

Summary
Background
The spindle checkpoint ensures accurate chromosome transmission by delaying chromosome segregation until all chromosomes are correctly aligned on the mitotic spindle. The checkpoint is activated by kinetochores that are not attached to microtubules or are attached but not under tension and arrests cells at metaphase by inhibiting the anaphase-promoting complex (APC) and its co-activator Cdc20. Despite numerous studies, we still do not understand how the checkpoint proteins coordinate with each other to inhibit APCCdc20 activity.
Results
To ask how the checkpoint components induce metaphase arrest, we constructed fusions of checkpoint proteins and expressed them in the budding yeast, Saccharomyces cerevisiae, to mimic possible protein interactions during checkpoint activation. We found that expression of a Mad2-Mad3 protein fusion or non-covalently linked Mad2 and Mad3, but not the overexpression of the two separate proteins, induces metaphase arrest that is independent of functional kinetochores or other checkpoint proteins. We further showed that artificially tethering Mad2 to Cdc20 also arrests cells in metaphase independently of other checkpoint components.
Conclusion
Our results suggest that Mad3 is required for the stable binding of Mad2 to Cdc20 in vivo, which is sufficient to inhibit APC activity and is the most downstream event in spindle checkpoint activation.
Introduction
Faithful segregation of genetic material during cell division is essential for the viability of all organisms. For each chromosome, DNA replication creates two identical copies, which are segregated from each other at mitosis. Segregation is directed by the kinetochore, a specialized multi-protein structure that assembles on centromeric DNA and binds to and moves along microtubules. Normal segregation depends on the two sister kinetochores attaching to microtubules from opposite spindle poles during mitosis. Eukaryotes use a control circuit called the spindle checkpoint to ensure accurate segregation. During unperturbed mitosis, an E3 ubiquitin ligase known as the anaphase-promoting complex (APC) and its co-activator Cdc20 triggers anaphase and chromosome segregation by catalyzing the ubiquitination and destruction of securin (Pds1 in budding yeast) (Figure 1A). The absence of microtubule attachment [1, 2] or the lack of tension at the kinetochore (because of chromosome failing to attach to opposite spindle poles) [3-5] activates the checkpoint, which arrests cells at the metaphase-to-anaphase transition by targeting APC and Cdc20 for inhibition (for reviews see [6, 7]). In the budding yeast, Saccharomyces cerevisiae, the key players of the spindle checkpoint include Mad1, Mad2, Mad3, Bub1, Bub3, Mps1, and Ipl1, all of which are highly conserved among eukaryotes [1, 2, 8, 9].
Figure 1.

A model for spindle checkpoint activation (adapted from [6, 7]). (A) During mitosis, when all chromosomes are properly attached to microtubules, the anaphase-promoting complex (APC) and its co-activator Cdc20 polyubiquitinate different substrates such as securin (Pds1 in budding yeast), leading to its destruction and anaphase onset. (B) The spindle checkpoint is activated by the absence of microtubule attachment or the lack of tension at the kinetochore. Mad1-Mad2 complexes associate with unattached kinetochores and recruit the “open” Mad2 conformer (O-Mad2), facilitating the formation of a “closed” Mad2 (C-Mad2)-Cdc20 complex. The closed Mad2-Cdc20 complex associates with Mad3 and Bub3, which can be recruited to the kinetochores by interactions with Bub1 (not shown), to form the mitotic checkpoint complex (MCC). Inhibition of APC activity by the MCC arrests the cells in metaphase and gives the cells time to correct attachment errors at the kinetochores.
Although the checkpoint proteins have been studied extensively, we lack a molecular description of how events at the kinetochore are converted into inhibition of the APC. Several models have been described including the conformational change (Mad2-template) model [6], which proposes that Mad1-Mad2 complexes associate with kinetochores that lack microtubule attachments and recruit an “open” Mad2 conformer (O-Mad2), facilitating the formation of the “closed” Mad2 (C-Mad2)-Cdc20 complex (Figure 1B). Besides the recruitment of Mad1 and Mad2 to unattached kinetochores, experiments such as fluorescent protein localization and coimmunoprecipitation (co-IP) have shown that in budding yeast both Bub1 and Bub3 can associate with kinetochore [10] and Mad1 [11], while Mad3 can interact with both Mad2 and Bub3 [12]. This complicated network of interactions can potentially bring different checkpoint proteins together at the kinetochores in response to attachment errors and lead to formation of additional inhibitory complexes. One example is the mitotic checkpoint complex (MCC), which is proposed to consist of Mad2, Mad3, Bub3 and Cdc20 and has been shown to be a potent inhibitor of APCCdc20 [13, 14] (Figure 1B). Inhibition of APC activity arrests cells in metaphase and provides the cells a chance to correct the attachment errors at the kinetochores. The spindle checkpoint hence ensures that cells only progress through mitosis when all chromosomes are properly attached.
The initial studies that identified Cdc20 as the target of the spindle checkpoint showed that both Mad2 and Mad3 bind to Cdc20 [15]. We have investigated the consequences of this binding by manipulating the linkage between Mad2, Mad3, and Cdc20. Expressing physically-linked Mad2 and Mad3 induces a metaphase arrest that does not require functional kinetochores or other checkpoint proteins, indicating the Mad2-Mad3 fusion alone is sufficient to inhibit APC activity. We also show that tethering Mad2 directly to Cdc20 can lead to similar arrest that does not require Mad3 or other checkpoint components, supporting the idea that the Mad2-Mad3 fusion induces metaphase arrest by promoting an intimate association between Mad2 and Cdc20. Our results suggest that the most downstream event in spindle checkpoint activation is the cooperative binding of Mad2 and Mad3 to Cdc20.
Results
Expressing physically-linked Mad2 and Mad3 leads to metaphase arrest
Previous studies in budding yeast showed that both Mad2 and Mad3 are part of the MCC and associate with Cdc20 [12, 13, 15]. If Mad2 and Mad3 can both bind to Cdc20, checkpoint activation could strengthen the interaction between Mad2 and Mad3, making them bind more avidly to Cdc20 and arresting cells in mitosis. In this scenario, an engineered Mad2-Mad3 complex would artificially activate the spindle checkpoint. To test this idea, we asked if overexpressing different versions of Mad2 and Mad3 from the GAL1 promoter could arrest wild-type yeast cells going through a synchronous cell cycle. We integrated the constructs for overexpression into the yeast genome without modifying the endogenous spindle checkpoint genes. The GAL1 promoter is transcribed in the presence of galactose and inhibited by glucose. We arrested the cells in G1 with α-factor, then released them into media with either glucose or galactose, and looked at them three hours later. Cells that are cycling normally have no buds or buds that are clearly smaller than the mother cell, whereas cells that have trouble progressing through mitosis are enlarged and have distinctive large buds that approach the size of the mother cell. Overexpressing Mad2, Mad3, or both Mad2 and Mad3 together had little effect on the cells; more than 90% of the population continued to cycle when grown in galactose-containing medium (Figure 2A). In contrast, overexpressing a Mad2-Mad3 protein fusion, with the C terminus of Mad2 fused to the N terminus of Mad3 by an 8-amino acid linker, led to accumulation of large budded cells, a hallmark of metaphase arrest (Figure 2A). Expressing a fusion with Mad3 at the N terminus (Mad3-Mad2) produced similar result (data not shown). Cells arrested rapidly, as large budded cells started to accumulate at the first mitosis after inducing the Mad2-Mad3 fusion protein (Figure 2B). Overexpression of a Mad2-Mad3 fusion is therefore able to induce a strong metaphase arrest that does not occur when Mad2 and Mad3 are overexpressed as two independent proteins in the same cell.
Figure 2.
