

Abstract
Almost all naturally occurring metalloproteases are monozinc enzymes. The zinc in any number of zinc metalloproteases has been substituted by some other divalent cation. Almost all Co(II)- or Mn(II)-substituted enzymes maintain the catalytic activity of their zinc counterparts. However, in the case of Cu(II) substitution of zinc proteases, a great number of enzymes are not active, for example, thermolysin, carboxypeptidase A, endopeptidase from Lactococcus lactis, or aminopeptidase B, while some do have catalytic activity, for example, astacin (37%) and DPP III (100%). Based on structural studies of various metal-substituted enzymes, for example, thermolysin, astacin, aminopeptidase B, dipeptidyl peptidase (DPP) III, and del-DPP III, the metal coordination geometries of both active and inactive Cu(II)-substituted enzymes are shown to be the same as those of the wild-type Zn(II) enzymes. Therefore, the enzyme activity of a copper-ion-substituted zinc metalloprotease may depend on the flexibility of catalytic domain.
1. Introduction
Proteolytic enzymes are recognized by their catalytic type, that is, aspartic, cysteine, metallo, serine, threonine, and others as yet unclassified. The largest number of proteolytic enzymes are classified as metalloproteases [1]. Almost all metalloproteases contain one or two zinc ions, and several enzymes contain one or two cobalt or manganese ions. The HExxH motif forming an α-helix is well conserved in many monozinc enzymes as the active site in which the two histidine residues coordinate with the zinc ion [2]. Some other monozinc proteases have different zinc-binding motifs, for example, HxxE(D)-aan-H in the carboxypeptidase family or HxD-aa12-H-aa12-H in the matrix metalloprotease family [2]. Dipeptidyl peptidase (DPP) III also has a unique zinc-binding motif, which was classified as family M49 in 1999 by MEROPS (peptidase database) after rat DPP III had been cloned and the HELLGH motif of DPP III was identified as an active site coordinated with a zinc ion [3, 4]. Although the motif HELLGH could not be found in any other metalloproteases, it exists in three kinds of monooxygenases (tyrosine, phenylalanine, and tryptophan hydroxylases) as an iron-binding site, as revealed by a search of the NBRF-PIR protein sequence database.
Zinc atoms in several zinc metalloproteases, for example, astacin [5], carboxypeptidase A [6], and thermolysin [7, 8], have been substituted by other divalent cations to probe the role of the metal for catalysis and structure. Some of these enzymes, for example, DPP III and astacin, were shown to have high metal substitution tolerance by metal substitution studies [9]. However, it is difficult to determine the relationship between the metal tolerance and the metal coordination structure of zinc metalloproteases.
Here, we show the metal coordination structure of the unique zinc-binding motif of DPP III, in which the zinc-binding motif is stabilized by several hydrogen bonds with acidic amino acid residues surrounding the zinc-binding motif, in order to clarify the relationship between the metal tolerance and the structure of the zinc-binding domain. The metal tolerances of both DPP III and del-DPP III, whose active site converts into a normal zinc-binding motif (HExxH), are shown here and compared with those reported for other metalloproteases [10]. Finally, we discuss the relationship between the catalytic activities and metal coordination structures of metal-substituted enzymes.
2. Identification of a Zinc-Binding Motif in DPP III
We start with the identification of the zinc-binding motif in DPP III, which will be used for further investigation of the relationship between the metal tolerance and the metal coordination structures of DPP III. The deduced amino acid sequences from cDNA for human, rat, and fruit fly DPP IIIs are 723–738 amino acids long and conserve the amino acid sequence HELLGH-aa52-E [3, 11, 12], which resembles the HExxH-aan-E zinc-binding motif conserved in many metalloproteases, such as thermolysin [13] and leukotriene A4 hydrolase [14]. Site-directed mutagenesis was performed on rat DPP III in order to testify that the HELLGH-aa52-E is a zinc-binding domain. Site-directed mutagenesis studies have clearly shown that the H450Y, H455Y, and E508A mutants, which lack zinc ions, lose their catalytic activity [4]. Replacement of Glu451 in these mutants with an alanine or an aspartic acid restores a mol of zinc ion per mol of protein but does not restore catalytic activity [4]. These results show that the H450ELLGH-aa52-E508 motif is a catalytic domain of which His450, His455, and Glu508 are ligands of a zinc ion and of which Glu451 is a catalytic amino acid residue, in the same way that the H142ExxH-aa19-E166 motif of thermolysin is a catalytic domain of which His142, His146, and Glu166 are ligands of a zinc ion and Glu143 is a catalytic amino acid residue.
