

Abstract
Small-molecule compounds remain the major source of therapeutic and preventative drugs. Developing new drugs against a protein target often requires screening large collections of compounds with diverse structures for ligands or ligand fragments that exhibit sufficiently affinity and desirable inhibition effect on the target before further optimization and development. Since the number of small molecule compounds is large, high-throughput screening (HTS) methods are needed. Small-molecule microarrays (SMM) on a solid support in combination with a suitable binding assay form a viable HTS platform. We demonstrate that by combining an oblique-incidence reflectivity difference optical scanner with SMM we can screen 10,000 small-molecule compounds on a single glass slide for protein ligands without fluorescence labeling. Furthermore using such a label-free assay platform we can simultaneously acquire binding curves of a solution-phase protein to over 10,000 immobilized compounds, thus enabling full characterization of protein-ligand interactions over a wide range of affinity constants.
Introduction
In drug discovery, affinity and inhibition screening of thousands to millions of molecules against a protein target or a set of protein targets is routinely required to identify suitable ligands as candidates for further mechanistic and developmental studies. The number of biomolecular binding assays involved is further increased by post-translational modification of proteins such as glycosylation and phosphorylation. It has come to be expected that such a massive amount of screening assays should be done in a highly parallel manner so that endpoints or preferably kinetic constants of thousands of binding reactions are determined simultaneously. Over the last decade, microarrays1–8 and micro-fluidics9–12 have emerged as two major enabling platforms for high-throughput screening. They afford a large number of biochemical reactions and their detection to take place either concurrently (in cases of microarrays) or sequentially at a high rate (in cases of microfluidics). In addition to high throughputs and time-saving, these platforms consume far less reagents than conventional assays. These attributes make them indispensable tools of drug discovery.
Identifying ligands with desirable affinity to a protein target from a large numberof synthetic and natural compounds including aptamers and carbohydrates is facilitated by microarray-based binding assays where ligand candidates are immobilized on a solid support as an addressable array with thousands or tens of thousands distinct features. By exposing the array to a solution-phase protein probe, as many binding reactions occur concurrently. It is advantageous to immobilize small molecules instead of protein targets for two reasons: (1) the number of small molecule compounds is much larger than that of protein targets and thus for high throughput consideration it is sensible to immobilize small molecules as large microarrays and keep relative few protein targets in solution phase; (2) small molecules are more rigid than proteins and thus less prone to conformational change when immobilized through a flexible linker to a solid support. One should expect the immobilized small molecules to better maintain their innate structural and chemical properties. Koehler and coworkers showed that ~ 70% of the compounds from NIH and commercial vendors without a “common” surface-anchoring residue can be immobilized on an isocyanate-functionalized solid support through non-specific reaction of nucleophile groups on these compounds.7, 13 As long as key structural residues on some (if not all) of the “un-immobilizable” compounds are represented in one form or another by the “immobilizable” compounds, the “un-immobilized” compounds are in essence not excluded from screening.
Binding of a protein target to small molecule microarrays (SMM) has mostly been detected ex situ with fluorescence-based methods, by labeling either the protein with fluorescent tags (including incorporation of GFP) or a secondary probe that recognizes the primary protein.3, 7, 13 The combination of SMM and fluorescence-based detection yields endpoints of binding reactions under a specific set of conditions including protein concentration, buffer, incubation time, and post-incubation processing before fluorescence readout. There are drawbacks in fluorescence-based SMM detection: (1) labeling protein targets with fluorescent tags requires extra reaction steps, and the efficiency of labeling varies from protein to protein; (2) directly labeling a protein alters the protein affinity profile, in ways often uncharacterized14, 15; (3) ex-situ measurements render the readout a function of assay conditions and limited to ligands with high affinity or small dissociation rates9, 14; (4) end-point measurements yield significantly limited information on binding kinetics and as a result offer incomplete characterization of protein-ligand interactions14.