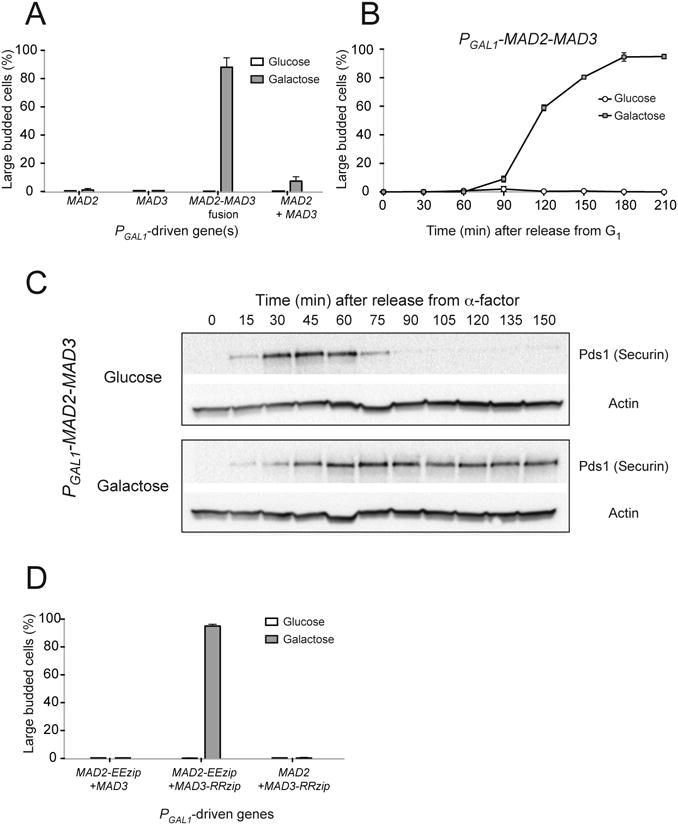
Overexpressing a Mad2-Mad3 protein fusion leads to metaphase arrest. (A) Cells with the indicated PGAL1-driven genes were grown to mid-log phase, arrested in G1 with α-factor, and were released into media with either glucose or galactose. After 3 hours of growth, the percentage of large budded cells was determined by light microscopy as a measure of metaphase arrest. Error bars represent the standard deviation of three independent trials. Two hundred cells were counted for each trial. (B) PGAL1-MAD2-MAD3 cells were released from G1 arrest into glucose- or galactose-containing media. Samples were collected at the indicated time point and the percentage of large budded cells was determined by light microscopy. Error bars represent the standard deviation of three independent trials. Two hundred cells were counted at each time point for each trial. (C) Cell cycle progression of PGAL1-MAD2-MAD3 cells was monitored by Western blots (n=3), which detect securin (Pds1), a protein that is destroyed as cells enter anaphase. Cells were grown to mid-log phase and arrested in G1 with α-factor, and were released into media with either glucose (top) or galactose (bottom). Western blots against Myc (to visualize Myc-tagged securin) or actin (loading control) were performed. When the cells were grown in glucose, securin level first increased and then dropped rapidly as cells progressed into anaphase. When the Mad2-Mad3 fusion was overexpressed in the presence of galactose, securin was stabilized, indicating that the cells were arrested in metaphase. (D) Overexpressing Mad2 and Mad3 linked by leucine zippers also induces metaphase arrest. Cells with the indicated PGAL1-driven genes were released from G1 arrest into glucose- or galactose-containing media. The percentage of large budded cells was determined by light microscopy after 3 hours of growth. Error bars represent the standard deviation of three independent trials. Two hundred cells were counted for each trial.
We confirmed that expressing Mad2-Mad3 fusions prevents APC activation by using a biochemical marker for the exit into anaphase. The disappearance of securin (named Pds1 in budding yeast), a target of APCCdc20, triggers the metaphase-to-anaphase transition and we followed the level of this protein by monitoring the level of epitope-tagged securin (Pds1-18×Myc) on Western Blots. When the Mad2-Mad3 fusion was not expressed (glucose), securin levels rose and fell after cells were released from G1 arrest, indicating normal cell cycle progression (Figure 2C, top). Overexpressing the Mad2-Mad3 fusion (galactose) stabilized securin (Figure 2C, bottom).
We concluded that overexpressing the Mad2-Mad3 fusion inhibits APC and arrests cells in metaphase. To avoid the possibility that this arrest reflects some peculiarity of how Mad2 and Mad3 were fused together, we tested the effect of linking Mad2 and Mad3 using a different, non-covalent method. We fused Mad2 and Mad3 to engineered leucine zipper sequences (EEzip and RRzip) that allow the formation of stable heterodimers between two zippers bearing opposing charges [16]. When we expressed Mad2-Mad3 heterodimers (by releasing strains containing both PGAL1-MAD2-EEzip and PGAL1-MAD3-RRzip into galactose-containing medium), 90% of the cells arrested in metaphase (Figure 2D). The phenotype is not observed when Mad2-EEzip was expressed with untagged Mad3 and vice versa, indicating that the arrest depends on interaction between Mad2 and Mad3 through the leucine zipper (Figure 2D). The results confirm that overexpressing Mad2 and Mad3 can only induce metaphase arrest if the two proteins are held in close proximity.
Mad2-Mad3 fusion does not affect mitotic spindle structure
The Mad2-Mad3 fusion could lead to metaphase arrest in two ways: directly, by inhibiting APCCdc20 activity, or indirectly, by detaching microtubules from kinetochores, thus activating the spindle checkpoint. To determine the mode of checkpoint activation, we looked at chromosome bi-orientation in cells arrested in metaphase. A chromosome is said to bi-orient when the two sister kinetochores attach to microtubules from opposite spindle poles. The sister kinetochores can be pulled apart from each other by the spindle, and the separation, which can extend to about 0.5 μm, can be visualized by expressing a GFP-Lac repressor fusion protein (GFP-LacI) and placing a lactose operator (LacO) array near the centromere [17-20]. Since bi-orientation is generated when chromosomes align correctly on the spindle, fewer chromosomes will bi-orient in cells arrested by Mad2-Mad3 overexpression if the fusion disrupts kinetochore-microtubule attachments or causes both sister kinetochores to attach to the same spindle pole. We integrated PGAL1-MAD2-MAD3 into a strain that expresses GFP-LacI, has a LacO array near the centromere of chromosome XV, and CDC20 under the methionine-repressible MET3 promoter (PMET3-CDC20). When the cells were grown in medium with glucose and methionine, Cdc20 was not expressed and the cells arrested in metaphase because they cannot activate the APC. Cdc20 depletion leads to metaphase arrest without disrupting the mitotic spindle, and around 70% of the cells arrested by this mechanism contained two GFP dots, indicating that chromosome XV bi-oriented (Figure 3A). In about 30% of the cells, the separation between the two sister kinetochores is too small to allow the two LacO arrays to be resolved into two dots (Figure 3A). The percentage of cells with two GFP dots was statistically indistinguishable when cells were arrested by overexpression of the Mad2-Mad3 fusion (by growing in medium with galactose and no methionine) (Figure 3A). The results show that chromosomes bi-orient normally during arrest induced by the Mad2-Mad3 fusion, suggesting that the fusion leads to metaphase arrest directly and does not disrupt normal spindle structure.
Figure 3.
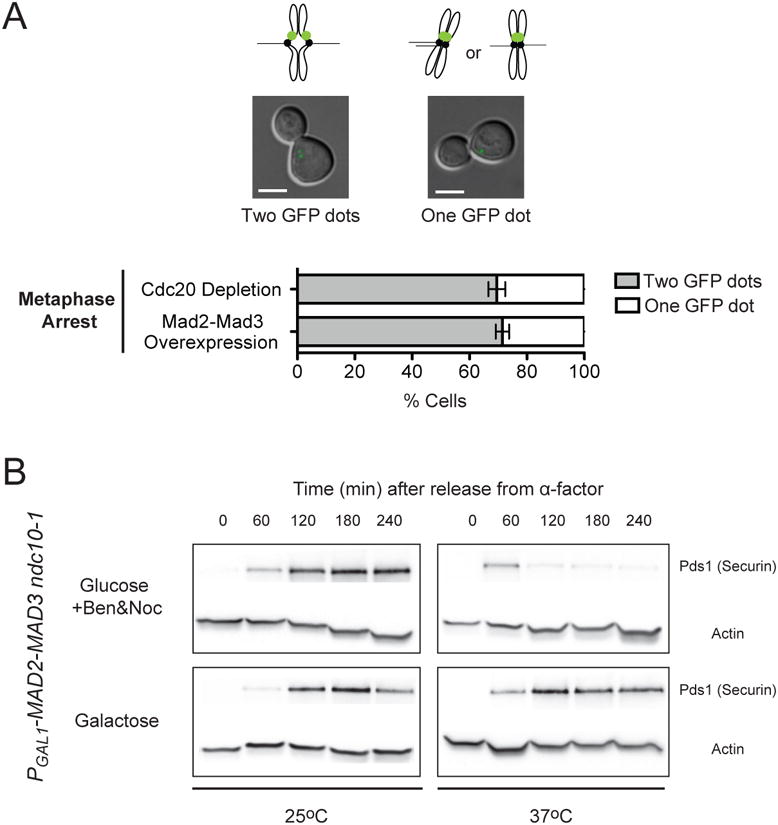
Metaphase arrest by the Mad2-Mad3 fusion is independent of kinetochore-microtubule attachments. (A) Chromosomes bi-orient in cells overexpressing the Mad2-Mad3 fusion. To achieve metaphase arrest, cells carrying PGAL1-MAD2-MAD3, GFP-LacI, a LacO array located near the centromere of chromosome XV, and PMET3-CDC20 were released from G1 arrest into either medium with glucose and methionine (Cdc20 depletion) or medium with galactose and no methionine (Mad2-Mad3 overexpression). Differential interference contrast (DIC) and GFP images of the cells were taken 3 hours after their release from G1. One or two GFP dots can be seen in cells arrested in metaphase. Cells have two GFP dots when chromosome XV bi-orients and the two sister kinetochores are separated by the spindle. Cells have one GFP dot when chromosome XV mono-orients (sister kinetochores attaching to the same spindle pole) or bi-orients but the two sister kinetochores are not pulled apart enough to allow resolution of two separate dots. Representative images of metaphase-arrested cells with one or two GFP dots are shown (Scale bar, 5 μm). Bar graph shows the percentage of cells with one or two GFP dots when they were arrested by Cdc20 depletion or overexpression of the Mad2-Mad3 fusion. Error bars represent the standard deviation of three independent trials. Two hundred cells were counted for each trial. Around 70% of cells arrested by Cdc20 depletion had two GFP dots. The percentage of cells that showed two GFP dots when the cells were arrested by the Mad2-Mad3 fusion is statistically indistinguishable (P=0.415; two-tailed Student's t test). Cells therefore show normal chromosome bi-orientation when they are arrested in metaphase by the Mad2-Ma3 fusion, suggesting that the fusion does not disrupt kinetochore-microtubule connections. (B) Metaphase arrest by the Mad2-Mad3 fusions does not require functional kinetochores. Cell cycle progression of cells with PGAL1-MAD2-MAD3 and ndc10-1 (a mutation that inactivates kinetochore at 37°C) was measured by Western blots (n=3). Cells were released from G1 arrest into media with either glucose with benomyl and nocodazole (top) or galactose (bottom) at 25°C or 37°C. Western blots against Myc (to visualize Myc-tagged securin) or actin (loading control) were performed. Securin was stabilized in galactose-containing medium even at 37°C, indicating that the metaphase arrest is independent of functional kinetochores.