The 1.95-Å crystal structure of yeast DPP III representing a prototype for the M49 family of metalloproteases was resolved by Baral et al. [15]. It shows a novel protein fold with two domains forming a wide cleft containing the catalytic metal ion. However, the three-dimensional structure of zinc coordination (His460, His465, and Glu517) and the catalytic active (Glu461) residues are structurally conserved, similar to those presented in many metalloproteases, such as thermolysin [13]. The HELLGH motif and the third ligand (Glu517) of DPP III construct a helix α14 and a helix α16, respectively [15]. The 3D structure of DPP III is similar to that of thermolysin [13] or leukotriene A4 hydrolase [16], the zinc-binding domain of which is constructed of two α-helixes, for HExxH (containing two zinc ligands) and xNEx (third ligand).
Figure 1 shows the superimposition of the active sites of rat DPP III and thermolysin. The helix α14 of DPP III has a slightly larger loop than that of thermolysin, and the glutamic acid on the motif comes close to zinc ion comparing with the glutamic acid on the normal helix of thermolysin [13, 17].
Figure 1.

The superimposition of the active sites of rat DPP III and thermolysin. Zinc ion is shown as a green sphere, and amino acid side chains are shown as sticks colored red for oxygen and blue for nitrogen. Metal coordinates in light blue and hydrogen bonds in yellow are indicated by dashed lines. Carbon atom and amino acid chain are shown colored white for thermolysin and magenta for DPP III. Metal coordination bonds are indicated by light blue dashed lines.
3. Stabilization of the Coordination between Ligands and Metal
In the 3D structural model of the zinc-binding domain of many zinc enzymes—neprilysin [18], thermolysin [13], carboxypeptidase A [19], leukotriene A4 hydrolase [16], aspzincin [20], and DPP III [17]—the His, His, and Glu residues that coordinate with the zinc ion are engaged in hydrogen bonds with one or two acidic amino acid residues (Glu or Asp) or other carbonyl oxygen atoms (Table 1). 3D structural models of catalytic domains of thermolysin (PDB: 1KEI), peptidyl-Lys metallopeptidase (PDB: 1GE6), and carboxypeptidase A (PDB: 1YME) are shown in Figure 2. In thermolysin (a), the oxygen atoms of Asp165 and Asp170 are engaged in hydrogen bonding with the nitrogen atoms of His146 and His142, respectively. Asp154 and Thr128 of peptidyl-Lys metalloendopeptidase (b) and Asp142 of carboxypeptidase A (c) are also engaged in hydrogen bonding with His117, His121, and His69, respectively. It was proved through the mutational studies of rat DPP III that this network of hydrogen bonds close to the zinc-binding motif plays an important role in stabilizing the coordination of the zinc ion to the protein [17]. The hydrogen bonds surrounding the zinc-binding motif of rat DPP III are shown in Figure 3, and the kinetic parameters, zinc contents and zinc dissociation constants of the several mutants are shown in Table 2. The replacement of Glu507 and Glu512, the oxygen atoms of which bind with the nitrogen atoms of His455 and His450, respectively, increases the dissociation constants by factors of 10~105 and correlatively reduces the zinc contents and enzyme activities. The hydrogen bonds between acidic amino acid residues and zinc ligands (His, His, and Glu) may stabilize the coordination of the zinc ion with the protein of the metalloprotease.
Table 1.
The zinc coordination residues and the residues that fix the coordination with hydrogen bonds.
| Zinc metalloprotease | Coordination residues | Residues that form the hydrogen bond with the coordination residues | PDB no. |
|---|---|---|---|
| (1) Thermolysin type | (HExxH- aan-E) | α-helix-aan-α-helix | |
| Thermolysin | His142, His146 Glu166 | Asp170-2.8 Å-His142
Asn165-2.8 Å-His146 |
1KEI |
| Vibriolysin | His345, His349 Glu369 | Asp373-2.8 Å-His345
Asn368-2.8 Å-His349 |
3NQX |
| Staphylococcus aureus metalloproteinase | His144, His148 Glu168 | Asn167-2.8 Å-His148
Asp172-2.8 Å-His144 |
1BQB |
| Zinc aminopeptidase | His265, His269 Glu288 | Phe272(C=O)-2.9 Å-His269 | 1Z1W |
| Leukotriene A4 hydrolase | His295, His299 Glu318 | Glu325-2.8 Å-His295
Gly303(C=O)-2.6 Å-His299 |
1SQM |
| Human thimet oligopeptidase | His473, His477 Glu502 | Glu509-2.6 Å-His473 | 1SQM |
| Human neutral endopeptidase (Neprilysin) | His583, His587 Glu646 | Asp650-2.9 Å-His583
Asp590-2.7 Å-His587 |
1DMT |
|
| |||
| (2) Endopeptidase type | (HExxH-aan-E or D) α-helix-aan-random coil | ||
| Peptidyl-Lys metalloendopeptidase | His117, His121 Asp130 | Asp154-2.7 Å-His117
Thr128(C=O)-2.8 Å-His121 |
1GE6 |
|
| |||
| (3) Carboxypeptidase A type | β-sheet-aan-random coil | ||
| Carboxypeptidase A | His69, His196 Glu72 | Asp142-2.7 Å-His69 | 1YME |
| Putative lysostaphin peptidase | His232, His311 Asp236 | Glu315-2.6 Å-His311
Gly216(C=O)-2.8 Å-His232 |
2GU1 |
Figure 2.