We describe a combination of an ellipsometry-based optical scanner16, 17 and small-molecule microarrays on isocyanate-functionalized glass surfaces as a non-fluorescence-based assay platform in high-throughput screening of small molecule libraries for protein ligands. This platform avoids the drawbacks associated with the fluorescence-based detection. The in-situ nature of the ellipsometry detection enables measurements of binding curves and in turn determination of binding kinetic constants over a wide dynamic range.17, 18
Small molecule microarray of a NCI/DTP compound library
For small molecule compounds, we use five compound sets from NCI/DTP Open Repository: (1) Challenge set with 57 compounds (10 mM in DMSO, 20 μL); (2) Natural Products set with 235 compounds (10 mM in DMSO, 20 μL); (3) Structural Diversity set with 1,990 compounds (10 mM in DMSO, 20 μL); (4) Mechanistic Diversity set with 879 compounds (1 mM in DMSO, 20 μL); (5) an Open set with 4,800 compounds (1 mM in DMSO, 20 μL). There are a total of 7,761 compounds that come in 96-well plates. Directly from the stock solutions from NCI and using an OmniGrid100 Arrayer (DIGILAB, MA) with 8 printing tips, we immobilized these compounds into two small molecule microarrays on separate isocyanate-functionalized glass slides, prepared with a protocol developed by Bradner and coworkers with minor modifications13. We printed one half of the NCI compounds in duplicate on one slide along with a set of control compounds, and the other half of the NCI compounds also in duplicate on the second slide along with the same set of control compounds. Each slide contained a total of ~ 10,000 printed features over an area of 2 cm × 4 cm. A printed glass slide was assembled with a fluid chamber so that the printed microarray could be processed while being imaged or read out in real time with an ellipsometry-based scanner.16, 17
Optical image of NCI compound microarrays on functionalized glass slides acquired with an ellipsometry-based optical scanner
We detected printed NCI microarrays and subsequent reactions of these microarrays with solution-phase protein probes using an oblique-incidence reflectivity difference scanning microscope, - a special form of scanning ellipsometry.16, 17 The oblique-incidence reflectivity difference (OI-RD) signal is defined as the difference of fractional reflectivity changes between the p-polarized and s-polarized components of a monochromatic light off a solid surface when it is covered with an immobilized molecular layer, (rp − rp0)/rp0 − (rs − rs0)/rs0 ≡ Δp − Δs 19, 20. rp0 and rs0 are the complex reflectivities of the bare surface; rp and rs are the reflectivities of the surface when it is covered with the immobilized molecular layer. Δp − Δs is proportional to the surface mass per unit area Γ of the immobilized molecular layer just as surface-plasmon resonance (SPR) responses.16, 17, 19, 21 Using an OI-RD scanning microscope, we directly measure Δp - Δs from immobilized NCI compound microarrays on functionalized glass slides.
Figure 1 shows the OI-RD image (equivalent to an image of the surface mass density Γ16, 17, 19 of an NCI compound microarray taken before the array was further processed. The features along the perimeter are bovine serum albumin (BSA); the features immediately inside BSA features are biotin-conjugated BSA; both BSA and biotin-BSA are control features. Further inside are ~ 3,900 NCI compounds printed in vertical pairs. They form 24 rectangular blocks. Printed horizontally between blocks of the NCI compounds are other control compounds.
Figure 1.

OI-RD image of a printed NCI compound microarray with ~ 10,800 spots immobilized on isocyanate-functionalized glass slide before further processing. The microarray contains 3,900 NCI/DTP compounds printed in vertical duplicate that form 24 blocks. The double borders (BSA and biotin-conjugated BSA) and spots as double lines between the blocks are control features. The image covers an area of 2 cm× 4 cm.
Binding reaction of human kinase insertion domain receptor (KDR) with 7,761 NCI compounds on microarrays – label-free endpoint measurement
Human kinase insertion domain receptor (KDR or VEGFR2) fused to the Fc fragment of rabbit IgG was purchased from Epitomics (Burlingame, CA) as the protein probe. On fresh NCI compound microarrays, we performed reactions of KDR with 7,761 compounds by incubating the microarrays in a KDR solution at 0.0068 mg/ml in 1× PBS for 60 minutes and then washing the microarrays with 1× PBS. Figure 2 shows the change in OI-RD image of one NCI compound microarray after the reaction. The distinguishable pairs in the vertical direction reveal the few compounds that captured KDR. Fusing this method, we identified 27 “hits” from a total of 7,761 NCI compounds that reacted with KDR with high affinity.
Figure 2.

Change in OI-RD image of the NCI compound microarray acquired in 1× PBS after incubation in a solution of KDR 0.0068 mg/ml in 1× PBS for 60 minutes. The distinguishable doublets in vertical directions reveal the compounds that have captured KDR from the solution. We can identify 14 “hits” out of ~ 3,900 immobilized NCI compounds on this microarray.