We next asked if the Mad2-Mad3 fusion could arrest cells that lacked kinetochores. The ability of microtubule poisons to activate the spindle checkpoint depends on the presence of functional kinetochores [21, 22]. Thus, demonstrating kinetochore-independent arrest would strengthen the conclusion that the Mad2-Mad3 fusion does not activate the checkpoint by disrupting kinetochore-microtubule connections. We looked at the phenotype of Mad2-Mad3 overexpression in the absence of functional kinetochores by using an ndc10-1 strain. Ndc10 is a member of the CBF3 complex of the budding yeast kinetochore, which recognizes the centromeric DNA sequence and acts as the primary link between the chromosome and microtubule binding complexes of the kinetochore [23]. At the restrictive temperature (37°C), ndc10-1 cells lack functional kinetochores and are therefore unable to activate the spindle checkpoint even in the presence of the microtubule deploymerizing drugs benomyl and nocodazole [21, 22]. If the Mad2-Mad3 fusion activates the checkpoint by disrupting microtubule attachment to the kinetochores, we should not observe metaphase arrest when the fusion is overexpressed in ndc10-1 cells at 37°C. To test this prediction, we released ndc10-1 cells carrying PGAL1-MAD2-MAD3 from a G1 arrest at 25°C or 37°C and monitored the level of securin. In cells that were released into glucose-containing medium (to inhibit expression of Mad2-Mad3) with benomyl and nocodazole (to depolymerize microtubules) at 37°C, securin levels rose and fell, showing that these cells failed to activate the spindle checkpoint (Figure 3B) and confirming previous reports that kinetochores are required for normal checkpoint activation [21, 22]. In contrast, when these cells overexpressed the Mad2-Mad3 fusion because we released them into galactose-containing medium at 37°C, securin was stabilized (Figure 3B). These results show that the Mad2-Mad3 fusion can arrest cells in metaphase even in the absence of functional kinetochores, supporting the idea that the fusion protein is acting downstream of the events at the kinetochores and directly inducing metaphase arrest without disrupting microtubule attachments to chromosomes.
Metaphase arrest by Mad2-Mad3 fusions does not require other checkpoint proteins
The Mad2-Mad3 fusion arrests cells in metaphase even when kinetochores are not assembled, showing that it bypasses at least one step of the normal spindle checkpoint and prompting us to ask whether other checkpoint components are similarly dispensable. We integrated the PGAL1-MAD2-MAD3 construct into yeast strains with different spindle checkpoint genes deleted (mad1Δ, mad2Δ, mad3Δ, bub1Δ, and bub3Δ) and tested the effect of expressing Mad2-Mad3 by releasing the cells from G1 arrest into galactose-containing medium. In all cases, more than 70% of the population accumulated as large budded cells, indicating that the ability of the Mad2-Mad3 fusion to induce metaphase arrest does not require the presence of these checkpoint proteins (Figure 4A). We noticed that bub1Δ and bub3Δ strains had a slightly lower percentage of large budded cells after 3 hours. The most likely explanation is that bub1Δ and bub3Δ cells grow more slowly and have a higher rate of death than wild type cells because of aneuploidy due to their high chromosome loss rate [24, 25].
Figure 4.
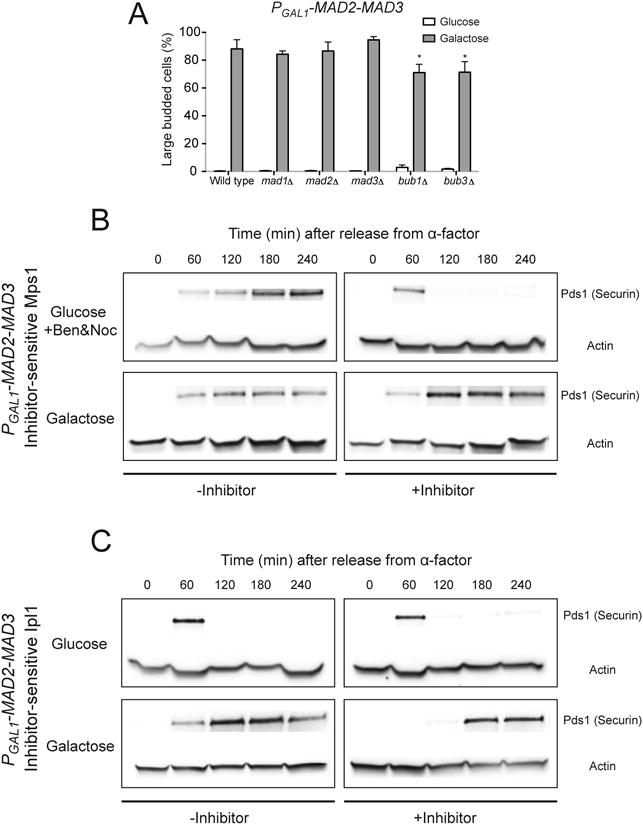
Metaphase arrest by Mad2-Mad3 fusions does not require other checkpoint components. (A) Cells with PGAL1-MAD2-MAD3 and deletion of the indicated checkpoint genes were released from G1 arrest into glucose- or galactose-containing media. The percentage of large budded cells was determined by light microscopy after 3 hours of growth. Error bars represent the standard deviation of three independent trials. Two hundred cells were counted for each trial. Asterisks indicate significant difference from wild-type control (*P < 0.05; two-tailed Student's t test). (B) Metaphase arrest by the Mad2-Mad3 fusion does not require Mps1 function. Cell cycle progression of cells with PGAL1-MAD2-MAD3 and mps1-as1 was monitored by Western blotting (n=3). Cells were released from G1 arrest into media with either glucose with benomyl and nocodazole (top) or galactose (bottom), in the absence (-Inhibitor) or presence (+Inhibitor) of 1NM-PP1, an inhibitor of the analog-sensitive Mps1. Western blots against Myc or actin (loading control) were performed. Both securin and Mps1-as1 are tagged with Myc in the strain, but only the bands corresponding to Myc-tagged securin are shown in the figure. Securin was stabilized in galactose-containing medium even in the presence of inhibitor, indicating that Mps1 activity is not needed for the metaphase arrest. The cause of the delay in accumulation of securin in cells grown in medium with galactose and the inhibitor is not known. (C) The Mad2-Mad3 fusion does not require Ipl1 activity to induce metaphase arrest. The cell cycle progression of cells with PGAL1-MAD2-MAD3 and ipl1-as5 was measured by Western blots (n=3). Cells were released from G1 arrest into media with either glucose (top) or galactose (bottom), in the absence (-Inhibitor) or presence (+Inhibitor) of 1NA-PP1, an inhibitor of the analog-sensitive Ipl1. Western blots against Myc (to visualize Myc-tagged securin) or actin (loading control) were performed. Securin was stabilized in galactose-containing medium in the presence of inhibitor, showing that Ipl1 activity is not needed for the metaphase arrest. The cause of the delay in accumulation of securin in cells grown in medium with galactose and the inhibitor is not known.