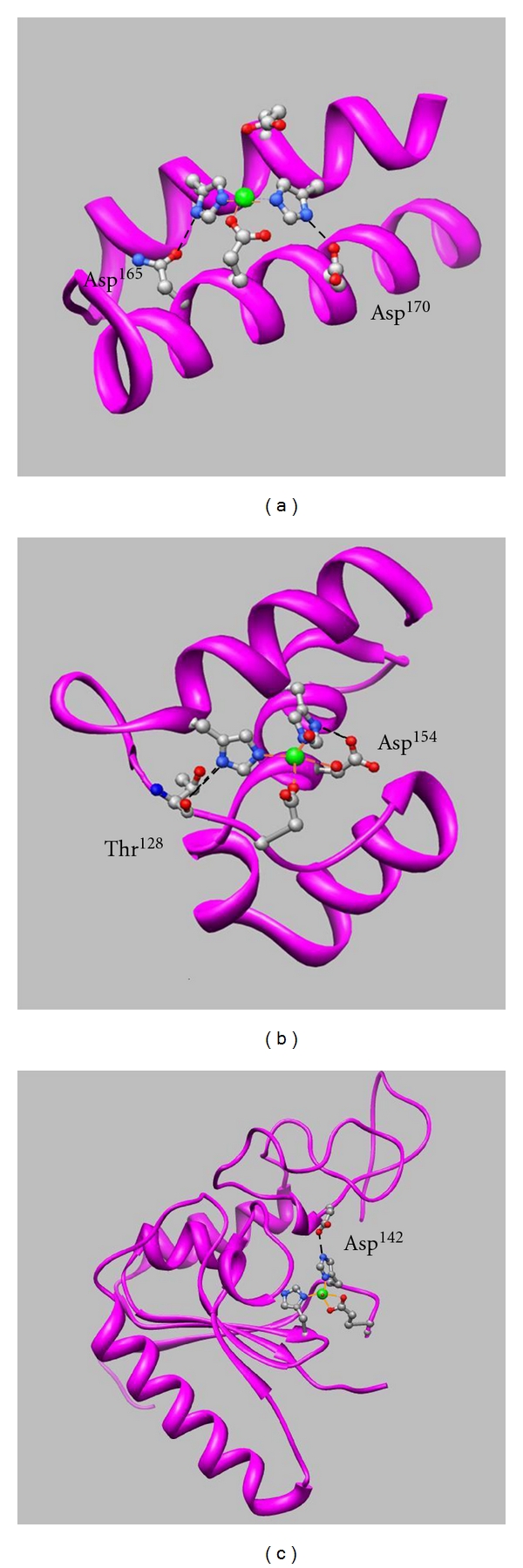
Three-dimensional structures of the catalytic domain models for thermolysin ((a): PDB 1KEI), peptidyl-Lys metallopeptidase ((b): PDB 1GE6), and carboxypeptidase A ((c): PDB 1YME). The zinc ion is shown as a green sphere, and amino acid side chains are shown as sticks colored red for oxygen, blue for nitrogen, and silver for carbon. Hydrogen bonds are indicated by dashed lines.
Figure 3.
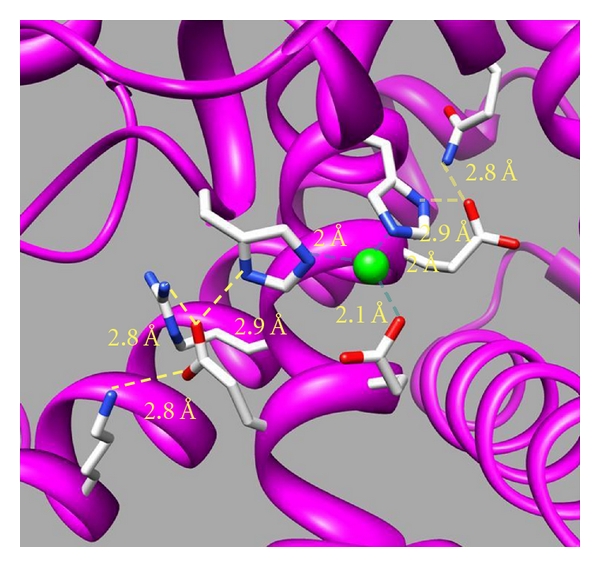
Molecular modeling of the catalytic site of rat DPP III. The model was generated as a template of the human DPP III crystal structure [39]. The zinc ion is shown as a green sphere, and amino acid side chains are shown as sticks colored red for oxygen, blue for nitrogen, and white for carbon. Metal coordinates in light blue and hydrogen bonds in yellow are indicated by dashed lines.
