

Abstract
Nuclear RNA processing requires dynamic and intricately regulated machinery composed of multiple enzymes and their cofactors. In this review, we summarize recent experiments using Saccharomyces cerevisiae as a model system that have yielded important insights regarding the conversion of pre-RNAs to functional RNAs, and the elimination of aberrant RNAs and unneeded intermediates from the nuclear RNA pool. Much progress has been made recently in describing the 3D structure of many elements of the nuclear degradation machinery and its cofactors. Similarly, the regulatory mechanisms that govern RNA processing are gradually coming into focus. Such advances invariably generate many new questions, which we highlight in this review.
Keywords: Cryptic unstable transcript, Exosome, mRNA, Mtr4p, Polyadenylation, rRNA, Small nuclear RNA, Small nucleolar RNA, TRAMP, tRNA
RNA PROCESSING AND DEGRADATION
Quality control of nuclear RNA requires both processing and surveillance pathways. In particular, rRNA, small nucleolar RNA (snoRNA), small nuclear RNA (snRNA), mRNA, and tRNA species are all transcribed as pre-RNAs, which must then be cleaved and/or trimmed to produce functional RNAs[1,2]. Likewise, many aberrant RNAs are detected by surveillance mechanisms and thereby eliminated from the nuclear RNA pool. Any byproducts from the conversion of pre-RNA to functional RNA must be rapidly degraded by exonucleolytic digestion. In addition, non-translated RNAs, such as cryptic unstable transcripts (CUTs), are subject to processing and degradation. The nuclear exosome is the major degradation machine involved in both pathways of nuclear RNA quality control. In Saccharomyces cerevisiae (S. cerevisiae), the nuclear exosome is a collection of six RNase PH homologues (Rrp41p, Rrp42p, Rrp43p, Rrp45p, Rrp46p and Mtr3p), which are inactive and form a scaffolding ring structure[3], three putative RNA binding proteins (Rrp4p, Rrp40p and Csl4p)[4], and two active 3′→5′ exonucleases, Rrp44p[3-5] and Rrp6p[6,7]. Many exosome substrates contain structured segments that preclude complete processing or degradation by the exosome alone, thus requiring cofactors to ensure generation of the desired end products. One of those cofactors, Mtr4p, is an indispensible partner of the exosome, and likely maintains the momentum of exonucleolytic activity as both Mtr4p and the exosome move through structured RNA substrates. This section describes the different types of RNA in S. cerevisiae whose processing is affected by the Mtr4p-exosome system.
rRNA
In eukaryotes, rRNA synthesis is initiated in the nucleolus by RNA polymerase I (Pol I), which synthesizes the 90S rRNA precursor. This precursor is co-transcriptionally cleaved by RNase III to release the initial 90S preribosomal particle, which contains the 35S pre-rRNA[8]. 35S pre-rRNA contains the sequences of three rRNAs that are the transcripts for 18S, 5.8S and 25S ribosomal subunits. These three transcripts are separated by two internal transcribed spacers (ITSs) and flanked by two external transcribed spacers (ETSs)[9]. Through a series of cleavage events, 35S pre-rRNA is converted into mature rRNAs (Figure 1). Pre-rRNA processing begins with cleavage at site A0, removing the 5′ ETS, at site A3, releasing 23S rRNA, and at site A2, generating both 20S and 27SA2 intermediates. 20S pre-rRNA is further processed in the cytoplasm to become mature 18S rRNA. 27SA2 continues maturation in the nucleus by two separate pathways: about 85% of 27SA2 is cleaved at site A3 and then rapidly trimmed to site B1S, while 15% of 27SA2 is cleaved directly at site BIL to become 27SBIL. 27SBIL/IS is then cleaved at sites C1, and C2[10]. These cleavage events produce 7SL/S and 25S rRNA. 7SL/S are then trimmed to produce the 6SL/S rRNA. Final maturation of 5.8S rRNA takes place in the cytoplasm[11]. 5.8S rRNA undergoes exonucleolytic processing to produce a mature 3′ end and further cleavage at site BIS to produce 5.8SS. Final maturation of 25S rRNA occurs through cleavage at site B2[7,10,12].
Figure 1.
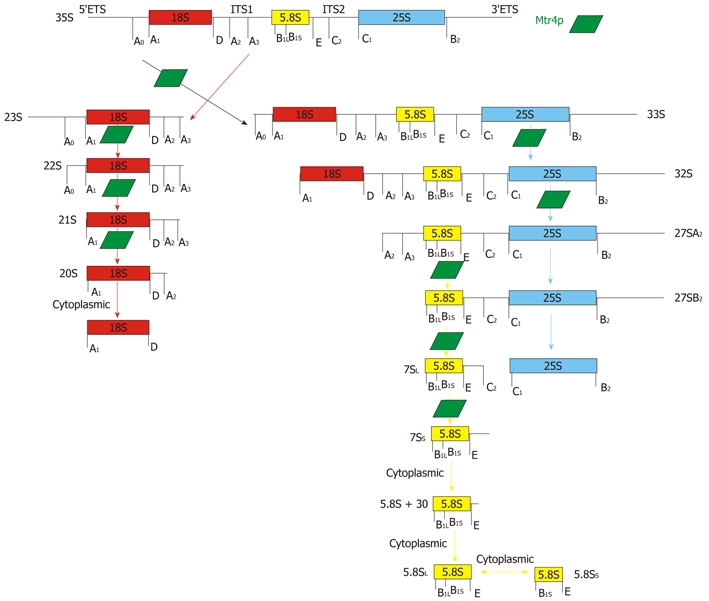
Schematic representation of rRNA processing pathway in Saccharomyces cerevisiae. The full-length rRNA precursor undergoes several cleavage steps to produce functional 18S, 5.8S and 25S rRNAs for 60S and 40S ribosomes. Green trapezoids indicate points in the processing pathway at which Mtr4p activity has been shown to play a role. Also indicated are steps of processing that have been found to be completed in the cytoplasm[7,11].
The processing events for rRNA are closely monitored by a series of decay factors that are involved when a cleavage reaction fails or stalls. Maturation and assembly of the rRNA subunits involves at least 170 accessory proteins with various functions including endo- and exoribonucleases, RNA helicases, other “assembly factors”, and just as many small nucleolar Ribonucleoproteins (snoRNPs)[8]. In addition to cleavage events, multiple modifications are required to produce mature rRNAs. The most common modifications are the isomerization of uridines to pseudouridines and methylation of select ribose 2′-hydroxyl groups. In S. cerevisiae, about 50 rRNA sites of each type are modified[12]. Once fully processed, 18S rRNA is located in the 40S small ribosomal subunit, while 5S, 5.8S and 25S rRNAs combine to create the 60S large ribosomal subunit. Once assembled, the pre-40S subunit is exported to the cytoplasm, whereas the pre-60S subunit requires nuclear maturation to be exported. Once in the cytoplasm, both pre-ribosome units undergo further processing and are assembled into mature subunits[13]. Mtr4p, a nuclear RNA helicase, is a major participant in the exoribonucleolytic rRNA processing events. Specifically, Mtr4p is required for proper processing following cleavage events which occur at A0, A2, A3, C1, B1S, and E sites[7,10] (Figure 1).
snoRNAs
In S. cerevisiae, most snoRNAs are transcribed from independent genes or excised from polycistronic transcripts. Polycistronic transcripts can contain as many as seven different snoRNAs. Processing of these transcripts is catalyzed by RNase three protein 1 (Rnt1p) and different 5′→3′ and 3′→5′ exonucleases[14]. Production of intronic snoRNAs is dependent on the RNA lariat-debranching enzyme Dbr1p. Depletion of this enzyme causes snoRNAs to become caught within the host lariats[15]. Following transcription, snoRNAs must be processed from both the 3′ and 5′ ends to generate a mature snoRNA. Transcription termination is dependent on a complex of two RNA-binding proteins, Nrd1p and Nab3p, and an RNA helicase, Sen1p. Nrd1p interacts with the C-terminal domain of RNA Pol II and the exosome, an RNA processing and degradation complex, to link termination with processing. snoRNA termination is followed by polyadenylation by the poly(A) polymerase Pap1p. It is this polyadenylation that initiates 3′ end processing by the exosome[16].
Mature transcripts of snoRNA can be structurally and functionally divided into three categories: C/D box-containing snoRNAs; H/ACA box-containing snoRNAs; and the mitochondrial RNA processing (MRP) snoRNA. The box C/D and box H/ACA sequences serve as family-specific nucleolar localization elements (NoLEs). Box C/D snoRNAs are intron-encoded and contain two distinct motifs located at the 5′ and 3′ ends of the mature RNA: box C (UGAUGA) and box D (CUGA). Boxes C′ and D′ are located within the interior of the snoRNA[15,17]. The C and D motifs are brought together in the pre-snoRNA to form a distinctive structural motif. This motif is required for processing, stability, and localization within the nucleolus. Most box C/D snoRNAs have long sequences complementary to rRNAs located immediately before the box D or D′ motif[15]. These sequences create duplex regions at sites of 2′-O-ribose methylation[17]. The box C/D snoRNAs are associated with the nucleolar protein 1 (Nop1p), which is required for many steps of ribosome assembly[14,17].
Box H/ACA snoRNAs contain a hinge-box (H-box of the sequence ANANNA) and a trinucleotide ACA sequence downstream of the 3′-terminal stem-loop[14]. H/ACA containing snoRNAs are required for pseudouridyl (Ψ) formation in rRNA[18]. In order to modify the rRNA, a box H/ACA snoRNA base pairs to nucleotides flanking the substrate uracil, allowing the modifying enzyme to be positioned for catalysis[19]. Box H/ACA snoRNAs are associated with four proteins: Cbf5p, Gar1p, Nhp2p, and Nop10p; the factor Cbf5p is responsible for the pseudouridylation of rRNA[20]. Box H/ACA snoRNAs can be further categorized by the presence or absence of a Cajal-body-specific localization signal (CAB). Those H/ACA snoRNAs with a CAB are considered small Cajal-body-specific RNAs (scaRNAs) and contain two CAB boxes within the terminal loops of both the 5′ and 3′ hairpins[21]. The third category of snoRNAs, MRP RNA, is the least characterized. This snoRNA is a component of the RNase MRP, a ribonucleoprotein enzyme that cleaves at pre-rRNA at A3[14]. Mutants of this RNA introduced into RNase MRP alter the ratio of the long and short forms of 5.8S rRNA (5.8SL and 5.8SS) produced in the cell[22].
SnoRNAs transit through the Cajal bodies to undergo final maturation before being transported to the nucleolus. The H/ACA box snoRNAs that contain the CAB binding sequence remain in the Cajal bodies. Once processed, snoRNAs participate in multiple steps of pre-ribosomal assembly prior to cytoplasmic export. Within the nucleolus, snoRNAs form snoRNPs which, along with ribosomal and nonribosomal proteins, help to assemble the 90S pre-ribosome in the nucleolus[8,12]. Mtr4p in conjunction with the exosome has been implicated in proper maturation of polycistronic transcripts and independently transcribed pre-snoRNAs, as well as intron-derived snoRNAs. Although Mtr4p is not thought to be a universal factor in all snoRNA processing, it has been implicated in proper 3′ end processing of snR44, snR73, snR72, U14, and snR33[23,24].
snRNAs
S. cerevisiae contains 24 different snRNAs, six of which are dispensable for growth[1]. Each snRNA contains a trimethyl cap at the 5′ terminus and is encoded by a single-copy gene. The major function of snRNAs is to catalyze the maturation of pre-mRNA to mRNA via the spliceosome. During spliceosome assembly, snRNAs and pre-mRNAs, along with at least 150 protein factors, undergo a variety of conformational changes to establish specific and critical RNA-RNA interactions between the snRNAs and the pre-mRNAs[25]. The five snRNAs within the major spliceosome are U1, U2, U4, U5, and U6. U1 and U2 are used predominantly for establishing critical contacts between the spliceosome and the pre-mRNA. These contacts include determinants of the correct splice site by interaction with both the intron and exon of the pre-mRNA[26]. U1 and U2 snRNAs bind to sequences at the site of 5′ cleavage and the branch point adenosine (A), while U5 participates in interactions involving the 3′ splice site[27]. U2, U5, and U6 are part of the active spliceosome.
S. cerevisiae snRNAs are transcribed predominantly by RNA Pol II, although U6 snRNA is transcribed by RNA Pol III. SnRNAs contain TATA boxes located upstream of their transcription start sites[1]. The U4 snRNA transcript is cleaved by Rnt1p, polyadenylated by Pap1p, and finally processed by Rrp6p, the exosome and Mtr4p to produce a functional snRNA[24]. Rnt1p, Mtr4p and the exosome appear to have some role in U5 snRNA processing, as a 3′-extended intermediate accumulates in both an rrp6-∆ strain and strains depleted of Rrp41p, Rrp45p, and Mtr4p[23]. Furthermore, the ratio of the long and short forms of U5 (U5L and U5S) is drastically shifted in favor of U5S in strains depleted of Rrp41p, Rrp45p, and Mtr4p and in strains lacking Rrp6p or Rnt1p[23]. The pathway by which other snRNAs are produced is less well defined, although all are thought to undergo 3′ end processing[23].
mRNA
Pol II is responsible for the transcription of pre-mRNAs. Unlike many RNAs, mRNA undergoes processing steps co-transcriptionally. Events such as 5′ end capping and some splicing take place while the RNA is still being synthesized. Pre-mRNA processing involves five activities: 5′ end capping, editing, splicing, 3′ end formation, and degradation. With the exception of editing, each of these processes can occur co-transcriptionally. The ability to process co-transcriptionally is in part due to the fact that Pol II stimulates the activity of many required processing factors[28]. The initial step in pre-mRNA processing is the addition of the 5′ end cap. This process is directly linked to Pol II via direct binding of the three capping enzymes to Pol II. Following phosphate removal, Ceg1p utilizes GTP to add a 5′ phosphate onto the 5′ end of the pre-mRNA. After the 5′ phosphate is added, a 5′ to 5′ linkage is completed, and Abd1p utilizes S-adenosylmethionine (SAM) to donate a methyl group to position 7 on the guanosine cap[29,30]. Completion of this process caps the protein with the m7G(5′)ppp(5′)X cap. Once the 5′ cap is in place splicing can begin.