Simultaneous measurement of 10,000 protein-ligand binding curves, – reaction of a glycan-binding protein with a host of glycans and protein targets
The change in optical image as displayed in Figure 2, albeit acquired label-free, is an endpoint measurement of protein-compound reactions after the solution of the protein probe has been replaced with 1× PBS for a period of time (~ 30 minutes). As a result, the image only reports binding reactions with dissociation times τd being comparable or longer than 30 minutes14 and thus with equilibrium dissociation constant Kd (inversely proportional to τd) in the range of nM. In order to capture binding reactions with Kd ranging from sub-mM to pM and fully characterize reaction kinetics, it is necessary to record binding curves of protein-compound reactions. To respond to such a need, we developed the capability of our OI-RD scanning microscope to simultaneously record binding curves of a protein probe with 10,000 immobilized ligand candidates. To illustrate the instrument capability, we fabricated a large microarray with 10,880 printed features printed on an epoxy-coated glass slide. The microarray consists of 16 “identically” printed sub-arrays. Each sub-array (Figure 3) consists of 24 distinct protein targets that are printed in quadruplet at 6 different target concentrations (0.5 μM to 16 μM).16, 19, 22 These 24 protein targets are: (1) human IgG (HM); (2) mouse IgG (MS); (3) rabbit IgG (RB); (4–6) polyclonal goat IgG against human/mouse/rabbit IgG (GT anti-HM, GT anti-MS, GT anti-RB); (7) Methamphetamine-BSA (Meth-BSA); (8) tetrahydrocannabinol-BSA (THC-BSA); (9) morphine-BSA (Morph-BSA); (10) Theophylline-BSA (THP-BSA); (11) phenobarbital-BSA (PB-BSA); (12) digoxin-BSA (Dig-BSA); (13) Metallothionein (Metal); (14–17) biotin-BSA (B-BSA) with the loading molar ratio of the NHS-biotin ester to BSA at 5×, 10×, 20×, and 40×; (18–21) iminobiotin-BSA (IB-BSA) with the loading molar ratio of the NHS-biotin ester to BSA at 5×, 10×, 20×, and 40×; (22) 2,4-dinitrophenol-BSA (DNP-BSA); (23) glucose-BSA (Glc-BSA); and finally (24) maltose-BSA (Mal-BSA). The remaining 1664 features are printed for controls.
Figure 3.
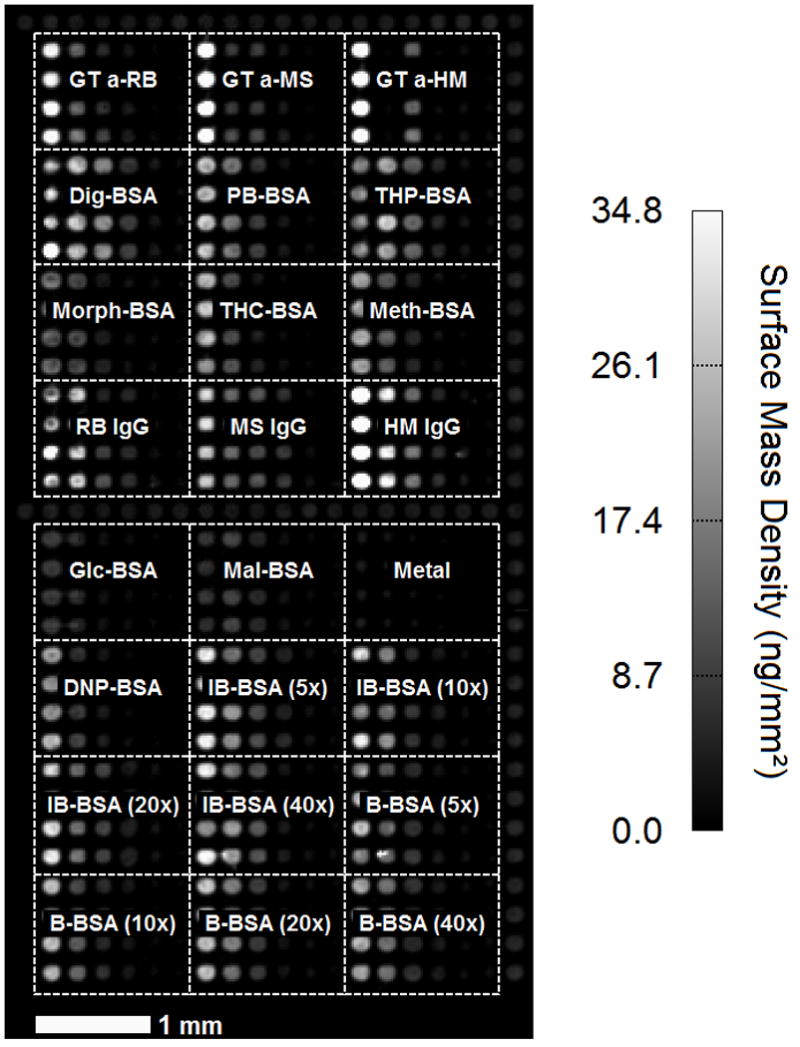
The surface mass density of a subarray that consists of 24 distinct molecular targets printed in quadruplet and in 6 target concentrations (16 μM, 8μM, 4 μM, 2 μM, 1 μM, 0.5 μM). 16 such subarrays along with the control features around the perimeters of these subarrays form a 10,880-feature molecular target microarray that is fabricated on an epoxy-functionalized glass slide over an area of ~ 2 cm × 4 cm. The surface mass density map is determined from the OI-RD images of the microarray 16, 19, 22 taken before it reacts with protein probes of interest such as ConA.
