

Abstract
Bifenthrin is one the most widespread pollutants and has caused potential effect on aquatic life and human health, yet little is known about microbial degradation in contaminated regions. A novel yeast strain ZS-02, isolated from activated sludge and identified as Candida pelliculosa based on morphology, API test and 18S rDNA gene analysis, was found highly effective in degrading bifenthrin over a wide range of temperatures (20–40°C) and pH (5–9). On the basis of response surface methodology (RSM), the optimal degradation conditions were determined to be 32.3°C and pH 7.2. Under these conditions, the yeast completely metabolized bifenthrin (50 mg·L−1) within 8 days. This strain utilized bifenthrin as the sole carbon source for growth as well as co-metabolized it in the presence of glucose, and tolerated concentrations as high as 600 mg·L−1 with a q max, K s and K i of 1.7015 day−1, 86.2259 mg·L−1 and 187.2340 mg·L−1, respectively. The yeast first degraded bifenthrin by hydrolysis of the carboxylester linkage to produce cyclopropanecarboxylic acid and 2-methyl-3-biphenylyl methanol. Subsequently, 2-methyl-3-biphenylyl methanol was further transformed by biphenyl cleavage to form 4-trifluoromethoxy phenol, 2-chloro-6-fluoro benzylalcohol, and 3,5-dimethoxy phenol, resulting in its detoxification. Eventually, no persistent accumulative product was detected by gas chromatopraphy-mass spectrometry (GC-MS) analysis. This is the first report of a novel pathway of degradation of bifenthrin by hydrolysis of ester linkage and cleavage of biphenyl in a microorganism. Furthermore, strain ZS-02 degraded a variety of pyrethroids including bifenthrin, cyfluthrin, deltamethrin, fenvalerate, cypermethrin, and fenpropathrin. In different contaminated soils introduced with strain ZS-02, 65–75% of the 50 mg·kg−1 bifenthrin was eliminated within 10 days, suggesting the yeast could be a promising candidate for remediation of environments affected by bifenthrin. Finally, this is the first described yeast capable of degrading bifenthrin.
Introduction
Synthetic pyrethroid insecticides are frequently and widely used worldwide. These kinds of pesticides are synthetic analogs of the naturally occurring toxin, derived from the flowers of Chrysanthemum cinerariaefolium [1]. They have been classified as type I or type II based on the toxic symptoms and the absence or presence of a cyano group at the carboxyl alpha position. Recently, with the restriction on the use of organophosphates and carbamates, pyrethroids have generally been regarded as the replacements [2]. At present, they are the dominant insecticides among retail sales to consumers [3], and their usage is expected to further increase [4], [5]. Due to these uses, pyrethroids have been reported in nearly all sediment samples tested from urban creeks [2], [3], [6], [7], [8].
The accumulation and widespread use of pyrethroids in agriculture have increased the public concern on potential human health risks that may result from chronic dietary exposure to pyrethroid residues on food [9], [10]. For instance, numerous studies have shown that they may have neurotoxicity [11], [12], immunotoxicity [13], [14], reproductive toxicity [5], [15], and genotoxicity [16], [17] on non-target organisms. In addition, some current-use pyrethroids have recently been listed as potential endocrine disruptors by the Environmental Protection Agency (EPA) of the USA [18]. All these findings indicated that pyrethroids might be potentially harmful to human health. It is thus critically essential to develop an efficient and cost-effective remediation strategy to eliminate pyrethroids from the environments and agricultural products.
Bifenthrin[2-methylbiphenyl-3-ylmethyl-(Z)-(1RS)-3-(2-chloro-3,3,3-trifluoroprop-1-enyl)-2,2-dimethylcyclopropane carboxylate] is one of the most popular pyrethroids effective against a broad spectrum of insect pests of economically important crops [5], [16]. It is also extensively used for the control of residential pests such as termites in urban areas [19]. Its half-life in soil is usually between 65 and 125 days, but can range from 2 weeks to over 1 year, depending on the soil type, moisture, pH, temperature, and other conditions [20]–[22]. The World Health Organization (WHO) has classified bifenthrin as Toxicity Class II moderately hazardous [23]. In urban sediments, bifenthrin contributed the most to the observed toxicity among pyrethroids [2], [6], [8]. Therefore, bifenthrin was the pyrethroid of greatest concern with regards to aquatic life toxicity and widespread occurrence in water and soil [2], [8].