Splicing takes place both co-transcriptionally and post-transcriptionally. Post-transcriptional splicing event sites are marked co-transcriptionally. Splicing is initiated by recruitment of the spliceosome, which is composed of snRNAs and a variety of proteins[27]. After splicing has been completed, the 3′ end of the mRNA must also be processed. 3′ end formation of mRNA in S. cerevisiae requires three elements: (1) the efficiency element; UAG...UAUGUA and similar sequences, which enhances the efficiency of downstream positioning elements; (2) the positioning element AATAAA which positions the poly(A) site; and (3) the poly(A) site, which contains a pyrimidine (Py)(A)n[31,32]. The efficiency element includes the sequences UAG...UAUGUA, UAUAUA, UUUUUAUA, and UACAUA. These elements act in concert with the efficiency element to determine where on the mRNA the poly(A) site is located. Once bound, Pap1p is able to polyadenylate the 3′ end of the mRNA[33]. mRNAs that require splicing yet are unspliced are rapidly degraded in the nucleus by the exosome through 3′→5′ degradation and to a lesser extent by Rat1p directed 5′→3′ degradation[34]. Mtr4p has been implicated in the degradation of both mRNAs which lack a full length poly(A) tail[35] and mRNAs that are hyperadenylated[36].
tRNA
There are 274 tRNA genes in S. cerevisiae and each is transcribed by Pol III. All tRNA transcription occurs within the nucleolus[37-39]. Following transcription, tRNA undergoes extensive folding and modification before export. Over 100 kinds of nucleotide modifications occur on tRNA. These modifications are required for proper folding of the tRNA into the canonical L-shaped tertiary structure as well as for the function of tRNAs in decoding. Lack of these modifications leads to degradation of the tRNA. For example, the 1-methyladenosine tRNA methyltransferase Gdc10p/Gdc14p modifies A58 of tRNAiMet to a 1-methyladenosine (m1A58). When this modification is not made pre-tRNAiMet is degraded by Mtr4p, the Trf4/5p, Air1/2p, and Mtr4p Complex (TRAMP), Rrp6p and the exosome[13,40,41].
CUTs
CUTS are a class of Pol II transcripts in S. cerevisiae. A typical CUT has a capped 5′ end and can vary in length depending on when transcription stops. This creates heterogeneity and multiple polyadenylated 3′ ends[42]. There have been between 50 000 and 67 000 CUTs identified and these can be grouped into 1779 clusters. Of the clusters, 1496 do not correspond to open reading frames or non-coding RNAs (ncRNAs). Each CUT is between 200 and 500 nucleotides in length and undergoes Nrd1p/exosome/TRAMP-dependent degradation. One hundred and six of the clusters are mapped to ncRNA, and 134 are located within intron-containing pre-mRNAs. Forty-three clusters remain unclassified but some of the members of these clusters are transcription start sites[43].
CUTs are produced from inter- and intragenic regions of the genome. These regions may represent 10% of the intergenic transcripts in S. cerevisiae[42]. CUTs are derived from a variety of specific transcripts and are most prevalent in nucleosome-free regions. Most of the sites which produce CUTs are within the promoter regions of bona fide genes[43]. CUTs derived from gene promoters are believed to have a regulatory role, conferred by their co-transcription. However, most identified CUTs are antisense to the flanking genes and appear to arise via promoter divergent transcription[43]. Specifically, it is believed that CUTs may act as gene regulators in a pathway that resembles RNA interference (RNAi) despite the lack of evidence for an RNAi pathway in S. cerevisiae[44]. In fact, several examples of CUT-dependent RNA transcription regulation have been cited recently[45-47].
Regulation of CUTs is directed by the RNA binding proteins Nrd1p and Nab3p. Nrd1p and Nab3p are located on chromosomes near the sites of CUT transcription, and through this proximity they are able to bind the CUTs and cause termination. Nrd1 interacts with the CTD of RNA Pol II and also binds Nab3p. Nab3p binds the RNA and Nrd1p; this combination of events leads to the termination of CUTs. Interactions of both Nrd1p and Nab3p with the exosome propagate degradation[48]. Degradation is dependent upon polyadenylation of the CUT by the poly(A) polymerase Trf4p but the mechanism whereby the 3′ end is made available to Trf4p is unknown. In some instances, Mtr4p is involved in the degradation of CUTs, through stimulation of the exonuclease activity of Rrp6p, as well as through its role as a member of the TRAMP complex[42,49].
PATHWAYS FOR NUCLEAR RNA PROCESSING AND DEGRADATION
The processing and degradation of RNAs is essential to cell survival. Multiple RNAs are transcribed as pre-RNAs that require processing to convert them to functional RNAs. In particular, each rRNA, snRNA and snoRNA must have 3′ end extensions removed to yield functional RNAs[2]. In addition, aberrant RNAs and byproducts from RNA processing events must be removed from the cellular milieu. Two pathways exist for nuclear RNA processing and degradation, the 5′→3′ exonucleolytic pathway and the more prevalent 3′→5′ exonucleolytic pathway. 5′→3′ degradation is performed by Rat1p, the Rat1p-interacting protein Rai1p, and additional cofactors. Rat1p is a 5′→3′ RNA exonuclease, and Rai1p interacts with Rat1p to stabilize its exonuclease activity in vitro[50]. Specific cofactors are recruited by the 5′→3′ exonuclease machinery for degradation of each type of RNA. 3′→5′ degradation is performed by the nuclear exosome, Mtr4p, and a series of RNA species specific cofactors. The nuclear exosome is a collection of six RNase PH homologs[3], three putative RNA binding proteins[51] and two active 3′→5′ exonucleases. In this section the functions, characteristics, cofactors, and activities of each RNA processing and degradation pathway are discussed.
5′→3′ nuclear RNA processing and degradation by Rat1p
Transcription of RNAs involves initiation, elongation and termination of the RNA. The final step of termination is controlled by two different pathways: either the poly(A)-site-independent pathway, or the poly(A)-site-dependent pathway. Poly(A)-independent transcription termination requires the helicase Sen1p, and sequence-specific RNA binding proteins, Nab3p and Nrd1p[52]. The poly(A) independent termination complex interacts with the exosome and undergoes 3′→5′ degradation, which is further discussed in the following section.
Poly(A)-site-dependent termination is less well characterized, and the components required are not all known. At present, three theories on how termination is achieved exist: the “torpedo model”[53,54]; the “allosteric model”[55]; and the “hybrid model”[56]. The torpedo model suggests that the 5′-end of the RNA, following the poly(A) cleavage site is used as a substrate for an exonuclease. When the exonuclease catches up with the polymerase it releases the DNA. In this model, the exonuclease acts as a torpedo jettisoning the polymerase from the DNA and thereby terminating transcription[53,54] (Figure 2A). The allosteric model proposes that the polymerase is altered by conformational changes that take place because of poly(A) site recognition, or loss of anti-termination factors by recognition of the poly(A) site. In this model processivity is lost and termination is gradual[55] (Figure 2B). The current opinion is that these two methods work together cohesively in a hybrid model. The polymerase changes conformation upon poly(A) site recognition, and Rat1p and Pcf11p cause a pause in transcription. In this model, the Rat1p/Rai1p complex is used to remove RNA transcribed past the poly(A) site[56] (Figure 2C). In each of the models, Rat1p acts as the primary exonuclease in the nucleus, whereas Xrn1p is the cytoplasmic nuclease, and also plays a minor role in nuclear degradation.
Figure 2.
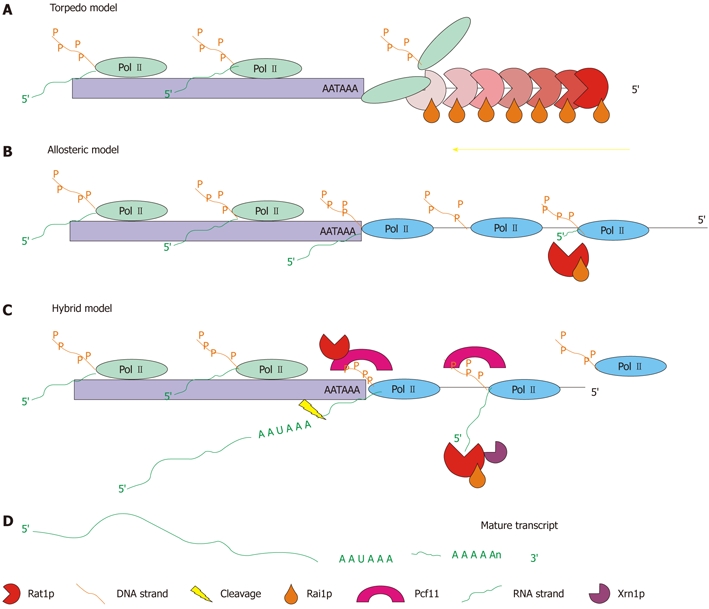
Schematic representation of the three possible modes of operation for the 5′→3′ exonuclease pathway. A: Torpedo model-the Rat1p/Rai1p complex acts as a wedge to torpedo the polymerase from the DNA strand and terminate transcription[53,54]; B: Allosteric model-the positioning element for the poly(A) site (AATAAA) causes a change in the conformation of the polymerase causing a gradual stop in transcription. The Rat1p/Rai1p complex is used to eliminate RNA downstream of the poly(A) site[55]; C: Hybrid model-the positioning element for the poly(A) site causes a change in the conformation of the polymerase, while Rat1p and Pcf11 also cause a pause in transcription. Termination is gradual but faster than that in the allosteric model. Rat1p/Rai1p are still utilized to remove RNA transcribed downstream of the poly(A) site (adapted from[56]); D: The end result of each model is a mature transcript.
Both Rat1p and Xrn1p are 5′→3′ exoribonucleases which processively degrade RNA containing a 5′-monophosphate[52]. Rat1p has also been implicated in termination of RNA Pol I[57], Pol II[58] and Pol III transcripts[58,59]. Rat1p requires a cofactor, Rai1p, to enhance its activity[50]. The Rat1p/Rai1p complex is responsible for both binding and degrading the RNA transcript once the polymerase has progressed past the poly(A) site on the template strand. When the Rat1p/Rai1p complex catches up with the polymerase, transcription is terminated. It has been suggested that the Rat1p/Rai1p complex is able to jettison the polymerase, but evidence for this action has not been found[57].
The Rat1p/Rai1p complex is involved in Pol I pre-rRNA transcription and co-transcriptional cleavage by Rnt1p, an endonuclease. Rnt1p generates a loading site for the Rat1p/Rai1p complex. In the absence of the Rat1p/Rai1p complex, pre-rRNA transcription terminates predominantly at the “fail safe” (T2) site of termination rather than the primary termination site (T1), located approximately 93 nucleotides downstream from the 3′ end of the 25S sequence[60]. Rat1p is also responsible for 5′ maturation of 25S rRNA. Depletion causes an accumulation of 26S rRNA, implicating Rat1p in cleavage at the C2 site in ITS2 (Figure 1)[57]. Rat1p is required for proper maturation of 5.8S rRNA. The predominant form of 5.8S rRNA is in the short form or 5.8SS. In the absence of Rat1p the predominant form is the 5.8SL form, indicating that Rat1p is involved in exonucleolytic cleavage at the B1s site (Figure 1). Depletion of Rai1p exacerbates the accumulation of 5.8SL[57]. The physiological relevance of the 5.8SS and 5.8SL forms is unknown. Rat1p also interacts with Rrp17p, a second 5′→3′ exonuclease, during 5′ digestion of both 5.8S and 25S rRNA. Rrp17p binds to late pre-60S ribosomal subunits, accompanying them from the nucleolus to the nuclear periphery[61].
The Rat1p/Rai1p complex has also been implicated in proper termination of Pol II pre-mRNA transcription. Rat1p interacts with the largest subunit of RNA Pol II (RBP1), and through this interaction is able to promote transcription termination upstream of the polyadenylation site. Additionally, Rat1p is responsible for 5′→3′ degradation of uncapped mRNA[54]. When functioning in Pol II transcription, Rat1p requires additional cofactors for binding and termination. Rtt103 is bound to the CTD of Pol II when serine 2 is phosphorylated. Serine 2 phosphorylation takes place following early elongation and is a known binding site of termination factors. The Rat1p/Rai1p complex co-purifies with Rtt103p and this interaction is thought to be critical for recruitment of Rat1p to 3′ end cleavage sites in gene terminator regions[58]. Rat1p and Rai1p also interact with Pcf11p, a 3′ end processing factor known for dismantling Pol II. This interaction may be critical to the termination function of Rat1p[62].
Rat1p is the major component of the rapid tRNA decay (RTD) pathway that degrades mis-modified tRNA. In the RTD pathway Rat1p interacts with Met22p, a 3′(2′)5′-bisphosphate nucleotidase. The RTD pathway is responsible for degradation of tRNAVal(AAC) lacking 7-methylguanine 46 (m7G46) and 5-methylcytosine 49 (m5C49) modifications, as well as tRNASer(UGA)(CGA) and tRNALeu(GAG) which are mis-modified[63]. In summary, the 5′→3′ nuclear RNA processing and degradation pathway is the not the predominant pathway for nuclear RNA degradation. However, the 5′→3′ pathway does play a critical role in RNA Pol I, II, and III transcription termination, transcript processing, and degradation of aberrant transcripts.
3′→5′ nuclear RNA processing and degradation by the exosome
The exosome is a complex comprising 10 components as well as a variety of cofactors. The central core of the exosome contains six RNase PH homologs that form a ring structure. These six proteins (Rrp41p, Rrp42p, Rrp43p, Rrp45p, Rrp46p, and Mtr3p) are catalytically inactive and act as a scaffold for exosome activity[3]. The six central core proteins are structurally related to the exonucleases found in both bacteria (PNPase or degradosome), and Archaea (aExosome). The bacterial PNPase is a homotrimer. Each monomer is a single polypeptide whose sequence contains two tandem RNase PH domains linked by an α domain. These two domains are followed by a K-homology (KH) and an S1 RNA binding domain[64]. The monomers come together to form a ring structure able to accommodate only single-stranded RNA (ssRNA) (Figure 3A). The archaeal exosome is a trimer of heterodimers made up of aRrp41p and aRrp42p, which both contain RNase PH domains. The trimer forms a ring structure. aRrp41p is the catalytically active subunit and aRrp42p contributes to RNA binding. RNA binding is facilitated by a “cap” ring composed of aRrp4p and aCsl4p. This ring is located on the face of the ring opposite the active sites of aRrp41p[65] (Figure 3B). The heterotrimer of dimers creates a pore in the center which contains three phosphorolytic active sites. This pore is 8-10 Å in width, capable of accommodating only ssRNA[66,67]. The structure of the archaeal exosome indicates that the ring structure interacts with only the backbone of the RNA. Therefore, interactions of the RNA with the pore do not confer substrate specificity[68]. In the yeast exosome Rrp41p, Rrp46p and Mtr3p are the aRrp41p-like subunits and Rrp42p, Rrp43p and Rrp46p are the aRrp42p-like subunits. The yeast exosome core is formed by intracomplex interactions between Rrp43p-Rrp46p[69], Rrp43-Rrp45[70], Rrp41p-Rr45p, and Rrp42p-Mtr3p[71] (Figure 3C). Each of these subunits is required for viability in yeast[72]. In the yeast exosome, none of these subunits are catalytically active. The primary function of the exosome core is to serve as scaffolding, allowing the exosome structure to be built upon its core. Like the archaeal exosome, the ring still forms a pore that can only accommodate ssRNA. However, a feature unique to the eukaryotic exosomes is that the exosome core acts to regulate the efficiency of the exonuclease Rrp44p.
Figure 3.