We used concanavalin A (ConA) as the protein probe. ConA is known to react specifically with glucose. Unlike monoclonal antibodies, ConA is also expected to react with other molecular motifs as we will reveal here. Instead of acquiring OI-RD images before and after incubation with a ConA solution at a concentration of 300 nM in 1× PBS, we measured the difference between the optical signal from one pixel inside the target area and the average of the optical signals from two pixels in the neighboring unprinted area as a background-corrected readout. It took ~ 20 seconds to sequentially read from 10,880 immobilized targets for one time point. By repeating the time-point measurement during incubation phase (for 30 minutes after the protein probe solution replaces 1× PBS) and subsequent dissociation phase (for 90 minutes after the protein probe solution is replaced by 1× PBS again), we acquired 10,880 ConA binding curves in one experiment at a probe concentration of 300 nM. Figure 4 displays 576 out of 10,880 binding curves of ConA to the protein target microarray. As expected, ConA reacted with glucose-BSA and yet not with maltose-BSA, indicating the specific affinity to glucose. However it is clear that ConA reacted non-specifically with all IgG molecules. There is also evidence of non-specific reaction with drug-BSA conjugates. We note that even though the density of the immobilized glucose-BSA and the amount of the captured ConA by glucose-BSA changed by more than a factor of 10, the temporal behaviors of the binding curves remained unchanged as one should expect. As a result the binding kinetics extracted from these binding curve sets do not depend on which pixel inside the target area is used to yield the binding curve.17
Figure 4.

A small portion of 10,880 simultaneously acquired binding curves of ConA to as many immobilized targets on an epoxy-functionalized glass slide surface at ConA concentration of 300 nM. In each panel, the vertical axis is the surface mass density of the captured ConA and has the full scale of 10 ng/mm2. The horizontal axis is in minutes and has a full scale of 120 minutes. The incubation phase starts at t = 0 and lasts for 30 minutes. The dissociation phase starts at t = 30 minutes and lasts for 60 minutes. In addition to specific reactions with glucose, ConA also reacted with the immobilized IgG molecules.
Our present microarray-based binding curve detection technology is moving microarrays from a highly parallel end-point assay platform to an as highly parallel kinetic constant assay platform that directly yields equilibrium dissociation constants. It is an effective tool for affinity ranking of ligands of a protein probe with Kd ranging from pM to sub-mM. It enables simultaneous characterization of both specific and non-specific reactions of a protein probe with a wide variety of immobilized targets as we illustrated here. With 2 ~ 3 such OI-RD optical scanners simultaneously in operation, we should be able to measure equilibrium dissociation constants of a protein probe against 100,000 compounds in a couple of days.
Acknowledgments
This work was supported by NIH under NIH-R01-HG003827 and by the University of California under UC Discovery Grant #bio09-156225.