Next we tested the requirement for two components of the spindle checkpoint, Mps1 and Ipl1, that have other essential functions. Mps1 is a kinase that is required for the spindle checkpoint, duplication of the spindle pole bodies (SPBs), and mitotic spindle assembly and function [26, 27]. Ipl1/Aurora B is another protein kinase, which is required to activate the spindle checkpoint in the absence of mechanical tension at the kinetochores [5, 28]; it is also important for other processes such as spindle disassembly [29]. Since both Mps1 and Ipl1 are essential for cell viability, we used conditional alleles to inhibit their activities to ask whether they are required for the Mad2-Mad3 fusion to arrest cells. For Mps1, we used the analog-sensitive allele mps1-as1 [27], which contains an enlarged ATP-binding pocket that makes this engineered kinase uniquely sensitive to a bulky protein kinase inhibitor [30]. In the absence of the inhibitor, such engineered kinases are functional, and in its presence, they are the only protein kinase whose activity is inhibited. We released mps1-as1 cells carrying PGAL1-MAD2-MAD3 from G1 arrest into media with or without the inhibitor (1NM-PP1, 1-(1, 1-dimethylethyl)-3-(1-naphthalenylmethyl)-1H-pyrazolo[3, 4-d]pyrimidin-4-amine) and monitored securin levels. Cells treated with the inhibitor that were grown in glucose-containing medium (to inhibit expression of Mad2-Mad3) with benomyl and nocodazole (to depolymerize microtubules) showed the normal rise and fall in securin (Figure 4B). This result confirmed that Mps1-as1 cannot function in the presence of the inhibitor and that cells normally cannot activate the spindle checkpoint in the absence of Mps1 activity. On the other hand, cells that were grown in galactose-containing medium (to express Mad2-Mad3) stabilized securin even in the presence of inhibitor (Figure 4B), indicating strong inhibition of APC activity by the Mad2-Mad3 fusion, despite the inactivation of Mps1. We performed similar experiments to test the requirement for Ipl1 by using the analog-sensitive allele, ipl1-as5 [28]. We released ipl1-as5 cells carrying PGAL1-MAD2-MAD3 from G1 arrest into media with or without a slightly different inhibitor (1NA-PP1, 1-(1,1-dimethylethyl)-3-(1-naphthalenyl)-1H-pyrazolo[3, 4-d]pyrimidin-4-amine). Overexpressing the Mad2-Mad3 fusion stabilized securin even when Ipl1 was inactivated by adding the inhibitor (Figure 4C). We confirmed that Ipl1 activity in the strain is indeed sensitive to the inhibitor as they did not proliferate in the presence of 1NA-PP1 (data not shown). Our results thus show that both Mps1 and Ipl1 activity are dispensable for metaphase arrest by the Mad2-Mad3 fusion.
Directly tethering Mad2 to Cdc20 induces metaphase arrest
Why does fusing Mad2 to Mad3 activate the checkpoint when the overexpression of the two separate proteins does not? One possibility is that each protein binds independently but weakly to Cdc20, whereas activation of the checkpoint requires stable binding of Mad2 or Mad3 to the Cdc20. In this scenario, the Mad2-Mad3 fusion would bind Cdc20 strongly since it would have a higher avidity for Cdc20 compared to Mad2 or Mad3 alone. Both the Mad2 and Mad3 region of the fusion protein can bind Cdc20, and either the Mad2-Cdc20 or Mad3-Cdc20 interactions would prevent the Mad2-Mad3 fusion from completely dissociating from Cdc20. Thus the fusion protein would bind Cdc20 tightly and inhibit APCCdc20 activity even in the absence of other checkpoint components. This model also explains how Mad2 and Mad3 could associate with Cdc20 in budding yeast throughout the cell cycle [12, 15] without activating the checkpoint until some signals from the checkpoint increased the strengths of the interactions between Mad2, Mad3, and Cdc20.
If the stable binding of Mad2-Mad3 to Cdc20 causes metaphase arrest, we should be able to induce a similar phenotype by artificially tethering Mad2 or Mad3 to Cdc20 to form a tight complex. To test this hypothesis, we investigated the effect of heterodimerizing Mad2 or Mad3 with Cdc20 using the engineered leucine zipper system [16]. We fused the endogenous CDC20 to a negatively charged leucine zipper (making PCDC20-EEzip-CDC20 as the only copy of CDC20 in the cell) and integrated a construct with either MAD2 or MAD3 fused to a positively charged leucine zipper (RRzip) under the GAL1 promoter (while leaving the endogenous MAD2 and MAD3 loci intact). We released the cells from G1 arrest into galactose-containing medium to overexpress the checkpoint fusion constructs, which would then be tethered to EEzip-Cdc20. In cells with PCDC20-EEzip-CDC20 and PGAL1-MAD3-RRzip, the majority of the population continued to cycle when they were grown in medium with galactose, showing that binding of Mad3 to Cdc20 alone was unable to stop the cell cycle (Figure 5A). In contrast, around 90% of cells expressing both PCDC20-EEzip-CDC20 and PGAL1-MAD2-RRzip arrested in mitosis (Figure 5A). The result shows that simply tethering Mad2 to Cdc20 leads to strong metaphase arrest. To confirm that the arrest is due to binding of Mad2 to Cdc20 via the leucine zippers, we performed the same experiment with a strain that expresses untagged Cdc20 in addition to EEzip-Cdc20 and Mad2-RRzip. In this case, the cells continued to cycle even in galactose-containing medium (Figure 5A), showing that the metaphase arrest is due to direct binding of Mad2 to Cdc20 and can be overcome by Cdc20 that is not tethered to Mad2. The normal cell cycle observed in cells with free (untagged) Cdc20 also suggests that tethering Mad2 to Cdc20 does not have any obvious detrimental effects besides inhibition of APC activity.
Figure 5.
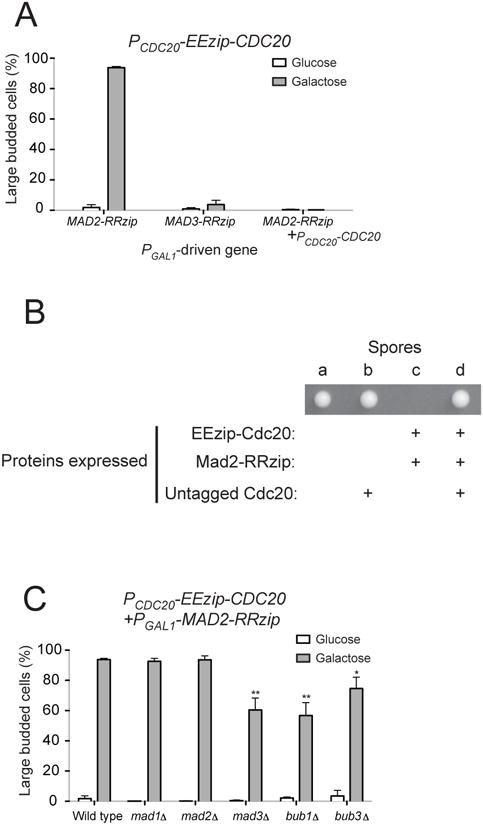
Tethering Mad2 directly to Cdc20 leads to metaphase arrest. (A) Cells with PCDC20-EEzip-CDC20 and the indicated PGAL1-driven genes were released from G1 arrest into glucose- or galactose-containing media. The percentage of large budded cells was determined by light microscopy after 3 hours of growth. Error bars represent the standard deviation of three independent trials. Two hundred cells were counted for each trial. Cells in the last column contain PCDC20-EEzip-CDC20, PGAL1-MAD2-RRzip, and an untagged copy of Cdc20 (PCDC20-CDC20) and thus cannot be arrested in mitosis by expressing Mad2-RRzip. (B) Cells expressing EEzip-Cdc20 and Mad2-RRzip from the endogenous CDC20 and MAD2 promoters display growth defects that can be overcome by expressing untagged Cdc20. Diploids that are heterozygous for three manipulated genes, PCDC20-EEzip-CDC20, PMAD2-MAD2-RRzip, and PCDC20-CDC20@URA3, were sporulated and a total of 15 tetrads were dissected (also see Table 1). Representative image of a tetrad on rich, glucose-containing plate after 2 days of growth at 30°C is shown. The “+” signs indicate proteins that are expressed based on the genotypes of each spore, which were determined by replica plating the tetrad onto dropout or drug plates. The genotype of spore c, which failed to form visible colonies, was inferred from the genotypes of other spores from the same tetrad. (C) The metaphase arrest produced by tethering Mad2 to Cdc20 does not require other checkpoint components (also see Figure S1). Cells with PCDC20-EEzip-CDC20, PGAL1-MAD2-RRzip, and deletion of the indicated checkpoint genes were released from G1 arrest into glucose-or galactose-containing media. The percentage of large budded cells was determined by light microscopy after 3 hours of growth. Error bars represent the standard deviation of three independent trials. Two hundred cells were counted for each trial. Asterisks indicate significant difference from wild-type control (*P < 0.05 and **P < 0.01; two-tailed Student's t test).