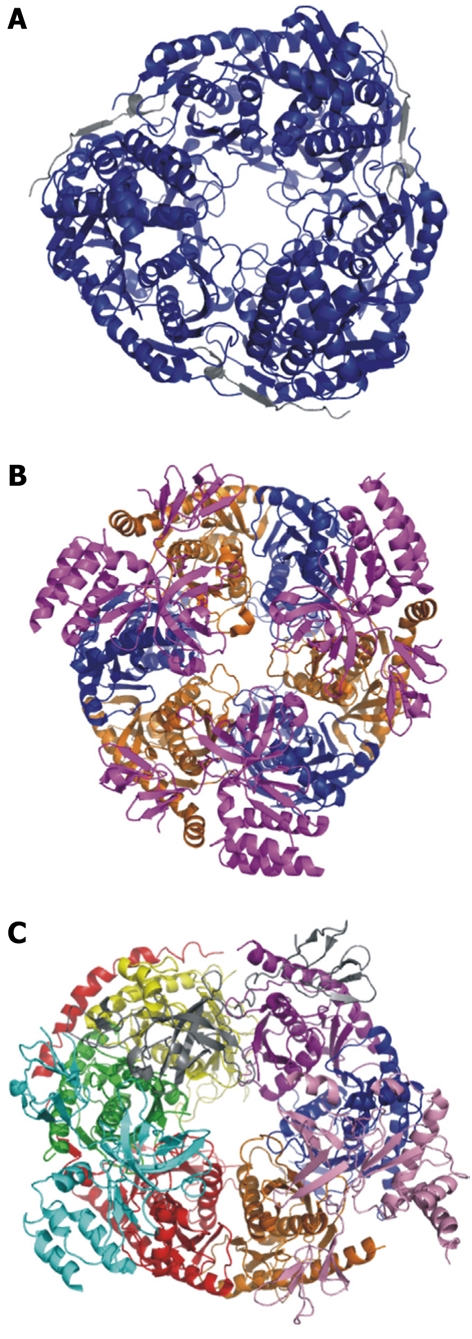
Crystal structures of the bacterial PNPase, archaeal exosome, and human exosome. A: Escherichia coli PNPase. Pictured in blue is the polynucleotide nucleotidyltransferase domain. In grey is the associated RNAse E domain (RCSB # 3GCM). The S1 and KH domains are not pictured because they are not included in the crystallized complex; B: Archaeal exosome. In blue is aRrp42, in orange is aRrp41. These two proteins are the exonucleolytic core proteins. In magenta is the aRrp4 cap protein (RCSBID: 2BA0); C: Human exosome. In orange is Rrp41p, in Blue is Rrp42p, in yellow is Rrp43p, in red is Rrp45p, in green is Rrp46p, in purple is Mtr3p. These six proteins make up the scaffolding ring structure. Also pictured are the cap proteins; in pink is Rrp4, in cyan is Rrp40, and in grey is Csl4 (RCSBID: 2NN6). The S. cervisiae exosome has not been crystalized but the human and yeast exosomes have approximately 51% sequence identity[51,66,143].
The top side of the scaffold ring interacts with three putative RNA binding proteins: Rrp4p, Rrp40p, and Csl4p[4]. These three proteins form a cap ring on the scaffold ring structure (Figure 3C). This cap structure also resembles the degradation machinery in bacteria and archaea. In the PNPase the monomers contain two RNA binding motifs[64]. In archaea the exosome contains a cap structure heterotrimer of aRrp4p or aCsl4p, both of which are RNase II-like RNA binding proteins. aRrp4p contains a KH RNA binding motif and aCsl4p contains an S1 RNA-binding motif and is a zinc finger protein[65]. The presence of RNA binding domains in the cap binding proteins is believed to facilitate substrate recognition and specificity[73]. Specifically, poly(A)-deficient RNA is degraded most efficiently by an exosome capped with aCsl4p. An exosome capped with aRrp4p is able to degrade poly(A)-tailed substrates[74]. The presence of aRrp4 significantly increases the affinity of the exosome for long RNA stretches[74,75]. The cap ring secures the RNA, while the three active sites in the archaeal core cleave the RNA. The archaeal exosome is highly processive, likely due to the RNA binding close to the active sites[76]. In the yeast exosome, the cap is a heterotrimer. Rrp4p interacts with Rrp41p, Mtr3p, Rrp44p, and Rrp6p[70], but does not bind RNA[77]. The association of Csl4p to the exosome is salt dependent. Csl4p is shown to be in sub-stoichiometric ratios to the other components of the exosome as determined by electron microscopy (EM). This observation shows Csl4p to be dispensable for the structural integrity of the exosome, but does not preclude it as being necessary for cofactor recruitment[78]. We have tested Csl4p for RNA binding activity and found that protein unable to bind RNA (Bernstein, unpublished data). Csl4p has been shown to interact with Mtr3p and is proposed to stabilize the heterodimers of the exosome core ring structure (Figure 3C).
On the bottom of the core ring structure is an active 3′→5′ exonuclease Dis3p (Rrp44p), which is responsible for some of the activity of the exosome[3-5]. Rrp44p is a member of the RNase RNR superfamily of 3′ exonucleases and is most closely related to RNase II and RNase R[76]. The domain structure of Rrp44p includes an amino terminal PilusT N-terminal (PIN) domain followed by a loop region that connects to two cold shock domains (CSDs), RNA binding (RNB) domain, and a C-terminal S1 domain (Figure 4A). The PIN domain is exclusive to Rrp44p and homologs of this protein. This domain provides Mn2+-dependent endonuclease activity to Rrp44p. The two CSDs and the S1 domain are each RNB domains. The RNB domain includes the catalytic site for exonuclease activity[79]. Rrp44p anchors to the exosome core through interactions with Rrp45p, Rrp43p and Rrp41p. The CSDs interact primarily with Rrp45p and to a lesser extent with Rrp43p, creating a direct route for RNA to pass from the exosome to Rrp44p. The “head domain,” or amino-terminal domain (NTD) of Rrp44p interacts with Rrp41p[78]. Each of the domains of Rrp44p is functionally separate[80]. Inactivating both the endonuclease and exonuclease activity of Rrp44p is lethal[79]. EM images of Rrp44p interacting with the core exosome show that RNA could enter the Rrp44p active sites in two ways: (1) through the exosome core, when at least 7 nt of ssRNA are present; or (2) directly into Rrp44p[76,78] (Figure 4B). The RNA recruitment channel of Rrp44p opens towards the exosome core, suggesting that RNA is “handed over” from the exosome to Rrp44p for processing or degradation[78]. At least 9 nt of ssRNA need to be available to thread through the pore of the exosome to Rrp44p[78]. Although the RNA only needs to be 9 nt in length to reach Rrp44p, 29-32 nt are required for activity[81]. Hand over delivery utilizes this property of the exosome to help select substrates. In the direct method, Rrp44p receives RNA through the cleft between the PIN domain and the other domains of Rrp44p. The PIN domain is located approximately 20 Å from the recruitment channel and may block some RNA from entering the catalytic site. The cleft that is created between the PIN domain and the rest of Rrp44p could easily accommodate ssRNA, and could fit double stranded (dsRNA) RNA, but any RNA with tertiary structures would be excluded. The position of the PIN domain is thought to provide a mechanism of selecting targets for processing and/or degradation[78]. Rrp44p is known to discriminate between substrates, showing specificity for hypomodified tRNAimet. This recognition is dependent upon amino acids in the recruitment channel. Even though Rrp44p can recognize hypomodified tRNAimet, full degradation of tRNA requires the exosome and Mtr4p[80].
Figure 4.

Rrp44p. A: Crystal structure of Saccharomyces cerevisiae Rrp44p bound to RNA. CSD1 is colored red, CSD2 is colored teal, the RNB domain is blue, and the S1 domain is magenta. The PIN domain is not pictured because this domain is not included in the crystallized construct (RCSB # 2VNU); B: Schematic representation of the two mechanisms whereby Rrp44p is able to degrade RNA. In Scheme 1 the RNA is fed through the exosome to Rrp44p. Scheme 2 shows the RNA being degraded directly by Rrp44p[4,144].
In addition to Rrp44p, a second active exonuclease named Rrp6p[6,7] interacts with the exosome to promote the processing and degradation of RNAs. Unlike Rrp44p, Rrp6p associates exclusively with the nuclear exosome. Rrp6p is a member of death effector domain containing protein (DEDD) family of 3′→5′ exonucleases[82]. Catalysis by Rrp6p requires two divalent metal ions[83,84]. The domain structure of Rrp6p includes an NTD, an exonuclease domain, and a helicase and RNase D C-terminal (HRDC) domain[76]. The NTD includes a polycystin 2 N-terminus (PC2NT) domain, which creates a binding site for Rrp47p, a cofactor of the exosome. Rrp47p promotes the catalytic activity of Rrp6p as well as allowing Rrp6p to bind RNA with secondary structures[85]. The exonuclease domain, located in the NTD, is responsible for the catalytic activity of Rrp6p[86]. The HRDC domain provides Rrp6p with a potential helicase domain, an RNase D domain, and is responsible for the RNA binding capabilities of Rrp6p[76] (Figure 5).
Figure 5.

Rrp6p. A: Crystal structure of Saccharomyces cerevisiae Rrp6p. The NTD is colored yellow, the exonuclease domain is colored green, and the HRDC domain is colored purple (RCSBID: 2HBJ); B: Schematic representation of the two different ways that Rrp6p functions in RNA degradation and processing. Scheme 1: Rrp6p interacts with the nuclear exosome to facilitate processing or degradation; Scheme 2: Rrp6p interacts with Rrp47 allowing Rrp6p to bind to RNA with secondary structure[145].
The CTD of Rrp6p interacts with the exosome[7] and might contribute to substrate preference[76]. Although Rrp6p is not required for yeast viability[87], a loss in the protein causes slow growth, temperature sensitive lethality, and RNA synthesis defects[85]. Rrp6p is implicated in processing of 3′-extended 5.8S rRNA. Processing intermediates including 23S, 21S pre-rRNAs and unadenylated rRNA fragments from the D-BIL cleavage sites accumulate in the absence of Rrp6p. These intermediates are degraded by Rrp6p and Rat1p, independent of the exosome[88]. In addition, Rrp6p is implicated in the 3′ maturation events of 5S rRNA[76], maturation of U14 snRNA, Nop1 snoRNP[89], and many other 3′-extended and polyadenylated snRNAs and box C/D snoRNAs[85]. Maturation of RNase P RNA, and mRNA surveillance are also dependent upon Rrp6p[76]. Rrp6p is involved in mRNA quality control by degradation of transcripts at the site of transcription, a surveillance activity that requires poly(A) polymerase and the mRNA cap-binding complex[85,87,90,91]. Rrp6p is also involved in the degradation of CUTs following polyadenylation[42,89].
In summary, the exosome is responsible for 3′→5′ processing and degradation of multiple RNAs. The ability of the exosome to process or degrade RNA is dependent on association of a substrate specific cofactor to enhance activity, and the helicase activity of Mtr4p to remove secondary structure in substrates. Together, each of these components comes together to create an efficient pathway for processing and degradation[76,80] (Figure 6).
Figure 6.

Schematic representation of the basic mode of operation of the 3′→5′ RNA exosome. The figure highlights the need for a cofactor to stimulate the exonuclease activity of the exosome and the need for an RNA helicase to remove secondary structure to allow proper processing or degradation. The figure shows a representative stem loop structured RNA but any RNA with secondary structure could undergo the same remodeling to complete processing or degradation. The cofactor shown is a representative of all known and unknown cofactors. The exosome shows weak exonuclease activity in vitro, yet rapid degradation is seen in vivo, indicating that cofactors are required for this activity[76,80].
COFACTORS OF THE EXOSOME
The exosome is influenced by a series of cofactors that presumably promote specificity or target the exosome to a particular site of processing or degradation. The current theory is that processing is the primary mode of action of the exosome[76]. The exosome has weak exonuclease activity in vitro but rapid degradation is seen in vivo. This suggests that cofactors are required to stimulate activity in the presence of targets[92].
Rrp47p
Rrp47p associates with the exosome at sub-stoichiometric levels and association is dependent on low Mg2+ concentrations[93]. Early steps in processing rRNA and some snoRNAs by the exosome require an interaction with Rrp47p. In particular, Rrp47p participates in exosome-mediated processing of 35S, 23S, and 21S rRNA[93]. Rrp47p also interacts directly with the N terminus of Rrp6p through the PC2NT domain in the absence of the exosome[85]. Specifically, the interaction of Rrp47p with Rrp6p allows Rrp6p to bind structured RNAs including double-stranded RNA and tRNAPhe[85]. Rrp47p is involved in 3′-end processing of snoRNAs, but is not required for trimming the final few nucleotides. Processing of U4 and U5 snRNAs are also dependent on Rrp47p and Rrp6p. Although Rrp47p is involved in many of the same processing events as Rrp6p, there is no evidence that Rrp47p itself is an exonuclease. The level of Rrp47p protein in the cell is highly dependent on the presence of Rrp6p, but Rrp6p protein levels do not change significantly if Rrp47p is deleted[85]. Rrp47p is also not required for cell vitality or association of Rrp6p with the exosome[93].
Mpp6p
Mpp6p is a nuclear RNA binding protein which interacts with the exosome, Rrp44p, Rrp47p, Nrd1p, TRAMP and Rrp6p. Mpp6p is non-essential yet robustly interacts with the exosome in an RNA-independent manner. Interestingly, loss of Mpp6p is lethal in combination with loss of either Rrp47p or Rrp6p, indicating that these proteins function in similar steps of processing or degradation. Mpp6p is known to function in surveillance of pre-rRNA, which is the most likely site of interaction with Rrp47p and Rrp6p. In particular, Mpp6p is involved in processing of 5.8S+30 pre-rRNA and, to a lesser extent, 23S pre-rRNA, in addition to some other uncharacterized processing intermediates. Although Mpp6p and Rrp47p both interact with Rrp6p, each is believed to serve a different purpose. The specificity of these proteins may come from the different RNA preferences of the proteins[94]. For example, Mpp6p shows a preference for poly(U) stretches, whereas Rrp47 binds preferentially to structured RNAs. In addition to a role in pre-rRNA processing, Mpp6p is involved in pre-mRNA and mRNA surveillance. Degradation of CUTs is facilitated by targeting via Mpp6p, Rrp47p, TRAMP and Nrd1p/Nab3p to the exosome and Rrp6p[94].
Nrd1p/Nab3p
The Nrd1p/Nab3p complex interacts with the exosome to promote proper 3′ end formation of several mRNAs[95], snRNAs[23,24,86,95], snoRNAs[95] and CUTs[48,49]. Nrd1p is an RNA binding protein that interacts with the CTD of Pol II via its NTD[96]. Nrd1p works in association with another RNA binding protein, Nab3p. Nab3p forms a direct interaction with Ctk1p[97], a catalytic subunit of a kinase that phosphorylates the CTD of Pol II[98]. In order to act efficiently on the 3′ ends of RNA substrates, the Nrd1p complex also requires the putative helicase Sen1p[97,99]. The final component known to be required for the Nrd1p complex is Ess1p, a prolyl isomerase. Ess1p binds to the CTD of Pol II and regulates transcription by causing conformational changes by isomerization of serine-proline bonds in the CTD[100]. The Nrd1p complex and all of its cofactors are responsible for directing transcription termination of non-polyadenylated transcripts[95]. These transcripts can then be targeted by the Nrd1p complex to TRAMP for polyadenylation and the exosome for processing or degradation[48]. In the presence of CUTs the Nrd1p complex is required for proper 3′ end formation[49] (Figure 7).