Table 2.
Kinetic parameters for the hydrolysis of Arg-Arg-NA, zinc contents, and zinc dissociation constants of wild-type and mutated rat DPP IIIs.
| Enzymes |
k
cat/K
M × 10−4
(M−1 s−1) |
Zinc content (mol/mol of proteina) | Zinc dissociation constant (M) (K d) |
|---|---|---|---|
| Wild-type | 73.6 ± 6.9 | 1.02 ± 0.15 | (4.5 ± 0.1) × 10−13 |
| E507D | 22.8 ± 1.9 | 0.65 ± 0.07 | (1.0 ± 0.2) × 10−11 |
| E507A | 4.43 ± 0.41 | 0.29 ± 0.04 | (1.0 ± 0.2) × 10−8 |
| E512D | 21.0 ± 0.19 | 0.45 ± 0.06 | (1.4 ± 0.1) × 10−12 |
| E512A | 2.45 ± 0.28 | 0.08 ± 0.01 | (2.6 ± 0.7) × 10−9 |
aValues are means ± SD of two separately prepared enzymes with duplicate determinations.
4. Metal Substitutions of Monozinc Metalloproteases
Almost all metalloproteases are monozinc enzymes. Some enzymes contain two zinc ions for catalytic domains, for example, human renal dipeptidase [36], and a few are dicobalt or dimanganese enzymes, for example, Pyrococcus furiosus methionine aminopeptidase [37] or Escherichia coli proline aminopeptidase [38], respectively.
The zinc in numerous zinc metalloproteases has been substituted by several divalent cations. The cobalt(II)- or manganese(II)-substituted enzymes showed nearly restored catalytic activity or even excess activity from apoenzyme, as seen in Table 3.
Table 3.
Reactivated Co(II) and Mn(II) enzymes substituted from apo-metalloproteases.
| Clan | Subclan | Name of enzyme | Replaced Ion | Reference |
|---|---|---|---|---|
| MA | E | Aminopeptidase Ey | Co2+, Mn2+ | Tanaka and Ichishima [21] |
| MA | E | Aminopeptidase B | Co2+ | Hirose et al. [22] |
| MA | E | Saccharolysin | Co2+, Mn2+ | Achstetter et al. [23] and Büchler et al. [24] |
| MA | E | Lysyl aminopeptidase | Co2+, Mn2+ | Klein et al. [25] |
| MA | E | Oligopeptidase F | Co2+, Mn2+ | Yan et al. [26] and Monnet et al. [27] |
| MA | E | Mycolysin (Thermolysin) | Co2+, Mn2+ Co2+ (200%), Mn2+ (10%) | Chang and Lee [28], and Holmquist and Vallee [29] |
| MA | E | Oligopeptidase O | Co2+
Mn2+ (50%) |
Tan et al. [30] and Baankreis et al. [31] |
| MA | E | Hyicolysin | Co2+ | Ayora and Götz [32] |
| ME | E | Legionella metalloendopeptidase | Mn2+ (69%) | Dreyfus and Iglewski [33] |
| MA | A | Epralysin | Co2+ (58%) | Diermayr et al. [34] |
| MA | M | Astacin | Co2+ (140%) | Gomis-Rüth et al. [5] |
| MA | M | MEPa (Gf b MEP) | Co2+
Mn2+ (200%) |
Nonaka et al. [35] |
| MA | M | Po c MEP | Co2+ (80%) Mn2+ (30%) |
Nonaka et al. [35] |
aPeptidyl-Lys metallopeptidase; b Grifola frondosa; c Pleurotus ostreatus.
Gomis-Rüth et al. [5] demonstrated in the metal substitution studies of astacin that Cu(II)-astacin displays enzyme activity of about 37%, while Ni(II)- and Hg(II)-astacin were almost inactive. In the crystal structure of Cu(II)-astacin, the metal ion is pentacoordinated with His92, His96, His102, Tyr149, and H2O, as in native Zn(II)-astacin or Co(II)-astacin; however, in the Ni(II)-astacin or Hg(II)-astacin, the metal ion is hexacoordinated with an additional solvent molecule or tetracoordinated with no ordered solvent molecule, respectively [5]. The restoration of catalytic activity in these substituted astacins was shown to be dependent on the metal coordination structure [5].