We then asked what would happen if Mad2-RRzip is expressed from the MAD2 promoter instead of being overexpressed. Since the level of Mad2 in budding yeast exceeds that of Cdc20 [31], all Cdc20 should be tethered by Mad2 when both EEzip-Cdc20 and Mad2-RRzip are expressed from their endogenous promoters. We mated haploid cells carrying both PCDC20-EEzip-CDC20 and PCDC20-CDC20 (at the URA3 locus) with cells carrying PMAD2-MAD2-RRzip. We then sporulated the diploids and dissected the tetrads to look at viability of the spores. Cells expressing both EEzip-Cdc20 and Mad2-RRzip failed to form visible colonies or only formed very small colonies (Figure 5B and Table 1). When the small colonies were examined microscopically, they were mostly made up of mitotically arrested cells (data not shown), suggesting that the cells had trouble progressing through mitosis. On the other hand, cells that expressed untagged Cdc20 in addition to EEzip-Cdc20 and Mad2-RRzip showed normal growth (Figure 5B and Table 1). Our results show that the metaphase arrest seen in our earlier experiments is not simply due to a high, non-physiological level of the fusion proteins, since wild type expression of Mad2 is sufficient to arrest cells when Mad2 is constitutively tethered to Cdc20.
Table 1. Colony size of spores with indicated genotypes from tetrad dissection (also see Figure 5B).
The genotypes of the spores that failed to form colonies or formed very small colonies were inferred from the genotypes of the other spores assuming that all three heterozygously modified genes (PCDC20-EEzip-CDC20, PMAD2-MAD2-RRzip, and PCDC20-CDC20@URA3) showed Mendelian (2:2) segregation.
| Colony size | ||
|---|---|---|
| Genotype | Very Small/No colony | Regular |
| PCDC20-EEzip-CDC20 PMAD2-MAD2-RRzip | 7 | 0 |
| PCDC20-EEzip-CDC20 PMAD2-MAD2-RRzip PCDC20-CDC20@URA3 | 0 | 12 |
Number of tetrads dissected : 15
Since Cdc20 is the target of the spindle checkpoint, the stable binding of Mad2 to Cdc20 should be the last step in spindle checkpoint activation. If the hypothesis is correct, the arrest induced by tethering Mad2 to Cdc20 will not require other checkpoint components. To test this prediction, we introduced PGAL1-MAD2-RRzip and PCDC20-EEzip-CDC20 into yeast strains with different checkpoint genes deleted (mad1Δ, mad2Δ, mad3Δ, bub1Δ, and bub3Δ). We looked at the effect of tethering Mad2 to Cdc20 by releasing these cells from G1 arrest into galactose-containing medium. In all five checkpoint mutants, at least 55% of the population accumulated as large budded cells, showing that the metaphase arrest does not require the presence of these checkpoint proteins (Figure 5C). We noted that the percentages of metaphase-arrested cells were significantly lower in mad3Δ, bub1Δ, and bub3Δ strains. The smaller number of large budded cells in bub1Δ and bub3Δ strains is again probably due to the growth defects exhibited by these strains. The weaker phenotype in mad3Δ cells, on the other hand, may suggest a role of Mad3 in strengthening the inhibition of Cdc20 by Mad2. The metaphase arrest also does not require Mps1, Ipl1, or functional kinetochores (Figure S1), further suggesting that tethering Mad2 to Cdc20 recapitulates a downstream event in checkpoint activation and leads to direct inhibition of APC.
The phenotypes of tethering Mad2 mutants to Cdc20 support the Mad2-template model
The phenotypes we produced by tethering Mad2 to Cdc20 can be rationalized by the Mad2-template model. This model is based on the structures of different conformations of Mad2 [32-35], structural analysis of the Mad1-Mad2 complex [34], and imaging the dynamics of checkpoint proteins at the kinetochore [36-40]. In the model, Mad1 dimers associate with unattached kinetochores and bind Mad2 (Figure 6A). This leads to the formation of Mad1-Mad2 complexes with Mad2 in the “closed” conformation (C-Mad2), which wraps around Mad1 or Cdc20. The complex in turn recruits a different conformer of Mad2, “open” Mad2 (O-Mad2), and facilitates its association with Cdc20 and conversion into closed Mad2. Mad2 can associate with Cdc20 throughout the budding yeast cell cycle [15], but this is insufficient to activate the checkpoint, likely because the interaction is too short-lived to allow the conformational change that is required to generate the closed Mad2-Cdc20 complex from open Mad2. The Mad1-Mad2 complex is therefore required to increase the rate of Mad2 conversion and produce the closed Mad2-Cdc20 complex [41]. The model predicts that if Mad2 can stably associate with Cdc20, it can eventually reach the closed conformation and inhibit APCCdc20 even in the absence of attachment errors or other checkpoint proteins. Our observation that tethering Mad2 to Cdc20 can directly induce metaphase arrest is consistent with the model.
Figure 6.

. Metaphase arrest induced by tethering Mad2 mutants to Cdc20 supports the Mad2-template model. (A) The Mad2-template model (adapted from [6]). Mad1 dimers associate with unattached kinetochores and bind Mad2, converting them from “open” (O-Mad2) to “closed” (C-Mad2) conformation. The Mad1-Mad2 complexes at the kinetochores (the “templates”) then recruit additional open Mad2, allowing the formation of closed Mad2-Cdc20 complexes. (B) The behavior of Mad2 mutants in the context of the Mad2-template model. (Top) Mad2ΔC lacks the C-terminal amino acid residues and cannot convert to the closed Mad2 conformation. It fails to activate the spindle checkpoint since it is unable to form a stable complex with Mad1 and to bind to and inhibit Cdc20. (Bottom) The double point mutant Mad2RQEA carries the mutations Arg126-Glu and Gln127-Ala. The changes inhibit the binding between free Mad2 and closed conformation of Mad2 found in the Mad1-Mad2 complex, which inactivates the spindle checkpoint by preventing the formation of C-Mad2-Cdc20 complexes. The mutations also affect the interaction of Mad2 with BUBR1 (mammalian version of Mad3) and the formation of stable MCC. (C) Effects of tethering Mad2 mutants to Cdc20. Cells with PCDC20-EEzip-CDC20 and the indicated PGAL1-driven genes were released from G1 arrest into glucose- or galactose-containing media. The percentage of large budded cells was determined by light microscopy after 3 hours of growth. Error bars represent the standard deviation of three independent trials. Two hundred cells were counted for each trial. Tethering the Mad2 mutant that can reach the closed conformation, but cannot induced conformational conversion in other Mad2 molecules (Mad2RQEA), does activate the checkpoint, but tethering the mutant that cannot achieve the closed conformation (Mad2ΔC) does not.
To further test the Mad2-template model, we tethered two previously studied Mad2 mutants to Cdc20. One mutant is Mad2 lacking its C-terminal 10 amino acid residues and was first characterized in HeLa cells [39]. Without this region, Mad2 is unable to close. The Mad2 mutant cannot activate the checkpoint as it fails to form a stable complex with Mad1 or to bind to and inhibit Cdc20 (Figure 6B). The corresponding Mad2 mutant in budding yeast (MAD2ΔC) also has no checkpoint function [42]. We integrated PGAL1-MAD2ΔC-RRzip into cells expressing EEzip-Cdc20. When the cells were grown in galactose-containing medium, the majority of the population continued to cycle (Figure 6C). Thus tethering Mad2ΔC to Cdc20 is unable to induce metaphase arrest, which supports the notion that only closed Mad2 can inhibit Cdc20.
Next we tested the Mad2 double point mutant Arg126-Glu/Gln127-Ala (Mad2RQEA). These two mutated residues have been conserved in eukaryotic evolution and are essential for the binding of soluble Mad2 to the closed conformation of Mad2 found in the Mad1-Mad2 complex [39, 42] (Figure 6B). Since this interaction is important for facilitating the formation of closed Mad2-Cdc20 complex, this Mad2 mutant does not have normal checkpoint function in either budding yeast [42] or HeLa cells [39]. These residues have also been shown to be important for the binding of Mad2 to BUBR1, the mammalian equivalent of Mad3, and the formation of MCC in vitro [43]. To test the effect of tethering Mad2RQEA to Cdc20, we integrated PGAL1-MAD2RQEA-RRzip into cells expressing EEzip-Cdc20. When the cells were released into galactose-containing medium, 95% of them accumulated at the large-budded state (Figure 6C). The result suggests that when Mad2 is directly tethered to Cdc20, the interaction between closed and open Mad2 is dispensable since Mad2 can eventually close and inhibit Cdc20, which is again consistent with the Mad2-template model. Since human Mad2RQEA fails to bind to BUBR1, our experiment strengthens the conclusion that the binding between Mad2 and Mad3 and the formation of stable MCC is not necessary for the metaphase arrest induced by tethering Mad2 to Cdc20.