Figure 7.
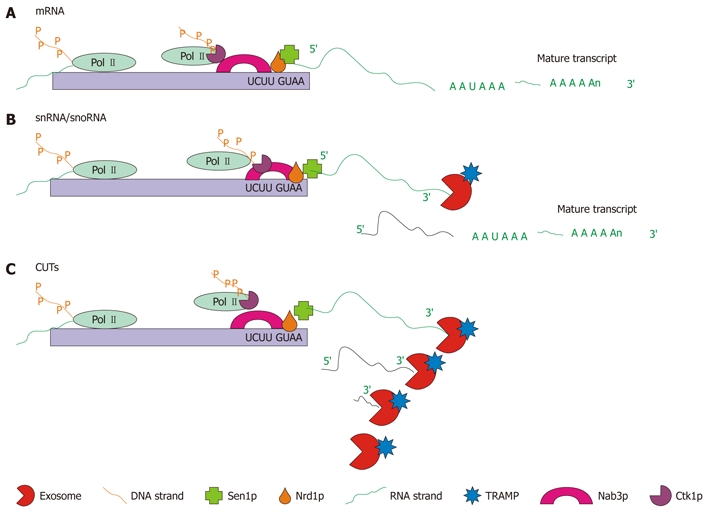
Schematic representation of the three types of Nrd1p/Nab3p-dependent RNA maturation. A: mRNA 3′ end trimming: for some mRNAs, the Nrd1p complex is required to complete transcription. Pictured is the Nrd1p complex associated with Pol II bound to the Nrd1p and Nab3p binding sites, which cause the Pol II molecule to lift off of the DNA and release the RNA. At this point a poly(A) polymerase (not shown) polyadenylates the end of the strand producing the mature mRNA; B: snRNA/snoRNA 3′ end trimming: snRNA and snoRNA, which are transcribed from autonomous transcription units, are terminated by the Nrd1p complex. Pictured is the Nrd1p complex which causes transcription termination. Following polyadenylation by the TRAMP complex and 3′ end trimming by the exosome, the mature transcript is formed; C: Cryptic unstable transcript (CUT) degradation: transcription of CUTs is also terminated by the Nrd1p complex. Pictured is the Nrd1p complex interacting with Pol II causing transcription termination. Following termination the transcript is degraded by the exosome in conjunction with the TRAMP complex[86,95].
Nop53p
Another exosome cofactor is Nop53p, which interacts with Rrp6p[101], Trf4p[101,102], and several rRNA assembly proteins[103]. Although no direct interaction between Nop53p and the exosome has been found, depletion of Nop53p causes functional defects in the exosome[101,104]. In addition, the defects seen in rRNA processing by depletion of Nop53p are much like those seen upon exosome depletion[101]. Nop53p is involved in the processing of 7S to 5.8S and 27S to 25S rRNA[101-104]. Nop53p interacts with these pre-rRNAs co-transcriptionally during 60S ribosome biogenesis[101]. Specifically, Nop53p associates with unassembled 60S ribosome subunits while they are located within the nucleolus and nucleoplasm[103-105]. Nop53p is a late-acting factor in 60S ribosome biogenesis, which is required for 60S ribosome subunit export competency[103,105]. The requirement of Nop53p for export has led to the assumption that Nop53p is involved in targeting aberrant pre-ribosomes for degradation by the exosome. This assumption is enhanced by the fact that polyadenylated pre-rRNA precursors of 7S and 27S pre-rRNAs are accumulated in the absence of Nop53p[101]. This indicates that these rRNAs have been tagged for degradation but are unable to be targeted to the degradation machinery due to the absence of Nop53p[105].
THE TRAMP COMPLEX
The major cofactor of the nuclear exosome is the TRAMP complex. The TRAMP complex was discovered using a yeast two-hybrid screen with Mtr4p as bait. This screen identified both Trf4p and Trf5p poly(A) polymerases as potential Mtr4p-interacting proteins. Further experiments confirmed that Trf4/5p interact with Mtr4p with a minimal site at amino acids (aa) 53-199 of Mtr4p. The third component of the TRAMP complex was found by an interaction with Lsm2p and Air2p. The immunoprecipitates containing this complex also contain Trf4p and Mtr4p[92]. Due to these sets of interactions, the TRAMP complex was named for the three proteins that comprise the complex; Trf4/5p, the poly(A) polymerases, Air1/2p, putative RNA binding proteins that contain a zinc ring finger domain, and Mtr4p, a nuclear RNA helicase[92]. Together, these three proteins form two distinct complexes: the TRAMP4 complex which includes Trf4p, Air1/2p and Mtr4p, and the TRAMP5 complex with Trf5p, Air1/2p, and Mtr4p. Each complex is competent for polyadenylation of various pre-rRNA[42,65,92,106-108], snRNA[42,92,109,110], snoRNA[108,109], tRNA[40,41,92,111], CUTs[42,108,109], and mRNA[112], as well as stimulating degradation by Rrp6p[42,106,107,109-111] and the exosome. The TRAMP4 complex is believed to be approximately 3-fold more prevalent than TRAMP5. Each complex is responsible for distinct surveillance and degradation activities. The complexes are functionally redundant in only select circumstances. The Trf4/5p and Air1/2p proteins are found in stoichiometric ratios in the cell[42]. Additionally, only a fraction of the Mtr4p in the cell is found in a TRAMP complex. Mtr4p can be removed from the TRAMP complex by a relatively moderate increase in ionic strength, disassociating at 500 mmol/L NaCl in vitro. This suggests that Mtr4p also has TRAMP-independent functions. Each of the TRAMP complexes has been found to be responsible for the surveillance and degradation of RNAs, but not for their maturation[92].
The TRAMP complex is required for 3′ end trimming of NAB2 mRNA[112], polyadenylation of hypo-modified tRNA[40,41,111], stimulation of the exonuclease activity of both Rrp6p and the exosome[92,106], and surveillance and degradation of various RNAs[92]. Nab2p is a nuclear protein required for mRNA export and poly(A) tail length determination. Due to its role in export of mRNA, Nab2p must be tightly regulated to avoid export of improper transcripts. Nab2p is responsible for regulating the transcription of its own mRNA (NAB2). Regulation is facilitated by a competition between proper 3′ end trimming and polyadenylation. The 3′ end of NAB2 mRNA contains a stretch of 26 adenosines (A26 site) and approximately 114 nt further downstream is a polyadenylation site. The balance between which of these sites is utilized for 3′ end formation determines the level of NAB2 transcripts. Nab2p binds to the A26 site of its own mRNA, and thereby recruits TRAMP and the exosome to degrade the transcript. The exosome and TRAMP are responsible for trimming the NAB2 mRNA back to the polyadenylation site, at which point either polyadenylation can occur or the transcript can be rapidly degraded by Rrp6p and TRAMP. Degradation by the exosome is caused by defects in 3′ end formation, and the rate of degradation is enhanced by TRAMP[112]. TRAMP is also involved in polyadenylating misfolded tRNA. Unmodified tRNAs are preferentially polyadenylated by TRAMP4. This indicates that TRAMP is able to differentiate between modified and unmodified tRNA, likely due to misfolding of the unmodified tRNA. TRAMP4 polyadenylates these misfolded tRNAs, utilizing the poly(A) polymerase activity of Trf4p. Trf4p is stimulated by Air1/2p and requires the presence of rATP, Mn2+ or Mg2+, and an ssRNA stretch. Polyadenylation targets these tRNAs to the exosome for degradation. The exosome is able to degrade the poly(A) tail and the single-stranded stretch but requires Mtr4p for degradation of the tRNA body[111]. Specifically, TRAMP is able to stimulate Rrp44p to degrade hypomodified tRNAiMet[40,41]. TRAMP has also been implicated in the polyadenylation and degradation of rRNA, snoRNA and CUTSs. In particular, TRAMP is known to polyadenylate 23S pre-rRNA and U14 snoRNA[92]. In addition to the roles of TRAMP in specific cases of RNA surveillance and degradation, the complex has a global role in stimulating the exonuclease activity of the exosome and Rrp6p. In the presence of TRAMP, the rate of Rrp6p hydrolytic exonuclease activity is increased approximately 10-fold. The enhancement of Rrp6p activity by TRAMP is independent of rATP and polyadenylation. TRAMP is also able to enhance the exonuclease activity of the exosome containing Rrp6p to degrade aberrant RNA[106] (Figure 8). Each of the proteins within the TRAMP complex also has functions that are unrelated to the complex which will be discussed further below.
Figure 8.
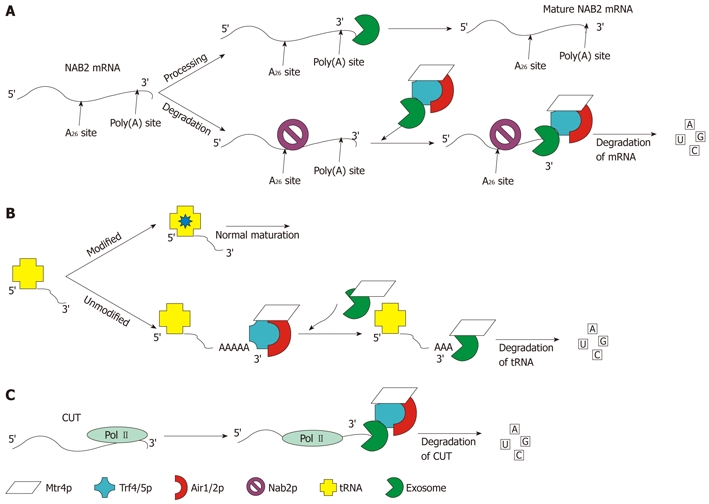
Schematic representation of the three well-characterized functions of the TRAMP4/5 complex. A: Processing and degradation of NAB2 mRNA: the level of NAB2 mRNA is controlled by Nab2p, which recruits TRAMP and the exosome by binding to the A26 site of the mRNA. Once TRAMP and the exosome are recruited the transcript is degraded; B: Degradation of un-modified/misfolded tRNA: tRNA that has not undergone modification at the correct time is preferentially polyadenylated by TRAMP. Once polyadenylated, the exosome aided by Mtr4p degrades the tRNA; C: ncRNA degradation: shown in panel C is the degradation of CUTs. The same pathway is followed for snRNA and snoRNA, which are processed and degraded by TRAMP and the exosome. Degradation of CUTs is performed co-transcriptionally as pictured. The levels of these RNAs cannot be detected without depletion of Rrp6p or Trf4p[92,111,112].
A recent structural study[113] has revealed the molecular architecture of the TRAMP4 core polyadenylation machinery (Figure 9). The complex, which contains the catalytic and central domains of Trf4p (residues 161-481; full-length Trf4p contains 584 residues) and the fourth and fifth zinc knuckles of Air2p (residues 119-198; full-length Air2p contains 344 residues), shows that the fifth zinc knuckle serves as a protein interaction module, as the residues most likely to interact with RNA are buried against the surface of Trf4p. In contrast, the fourth zinc knuckle has these same putative RNA-binding residues exposed, making this zinc knuckle a likely RNA-binding module. The TRAMP4 core (Trf4p161-481 plus Air2p119-198) can recognize and polyadenylate mutant tRNAAla[113], in agreement with the hypothesis that the fourth zinc knuckle plays a role in substrate binding. In addition, the first zinc knuckle plays a significant role in TRAMP4 activity. A fragment containing this zinc knuckle polyadenylated a greater fraction of mutant tRNAAla and appended longer poly(A) tails on those substrates than fragments that lacked the first zinc knuckle[113]. TRAMP requires a minimum 3′ overhang of 3 nt to initiate polyadenylation, but beyond that requirement, little is known regarding how TRAMP differentiates between substrate and non-substrate RNAs.
Figure 9.
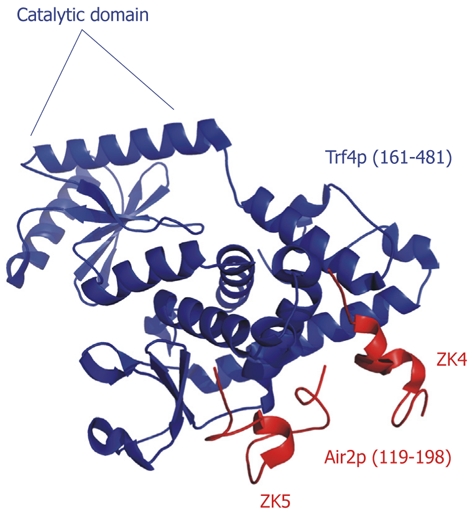
Crystal structure of the TRAMP4 core. The catalytic domain of Trf4p is indicated. The fourth and fifth zinc knuckles (ZK4 and ZK5) pack against the central domain of Trf4p (RCSBID: 3NYB)[113].
Trf4p and Trf5p
The Trf4p and Trf5p proteins are approximately 56% similar at the amino acid level[108] and 48% identical[110]. The two proteins have proven to be functionally redundant for the polyadenylation and degradation of some 3′ extended RNAs, including U14 snoRNA, 23S pre-rRNA, and the NEL025c CUT[108]. It is likely that Trf4p and Trf5p are responsible for promoting exosome degradation of other CUTs, including spliced introns[108]. This function is carried out by a polyadenylation independent mechanism. With the exception of these few examples, it is believed that Trf4p and Trf5p target distinct RNA populations. A recent study[108] has investigated the effects of depleting either Trf4p or Trf5p individually. Depletion of Trf4p caused 422 RNAs to accumulate, 72 of which were snoRNAs, intergenic RNA regions, autonomously replicating sequences or CUTs. The second largest population consisted of Ty1 retrotransposon transcripts. Ty1 retrotransposons are a class of RNAs that encode proteins and RNAs that assemble into virus-like particles. Depletion of Trf4p also results in accumulation of mRNAs encoding nuclear proteins. It appears that highly expressed and structured RNAs depend on the polyadenylation activity of Trf4p to be properly processed; this only accounts for approximately 10% of the Trf4p targets. One possibility is that the majority of Trf4p target RNAs recruit other factors that are required for their processing. Perhaps Trf4p catalytic activity is not essential for the proper processing of these targets, but helps ensure that they are processed in a timely manner. Trf4p is also required for degradation of subtelomeric RNAs, particularly those containing a repetitive helicase-encoding sequence called the Y′ sequence within terminal telomeric repeats. The polyadenylation activity of Trf4p is not required for this function but is beneficial. In the absence of Trf4p, telomeres are approximately 120 bp shorter than normal. This shortening is not related to the accumulation of subtelomeric RNA[108].
