(6):831–840. doi: 10.1093/jmedent/28.6.831. [DOI] [PubMed] [Google Scholar]
- Huang F, Zhou S, Zhang S, Wang H, Tang L. Temporal correlation analysis between malaria and meteorological factors in Motuo County, Tibet. Malar J. 2011;10:54. doi: 10.1186/1475-2875-10-54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bassiouny HK. Bioenvironmental and meteorological factors related to the persistence of malaria in Fayoum Governorate: a retrospective study. East Mediterr Health J. 2001;7(6):895–906. [PubMed] [Google Scholar]
- Ruiz MO, Chaves LF, Hamer GL, Sun T, Brown WM, Walker ED, Haramis L, Goldberg TL, Kitron UD. Local impact of temperature and precipitation on West Nile virus infection in Culex species mosquitoes in northeast Illinois, USA. Parasit Vectors. 2010;3(1):19. doi: 10.1186/1756-3305-3-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alemu A, Abebe G, Tsegaye W, Golassa L. Climatic variables and malaria transmission dynamics in Jimma town, South West Ethiopia. Parasit Vectors. 2011;4:30. doi: 10.1186/1756-3305-4-30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang GJ, Gao Q, Zhou SS, Malone JB, McCarroll JC, Tanner M, Vounatsou P, Bergquist R, Utzinger J, Zhou XN. Mapping and predicting malaria transmission in the People's Republic of China, using integrated biology-driven and statistical models. Geospat Health. 2010;5(1):11–22. doi: 10.4081/gh.2010.183. [DOI] [PubMed] [Google Scholar]
- Paaijmans KP, Read AF, Thomas MB. Understanding the link between malaria risk and climate. PNAS. 2009;106(33):13844–13849. doi: 10.1073/pnas.0903423106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arunachalam N, Samuel PP, Hiriyan J, Rajendran R, Dash AP. Short report: observations on the multiple feeding behavior of Culex tritaeniorhynchus (Diptera: culicidae), the vector of Japanese encephalitis in Kerala in southern India. Am J Trop Med Hyg. 2005;72(2):198–200. [PubMed] [Google Scholar]