Discussion
We showed that expressing a Mad2-Mad3 protein fusion arrests budding yeast in metaphase. The arrest does not require other checkpoint proteins and is not due to disruption of microtubule attachments to kinetochores. We obtained similar results by non-covalently linking Mad2 to Mad3 using leucine zippers, indicating that constitutive association between Mad2 and Mad3 is sufficient to prevent progression through mitosis. Finally, we showed that directly tethering Mad2 to Cdc20 also arrests cells in metaphase and that this arrest is independent of other checkpoint proteins. Our results support a model in which Mad2 and Mad3 are the most downstream components of the checkpoint pathway and cooperate to bind to Cdc20 and inhibit the APC.
Linking Mad2 and Mad3 arrests cells in the absence of spindle damage
A high level of Mad2 protein arrests cells in metaphase in a variety of organisms including Xenopus embryos [44, 45], fission yeast [46], and tissue culture cells [36, 39, 47]. The exact mechanism leading to the arrest likely differs between organisms; the arrest only requires Mad3 in fission yeast [48] and is independent of Mad1 in Xenopus embryos [44], but it requires Mad1 in tissue culture cells [47]. Our experiments show that in budding yeast, a high level of Mad2 alone is not enough to induce metaphase arrest, whereas expression of physically-linked Mad2 and Mad3 arrests cells independently of other checkpoint components. The results in different species may reflect differences in the interactions of Mad2 with other checkpoint proteins, the maximum expression of Mad2 that can be obtained, or the relative importance of Mad2 in checkpoint activation.
Mps1 overexpression in budding yeast can activate the spindle checkpoint without disrupting the mitotic spindle [49]. Unlike the metaphase arrest caused by linking Mad2 to Mad3, the effect of Mps1 overexpression depends on other checkpoint proteins; checkpoint mutants overexpressing Mps1 progress through mitosis without significant delay. Mps1 therefore likely represents an upstream activator that coordinates with other checkpoint proteins to activate the spindle checkpoint, while the Mad2-Mad3 fusion acts as a downstream effector that inhibits APCCdc20 even in the absence of other checkpoint proteins and functional kinetochores. Unlike many other checkpoint complexes previously identified in vivo [11-13], the Mad2-Mad3 fusion represents a minimal complex that can induce metaphase arrest independently of other known checkpoint components. Because we inactivated the other checkpoint proteins individually to test their requirements for arrest by Mad2-Mad3 fusions, we cannot rigorously exclude the possibility that two or more of them play a redundant role in helping the fusion to inhibit Cdc20, but we believe that Mad2-Mad3 is a direct inhibitor of APCCdc20 and functions downstream of events at the kinetochore.
Stable binding of Mad2 to Cdc20 can lead to metaphase arrest
We believe the Mad2-Mad3 fusion arrests cells because its high avidity for Cdc20 allows Mad2 to stay in close proximity to Cdc20 for long enough for Mad2 to adopt the closed conformation and inhibit Cdc20. Tethering Mad2 to Cdc20 using leucine zippers arrests cells in metaphase, supporting the hypothesis. Expressing both Mad2 and Cdc20 fused to leucine zippers from their endogenous promoters is sufficient to induce metaphase arrest in cells, showing that a physiological level of Mad2 can inhibit Cdc20 if the two proteins are forced to stably associate with each other. The inhibition by Mad2 is likely direct, as all other known checkpoint proteins are dispensable for the arrest. We noticed that the metaphase arrest is weaker, but not absent, in mad3Δ cells. This result suggests that when Mad2 is stably associated with Cdc20, it can directly inhibit Cdc20 and recruit Mad3 to further strengthen the inhibition, and that while the recruitment of Mad3 potentiates Cdc20 inhibition, it is not essential to inactivate the APC in a significant fraction of cells. Consistent with this claim, most known checkpoint proteins, including Mad2, are necessary for the stable interaction between Cdc20 and Mad3 in budding yeast, whereas the binding of Mad2 to Cdc20 only requires Mad1 and Mps1 [12, 13, 15]. Our results support the hypothesis that Mad2 and Mad3 cooperate to bind to and inhibit Cdc20, and suggest that this event represents the last and essential step in spindle checkpoint activation. We believe that Mad3 has an auxiliary role and becomes dispensable when Mad2 can constitutively associate with Cdc20.
Tethering Mad3 to Cdc20 alone is unable to induce metaphase arrest in cells. We favor the interpretation that the main role of Mad3 is to promote the inhibition of Cdc20 by Mad2. In contrast, in vitro experiments have shown that BUBR1, the mammalian version of Mad3, could inhibit APCCdc20 alone and also act synergistically with Mad2 to repress APC activity [50, 51]. In budding yeast, Mad3 can also inhibit APCCdc20 in vitro in the absence of added Mad2 [Schuyler S, personal communication]. Several factors could account for the discrepancies between the in vitro data and our observations. The in vitro experiments, which contain reticulocyte lysate and APC purified from yeast or mammalian cells, may be contaminated by a low level of Mad2 that complicated the results. We also cannot exclude the possibility that inhibition of Cdc20 by Mad3 requires a specific orientation of the two proteins that cannot be achieved when they are tethered together by leucine zippers.
Implications for the Mad2-template model
The Mad2-template model is a prominent model that explains how the checkpoint proteins respond to events at the kinetochore and activate the spindle checkpoint. The model predicts that the requirements for kinetochores and other checkpoint proteins can be bypassed if Mad2 can stably associate with Cdc20, which is consistent with our result that tethering Mad2 to Cdc20 can directly induce metaphase arrest. The phenotypes observed when we tethered two known Mad2 mutants (Mad2ΔC and Mad2RQEA) to Cdc20 further support the Mad2-template model and strengthen our claim that the binding of Mad2 to Cdc20 is the most downstream event in checkpoint activation.
One extension from the Mad2-template model is that the closed Mad2-Cdc20 complex can recruit open Mad2 and trigger the production of additional closed Mad2-Cdc20, thereby amplifying the checkpoint signal [6]. When we tethered Mad2 to Cdc20 in cells that also express untagged Cdc20 (which cannot be tethered), the cells failed to arrest in metaphase, indicating that the Mad2-Cdc20 complex is unable to inhibit the untagged Cdc20. Amplification from closed Mad2-Cdc20 complexes is therefore unlikely to be a factor in further amplifying the checkpoint signal, and alternative mechanisms are required to ensure complete inhibition of APCCdc20 during normal checkpoint activation. Our results in cells with untagged Cdc20 also argue that tethering Mad2 to Cdc20 leads to direct inhibition of APC and does not have any obvious side effects as these cells progressed through the cell cycle normally.
Overall our results support the model that Mad2-Mad3 fusions and the association of Mad2 with Cdc20 inhibit APC activity by acting downstream of all other known checkpoint components. The two systems represent new ways for studying APC inhibition in vivo independently of other checkpoint proteins and upstream kinetochore signals, which may allow us to better understand the molecular details of spindle checkpoint activation.
Experimental Procedures
Yeast strains and methods
Strains used in this study are listed in Table S1. All strains are derivatives of W303 (ade2-1 his3-11,15 leu2-3,112 trp1-1 ura3-1 can1-100). Strains were constructed using standard genetic techniques. All media were prepared using established recipes [52], and contain 2% wt/vol of the indicated sugar as the carbon source. To prepare media containing benomyl and nocodazole, DMSO stocks of methyl 1-(butylcarbamoyl)-2-benzimidazolecarbamate (benomyl) (Sigma-Aldrich, St. Louis, MO) and nocodazole (Sigma-Aldrich, St. Louis MO) were added to yeast extract and peptone (YEP) with 2% wt/vol glucose to a final concentration of 30μg/ml of each drug. Diploid strains were sporulated in liquid culture by growing to saturation in YEP with 2% wt/vol glucose, diluting into YEP with 2% wt/vol potassium acetate for 12 hours at 30°C, washing with water, and resuspending in 2% potassium acetate at 25°C.
Cell cycle analysis by light microscopy
To look at the effect of overexpressing different checkpoint constructs on cell cycle progression, cells were first grown to mid-log phase (107 cells/ml) in YEP with 2% raffinose (wt/vol). Cells were then arrested in G1 by adding 10μg/ml α-factor (Bio-Synthesis, Lewisville, TX) and incubated for 2 hours at 30oC. Cells were washed four times to remove α-factor and resuspended in YEP with either 2% glucose (wt/vol) or 2% galactose (wt/vol). After growing for 3 hours at 30°C, the cultures were briefly sonicated to separate cells that fail to dissociate completely after division and the percentage of large-budded cells in each sample was determined by light microscopy. For the time course experiment in Figure 2B, the cultures were handled the same way except samples were taken every 30 min after releasing from G1 arrest and counted.