Depletion of Trf5p resulted in the accumulation of only 269 RNAs, of which 11 were ncRNA, one was a snoRNA, one was a CUT, and two were intergenic regions (IGRs). Trf5p depletion causes a slight decrease in Ty1 retrotransposon. The opposing effects of Trf4p and Trf5p on Ty1 levels indicate that they act via an unknown mechanism to regulate expression of the TY1 locus. The majority of the RNAs affected by Trf5p depletion are mRNAs that encode cytoplasmic translation proteins. Trf5p is involved in degradation of subtelomeric RNAs, by acting on the factors that regulate chromatin silencing[108].
In addition to roles in RNA surveillance and degradation, Trf4p and Trf5p are also critical factors in genome stability through interactions with transcripts encoding DNA replication-dependent histones. In order to maintain histone levels, cells regulate their expression by coupling transcription with the rate of DNA synthesis[114]. Trf4p and Trf5p bind to chromatin to facilitate maintenance of histone levels. Depletion of Trf4p causes defects in DNA metabolism including: hyper-recombination in rDNA; sensitivity to DNA-damaging agents[115-119]; chromosome condensation; cohesion defects[116,120,121]; and delay of entry into and progression of S-phase[122]. Trf4p interacts with various proteins involved in histone acetylation[122], DNA-damage checkpoints, histone protein regulation[123], histone chaperones, and nucleosome-assembly factors[124,125]. Trf5p interacts with Pol ε, a replicative DNA polymerase[126]. Although Trf4p and Trf5p are involved in maintenance of the levels of replication-dependent core histone mRNA, they are not responsible for the polyadenylation of those mRNAs[122].
Trf4p has been shown to possess other functions independent of Trf5p. Trf4p is involved in the degradation of Pol II CUTs and other Pol II and Pol III ncRNA transcripts, as well as the polyadenylation of Rnt1p cleavage products. In concert with Rrp6p, these polyadenylated transcripts, U3/U4 snoRNA, and snR40 snRNA are degraded[109]. Pre-ribosomes that have undergone surveillance and require degradation are polyadenylated by Trf4p and concentrated in the No-body, a sub-nucleolar region. Trf4p and Rrp6p are required for concentration of export blocked 60S pre-ribosomes in the No-body[107]. Trf4p is implicated in targeting of RNAs for degradation by the exosome via a mechanism that would limit the levels of inappropriate Pol-II-transcribed IGRs[42].
Trf5p also has functions distinct from those of Trf4p. Trf5p is the poly(A) polymerase that targets 5.8S+30 pre-rRNA[110], and 23S rRNA[127]. Mis-assembled pre-rRNAs that become the RNA component of the small ribosomal subunit are also polyadenylated by Trf5p and then degraded by the exosome. Mis-assembly is determined by the kinetics of assembly of the pre-rRNA processing complexes on the 5′ terminal end[127].
In summary, both Trf4p and Trf5p are extremely important for the correct surveillance and processing of a variety of RNAs and pre-RNAs. These two proteins interact closely with Rrp6p and the nuclear exosome to degrade RNAs that have been polyadenylated. Direct binding of Trf4/5p to Air1/2p is often seen during polyadenylation of substrate RNAs[42,128].
Air1p and Air2p
The Air1p and Air2p proteins were discovered in a two-hybrid screen for novel factors in yeast that interact with arginine rich domains (RGGs) of heterogeneous nuclear RNPs (hnRNPs). Air1p was found to interact with Hmt1p, a protein that methylates Npl3p, thereby allowing Npl3p to exit the nucleus[129]. hnRNPs are involved in mRNA processing and export. They are post-translationally modified by methylation at arginine residues within the RGG domain[130]. Methyl groups are donated by SAM and placed on the nitrogen group of an arginine residue[131]. Air1p binds to Hmt1p, thereby blocking the ability of Hmt1p to bind and methylate Npl3p. Without methylation Npl3p cannot export mRNA from the nucleus. Following the discovery of Air1p, a genome search has found a paralog, which has been named Air2p. Air2p is also able to bind Hmt1p and block methylation of Npl3p[130]. Air1/2p were later discovered to bind the large mobile (LM) proteins in the LM2-8 complex. This interaction has led to the discovery of their role in the TRAMP complexes[132].
Mtr4p
Helicases are enzymes that catalyze the unwinding of double-stranded nucleic acids (dsNAs), and are classified into five families, named super family (SF) 1-5. Mtr4p belongs to SF2, and the subfamily of DExH-box helicases. DExH-box helicases are involved in all processes involving RNA metabolism including: transcription, editing, splicing, ribosome biogenesis, RNA export, translation, RNA turnover, and organelle expression[133]. In addition, DExH-box helicases act with cofactors that are responsible for recruitment to a complex, bridging between the helicases and the target RNA, and activation of the helicases through physical interaction, although none of these activities are mutually exclusive[134].
Mtr4p was discovered in two independent studies, each of which was searching for proteins with different functions. In the first study[135], mRNA transport proteins were depleted. Of particular interest was the depletion of Mtr4p, which caused accumulation of polyadenylated RNA in the nucleolus[135]. This study examined the conserved regions within Mtr4p that are involved in helicase activity and nucleolar targeting. The second study[10] identified Mtr4p in a screen for genes that require overexpression of yeast transcription initiation factor eIF4B protein 3 (Tif3p). That study looked at the functional role of Mtr4p in biogenesis of 60S ribosomal subunits[10].
Mtr4p is an ATP-dependent RNA helicase[41,136] localized to the nucleolus[135], and is a member of the Ski2 subfamily of RNA helicases, which is named after the yeast helicase Ski2p. Ski2 family members contain eight conserved sequence motifs that are thought to be important for helicase activity. The Ski2 family of helicases also contains a large conserved motif at the end of their CTD, the DSHCT domain[137], which is an eight-helix bundle comprising residues 912-1073[138]. The CTD of Mtr4p also contains a bipartite nuclear localization signal (NLS) and an arginine/lysine-rich domain. Although the NLS sequence located between aa 855-869 of Mtr4p is able to confer nuclear localization to β-galactosidase, this sequence is not essential for Mtr4p nuclear localization. Surprisingly, the arginine/lysine domain acts as the NLS for Mtr4p[135]. Remarkably, recent structural and functional studies have implicated residues in the CTD in RNA-binding([139,140]; discussed further below). The N-terminal half, however, contains all of the helicase motifs (Figure 10). Our studies have shown that, despite the presence of all the helicase motifs, the N-terminal half of Mtr4p is not functional. This protein fragment cannot hydrolyze ATP and has impaired RNA binding (Bernstein, unpublished data).
Figure 10.

Ski2 family helicase motifs. A multiple sequence alignment of the Ski2 family conserved helicase motifs and the C-terminal domain terminal sequence. Shown are alignments of the Ski2 family of helicases with the cytoplasmic Ski2p Saccharomyces cerevisiae helicase and Mtr4p. In each of the pictured regions, the two related helicases are highly conserved.
A 2006 study[141] has investigated the importance of the sequences in three of the helicase motifs. In this study, two mutations were made in motif I, (K177A and K177R), one mutation (D262A) in motif II, and the S293L motif III mutation was engineered. The effects of the mutations were measured by dominant negative growth defects. In a dominant negative growth defect, the mutated protein may sequester the endogenous substrate by binding to it but being unable to release it. In Mtr4p, mutations made in motifs I and III both resulted in dominant negative growth defects. Surprisingly, mutating the D of the critical ATP binding and hydrolysis DExH sequence in motif II had no effect on growth[141]. We have shown that the D262A mutant is ATPase deficient and binds RNA (Bernstein, unpublished data).
Mtr4p structures
Two recent structural studies have revealed the architecture of Mtr4p and highlighted motifs important for binding both nucleotide and RNA substrates. The structure of apo-Mtr4p[138] reveals that the canonical recA-like core domains are decorated with both winged-helix and seven-helix bundle domains (similar to those found in the related archaeal Hel308 helicase) and a novel arch domain (Figure 11). Such unique sequence and structural features of Mtr4p and related helicases likely contribute to attributes that differentiate them from other enzymes in the same superfamily. In particular, the arch domain is both unique to the Ski2-like helicases and essential for Mtr4p function. Surprisingly, the arch domain has little effect on either ATPase or unwinding activity, but is essential for proper yeast growth and 5.8S rRNA processing[138].
Figure 11.
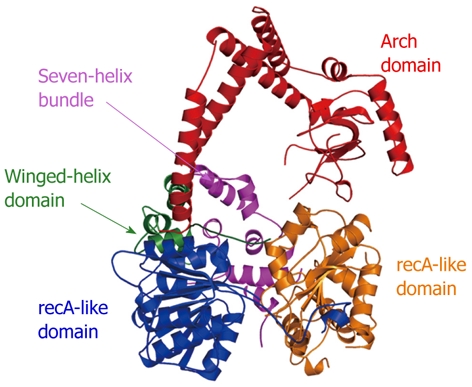
Crystal structure of apo-Mtr4p. The individual domains (recA-like, winged-helix, seven-helix bundle, and arch) are labeled and colored similarly to the scheme in[138] (RCSBID: 3L9O). The arch domain, despite being in an ideal position to promote protein interactions, does not appear to interact with the TRAMP complex[140].
The structure of Mtr4p bound to ADP and a short poly(A) RNA has also recently been solved[140]. The adenine base of ADP is sandwiched between the side chains of R547 and F148. The exocyclic amino group of adenine is recognized by Q154, which is part of the Q-motif. Both K177 of motif I and D262 of motif II are in position to interact with the γ-phosphate of bound ATP. The RNA substrate binds in a cleft between the two RecA-like domains. The 5′-most visible nucleotide (in this context, “visible” means that there is observed electron density for that nucleotide in the structure, making this position 5 in the 10 nucleotide poly(A), called A10, in the crystal) packs against W524 and G526, which form part of a β-hairpin that is similarly situated in both the archaeal Hel308 and Prp43p helicases. Several residues in the cleft between the recA-like domains make contact with the sugar-phosphate of the bound RNA. Surprisingly, despite the preferential binding of Mtr4p to poly(A) RNA, few contacts between Mtr4p and the adenine bases in the bound RNA exist. In the first of two monomers, in the asymmetric unit of the crystal structure, the 3′-end of the A10 substrate in the co-crystal abuts the interface between domains 1 and 4, with interactions between R272 and O4′ of the 3′-ribose, and between R1030 and the exocyclic amino group of the 3′ adenosine. In the second monomer, E947 and R1030 interact with the fourth and fifth visible base in the bound substrate, respectively. E947 interacts with the exocyclic amino group, and R1030 interacts with N3, a different position than that observed in the first monomer (Figure 12).
Figure 12.
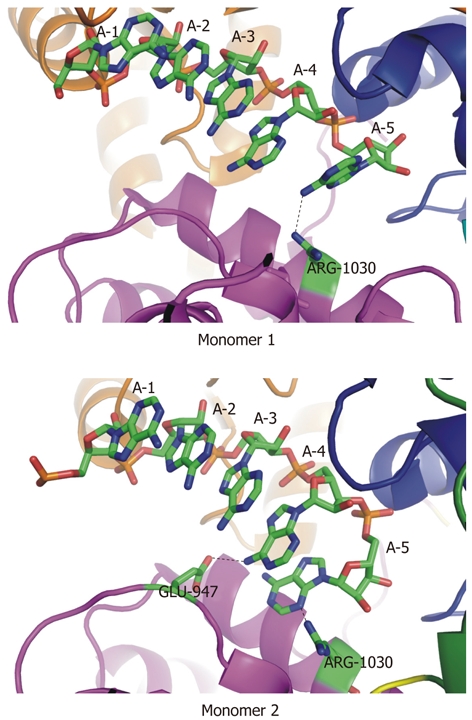
Two unique sets of contacts between Mtr4p and a short poly(A) substrate. In two independent monomers in the asymmetric unit of the crystal structure (RCSBID: 2XGJ), two different sets of interactions with adenine bases at the 3′-end of the bound RNA are observed. A: In monomer 1, R1030 interacts with the exocyclic amino group of A5; B: In monomer 2, E947 interacts with the exocyclic amino group of A4 and R1030 interacts with the N3 nitrogen of A5.
Perhaps this plasticity in the Mtr4p-poly(A) interface contributes to some of the unique functional properties observed in vitro[136,142] and in vivo[139].
Involvement of Mtr4p in RNA processing and degradation
Mtr4p is involved in a variety of RNA processing and degradation events including the processing of rRNA[7,10,141], snoRNA[23,24], snRNA[24], mRNA[35], the degradation of mis-modified tRNA[13,40,41], and degradation of CUTs[42,49]. During rRNA processing, Mtr4p is required for biogenesis of 60S ribosomal subunits. Depletion of Mtr4p leads to a decrease in the level of mature 60S ribosomal subunits. This decrease is due to the involvement of Mtr4p in processing at the A0, A1, A2, BIL, BIS, C2, and E cleavage sites[7,11] (Figure 1). Improper processing at these sites leads to accumulation of 35S, 23S, 22S, 21S, 7SL, and 7SS pre-rRNAs, and retention of the 5′ region of ITS2, A0, and ETS. During these processing events, Mtr4p interacts with the exosome but is not stably associated[10]. Although Mtr4p is not thought to be a universal factor in all snoRNA processing, it has been implicated in proper 3′ end processing of snR44, snR73, snR72, U14, and snR33[23,24]. Mtr4p activity is required for proper processing of both U4 and U5 snRNAs[23]. Mtr4p has also been implicated in degradation of mRNAs which that lack a full-length poly(A) tail or have been hyperadenylated[35,36]. Degradation of hypomodified tRNAiMet is dependent upon the helicase activity of Mtr4p[41]. In the absence of the m1A58 modification, hypomodified tRNAiMet is polyadenylated by Trf4p. After polyadenylation, the exosome is recruited to degrade the tRNA. In the absence of Mtr4p, the poly(A) tail is readily removed by the exosome but the body of the tRNA is unable to be degraded[40]. In some instances, Mtr4p is involved in the degradation of CUTs, through stimulation of the exonuclease activity of Rrp6p, as well as in the TRAMP complex[42,49]. The involvement of Mtr4p in such a wide variety of RNA processing and degradation events indicates the critical role this protein plays in cellular function. This is highlighted by the fact that deletion of the gene is lethal[10]. One essential function of Mtr4p is helicase activity; however, it is not the only function, as we discuss below.
A recent study[139] has discovered a novel Mtr4p activity as a regulator of TRAMP polyadenylation. Tight regulation of TRAMP polyadenylation likely protects against unnecessary ATP consumption and spurious polyadenylation (and subsequent exonucleolytic cleavage by the exosome). In vitro, TRAMP stimulates the accumulation of polyadenylated substrates containing very short (3-5 adenylates) poly(A) tails. This correlates quite well with the observed distribution of polyadenylated tRNAiMet
in vivo. Although the short poly(A)-tailed RNAs accumulate rapidly, they are extended quite slowly. Current structural, biochemical and biophysical data all indicate that Mtr4p interrogates the 3′-end of potential substrates. The specificity of Mtr4p for poly(A) RNA[136,142] likely facilitates this regulation once the poly(A) tail reaches a critical length of four or five adenylates. Intriguingly, residues outside the canonical helicase motifs in the conserved DSHCT domain appear to be important for this Mtr4p activity. In particular, E947, which contacts an adenine base in the Mtr4p-poly(A) crystal structure[140], when mutated to alanine relieves some of the Mtr4p-mediated regulatory pressure on TRAMP. It will be interesting to see what role (if any) the DSHCT domain plays in the recognition of other Mtr4p (i.e., non-TRAMP) substrates. Mtr4p restricts, but does not prevent further polyadenylation by TRAMP, suggesting that a given potential substrate will have to be relatively long-lived to accumulate TRAMP-appended poly(A) tails of significant length (Figure 13). Thus, it is plausible to assume that a long-lived unprocessed (or undegraded) potential RNA substrate is indicative of a bottleneck in exosome-mediated processing (or decay). Such a situation should lead to extended poly(A) tails, which could lead to tighter Mtr4p binding and decreased Mtr4p dissociation[136,142], and thus increase the opportunities for that complex to encounter and thus be processed by the exosome. Such a remarkable Mtr4p-mediated regulatory circuit could ensure that extended poly(A) tails are only appended to RNAs that have passed their expiration date.