Bifenthrin released to surface waters or sediments is subjected to hydrolysis, photodecomposition, volatilization, and aerobic degradation by microorganisms [20]. Microbial degradation is considered to be the most significant process determining the fate of bifenthrin and other pyrethroids in nature [24], and has received increasing attention as an effective, cheap, and safe approach to clean up contaminated environments [25]. To date, some pyrethroid-degrading microorganisms such as Bacillus cereus [26], Pseudomonas fluorescens [27], Micrococcus sp. CPN 1 [28], Sphingobium sp. JZ-2 [29], Serratia sp. JCN13 [30], Ochrobactrum tritici pyd-1 [31], and Cladosporium sp. HU [32] have been isolated. However, there was little information available on bifenthrin-degrading microorganism.
In the present study, a novel yeast Candida pelliculosa ZS-02 capable of degrading bifenthrin was isolated and characterized. The objective of this study was to optimize its degradation conditions, investigate its degradation pathway and evaluate its potential in bioremediation of bifenthrin-contaminated soils. Finally, obtained information illustrated that the isolated strain might have potential for use in bioremediation of bifenthrin-contaminated environments.
Materials and Methods
Chemicals and media
Technical grade bifenthrin (98% purity), cyfluthrin (95% purity), deltamethrin (98% purity), fenvalerate (91.2% purity), cypermethrin (92.9% purity), and fenpropathrin (93% purity) used in this study were obtained from Zhongshan Aestar Fine Chemical Inc., Ltd, China. Chromatographic grade acetonitrile were purchased from Sigma-Aldrich, USA. All other chemicals and solvents were purchased from Merck, Germany.
The mineral salt medium (MSM) containing (grams per liter) (NH4)2SO4, 2; MgSO4·7H2O, 0.2; CaCl2·2H2O, 0.01; FeSO4·7H2O, 0.001, Na2HPO4·12H2O, 1.5; and KH2PO4, 1.5; and yeast peptone dextrose (YPD) medium containing (grams per liter) yeast extract, 10; peptone, 20; and dextrose (or glucose), 20 were used for the isolation and cultivation of bifenthrin-degrading yeast, respectively. The final pH was adjusted to 7.2. Both media were autoclaved for 20 min at 121°C separately.
Isolation and screening of bifenthrin-degrading microorganisms
Activated sludge samples were collected as the inoculum from an aerobic pyrethroid-manufacturing wastewater treatment system located in Zhongshan (Guangdong, China), which had produced pyrethroids for many years. 30-mL of activated sludge was transferred into 250-mL Erlenmeyer flasks containing 50 mL sterilized MSM enrichment medium. Bifenthrin dissolved in acetone solution was added to a final concentration of 50 mg·L−1 as the sole carbon source. The enrichment culture was incubated for 7 days at 30±1°C with shaking at 150 rpm. A 5-mL from each enrichment culture was transferred into 50 mL of fresh enrichment medium containing 100 mg·L−1 of bifenthrin and incubated for another 7 days. Three additional successive transfers were made into media containing 200, 400, and 600 mg·L−1 of bifenthrin. The final cultures were serially diluted and spread on YPD agar plates. The plates were incubated for 5 days at 30°C, and colonies were picked and purified by re-streaking 3 times as described by Chen et al. [32], [33]. The abilities of isolates to degrade bifenthrin were determined by high performance liquid chromatography (HPLC) (Agilent, USA) according to Chen et al. [34].
Characterization and identification of the bifenthrin-degrading isolates
One bifenthrin-degrading isolate that showed highest degradation efficiency was selected for further study. The isolate was grown on YPD agar plates at 30°C for 5 days and its morphology was investigated with a light microscope (Olympus, Japan) and scanning electron microscope (XL-30 ESEM, Philips Optoelectronics Co., Ltd, Holland). Colony morphology was observed on YPD agar plates incubated at 30°C at 1, 3, 5, and 7 days according to Barnett et al. [35]. The isolate was also subjected to sugar fermentation pattern analysis using API 20C AUX system (bioMérieux, France) according to the instructions of the manufacturer.