Cell cycle analysis by Western blots
To monitor cell cycle progression by Western blots, cells were grown and arrested in G1 as described above, and released into the indicated media. 1μg/ml α-factor was used for bar1Δ strains. For experiments with ndc10-1 strains, cells were grown to mid-log phase (107 cells/ml) in YEP with 2% raffinose (wt/vol) at 25°C. Cells were then arrested in G1 by adding 10μg/ml α-factor and incubated for 2 hours at 25°C, and shifted to 37°C for 30 min to inactivate ndc10-1. Cells were washed four times to remove α-factor and resuspended in the indicated media at 37°C. 10μg/ml α-factor was added at 60 min after release from G1 arrest in all Western blot experiments to prevent cells from progressing into the next S phase.
For experiments with mps1-as1 strains, DMSO (-Inhibitor) or 10μM of 1NM-PP1 (+Inhibitor) was added to the media after releasing the cells from G1 arrest to inhibit the activity of Mps1-as1. For experiments with ipl1-as5 strains, DMSO (-Inhibitor) or 50μM of 1NA-PP1 (+Inhibitor) was added to inhibit the activity of Ipl1-as5.
In all Western blot experiments, 1ml samples of the culture were collected at the indicated time points, and the cells were pelleted by centrifugation for 1 min at room temperature. The supernatant was removed, and cell pellets were stored at -80°C.
Cell pellets were lysed using a NaOH/β-mercaptoethanol-based protocol [53]. Proteins samples were loaded onto and separated in 10% Criterion Tris-HCl polyacrylamide gel (Bio-Rad, Hercules, CA). Proteins were transferred overnight to nitrocellulose (Whatman, Picataway, NJ). Western blotting for Myc-tagged securin were performed using anti-Myc 9E10 antibodies (Roche Applied Science, Indianapolis, IN) at a 1:500 dilution, and actin was detected with anti-actin antibodies (Abcam, Cambridge, MA) used at a 1:2000 dilution. Horseradish peroxidase-conjugated goat anti-mouse (Santa Cruz Biotechnology, Santa Cruz, CA) was used as the secondary antibody at a 1:2000 dilution. The secondary antibody was detected by SuperSignal West Dura Chemiluminescent Substrate (Thermo Fisher Scientific, Rockford, IL) and the blot was imaged with an AlphaImager (ProteinSimple, Santa Clara, CA).
Fluorescence Microscopy
To look at cells arrested in metaphase by the Mad2-Mad3 fusions using live-cell microscopy, cells were synchronized in G1 with 1μg/ml α-factor and then subjected to a constant flow of indicated media for 3 hours at room temperature using the ONIX microfluidic perfusion platform (CellASIC, Hayward, CA). Fluorescence microscopy was performed using Nikon Ti-E inverted microscope (Nikon, Melville, NY) equipped with a 60× objective (PlanApo, numerical aperture 1.4, oil), GFP filter (Chroma Technology, Bellow Falls, VT), and a CoolSNAP charge-coupled device camera (Photometrics, Tucson, AZ). Z-stacks of 25 sections were acquired using exposure times of 350 ms in Metamorph (Molecular Devices, Sunnyvale, CA). Z-stacks were combined into a single maximum intensity projection with ImageJ (NIH).
Supplementary Material
Highlights.
-Expression of physically-linked Mad2 and Mad3 arrests cells in metaphase
-The metaphase arrest does not require other checkpoint components
-Tethering Mad2 to Cdc20 also arrests cells independently of other checkpoint proteins
Acknowledgments
We are grateful to S. Biggins, V. Denic, A. Hyman, and M. Winey for strains and plasmids and K. Shokat for generous gifts of reagents. We thank A. Amon, S. Biggins, K. Hardwick, A. Musacchio, E. Salmon, and members of the Murray Lab for reading and commenting on the manuscript. This work is supported by funds from the National Institutes of Health (GM043987) to A.W.M.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Li R, Murray AW. Feedback control of mitosis in budding yeast. Cell. 1991;66:519–531. doi: 10.1016/0092-8674(81)90015-5. [DOI] [PubMed] [Google Scholar]
- 2.Hoyt MA, Totis L, Roberts BT. S. cerevisiae genes required for cell cycle arrest in response to loss of microtubule function. Cell. 1991;66:507–517. doi: 10.1016/0092-8674(81)90014-3. [DOI] [PubMed] [Google Scholar]
- 3.Li X, Nicklas RB. Mitotic forces control a cell-cycle checkpoint. Nature. 1995;373:630–632. doi: 10.1038/373630a0. [DOI] [PubMed] [Google Scholar]
- 4.Stern BM, Murray AW. Lack of tension at kinetochores activates the spindle checkpoint in budding yeast. Curr Biol. 2001;11:1462–1467. doi: 10.1016/s0960-9822(01)00451-1. [DOI] [PubMed] [Google Scholar]
- 5.Biggins S, Murray AW. The budding yeast protein kinase Ipl1/Aurora allows the absence of tension to activate the spindle checkpoint. Genes and Development. 2001;15:3118–3129. doi: 10.1101/gad.934801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Musacchio A, Salmon ED. The spindle-assembly checkpoint in space and time. Nat Rev Mol Cell Biol. 2007;8:379–393. doi: 10.1038/nrm2163. [DOI] [PubMed] [Google Scholar]
- 7.Murray AW. A brief history of error. Nat Cell Biol. 2011;13:1178–1182. doi: 10.1038/ncb2348. [DOI] [PubMed] [Google Scholar]
- 8.Weiss E, Winey M. The Saccharomyces cerevisiae spindle pole body duplication gene MPS1 is part of a mitotic checkpoint. The Journal of Cell Biology. 1996;132:111–123. doi: 10.1083/jcb.132.1.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Biggins S, Severin FF, Bhalla N, Sassoon I, Hyman AA, Murray AW. The conserved protein kinase Ipl1 regulates microtubule binding to kinetochores in budding yeast. Genes and Development. 1999;13:532–544. doi: 10.1101/gad.13.5.532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Gillett ES, Espelin CW, Sorger PK. Spindle checkpoint proteins and chromosome-microtubule attachment in budding yeast. The Journal of Cell Biology. 2004;164:535–546. doi: 10.1083/jcb.200308100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Brady DM, Hardwick KG. Complex formation between Mad1p, Bub1p and Bub3p is crucial for spindle checkpoint function. Curr Biol. 2000;10:675–678. doi: 10.1016/s0960-9822(00)00515-7. [DOI] [PubMed] [Google Scholar]
- 12.Hardwick KG, Johnston RC, Smith DL, Murray AW. MAD3 encodes a novel component of the spindle checkpoint which interacts with Bub3p, Cdc20p, and Mad2p. The Journal of Cell Biology. 2000;148:871–882. doi: 10.1083/jcb.148.5.871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Fraschini R, Beretta A, Sironi L, Musacchio A, Lucchini G, Piatti S. Bub3 interaction with Mad2, Mad3 and Cdc20 is mediated by WD40 repeats and does not require intact kinetochores. EMBO J. 2001;20:6648–6659. doi: 10.1093/emboj/20.23.6648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sudakin V, Chan GK, Yen TJ. Checkpoint inhibition of the APC/C in HeLa cells is mediated by a complex of BUBR1, BUB3, CDC20, and MAD2. The Journal of Cell Biology. 2001;154:925–936. doi: 10.1083/jcb.200102093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hwang LH, et al. Budding yeast Cdc20: a target of the spindle checkpoint. Science. 1998;279:1041–1044. doi: 10.1126/science.279.5353.1041. [DOI] [PubMed] [Google Scholar]
- 16.Moll JR, Ruvinov SB, Pastan I, Vinson C. Designed heterodimerizing leucine zippers with a ranger of pIs and stabilities up to 10(-15) M. Protein Sci. 2001;10:649–655. doi: 10.1110/ps.39401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Straight AF, Belmont AS, Robinett CC, Murray AW. GFP tagging of budding yeast chromosomes reveals that protein-protein interactions can mediate sister chromatid cohesion. Curr Biol. 1996;6:1599–1608. doi: 10.1016/s0960-9822(02)70783-5. [DOI] [PubMed] [Google Scholar]
- 18.Goshima G, Yanagida M. Establishing biorientation occurs with precocious separation of the sister kinetochores, but not the arms, in the early spindle of budding yeast. Cell. 2000;100:619–633. doi: 10.1016/s0092-8674(00)80699-6. [DOI] [PubMed] [Google Scholar]
- 19.He X, Asthana S, Sorger PK. Transient sister chromatid separation and elastic deformation of chromosomes during mitosis in budding yeast. Cell. 2000;101:763–775. doi: 10.1016/s0092-8674(00)80888-0. [DOI] [PubMed] [Google Scholar]