Figure 13.
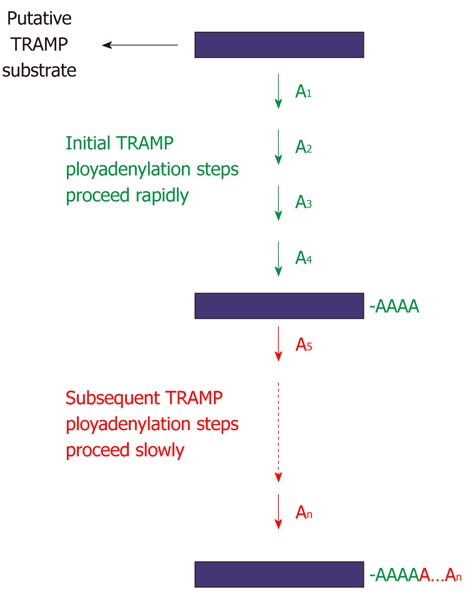
Regulation of TRAMP4 polyadenylation by Mtr4p. Mtr4p has a novel function as a regulator of TRAMP polyadenylation. Addition of the first four adenylates to a TRAMP substrate is accelerated by Mtr4p, whereas the subsequent adenylates (i.e., A5 to An) are added more slowly, suggesting that interrogation of the 3′-end of a substrate by Mtr4p plays a significant role in this regulation[139].
CONCLUSION
RNA processing and degradation are complex processes involving a number of proteins and RNAs working in concert to produce mature, functional RNA. This complex network of events can be broken down by RNA type and processing or degradation pathways affecting that RNA. Understanding the mechanism whereby an RNA is selected for processing or degradation, and which pathway that RNA takes are essential to determining how the RNA population is controlled to ensure proper cell function. RNA processing and degradation use the same basic machinery, yet have different endpoints. Thus, both processes must be tightly regulated. Such regulation likely involves an intricate network of modulating protein-protein and protein-RNA interactions. The discovery of a novel regulatory function for the Mtr4p helicase that does not involve unwinding activity, but rather its ability to bind short poly(A) 3′-ends, is an indication of the high level of complexity inherent in the regulatory schemes that ensure proper nuclear RNA processing.
ACKNOWLEDGMENTS
The authors would like to thank Gerald M Wilson for helpful comments and suggestions regarding this manuscript.
Footnotes
Supported by The Marlene and Stewart Greenebaum Cancer Center (Toth EA)
Peer reviewers: Luiz Otavio Penalva, PhD, Assistant Professor, Children’s Cancer Research Institute-UTHSCSA, Department of Cellular and Structural Biology, Mail Code 7784 -7703 Floyd Curl Dr., San Antonio, TX 78229-3900, United States; Herve Seligmann, PhD, Center for Ecological and Evolutionary Synthesis, Department of Biology, University of Oslo, Blindern, 3016 Oslo, Norway
S- Editor Cheng JX L- Editor Kerr C E- Editor Zheng XM
References
- 1.Siliciano PG, Jones MH, Guthrie C. Saccharomyces cerevisiae has a U1-like small nuclear RNA with unexpected properties. Science. 1987;237:1484–1487. doi: 10.1126/science.3306922. [DOI] [PubMed] [Google Scholar]
- 2.Butler JS. The yin and yang of the exosome. Trends Cell Biol. 2002;12:90–96. doi: 10.1016/s0962-8924(01)02225-5. [DOI] [PubMed] [Google Scholar]
- 3.Dziembowski A, Lorentzen E, Conti E, Séraphin B. A single subunit, Dis3, is essentially responsible for yeast exosome core activity. Nat Struct Mol Biol. 2007;14:15–22. doi: 10.1038/nsmb1184. [DOI] [PubMed] [Google Scholar]
- 4.Lorentzen E, Basquin J, Conti E. Structural organization of the RNA-degrading exosome. Curr Opin Struct Biol. 2008;18:709–713. doi: 10.1016/j.sbi.2008.10.004. [DOI] [PubMed] [Google Scholar]
- 5.Mitchell P, Petfalski E, Shevchenko A, Mann M, Tollervey D. The exosome: a conserved eukaryotic RNA processing complex containing multiple 3’ → 5’ exoribonucleases. Cell. 1997;91:457–466. doi: 10.1016/s0092-8674(00)80432-8. [DOI] [PubMed] [Google Scholar]
- 6.Burkard KT, Butler JS. A nuclear 3’-5’ exonuclease involved in mRNA degradation interacts with Poly(A) polymerase and the hnRNA protein Npl3p. Mol Cell Biol. 2000;20:604–616. doi: 10.1128/mcb.20.2.604-616.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Callahan KP, Butler JS. Evidence for core exosome independent function of the nuclear exoribonuclease Rrp6p. Nucleic Acids Res. 2008;36:6645–6655. doi: 10.1093/nar/gkn743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Henras AK, Soudet J, Gérus M, Lebaron S, Caizergues-Ferrer M, Mougin A, Henry Y. The post-transcriptional steps of eukaryotic ribosome biogenesis. Cell Mol Life Sci. 2008;65:2334–2359. doi: 10.1007/s00018-008-8027-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Fang F, Phillips S, Butler JS. Rat1p and Rai1p function with the nuclear exosome in the processing and degradation of rRNA precursors. RNA. 2005;11:1571–1578. doi: 10.1261/rna.2900205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.de la Cruz J, Kressler D, Tollervey D, Linder P. Dob1p (Mtr4p) is a putative ATP-dependent RNA helicase required for the 3’ end formation of 5.8S rRNA in Saccharomyces cerevisiae. EMBO J. 1998;17:1128–1140. doi: 10.1093/emboj/17.4.1128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Thomson E, Tollervey D. The final step in 5.8S rRNA processing is cytoplasmic in Saccharomyces cerevisiae. Mol Cell Biol. 2010;30:976–984. doi: 10.1128/MCB.01359-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Fromont-Racine M, Senger B, Saveanu C, Fasiolo F. Ribosome assembly in eukaryotes. Gene. 2003;313:17–42. doi: 10.1016/s0378-1119(03)00629-2. [DOI] [PubMed] [Google Scholar]
- 13.Andersen KR, Jensen TH, Brodersen DE. Take the ”A” tail--quality control of ribosomal and transfer RNA. Biochim Biophys Acta. 2008;1779:532–537. doi: 10.1016/j.bbagrm.2008.06.011. [DOI] [PubMed] [Google Scholar]
- 14.Filipowicz W, Pelczar P, Pogacic V, Dragon F. Structure and biogenesis of small nucleolar RNAs acting as guides for ribosomal RNA modification. Acta Biochim Pol. 1999;46:377–389. [PubMed] [Google Scholar]
- 15.Ooi SL, Samarsky DA, Fournier MJ, Boeke JD. Intronic snoRNA biosynthesis in Saccharomyces cerevisiae depends on the lariat-debranching enzyme: intron length effects and activity of a precursor snoRNA. RNA. 1998;4:1096–1110. doi: 10.1017/s1355838298980785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Grzechnik P, Kufel J. Polyadenylation linked to transcription termination directs the processing of snoRNA precursors in yeast. Mol Cell. 2008;32:247–258. doi: 10.1016/j.molcel.2008.10.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Nicoloso M, Qu LH, Michot B, Bachellerie JP. Intron-encoded, antisense small nucleolar RNAs: the characterization of nine novel species points to their direct role as guides for the 2’-O-ribose methylation of rRNAs. J Mol Biol. 1996;260:178–195. doi: 10.1006/jmbi.1996.0391. [DOI] [PubMed] [Google Scholar]
- 18.Ni J, Tien AL, Fournier MJ. Small nucleolar RNAs direct site-specific synthesis of pseudouridine in ribosomal RNA. Cell. 1997;89:565–573. doi: 10.1016/s0092-8674(00)80238-x. [DOI] [PubMed] [Google Scholar]
- 19.Lafontaine DL, Bousquet-Antonelli C, Henry Y, Caizergues-Ferrer M, Tollervey D. The box H + ACA snoRNAs carry Cbf5p, the putative rRNA pseudouridine synthase. Genes Dev. 1998;12:527–537. doi: 10.1101/gad.12.4.527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Leulliot N, Godin KS, Hoareau-Aveilla C, Quevillon-Cheruel S, Varani G, Henry Y, Van Tilbeurgh H. The box H/ACA RNP assembly factor Naf1p contains a domain homologous to Gar1p mediating its interaction with Cbf5p. J Mol Biol. 2007;371:1338–1353. doi: 10.1016/j.jmb.2007.06.031. [DOI] [PubMed] [Google Scholar]
- 21.Henras AK, Dez C, Henry Y. RNA structure and function in C/D and H/ACA s(no)RNPs. Curr Opin Struct Biol. 2004;14:335–343. doi: 10.1016/j.sbi.2004.05.006. [DOI] [PubMed] [Google Scholar]
- 22.Hermanns P, Bertuch AA, Bertin TK, Dawson B, Schmitt ME, Shaw C, Zabel B, Lee B. Consequences of mutations in the non-coding RMRP RNA in cartilage-hair hypoplasia. Hum Mol Genet. 2005;14:3723–3740. doi: 10.1093/hmg/ddi403. [DOI] [PubMed] [Google Scholar]
- 23.Allmang C, Kufel J, Chanfreau G, Mitchell P, Petfalski E, Tollervey D. Functions of the exosome in rRNA, snoRNA and snRNA synthesis. EMBO J. 1999;18:5399–5410. doi: 10.1093/emboj/18.19.5399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.van Hoof A, Lennertz P, Parker R. Yeast exosome mutants accumulate 3’-extended polyadenylated forms of U4 small nuclear RNA and small nucleolar RNAs. Mol Cell Biol. 2000;20:441–452. doi: 10.1128/mcb.20.2.441-452.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Xu D, Field DJ, Tang SJ, Moris A, Bobechko BP, Friesen JD. Synthetic lethality of yeast slt mutations with U2 small nuclear RNA mutations suggests functional interactions between U2 and U5 snRNPs that are important for both steps of pre-mRNA splicing. Mol Cell Biol. 1998;18:2055–2066. doi: 10.1128/mcb.18.4.2055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Maniatis T, Reed R. The role of small nuclear ribonucleoprotein particles in pre-mRNA splicing. Nature. 1987;325:673–678. doi: 10.1038/325673a0. [DOI] [PubMed] [Google Scholar]
- 27.Guthrie C. Messenger RNA splicing in yeast: clues to why the spliceosome is a ribonucleoprotein. Science. 1991;253:157–163. doi: 10.1126/science.1853200. [DOI] [PubMed] [Google Scholar]
- 28.Neugebauer KM. On the importance of being co-transcriptional. J Cell Sci. 2002;115:3865–3871. doi: 10.1242/jcs.00073. [DOI] [PubMed] [Google Scholar]
- 29.Furuichi Y, Shatkin AJ. Viral and cellular mRNA capping: past and prospects. Adv Virus Res. 2000;55:135–184. doi: 10.1016/S0065-3527(00)55003-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Shatkin AJ. Capping of eucaryotic mRNAs. Cell. 1976;9:645–653. doi: 10.1016/0092-8674(76)90128-8. [DOI] [PubMed] [Google Scholar]
- 31.Guo Z, Sherman F. 3’-end-forming signals of yeast mRNA. Mol Cell Biol. 1995;15:5983–5990. doi: 10.1128/mcb.15.11.5983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Millevoi S, Vagner S. Molecular mechanisms of eukaryotic pre-mRNA 3’ end processing regulation. Nucleic Acids Res. 2010;38:2757–2774. doi: 10.1093/nar/gkp1176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Keller W. No end yet to messenger RNA 3’ processing! Cell. 1995;81:829–832. doi: 10.1016/0092-8674(95)90001-2. [DOI] [PubMed] [Google Scholar]
- 34.Bousquet-Antonelli C, Presutti C, Tollervey D. Identification of a regulated pathway for nuclear pre-mRNA turnover. Cell. 2000;102:765–775. doi: 10.1016/s0092-8674(00)00065-9. [DOI] [PubMed] [Google Scholar]
- 35.Milligan L, Torchet C, Allmang C, Shipman T, Tollervey D. A nuclear surveillance pathway for mRNAs with defective polyadenylation. Mol Cell Biol. 2005;25:9996–10004. doi: 10.1128/MCB.25.22.9996-10004.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Hilleren P, McCarthy T, Rosbash M, Parker R, Jensen TH. Quality control of mRNA 3’-end processing is linked to the nuclear exosome. Nature. 2001;413:538–542. doi: 10.1038/35097110. [DOI] [PubMed] [Google Scholar]
- 37.Haeusler RA, Pratt-Hyatt M, Good PD, Gipson TA, Engelke DR. Clustering of yeast tRNA genes is mediated by specific association of condensin with tRNA gene transcription complexes. Genes Dev. 2008;22:2204–2214. doi: 10.1101/gad.1675908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Thompson M, Haeusler RA, Good PD, Engelke DR. Nucleolar clustering of dispersed tRNA genes. Science. 2003;302:1399–1401. doi: 10.1126/science.1089814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Wang L, Haeusler RA, Good PD, Thompson M, Nagar S, Engelke DR. Silencing near tRNA genes requires nucleolar localization. J Biol Chem. 2005;280:8637–8639. doi: 10.1074/jbc.C500017200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kadaba S, Wang X, Anderson JT. Nuclear RNA surveillance in Saccharomyces cerevisiae: Trf4p-dependent polyadenylation of nascent hypomethylated tRNA and an aberrant form of 5S rRNA. RNA. 2006;12:508–521. doi: 10.1261/rna.2305406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wang X, Jia H, Jankowsky E, Anderson JT. Degradation of hypomodified tRNA(iMet) in vivo involves RNA-dependent ATPase activity of the DExH helicase Mtr4p. RNA. 2008;14:107–116. doi: 10.1261/rna.808608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Wyers F, Rougemaille M, Badis G, Rousselle JC, Dufour ME, Boulay J, Régnault B, Devaux F, Namane A, Séraphin B, et al. Cryptic pol II transcripts are degraded by a nuclear quality control pathway involving a new poly(A) polymerase. Cell. 2005;121:725–737. doi: 10.1016/j.cell.2005.04.030. [DOI] [PubMed] [Google Scholar]
- 43.Neil H, Malabat C, d’Aubenton-Carafa Y, Xu Z, Steinmetz LM, Jacquier A. Widespread bidirectional promoters are the major source of cryptic transcripts in yeast. Nature. 2009;457:1038–1042. doi: 10.1038/nature07747. [DOI] [PubMed] [Google Scholar]
- 44.Aravind L, Watanabe H, Lipman DJ, Koonin EV. Lineage-specific loss and divergence of functionally linked genes in eukaryotes. Proc Natl Acad Sci USA. 2000;97:11319–11324. doi: 10.1073/pnas.200346997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Berretta J, Pinskaya M, Morillon A. A cryptic unstable transcript mediates transcriptional trans-silencing of the Ty1 retrotransposon in S. cerevisiae. Genes Dev. 2008;22:615–626. doi: 10.1101/gad.458008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Martens JA, Laprade L, Winston F. Intergenic transcription is required to repress the Saccharomyces cerevisiae SER3 gene. Nature. 2004;429:571–574. doi: 10.1038/nature02538. [DOI] [PubMed] [Google Scholar]