Meanwhile, almost all Cu(II)-substituted enzymes, such as thermolysin [7, 8], carboxypeptidase A [6], aminopeptidase B [22], or endopeptidase from Lactococcus lactis [30], show only partial activation or very low activities. The reason why these Cu(II) enzymes do not demonstrate catalytic activities may be that the coordination geometry of Cu(II) is more rigid than that of Zn(II) or Co(II).
In the case of DPP III, Co2+-, Ni2+- and Cu2+-DPP IIIs showed comparable catalytic activities to Zn2+-DPP III; the kinetic parameters are shown in Table 4 [9]. DPP III shows high flexibility of the metal ion for the catalytic activity compared with thermolysin or aminopeptidase B. Thermolysin or aminopeptidase B is a subclan MA (E) metalloprotease containing an HExxH-aan-E motif, and the 3D structure of the active domain is very similar to that of DPP III described above. The zinc ion in a subclan MA (E) metalloprotease or DPP III is tetracoordinated with three ligands (His, His, and Glu) and a water molecule. The metal-substituted (Co2+, Cu2+, or Ni2+) DPP III may have the same tetrahedral coordination structure as Zn2+-DPP III, so these enzymes are able to maintain the catalytic activity. The zinc in del-DPP III, whose active site converted into HExxH, was substituted with Co2+, Ni2+, or Cu2+to investigate the grounds for activation of the Cu2+-DPP III [10]. The Co2+-del-DPP III and Ni2+-del-DPP III showed comparable catalytic activity to that of Zn2+-del-DPP III, while the Cu2+-del-DPP III showed no catalytic activity, as in the case of thermolysin or aminopeptidase B [8–10].
Table 4.
Kinetic parameters for the hydrolysis of Arg-Arg-NA and metal contents of various metal-DPP IIIs.
| Enzyme |
K
M
(×10−5 M) |
k
cat
(s−1) |
k cat/K M(×104 M−1 s−1) | Metal content (mol/mol of protein) |
|---|---|---|---|---|
| Zn2+-DPP III | 8.1 (±1.0) |
7.1 (±0.2) |
8.8 | 0.8 (±0.1) |
| Co2+-DPP III | 8.2 (±0.9) |
7.0 (±0.1) |
8.5 | 1.0 (±0.1) |
| Cu2+-DPP III | 9.9 (±1.1) |
10.1 (±0.3) |
10.2 | 1.1 (±0.1) |
The EPR (electron paramagnetic resonance) parameters of various Cu2+-substituted metalloproteases are shown in Table 5. Each parameter is exactly alike between DPP III and thermolysin, aminopeptidase B, or del-DPP III [8–10, 22]. The results show that the Cu(II) coordination structures of the HExxH-aan-E and HExxxH-aa52-E motifs are very similar.
Table 5.
EPR parameters of Cu2+ proteases.
| g ⊥ | gll | All (×10−4 cm−1) | |
|---|---|---|---|
| Cu2+-DPP IIIa | 2.06 | 2.27 | 167 |
| Cu2+-del-DPP IIIb | 2.06 | 2.27 | 161 |
| Cu2+-thermolysinc | 2.06 | 2.26 | 163 |
| Cu2+-aminopeptidase Bd | 2.06 | 2.27 | 157 |
| Cu2+-carboxypeptidase Ae | 2.05 | 2.33 | 115 |
In the superimposition of the 3D structure models of active sites of DPP III and del-DPP III, the α-helix of DPP III, which is abnormally composed of 5 amino acid residues per one turn of the α-helix, is a larger loop than that of del-DPP III, the same as the case for the superimposition of the active sites of DPP III and thermolysin (Figure 1). The coordination geometries of the two enzymes are similar, while the position of Glu451, which is essential for the enzyme activity, is slightly closer to the copper ion in DPP III than in del-DPP III. The distances of oxygen atoms of the Glu451 residues of del-DPP III and wild-type DPP III are 4.8 Å and 3.2 Å from the zinc ion, respectively. Zn(II) coordination geometry is flexible, so both wild-type and del-DPP IIIs could have catalytic activity. However, the oxygen atom of Glu451 in Cu(II)-del-DPP III is not able to bind to the oxygen atom of the water molecule that is coordinated with the copper ion because the Cu(II) coordination geometry is very rigid. Therefore, the catalytic activity of Cu(II)-del-DPP III was diminished.