- 20.Tanaka T, Fuchs J, Loidl J, Nasmyth K. Cohesin ensures bipolar attachment of microtubules to sister centromeres and resists their precocious separation. Nat Cell Biol. 2000;2:492–499. doi: 10.1038/35019529. [DOI] [PubMed] [Google Scholar]
- 21.Tavormina PA, Burke DJ. Cell cycle arrest in cdc20 mutants of Saccharomyces cerevisiae is independent of Ndc10p and kinetochore function but requires a subset of spindle checkpoint genes. Genetics. 1998;148:1701–1713. doi: 10.1093/genetics/148.4.1701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Fraschini R, Beretta A, Lucchini G, Piatti S. Role of the kinetochore protein Ndc10 in mitotic checkpoint activation in Saccharomyces cerevisiae. Mol Genet Genomics. 2001;266:115–125. doi: 10.1007/s004380100533. [DOI] [PubMed] [Google Scholar]
- 23.Goh PY, Kilmartin JV. NDC10: a gene involved in chromosome segregation in Saccharomyces cerevisiae. The Journal of Cell Biology. 1993;121:503–512. doi: 10.1083/jcb.121.3.503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hughes TR, et al. Widespread aneuploidy revealed by DNA microarray expression profiling. Nat Genet. 2000;25:333–337. doi: 10.1038/77116. [DOI] [PubMed] [Google Scholar]
- 25.Warren CD, Brady DM, Johnston RC, Hanna JS, Hardwick KG, Spencer FA. Distinct chromosome segregation roles for spindle checkpoint proteins. Mol Biol Cell. 2002;13:3029–3041. doi: 10.1091/mbc.E02-04-0203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Winey M, Huneycutt BJ. Centrosomes and checkpoints: the MPS1 family of kinases. Oncogene. 2002;21:6161–6169. doi: 10.1038/sj.onc.1205712. [DOI] [PubMed] [Google Scholar]
- 27.Jones MH, et al. Chemical genetics reveals a role for Mps1 kinase in kinetochore attachment during mitosis. Curr Biol. 2005;15:160–165. doi: 10.1016/j.cub.2005.01.010. [DOI] [PubMed] [Google Scholar]
- 28.Pinsky BA, Kung C, Shokat KM, Biggins S. The Ipl1-Aurora protein kinase activates the spindle checkpoint by creating unattached kinetochores. Nat Cell Biol. 2006;8:78–83. doi: 10.1038/ncb1341. [DOI] [PubMed] [Google Scholar]
- 29.Buvelot S, Tatsutani SY, Vermaak D, Biggins S. The budding yeast Ipl1/Aurora protein kinase regulates mitotic spindle disassembly. The Journal of Cell Biology. 2003;160:329–339. doi: 10.1083/jcb.200209018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Bishop AC, et al. A chemical switch for inhibitor-sensitive alleles of any protein kinase. Nature. 2000;407:395–401. doi: 10.1038/35030148. [DOI] [PubMed] [Google Scholar]
- 31.Poddar A, Stukenberg PT, Burke DJ. Two complexes of spindle checkpoint proteins containing Cdc20 and Mad2 assemble during mitosis independently of the kinetochore in Saccharomyces cerevisiae. Eukaryotic Cell. 2005;4:867–878. doi: 10.1128/EC.4.5.867-878.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Luo X, et al. Structure of the Mad2 spindle assembly checkpoint protein and its interaction with Cdc20. Nat Struct Biol. 2000;7:224–229. doi: 10.1038/73338. [DOI] [PubMed] [Google Scholar]
- 33.Luo X, Tang Z, Rizo J, Yu H. The Mad2 spindle checkpoint protein undergoes similar major conformational changes upon binding to either Mad1 or Cdc20. Molecular Cell. 2002;9:59–71. doi: 10.1016/s1097-2765(01)00435-x. [DOI] [PubMed] [Google Scholar]
- 34.Sironi L, Mapelli M, Knapp S, De Antoni A, Jeang KT, Musacchio A. Crystal structure of the tetrameric Mad1-Mad2 core complex: implications of a “safety belt” binding mechanism for the spindle checkpoint. EMBO J. 2002;21:2496–2506. doi: 10.1093/emboj/21.10.2496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Luo X, et al. The Mad2 spindle checkpoint protein has two distinct natively folded states. Nat Struct Mol Biol. 2004;11:338–345. doi: 10.1038/nsmb748. [DOI] [PubMed] [Google Scholar]
- 36.Howell BJ, Hoffman DB, Fang G, Murray AW, Salmon ED. Visualization of Mad2 dynamics at kinetochores, along spindle fibers, and at spindle poles in living cells. The Journal of Cell Biology. 2000;150:1233–1250. doi: 10.1083/jcb.150.6.1233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Howell BJ, Moree B, Farrar EM, Stewart S, Fang G, Salmon ED. Spindle checkpoint protein dynamics at kinetochores in living cells. Curr Biol. 2004;14:953–964. doi: 10.1016/j.cub.2004.05.053. [DOI] [PubMed] [Google Scholar]
- 38.Shah JV, Botvinick E, Bonday Z, Furnari F, Berns M, Cleveland DW. Dynamics of centromere and kinetochore proteins; implications for checkpoint signaling and silencing. Curr Biol. 2004;14:942–952. doi: 10.1016/j.cub.2004.05.046. [DOI] [PubMed] [Google Scholar]
- 39.De Antoni A, et al. The Mad1/Mad2 complex as a template for Mad2 activation in the spindle assembly checkpoint. Curr Biol. 2005;15:214–225. doi: 10.1016/j.cub.2005.01.038. [DOI] [PubMed] [Google Scholar]
- 40.Vink M, et al. In vitro FRAP identifies the minimal requirements for Mad2 kinetochore dynamics. Curr Biol. 2006;16:755–766. doi: 10.1016/j.cub.2006.03.057. [DOI] [PubMed] [Google Scholar]
- 41.Simonetta M, et al. The influence of catalysis on mad2 activation dynamics. PLoS Biol. 2009;7:e10. doi: 10.1371/journal.pbio.1000010. 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Nezi L, Rancati G, De Antoni A, Pasqualato S, Piatti S, Musacchio A. Accumulation of Mad2-Cdc20 complex during spindle checkpoint activation requires binding of open and closed conformers of Mad2 in Saccharomyces cerevisiae. The Journal of Cell Biology. 2006;174:39–51. doi: 10.1083/jcb.200602109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Tipton AR, et al. BUBR1 and closed MAD2 (C-MAD2) interact directly to assemble a functional mitotic checkpoint complex. J Biol Chem. 2011;286:21173–21179. doi: 10.1074/jbc.M111.238543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Chen RH, Shevchenko A, Mann M, Murray AW. Spindle checkpoint protein Xmad1 recruits Xmad2 to unattached kinetochores. The Journal of Cell Biology. 1998;143:283–295. doi: 10.1083/jcb.143.2.283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Fang G, Yu H, Kirschner MW. Direct binding of CDC20 protein family members activates the anaphase-promoting complex in mitosis and G1. Molecular Cell. 1998;2:163–171. doi: 10.1016/s1097-2765(00)80126-4. [DOI] [PubMed] [Google Scholar]
- 46.He X, Patterson TE, Sazer S. The Schizosaccharomyces pombe spindle checkpoint protein mad2p blocks anaphase and genetically interacts with the anaphase-promoting complex. Proc Natl Acad Sci U S A. 1997;94:7965–7970. doi: 10.1073/pnas.94.15.7965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Sironi L, Melixetian M, Faretta M, Prosperini E, Helin K, Musacchio A. Mad2 binding to Mad1 and Cdc20, rather than oligomerization, is required for the spindle checkpoint. EMBO J. 2001;20:6371–6382. doi: 10.1093/emboj/20.22.6371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Millband DN, Hardwick KG. Fission yeast Mad3p is required for Mad2p to inhibit the anaphase-promoting complex and localizes to kinetochores in a Bub1p-, Bub3p-, and Mph1p-dependent manner. Mol Cell Biol. 2002;22:2728–2742. doi: 10.1128/MCB.22.8.2728-2742.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Hardwick KG, Weiss E, Luca FC, Winey M, Murray AW. Activation of the budding yeast spindle assembly checkpoint without mitotic spindle disruption. Science. 1996;273:953–956. doi: 10.1126/science.273.5277.953. [DOI] [PubMed] [Google Scholar]
- 50.Tang Z, Bharadwaj R, Li B, Yu H. Mad2-Independent inhibition of APCCdc20 by the mitotic checkpoint protein BubR1. Developmental Cell. 2001;1:227–237. doi: 10.1016/s1534-5807(01)00019-3. [DOI] [PubMed] [Google Scholar]
- 51.Fang G. Checkpoint protein BubR1 acts synergistically with Mad2 to inhibit anaphase-promoting complex. Mol Biol Cell. 2002;13:755–766. doi: 10.1091/mbc.01-09-0437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Sherman F, Fink G, Lawerence C. Methods in Yeast Genetics. New York: Cold Spring Harbor Laboratory Press; 1974. [Google Scholar]
- 53.Kushnirov VV. Rapid and reliable protein extraction from yeast. Yeast. 2000;16:857–860. doi: 10.1002/1097-0061(20000630)16:9<857::AID-YEA561>3.0.CO;2-B. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.