- 47.Hongay CF, Grisafi PL, Galitski T, Fink GR. Antisense transcription controls cell fate in Saccharomyces cerevisiae. Cell. 2006;127:735–745. doi: 10.1016/j.cell.2006.09.038. [DOI] [PubMed] [Google Scholar]
- 48.Arigo JT, Eyler DE, Carroll KL, Corden JL. Termination of cryptic unstable transcripts is directed by yeast RNA-binding proteins Nrd1 and Nab3. Mol Cell. 2006;23:841–851. doi: 10.1016/j.molcel.2006.07.024. [DOI] [PubMed] [Google Scholar]
- 49.Thiebaut M, Kisseleva-Romanova E, Rougemaille M, Boulay J, Libri D. Transcription termination and nuclear degradation of cryptic unstable transcripts: a role for the nrd1-nab3 pathway in genome surveillance. Mol Cell. 2006;23:853–864. doi: 10.1016/j.molcel.2006.07.029. [DOI] [PubMed] [Google Scholar]
- 50.Xue Y, Bai X, Lee I, Kallstrom G, Ho J, Brown J, Stevens A, Johnson AW. Saccharomyces cerevisiae RAI1 (YGL246c) is homologous to human DOM3Z and encodes a protein that binds the nuclear exoribonuclease Rat1p. Mol Cell Biol. 2000;20:4006–4015. doi: 10.1128/mcb.20.11.4006-4015.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Liu Q, Greimann JC, Lima CD. Reconstitution, activities, and structure of the eukaryotic RNA exosome. Cell. 2006;127:1223–1237. doi: 10.1016/j.cell.2006.10.037. [DOI] [PubMed] [Google Scholar]
- 52.Stevens A, Maupin MK. A 5’----3’ exoribonuclease of Saccharomyces cerevisiae: size and novel substrate specificity. Arch Biochem Biophys. 1987;252:339–347. doi: 10.1016/0003-9861(87)90040-3. [DOI] [PubMed] [Google Scholar]
- 53.Connelly S, Manley JL. A functional mRNA polyadenylation signal is required for transcription termination by RNA polymerase II. Genes Dev. 1988;2:440–452. doi: 10.1101/gad.2.4.440. [DOI] [PubMed] [Google Scholar]
- 54.Jimeno-González S, Haaning LL, Malagon F, Jensen TH. The yeast 5’-3’ exonuclease Rat1p functions during transcription elongation by RNA polymerase II. Mol Cell. 2010;37:580–587. doi: 10.1016/j.molcel.2010.01.019. [DOI] [PubMed] [Google Scholar]
- 55.Logan J, Falck-Pedersen E, Darnell JE, Shenk T. A poly(A) addition site and a downstream termination region are required for efficient cessation of transcription by RNA polymerase II in the mouse beta maj-globin gene. Proc Natl Acad Sci USA. 1987;84:8306–8310. doi: 10.1073/pnas.84.23.8306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Luo W, Johnson AW, Bentley DL. The role of Rat1 in coupling mRNA 3’-end processing to transcription termination: implications for a unified allosteric-torpedo model. Genes Dev. 2006;20:954–965. doi: 10.1101/gad.1409106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.El Hage A, Koper M, Kufel J, Tollervey D. Efficient termination of transcription by RNA polymerase I requires the 5’ exonuclease Rat1 in yeast. Genes Dev. 2008;22:1069–1081. doi: 10.1101/gad.463708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Kim M, Krogan NJ, Vasiljeva L, Rando OJ, Nedea E, Greenblatt JF, Buratowski S. The yeast Rat1 exonuclease promotes transcription termination by RNA polymerase II. Nature. 2004;432:517–522. doi: 10.1038/nature03041. [DOI] [PubMed] [Google Scholar]
- 59.Aldrich TL, Di Segni G, McConaughy BL, Keen NJ, Whelen S, Hall BD. Structure of the yeast TAP1 protein: dependence of transcription activation on the DNA context of the target gene. Mol Cell Biol. 1993;13:3434–3444. doi: 10.1128/mcb.13.6.3434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Lang WH, Reeder RH. The REB1 site is an essential component of a terminator for RNA polymerase I in Saccharomyces cerevisiae. Mol Cell Biol. 1993;13:649–658. doi: 10.1128/mcb.13.1.649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Oeffinger M, Zenklusen D, Ferguson A, Wei KE, El Hage A, Tollervey D, Chait BT, Singer RH, Rout MP. Rrp17p is a eukaryotic exonuclease required for 5’ end processing of Pre-60S ribosomal RNA. Mol Cell. 2009;36:768–781. doi: 10.1016/j.molcel.2009.11.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Zhang Z, Fu J, Gilmour DS. CTD-dependent dismantling of the RNA polymerase II elongation complex by the pre-mRNA 3’-end processing factor, Pcf11. Genes Dev. 2005;19:1572–1580. doi: 10.1101/gad.1296305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Chernyakov I, Whipple JM, Kotelawala L, Grayhack EJ, Phizicky EM. Degradation of several hypomodified mature tRNA species in Saccharomyces cerevisiae is mediated by Met22 and the 5’-3’ exonucleases Rat1 and Xrn1. Genes Dev. 2008;22:1369–1380. doi: 10.1101/gad.1654308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Aloy P, Ciccarelli FD, Leutwein C, Gavin AC, Superti-Furga G, Bork P, Bottcher B, Russell RB. A complex prediction: three-dimensional model of the yeast exosome. EMBO Rep. 2002;3:628–635. doi: 10.1093/embo-reports/kvf135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Houseley J, LaCava J, Tollervey D. RNA-quality control by the exosome. Nat Rev Mol Cell Biol. 2006;7:529–539. doi: 10.1038/nrm1964. [DOI] [PubMed] [Google Scholar]
- 66.Büttner K, Wenig K, Hopfner KP. Structural framework for the mechanism of archaeal exosomes in RNA processing. Mol Cell. 2005;20:461–471. doi: 10.1016/j.molcel.2005.10.018. [DOI] [PubMed] [Google Scholar]
- 67.Lorentzen E, Conti E. The exosome and the proteasome: nano-compartments for degradation. Cell. 2006;125:651–654. doi: 10.1016/j.cell.2006.05.002. [DOI] [PubMed] [Google Scholar]
- 68.Lorentzen E, Conti E. Structural basis of 3’ end RNA recognition and exoribonucleolytic cleavage by an exosome RNase PH core. Mol Cell. 2005;20:473–481. doi: 10.1016/j.molcel.2005.10.020. [DOI] [PubMed] [Google Scholar]
- 69.Oliveira CC, Gonzales FA, Zanchin NI. Temperature-sensitive mutants of the exosome subunit Rrp43p show a deficiency in mRNA degradation and no longer interact with the exosome. Nucleic Acids Res. 2002;30:4186–4198. doi: 10.1093/nar/gkf545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Luz JS, Tavares JR, Gonzales FA, Santos MC, Oliveira CC. Analysis of the Saccharomyces cerevisiae exosome architecture and of the RNA binding activity of Rrp40p. Biochimie. 2007;89:686–691. doi: 10.1016/j.biochi.2007.01.011. [DOI] [PubMed] [Google Scholar]
- 71.Ito T, Tashiro K, Muta S, Ozawa R, Chiba T, Nishizawa M, Yamamoto K, Kuhara S, Sakaki Y. Toward a protein-protein interaction map of the budding yeast: A comprehensive system to examine two-hybrid interactions in all possible combinations between the yeast proteins. Proc Natl Acad Sci USA. 2000;97:1143–1147. doi: 10.1073/pnas.97.3.1143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Allmang C, Petfalski E, Podtelejnikov A, Mann M, Tollervey D, Mitchell P. The yeast exosome and human PM-Scl are related complexes of 3’ → 5’ exonucleases. Genes Dev. 1999;13:2148–2158. doi: 10.1101/gad.13.16.2148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Lorentzen E, Dziembowski A, Lindner D, Seraphin B, Conti E. RNA channelling by the archaeal exosome. EMBO Rep. 2007;8:470–476. doi: 10.1038/sj.embor.7400945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Roppelt V, Klug G, Evguenieva-Hackenberg E. The evolutionarily conserved subunits Rrp4 and Csl4 confer different substrate specificities to the archaeal exosome. FEBS Lett. 2010;584:2931–2936. doi: 10.1016/j.febslet.2010.05.014. [DOI] [PubMed] [Google Scholar]
- 75.Lorentzen E, Walter P, Fribourg S, Evguenieva-Hackenberg E, Klug G, Conti E. The archaeal exosome core is a hexameric ring structure with three catalytic subunits. Nat Struct Mol Biol. 2005;12:575–581. doi: 10.1038/nsmb952. [DOI] [PubMed] [Google Scholar]
- 76.Ibrahim H, Wilusz J, Wilusz CJ. RNA recognition by 3’-to-5’ exonucleases: the substrate perspective. Biochim Biophys Acta. 2008;1779:256–265. doi: 10.1016/j.bbagrm.2007.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Oddone A, Lorentzen E, Basquin J, Gasch A, Rybin V, Conti E, Sattler M. Structural and biochemical characterization of the yeast exosome component Rrp40. EMBO Rep. 2007;8:63–69. doi: 10.1038/sj.embor.7400856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Wang HW, Wang J, Ding F, Callahan K, Bratkowski MA, Butler JS, Nogales E, Ke A. Architecture of the yeast Rrp44 exosome complex suggests routes of RNA recruitment for 3’ end processing. Proc Natl Acad Sci USA. 2007;104:16844–16849. doi: 10.1073/pnas.0705526104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Schneider C, Leung E, Brown J, Tollervey D. The N-terminal PIN domain of the exosome subunit Rrp44 harbors endonuclease activity and tethers Rrp44 to the yeast core exosome. Nucleic Acids Res. 2009;37:1127–1140. doi: 10.1093/nar/gkn1020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Schneider C, Anderson JT, Tollervey D. The exosome subunit Rrp44 plays a direct role in RNA substrate recognition. Mol Cell. 2007;27:324–331. doi: 10.1016/j.molcel.2007.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Bonneau F, Basquin J, Ebert J, Lorentzen E, Conti E. The yeast exosome functions as a macromolecular cage to channel RNA substrates for degradation. Cell. 2009;139:547–559. doi: 10.1016/j.cell.2009.08.042. [DOI] [PubMed] [Google Scholar]
- 82.Moser MJ, Holley WR, Chatterjee A, Mian IS. The proofreading domain of Escherichia coli DNA polymerase I and other DNA and/or RNA exonuclease domains. Nucleic Acids Res. 1997;25:5110–5118. doi: 10.1093/nar/25.24.5110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Beese LS, Steitz TA. Structural basis for the 3’-5’ exonuclease activity of Escherichia coli DNA polymerase I: a two metal ion mechanism. EMBO J. 1991;10:25–33. doi: 10.1002/j.1460-2075.1991.tb07917.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Morozov V, Mushegian AR, Koonin EV, Bork P. A putative nucleic acid-binding domain in Bloom’s and Werner’s syndrome helicases. Trends Biochem Sci. 1997;22:417–418. doi: 10.1016/s0968-0004(97)01128-6. [DOI] [PubMed] [Google Scholar]
- 85.Stead JA, Costello JL, Livingstone MJ, Mitchell P. The PMC2NT domain of the catalytic exosome subunit Rrp6p provides the interface for binding with its cofactor Rrp47p, a nucleic acid-binding protein. Nucleic Acids Res. 2007;35:5556–5567. doi: 10.1093/nar/gkm614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Lykke-Andersen S, Brodersen DE, Jensen TH. Origins and activities of the eukaryotic exosome. J Cell Sci. 2009;122:1487–1494. doi: 10.1242/jcs.047399. [DOI] [PubMed] [Google Scholar]
- 87.Greimann JC, Lima CD. Reconstitution of RNA exosomes from human and Saccharomyces cerevisiae cloning, expression, purification, and activity assays. Methods Enzymol. 2008;448:185–210. doi: 10.1016/S0076-6879(08)02610-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Canavan R, Bond U. Deletion of the nuclear exosome component RRP6 leads to continued accumulation of the histone mRNA HTB1 in S-phase of the cell cycle in Saccharomyces cerevisiae. Nucleic Acids Res. 2007;35:6268–6279. doi: 10.1093/nar/gkm691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Carneiro T, Carvalho C, Braga J, Rino J, Milligan L, Tollervey D, Carmo-Fonseca M. Depletion of the yeast nuclear exosome subunit Rrp6 results in accumulation of polyadenylated RNAs in a discrete domain within the nucleolus. Mol Cell Biol. 2007;27:4157–4165. doi: 10.1128/MCB.00120-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Das B, Butler JS, Sherman F. Degradation of normal mRNA in the nucleus of Saccharomyces cerevisiae. Mol Cell Biol. 2003;23:5502–5515. doi: 10.1128/MCB.23.16.5502-5515.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Kuai L, Das B, Sherman F. A nuclear degradation pathway controls the abundance of normal mRNAs in Saccharomyces cerevisiae. Proc Natl Acad Sci USA. 2005;102:13962–13967. doi: 10.1073/pnas.0506518102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.LaCava J, Houseley J, Saveanu C, Petfalski E, Thompson E, Jacquier A, Tollervey D. RNA degradation by the exosome is promoted by a nuclear polyadenylation complex. Cell. 2005;121:713–724. doi: 10.1016/j.cell.2005.04.029. [DOI] [PubMed] [Google Scholar]
- 93.Mitchell P, Petfalski E, Houalla R, Podtelejnikov A, Mann M, Tollervey D. Rrp47p is an exosome-associated protein required for the 3’ processing of stable RNAs. Mol Cell Biol. 2003;23:6982–6992. doi: 10.1128/MCB.23.19.6982-6992.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Milligan L, Decourty L, Saveanu C, Rappsilber J, Ceulemans H, Jacquier A, Tollervey D. A yeast exosome cofactor, Mpp6, functions in RNA surveillance and in the degradation of noncoding RNA transcripts. Mol Cell Biol. 2008;28:5446–5457. doi: 10.1128/MCB.00463-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Steinmetz EJ, Conrad NK, Brow DA, Corden JL. RNA-binding protein Nrd1 directs poly(A)-independent 3’-end formation of RNA polymerase II transcripts. Nature. 2001;413:327–331. doi: 10.1038/35095090. [DOI] [PubMed] [Google Scholar]