References
- 1.Falsey JR, Renil M, Park S, Li S, Lam KS. Peptide and small molecule microarray for high throughput cell adhesion and functional assays. Bioconjugate chemistry. 2001;12(3):346–53. doi: 10.1021/bc000141q. [DOI] [PubMed] [Google Scholar]
- 2.Liang PH, Wu CY, Greenberg WA, Wong CH. Glycan arrays: biological and medical applications. Current opinion in chemical biology. 2008;12(1):86–92. doi: 10.1016/j.cbpa.2008.01.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.MacBeath G, Koehler AN, Schreiber SL. Printing Small Molecules as Microarrays and Detecting Protein-Ligand Interactions en Masse. J Am Chem Soc. 1999;121:7967–7968. [Google Scholar]
- 4.MacBeath G, Schreiber SL. Printing proteins as microarrays for high-throughput function determination. Science. 2000;289(5485):1760–3. doi: 10.1126/science.289.5485.1760. [DOI] [PubMed] [Google Scholar]
- 5.Schena M. Microarray Analysis. Wiley; 2003. [DOI] [PubMed] [Google Scholar]
- 6.Uttamchandani M, Walsh DP, Yao SQ, Chang YT. Small molecule microarrays: recent advances and applications. Current opinion in chemical biology. 2005;9(1):4–13. doi: 10.1016/j.cbpa.2004.12.005. [DOI] [PubMed] [Google Scholar]
- 7.Vegas AJ, Fuller JH, Koehler AN. Small-molecule microarrays as tools in ligand discovery. Chemical Society reviews. 2008;37(7):1385–94. doi: 10.1039/b703568n. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zhu H, Bilgin M, Bangham R, Hall D, Casamayor A, Bertone P, Lan N, Jansen R, Bidlingmaier S, Houfek T, Mitchell T, Miller P, Dean RA, Gerstein M, Snyder M. Global analysis of protein activities using proteome chips. Science. 2001;293(5537):2101–5. doi: 10.1126/science.1062191. [DOI] [PubMed] [Google Scholar]
- 9.Maerkl SJ, Quake SR. A systems approach to measuring the binding energy landscapes of transcription factors. Science. 2007;315(5809):233–7. doi: 10.1126/science.1131007. [DOI] [PubMed] [Google Scholar]
- 10.Momez FA. Biological Application of Microfluidics. John Wiely and Sons; Hoboken, New Jersey: 2007. [Google Scholar]
- 11.Ottesen EA, Hong JW, Quake SR, Leadbetter JR. Microfluidic digital PCR enables multigene analysis of individual environmental bacteria. Science. 2006;314(5804):1464–7. doi: 10.1126/science.1131370. [DOI] [PubMed] [Google Scholar]
- 12.Thorsen T, Maerkl SJ, Quake SR. Microfluidic large-scale integration. Science. 2002;298(5593):580–4. doi: 10.1126/science.1076996. [DOI] [PubMed] [Google Scholar]
- 13.Bradner JE, McPherson OM, Koehler AN. A method for the covalent capture and screening of diverse small molecules in a microarray format. Nature protocols. 2006;1(5):2344–52. doi: 10.1038/nprot.2006.282. [DOI] [PubMed] [Google Scholar]
- 14.Fei YY, Sun YS, Li YH, Lau K, Yu H, Chokhawala HA, Huang SS, Landry JP, Chen X, Zhu XD. Fluorescent Labeling Agents Change Binding Profiles of Glycan-Binding Proteins. Molecular BioSystems. 2011;7(12):3343–3352. doi: 10.1039/c1mb05332a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Sun YS, Landry JP, Fei YY, Zhu XD, Luo JT, Wang XB, Lam KS. Effect of fluorescently labeling protein probes on kinetics of protein-ligand reactions. Langmuir: the ACS journal of surfaces and colloids. 2008;24(23):13399–405. doi: 10.1021/la802097z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Fei YY, Landry JP, Sun YS, Zhu XD, Luo JT, Wang XB, Lam KS. A novel high-throughput scanning microscope for label-free detection of protein and small-molecule chemical microarrays. Rev Sci Instrum. 2008;79(1):013708. doi: 10.1063/1.2830286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Landry JP, Fei YY, Zhu XD. Simultaneous Measurement of 10,000 Protein-Ligand Affinity Constants Using Microarray-based Kinetic Constant Assays. Assay and Drug Development Technologies. 2011 doi: 10.1089/adt.2011.0406. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sun YS, Landry JP, Fei YY, Zhu XD, Luo JT, Wang XB, Lam KS. Macromolecular scaffolds for immobilizing small molecule microarrays in label-free detection of protein-ligand interactions on solid support. Anal Chem. 2009;81(13):5373–80. doi: 10.1021/ac900889p. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Landry JP, Sun YS, Guo XW, Zhu XD. Protein reactions with surface-bound molecular targets detected by oblique-incidence reflectivity difference microscopes. Applied optics. 2008;47(18):3275–88. doi: 10.1364/ao.47.003275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zhu XD, Landry JP, Sun YS, Gregg JP, Lam KS, Guo XW. Oblique-incidence reflectivity difference microscope for label-free high-throughput detection of biochemical reactions in a microarray format. Appl Optics. 2007;46(10):1890–1895. doi: 10.1364/ao.46.001890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Zhu XD. Comparison of two optical techniques for label-free detection of biomolecular microarrays on solids. Opt Commun. 2006;259(2):751–753. [Google Scholar]
- 22.Fei YY, Schmidt A, Bylund G, Johansson DX, Henriksson S, Lebrilla C, Solnick JV, Boren T, Zhu XD. Use of Real-Time, Label-Free Analysis in Revealing Low-Affinity Binding to Blood Group Antigens by Helicobacter pylori. Anal Chem. 2011;83(16):6336–6341. doi: 10.1021/ac201260c. [DOI] [PMC free article] [PubMed] [Google Scholar]