Some other Cu(II)-substituted enzymes, for example, aminopeptidase Ey [21], vibriolysin [40], hyicolysin [32], and Legionella metalloendopeptidase [33], were shown to have enzyme activities. These enzymes are all classified in subclan MA (E), the same as thermolysin or aminopeptidase B. The metal coordination structures of these enzymes have not been shown in detail; however, the catalytic domain may be more flexible than that of thermolysin or aminopeptidase B, in the same way as Cu(II)-substituted DPP III.
5. Conclusions
In this paper, we compared metal flexibility with the geometry of metal coordination of metalloproteases, to investigate why DPP III shows metal tolerance. Metal substitution of Zn(II) by Co(II) or Mn(II) on metalloproteases generally maintains catalytic activity, because the metal coordination geometries of Zn(II), Co(II), and Mn(II) are flexible. Most Cu(II)-substituted enzymes could not restore the catalytic activities, because the Cu(II) coordination geometry is very rigid. However, Cu(II)-substituted DPP III showed the same catalytic activity as that of Zn(II)-DPP III. We then studied the metal flexibilities and metal coordination geometries of many metallopeptidases, especially DPP III and del-DPP III, but we could not prove a relation between the metal flexibility and the metal coordination geometry. The metal tolerance of DPP III might depend on the flexibility of the metal-binding motif, not on the metal coordination geometry. By comparison of the 3D structure of active sites of DPP III and del-DPP III, both coordination geometries are seen to be similar, while the positions of catalytic amino acid residues (Glu) on those zinc-binding motifs are slightly different. We conclude that the catalytic site of Cu(II)-DPP III could be flexible enough to form the catalytic complex, with substrate and H2O.
References
- 1.Rawlings ND, Barret AJ. Introduction: metallopeptidases and their clans. In: Barrett AJ, Rawlings ND, Woessner JF, editors. Handbook of Proteolytic Enzyme. 2nd edition. San Diego, Calif, USA: Academic Press; 2004. pp. 231–268. [Google Scholar]
- 2.David A. Catalytic mechanisms for metallopeptidases. In: Barrett AJ, Rawlings ND, Woessner JF, editors. Handbook of Proteolytic Enzyme. 2nd edition. San Diego, Calif, USA: Academic Press; 1998. pp. 268–289. [Google Scholar]
- 3.Fukasawa K, Fukasawa KM, Kanai M, Fujii S, Hirose J, Harada M. Dipeptidyl peptidase III is a zinc metallo-exopeptidase. Molecular cloning and expression. Biochemical Journal. 1998;329(2):275–282. doi: 10.1042/bj3290275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Fukasawa K, Fukasawa KM, Iwamoto H, Hirose J, Harada M. The HELLGH motif of rat liver dipeptidyl peptidase III is involved in zinc coordination and the catalytic activity of the enzyme. Biochemistry. 1999;38(26):8299–8303. doi: 10.1021/bi9904959. [DOI] [PubMed] [Google Scholar]
- 5.Gomis-Rüth FX, Grams F, Yiallouros I, et al. Crystal structures, spectroscopic features, and catalytic properties of cobalt(II), copper(II), nickel(II), and mercury(II) derivatives of the zinc endopeptidase astacin. A correlation of structure and proteolytic activity. Journal of Biological Chemistry. 1994;269(25):17111–17117. [PubMed] [Google Scholar]
- 6.Rosenberg RC, Root CA, Bernstein PK, Gray HB. Spectral studies of copper(II) carboxypeptidase A and related model complexes. Journal of the American Chemical Society. 1975;97(8):2092–2096. doi: 10.1021/ja00841a016. [DOI] [PubMed] [Google Scholar]
- 7.Monzingo AF, Matthews BW. Binding of N-carboxymethyl dipeptide inhibitors to thermolysin determined by X-ray crystallography: a novel class of transition-state analogues for zinc peptidases. Biochemistry. 1984;23(24):5724–5729. doi: 10.1021/bi00319a010. [DOI] [PubMed] [Google Scholar]
- 8.Bertini I, Canti G, Kozlowski H, Scozzafava A. Spectroscopic characterization of copper(II) thermolysin. Journal of the Chemical Society Dalton Transactions. 1979;(8):1270–1273. [Google Scholar]
- 9.Hirose J, Iwamoto H, Nagao I, et al. Characterization of the metal-substituted dipeptidyl peptidase III (rat liver) Biochemistry. 2001;40(39):11860–11865. doi: 10.1021/bi0110903. [DOI] [PubMed] [Google Scholar]
- 10.Hirose J, Kamigakiuchi H, Iwamoto H, et al. The metal-binding motif of dipeptidyl peptidase III directly influences the enzyme activity in the copper derivative of dipeptidyl peptidase III. Archives of Biochemistry and Biophysics. 2004;431(1):1–8. doi: 10.1016/j.abb.2004.07.033. [DOI] [PubMed] [Google Scholar]
- 11.Mazzocco C, Fukasawa KM, Raymond AA, Puiroux J. Purification, partial sequencing and characterization of an insect membrane dipeptidyl aminopeptidase that degrades the insect neuropeptide proctolin. European Journal of Biochemistry. 2001;268(18):4940–4949. doi: 10.1046/j.1432-1327.2001.02425.x. [DOI] [PubMed] [Google Scholar]
- 12.Homo sapiens dipeptidyl-peptidase 3 (DPP3), transcript variant 1, mRNA. NCBI Reference Sequence: NM 005700.3.