- 96.Yuryev A, Patturajan M, Litingtung Y, Joshi RV, Gentile C, Gebara M, Corden JL. The C-terminal domain of the largest subunit of RNA polymerase II interacts with a novel set of serine/arginine-rich proteins. Proc Natl Acad Sci USA. 1996;93:6975–6980. doi: 10.1073/pnas.93.14.6975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Conrad NK, Wilson SM, Steinmetz EJ, Patturajan M, Brow DA, Swanson MS, Corden JL. A yeast heterogeneous nuclear ribonucleoprotein complex associated with RNA polymerase II. Genetics. 2000;154:557–571. doi: 10.1093/genetics/154.2.557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Lee JM, Greenleaf AL. CTD kinase large subunit is encoded by CTK1, a gene required for normal growth of Saccharomyces cerevisiae. Gene Expr. 1991;1:149–167. [PMC free article] [PubMed] [Google Scholar]
- 99.Steinmetz EJ, Brow DA. Repression of gene expression by an exogenous sequence element acting in concert with a heterogeneous nuclear ribonucleoprotein-like protein, Nrd1, and the putative helicase Sen1. Mol Cell Biol. 1996;16:6993–7003. doi: 10.1128/mcb.16.12.6993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Singh N, Ma Z, Gemmill T, Wu X, Defiglio H, Rossettini A, Rabeler C, Beane O, Morse RH, Palumbo MJ, et al. The Ess1 prolyl isomerase is required for transcription termination of small noncoding RNAs via the Nrd1 pathway. Mol Cell. 2009;36:255–266. doi: 10.1016/j.molcel.2009.08.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Granato DC, Machado-Santelli GM, Oliveira CC. Nop53p interacts with 5.8S rRNA co-transcriptionally, and regulates processing of pre-rRNA by the exosome. FEBS J. 2008;275:4164–4178. doi: 10.1111/j.1742-4658.2008.06565.x. [DOI] [PubMed] [Google Scholar]
- 102.Ho Y, Gruhler A, Heilbut A, Bader GD, Moore L, Adams SL, Millar A, Taylor P, Bennett K, Boutilier K, et al. Systematic identification of protein complexes in Saccharomyces cerevisiae by mass spectrometry. Nature. 2002;415:180–183. doi: 10.1038/415180a. [DOI] [PubMed] [Google Scholar]
- 103.Sydorskyy Y, Dilworth DJ, Halloran B, Yi EC, Makhnevych T, Wozniak RW, Aitchison JD. Nop53p is a novel nucleolar 60S ribosomal subunit biogenesis protein. Biochem J. 2005;388:819–826. doi: 10.1042/BJ20041297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Granato DC, Gonzales FA, Luz JS, Cassiola F, Machado-Santelli GM, Oliveira CC. Nop53p, an essential nucleolar protein that interacts with Nop17p and Nip7p, is required for pre-rRNA processing in Saccharomyces cerevisiae. FEBS J. 2005;272:4450–4463. doi: 10.1111/j.1742-4658.2005.04861.x. [DOI] [PubMed] [Google Scholar]
- 105.Thomson E, Tollervey D. Nop53p is required for late 60S ribosome subunit maturation and nuclear export in yeast. RNA. 2005;11:1215–1224. doi: 10.1261/rna.2720205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Callahan KP, Butler JS. TRAMP complex enhances RNA degradation by the nuclear exosome component Rrp6. J Biol Chem. 2010;285:3540–3547. doi: 10.1074/jbc.M109.058396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Dez C, Houseley J, Tollervey D. Surveillance of nuclear-restricted pre-ribosomes within a subnucleolar region of Saccharomyces cerevisiae. EMBO J. 2006;25:1534–1546. doi: 10.1038/sj.emboj.7601035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.San Paolo S, Vanacova S, Schenk L, Scherrer T, Blank D, Keller W, Gerber AP. Distinct roles of non-canonical poly(A) polymerases in RNA metabolism. PLoS Genet. 2009;5:e1000555. doi: 10.1371/journal.pgen.1000555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Egecioglu DE, Henras AK, Chanfreau GF. Contributions of Trf4p- and Trf5p-dependent polyadenylation to the processing and degradative functions of the yeast nuclear exosome. RNA. 2006;12:26–32. doi: 10.1261/rna.2207206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Houseley J, Tollervey D. Yeast Trf5p is a nuclear poly(A) polymerase. EMBO Rep. 2006;7:205–211. doi: 10.1038/sj.embor.7400612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Vanácová S, Wolf J, Martin G, Blank D, Dettwiler S, Friedlein A, Langen H, Keith G, Keller W. A new yeast poly(A) polymerase complex involved in RNA quality control. PLoS Biol. 2005;3:e189. doi: 10.1371/journal.pbio.0030189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Roth KM, Byam J, Fang F, Butler JS. Regulation of NAB2 mRNA 3’-end formation requires the core exosome and the Trf4p component of the TRAMP complex. RNA. 2009;15:1045–1058. doi: 10.1261/rna.709609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Hamill S, Wolin SL, Reinisch KM. Structure and function of the polymerase core of TRAMP, a RNA surveillance complex. Proc Natl Acad Sci USA. 2010;107:15045–15050. doi: 10.1073/pnas.1003505107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Osley MA, Hereford L. Identification of a sequence responsible for periodic synthesis of yeast histone 2A mRNA. Proc Natl Acad Sci USA. 1982;79:7689–7693. doi: 10.1073/pnas.79.24.7689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Castaño IB, Brzoska PM, Sadoff BU, Chen H, Christman MF. Mitotic chromosome condensation in the rDNA requires TRF4 and DNA topoisomerase I in Saccharomyces cerevisiae. Genes Dev. 1996;10:2564–2576. doi: 10.1101/gad.10.20.2564. [DOI] [PubMed] [Google Scholar]
- 116.Edwards S, Li CM, Levy DL, Brown J, Snow PM, Campbell JL. Saccharomyces cerevisiae DNA polymerase epsilon and polymerase sigma interact physically and functionally, suggesting a role for polymerase epsilon in sister chromatid cohesion. Mol Cell Biol. 2003;23:2733–2748. doi: 10.1128/MCB.23.8.2733-2748.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Sadoff BU, Heath-Pagliuso S, Castaño IB, Zhu Y, Kieff FS, Christman MF. Isolation of mutants of Saccharomyces cerevisiae requiring DNA topoisomerase I. Genetics. 1995;141:465–479. doi: 10.1093/genetics/141.2.465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Walowsky C, Fitzhugh DJ, Castaño IB, Ju JY, Levin NA, Christman MF. The topoisomerase-related function gene TRF4 affects cellular sensitivity to the antitumor agent camptothecin. J Biol Chem. 1999;274:7302–7308. doi: 10.1074/jbc.274.11.7302. [DOI] [PubMed] [Google Scholar]
- 119.Wang Z, Castaño IB, Adams C, Vu C, Fitzhugh D, Christman MF. Structure/function analysis of the Saccharomyces cerevisiae Trf4/Pol sigma DNA polymerase. Genetics. 2002;160:381–391. doi: 10.1093/genetics/160.2.381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Carson DR, Christman MF. Evidence that replication fork components catalyze establishment of cohesion between sister chromatids. Proc Natl Acad Sci USA. 2001;98:8270–8275. doi: 10.1073/pnas.131022798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Wang Z, Castaño IB, De Las Peñas A, Adams C, Christman MF. Pol kappa: A DNA polymerase required for sister chromatid cohesion. Science. 2000;289:774–779. doi: 10.1126/science.289.5480.774. [DOI] [PubMed] [Google Scholar]
- 122.Reis CC, Campbell JL. Contribution of Trf4/5 and the nuclear exosome to genome stability through regulation of histone mRNA levels in Saccharomyces cerevisiae. Genetics. 2007;175:993–1010. doi: 10.1534/genetics.106.065987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Gunjan A, Verreault A. A Rad53 kinase-dependent surveillance mechanism that regulates histone protein levels in S. cerevisiae. Cell. 2003;115:537–549. doi: 10.1016/s0092-8674(03)00896-1. [DOI] [PubMed] [Google Scholar]
- 124.Schwabish MA, Struhl K. Asf1 mediates histone eviction and deposition during elongation by RNA polymerase II. Mol Cell. 2006;22:415–422. doi: 10.1016/j.molcel.2006.03.014. [DOI] [PubMed] [Google Scholar]
- 125.Tyler JK, Adams CR, Chen SR, Kobayashi R, Kamakaka RT, Kadonaga JT. The RCAF complex mediates chromatin assembly during DNA replication and repair. Nature. 1999;402:555–560. doi: 10.1038/990147. [DOI] [PubMed] [Google Scholar]
- 126.Tong AH, Lesage G, Bader GD, Ding H, Xu H, Xin X, Young J, Berriz GF, Brost RL, Chang M, et al. Global mapping of the yeast genetic interaction network. Science. 2004;303:808–813. doi: 10.1126/science.1091317. [DOI] [PubMed] [Google Scholar]
- 127.Wery M, Ruidant S, Schillewaert S, Leporé N, Lafontaine DL. The nuclear poly(A) polymerase and Exosome cofactor Trf5 is recruited cotranscriptionally to nucleolar surveillance. RNA. 2009;15:406–419. doi: 10.1261/rna.1402709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Haracska L, Johnson RE, Prakash L, Prakash S. Trf4 and Trf5 proteins of Saccharomyces cerevisiae exhibit poly(A) RNA polymerase activity but no DNA polymerase activity. Mol Cell Biol. 2005;25:10183–10189. doi: 10.1128/MCB.25.22.10183-10189.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Shen EC, Henry MF, Weiss VH, Valentini SR, Silver PA, Lee MS. Arginine methylation facilitates the nuclear export of hnRNP proteins. Genes Dev. 1998;12:679–691. doi: 10.1101/gad.12.5.679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Inoue K, Mizuno T, Wada K, Hagiwara M. Novel RING finger proteins, Air1p and Air2p, interact with Hmt1p and inhibit the arginine methylation of Npl3p. J Biol Chem. 2000;275:32793–32799. doi: 10.1074/jbc.M004560200. [DOI] [PubMed] [Google Scholar]
- 131.Aletta JM, Cimato TR, Ettinger MJ. Protein methylation: a signal event in post-translational modification. Trends Biochem Sci. 1998;23:89–91. doi: 10.1016/s0968-0004(98)01185-2. [DOI] [PubMed] [Google Scholar]
- 132.Fromont-Racine M, Mayes AE, Brunet-Simon A, Rain JC, Colley A, Dix I, Decourty L, Joly N, Ricard F, Beggs JD, et al. Genome-wide protein interaction screens reveal functional networks involving Sm-like proteins. Yeast. 2000;17:95–110. doi: 10.1002/1097-0061(20000630)17:2<95::AID-YEA16>3.0.CO;2-H. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Tanner NK, Linder P. DExD/H box RNA helicases: from generic motors to specific dissociation functions. Mol Cell. 2001;8:251–262. doi: 10.1016/s1097-2765(01)00329-x. [DOI] [PubMed] [Google Scholar]
- 134.Silverman E, Edwalds-Gilbert G, Lin RJ. DExD/H-box proteins and their partners: helping RNA helicases unwind. Gene. 2003;312:1–16. doi: 10.1016/s0378-1119(03)00626-7. [DOI] [PubMed] [Google Scholar]
- 135.Liang S, Hitomi M, Hu YH, Liu Y, Tartakoff AM. A DEAD-box-family protein is required for nucleocytoplasmic transport of yeast mRNA. Mol Cell Biol. 1996;16:5139–5146. doi: 10.1128/mcb.16.9.5139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Bernstein J, Patterson DN, Wilson GM, Toth EA. Characterization of the essential activities of Saccharomyces cerevisiae Mtr4p, a 3’-& gt; 5’ helicase partner of the nuclear exosome. J Biol Chem. 2008;283:4930–4942. doi: 10.1074/jbc.M706677200. [DOI] [PubMed] [Google Scholar]
- 137.Staub E, Fiziev P, Rosenthal A, Hinzmann B. Insights into the evolution of the nucleolus by an analysis of its protein domain repertoire. Bioessays. 2004;26:567–581. doi: 10.1002/bies.20032. [DOI] [PubMed] [Google Scholar]
- 138.Jackson RN, Klauer AA, Hintze BJ, Robinson H, van Hoof A, Johnson SJ. The crystal structure of Mtr4 reveals a novel arch domain required for rRNA processing. EMBO J. 2010;29:2205–2216. doi: 10.1038/emboj.2010.107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Jia H, Wang X, Liu F, Guenther UP, Srinivasan S, Anderson JT, Jankowsky E. The RNA helicase Mtr4p modulates polyadenylation in the TRAMP complex. Cell. 2011;145:890–901. doi: 10.1016/j.cell.2011.05.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Weir JR, Bonneau F, Hentschel J, Conti E. Structural analysis reveals the characteristic features of Mtr4, a DExH helicase involved in nuclear RNA processing and surveillance. Proc Natl Acad Sci USA. 2010;107:12139–12144. doi: 10.1073/pnas.1004953107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Bernstein KA, Granneman S, Lee AV, Manickam S, Baserga SJ. Comprehensive mutational analysis of yeast DEXD/H box RNA helicases involved in large ribosomal subunit biogenesis. Mol Cell Biol. 2006;26:1195–1208. doi: 10.1128/MCB.26.4.1195-1208.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Bernstein J, Ballin JD, Patterson DN, Wilson GM, Toth EA. Unique properties of the Mtr4p-poly(A) complex suggest a role in substrate targeting. Biochemistry. 2010;49:10357–10370. doi: 10.1021/bi101518x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Nurmohamed S, Vaidialingam B, Callaghan AJ, Luisi BF. Crystal structure of Escherichia coli polynucleotide phosphorylase core bound to RNase E, RNA and manganese: implications for catalytic mechanism and RNA degradosome assembly. J Mol Biol. 2009;389:17–33. doi: 10.1016/j.jmb.2009.03.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Lorentzen E, Basquin J, Tomecki R, Dziembowski A, Conti E. Structure of the active subunit of the yeast exosome core, Rrp44: diverse modes of substrate recruitment in the RNase II nuclease family. Mol Cell. 2008;29:717–728. doi: 10.1016/j.molcel.2008.02.018. [DOI] [PubMed] [Google Scholar]
- 145.Midtgaard SF, Assenholt J, Jonstrup AT, Van LB, Jensen TH, Brodersen DE. Structure of the nuclear exosome component Rrp6p reveals an interplay between the active site and the HRDC domain. Proc Natl Acad Sci USA. 2006;103:11898–11903. doi: 10.1073/pnas.0604731103. [DOI] [PMC free article] [PubMed] [Google Scholar]