- 13.Matthews BW. Structural basis of the action of thermolysin and related zinc peptidases. Accounts of Chemical Research. 1988;21(9):333–340. [Google Scholar]
- 14.Medina JF, Wetterholm A, Rådmark O, et al. Leukotriene A4 hydrolase: determination of the three zinc-binding ligands by site-directed mutagenesis and zinc analysis. In: Proceedings of the National Academy of Sciences of the United States of America, vol. 88; 1991; pp. 7620–7624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Baral PK, Jajčanin-Jozić N, Deller S, Macheroux P, Abramić M, Gruber K. The first structure of dipeptidyl-peptidase III provides insight into the catalytic mechanism and mode of substrate binding. Journal of Biological Chemistry. 2008;283(32):22316–22324. doi: 10.1074/jbc.M803522200. [DOI] [PubMed] [Google Scholar]
- 16.Thunnissen MMGM, Nordlund P, Haeggström JZ. Crystal structure of human leukotriene A4 hydrolase, a bifunctional enzyme in inflammation. Nature Structural Biology. 2001;8(2):131–135. doi: 10.1038/84117. [DOI] [PubMed] [Google Scholar]
- 17.Fukasawa KM, Hirise J, Hata T, Ono Y. In rat dipeptidyl peptidase III, His568 is essential for catalysis, and Glu507 or Glu512 stabilizes the coordination bond between His455 or His450 and zinc ion. Biochimica et Biophysica Acta. 2010;1804:2063–2069. doi: 10.1016/j.bbapap.2010.06.015. [DOI] [PubMed] [Google Scholar]
- 18.Oefner C, D’Arcy A, Hennig M, Winkler FK, Dale GE. Structure of human neutral endopeptidase (neprilysin) complexed with phosphoramidon. Journal of Molecular Biology. 2000;296(2):341–349. doi: 10.1006/jmbi.1999.3492. [DOI] [PubMed] [Google Scholar]
- 19.Rees DC, Lewis M, Lipscomb WN. Refined crystal structure of carboxypeptidase A at 1.54 A resolution. Journal of Molecular Biology. 1983;168(2):367–387. doi: 10.1016/s0022-2836(83)80024-2. [DOI] [PubMed] [Google Scholar]
- 20.Hori T, Kumasaka T, Yamamoto M, et al. Structure of a new ‘aspzincin’ metalloendopeptidase from Grifola frondosa: implications for the catalytic mechanism and substrate specificity based on several different crystal forms. Acta Crystallographica Section D. 2001;57(3):361–368. doi: 10.1107/s0907444900019740. [DOI] [PubMed] [Google Scholar]
- 21.Tanaka T, Ichishima E. Molecular properties of aminopeptidase Ey as a zinc-metalloenzyme. International Journal of Biochemistry. 1993;25(11):1681–1688. doi: 10.1016/0020-711x(93)90528-m. [DOI] [PubMed] [Google Scholar]
- 22.Hirose J, Ohsaki T, Nishimoto N, et al. Characterization of the metal-binding site in aminopeptidase B. Biological and Pharmaceutical Bulletin. 2006;29(12):2378–2382. doi: 10.1248/bpb.29.2378. [DOI] [PubMed] [Google Scholar]
- 23.Achstetter T, Ehmann C, Wolf DH. Proteinase yscD. Purification and characterization of a new yeast peptidase. Journal of Biological Chemistry. 1985;260(8):4585–4590. [PubMed] [Google Scholar]
- 24.Büchler M, Tisljar U, Wolf DH. Proteinase yscD (oligopeptidase yscD). Structure, function and relationship of the yeast enzyme with mammalian thimet oligopeptidase (metalloendopeptidase, EP 24.15) European Journal of Biochemistry. 1994;219(1-2):627–639. doi: 10.1111/j.1432-1033.1994.tb19978.x. [DOI] [PubMed] [Google Scholar]
- 25.Klein JR, Klein U, Schad M, Plapp R. Cloning, DNA sequence analysis and partial characterization of pepN, a lysyl aminopeptidase from Lactobacillus delbrueckii ssp. lactis DSM7290. European Journal of Biochemistry. 1993;260:4585–4590. doi: 10.1111/j.1432-1033.1993.tb18224.x. [DOI] [PubMed] [Google Scholar]
- 26.Yan TR, Azuma N, Kaminogawa S, Yamauchi K. Purification and characterization of substrate-size-recognizing metalloendopeptidase from Streptococcus cremoris H61. Applied and Environmental Microbiology. 1987;53:2296–2302. doi: 10.1128/aem.53.10.2296-2302.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Monnet V, Nardi M, Chopin A, Chopin MC, Gripon JC. Biochemical and genetic characterization of PepF, an oligopeptidase from Lactococcus lactis . Journal of Biological Chemistry. 1994;269(51):32070–32076. [PubMed] [Google Scholar]
- 28.Chang PC, Lee YHW. Extracellular autoprocessing of a metalloprotease from Streptomyces cacaoi . Journal of Biological Chemistry. 1992;267(6):3952–3958. [PubMed] [Google Scholar]
- 29.Holmquist B, Vallee BL. Metal substitutions and inhibition of thermolysin: spectra of the cobalt enzyme. Journal of Biological Chemistry. 1974;249(14):4601–4607. [PubMed] [Google Scholar]
- 30.Tan PST, Pos KM, Konings WN. Purification and characterization of an endopeptidase from Lactococcus lactis subsp. cremoris Wg2. Applied and Environmental Microbiology. 1991;57(12):3593–3599. doi: 10.1128/aem.57.12.3593-3599.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Baankreis R, Van Schalkwijk S, Alting AC, Exterkate FA. The occurrence of two intracellular oligoendopeptidases in Lactococcus lactis and their significance for peptide conversion in cheese. Applied Microbiology and Biotechnology. 1995;44(3-4):386–392. doi: 10.1007/BF00169933. [DOI] [PubMed] [Google Scholar]
- 32.Ayora S, Götz F. Genetic and biochemical properties of an extracellular neutral metalloprotease from Staphylococcus hyicus subsp. hyicus . Molecular and General Genetics. 1994;242(4):421–430. doi: 10.1007/BF00281792. [DOI] [PubMed] [Google Scholar]
- 33.Dreyfus LA, Iglewski BH. Purification and characterization of an extracellular protease of Legionella pneumophila . Infection and Immunity. 1986;51(3):736–743. doi: 10.1128/iai.51.3.736-743.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Diermayr P, Kroll S, Klostermeyer H. Influence of EDTA and metal ions on a metalloproteinase from Pseudomonas fluorescens biotype I. Biological Chemistry Hoppe-Seyler. 1987;368(1):57–61. doi: 10.1515/bchm3.1987.368.1.57. [DOI] [PubMed] [Google Scholar]
- 35.Nonaka T, Hashimoto Y, Takio K. Kinetic characterization of lysine-specific metalloendopeptidases from Grifola frondosa and Pleurotus ostreatus fruiting bodies. Journal of Biochemistry. 1998;124(1):157–162. doi: 10.1093/oxfordjournals.jbchem.a022074. [DOI] [PubMed] [Google Scholar]
- 36.Nitanai Y, Satow Y, Adachi H, Tsujimoto M. Crystal structure of human renal dipeptidase involved in β-lactam hydrolysis. Journal of Molecular Biology. 2002;321(2):177–184. doi: 10.1016/s0022-2836(02)00632-0. [DOI] [PubMed] [Google Scholar]
- 37.Tahirov TH, Oki H, Tsukihara T, et al. High-resolution crystals of methionine aminopeptidase from Pyrococcus furiosus obtained by water-mediated transformation. Journal of Structural Biology. 1998;121(1):68–72. doi: 10.1006/jsbi.1997.3940. [DOI] [PubMed] [Google Scholar]
- 38.Wilce MCJ, Bond CS, Dixon NE, et al. Structure and mechanism of a proline-specific aminopeptidase from Escherichia coli . Proceedings of the National Academy of Sciences of the United States of America. 1998;95(7):3472–3477. doi: 10.1073/pnas.95.7.3472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Dobrovetsky E, Dong A, Seitova A, et al. Structural Genomics Consortium (SGC) 2009. Crystal structure of human dipeptidyl peptidase III. [Google Scholar]
- 40.Miyoshi SI, Kawata K, Tomochika KI, Shinoda S. The hemagglutinating activity of Vibrio vulnificus metalloprotease. Microbiology and Immunology. 1999;43(1):79–82. doi: 10.1111/j.1348-0421.1999.tb02376.x. [DOI] [PubMed] [Google Scholar]