The isolate was confirmed by 18S rDNA sequence analysis. Total genomic DNA was prepared according to standard methods [36]. The 18S rDNA gene was amplified with the yeast universal primers EF4 (5′–GGAAGGGRTGTATTTATTAG–3′) and EF3 (5′–TCCTAAATGACCAAGTTTG–3′) [35]. Amplification was carried out in 50 µL reaction mixture containing 5 µL of 10×Ex Tap reaction buffer, 1 µL of 2.5 mmol·L−1 dNTP, 1 µL of 10 µmol·L−1 each primer, 1 µL of genomic DNA, 0.5 µL of 5 U·µL−1 Ex Tap DNA polymerase and 40.5 µL of ultrapure water. Reaction conditions consisted of initial denaturation at 94°C for 5 min, followed by 35 cycles of denaturation at 94°C for 1 min, annealing at 48°C for 1 min, and extension at 72°C for 2 min, with the last cycle followed by a ten-minute extension at 72°C. Polymerase chain reaction (PCR) product containing the amplified 18S rDNA gene fragment was purified with QIAquick Gel Extraction Kit spin column (Guangzhou Heda Technology Co. Ltd., China), ligated to the linear vector pMD20-T (TaKaRa Biotechnology Co. Ltd., China), and transformed into competent Escherichia coli DH5α cells. Positive clones were screened and sent to Shanghai Invitrogen Technology Co. Ltd., China, for sequencing. The 18S rDNA gene sequences with 1,452 bp were deposited at the GenBank under the accession number JN700989. The resulting sequence was compared with the genes available in the GenBank nucleotide library by a BLAST search through the National Center for Biotechnology Information (NCBI) internet site. Multiple alignments of 18S rDNA were carried out using CLUSTALX 1.8.1 software, and phylogenesis was analyzed using MEGA 4.0 software. An unrooted tree was built using the neighbor-joining method [37].
Inoculum preparation
To prepare the inoculum, the pure culture, obtained from individual colonies, was grown in YPD medium for 5 days, harvested by centrifugation at 4000×g for 5 min, washed twice with 0.9% sterile normal saline and re-suspended in MSM to set an OD600 of 0.3 by a spectrophotometer (Shimadzu, Japan). One percent of this suspension (1.0×107 CFU·mL−1) was used as the inoculum for studies.
Optimization of the bifenthrin-degrading conditions
The preliminary study indicated that temperature, pH and inoculum were significant variables for the degradation of bifenthrin by the yeast. In single-factor experiments, we determined the optimal ranges of the three factors, temperature: 20–40°C, pH: 5–9, and inoculum: OD600 = 0.1–0.5. These specified ranges were used as the reference levels. Furthermore, response surface methodology (RSM) based on the Box-Behnken design was used to optimize the main and interactive effects of the important parameters which significantly influenced the bifenthrin degradation by the yeast [38]. A three-variable Box-Behnken design consisting of 15 experimental runs with three replicates at the center point was generated by statistic analysis system (SAS) software (Version 9.0). The symbols and levels of three independent variables are shown in Table 1. The dependent variable was the degradation of 50 mg·L−1 bifenthrin in MSM by the yeast after 5 days of culture. Experiment was conducted according to Randomized block design and data were analyzed by response surface regression procedure of the SAS software to fit the following quadratic polynomial equation (Eq.(1)).
| (1) |
where Y i is the predicted response, X i and X j are variables, b 0 is the constant, b i is the linear coefficient, b ij is the interaction coefficient, and b ii is the quadratic coefficient.
Table 1. Box-Behnken experimental design and the response of dependent variable for bifenthrin degradation.
| Response | ||||
| Run | X 1 | X 2 | X 3 | Bifenthrin degradation (%)Y ZS-02 |
| 1 | −1 | −1 | 0 | 55.5±1.5o |
| 2 | −1 | 1 | 0 | 65.2±0.6n |
| 3 | 1 | −1 | 0 | 67.6±0.3m |
| 4 | 1 | 1 | 0 | 74.1±0.8i |
| 5 | 0 | −1 | −1 | 72.3±0.5j |
| 6 | 0 | −1 | 1 | 75.5±0.4g |
| 7 | 0 | 1 | −1 | 75.2±0.4h |
| 8 | 0 | 1 | 1 | 77.8±0.2f |
| 9 | −1 | 0 | −1 | 68.4±1.2l |
| 10 | 1 | 0 | −1 | 79.5±0.4e |
| 11 | −1 | 0 | 1 | 70.6±0.5k |
| 12 | 1 | 0 | 1 | 80.3±0.5d |
| 13 | 0 | 0 | 0 | 86.5±0.4a |
| 14 | 0 | 0 | 0 | 85.3±0.6b |
| 15 | 0 | 0 | 0 | 83.1±1.0c |
X 1: temperature, −1 (20°C), 0 (30°C), +1 (40°C); X 2: pH, −1 (5), 0 (7), +1 (9); X 3: inoculum, −1 (OD600 = 0.1), 0 (OD600 = 0.3), +1 (OD600 = 0.5). The data presented are means of three replicates with standard deviation, which is within 5% of the mean. Different letters indicate significant differences (p<0.05, LSD test).
Biodegradation of bifenthrin in liquid medium
Growth experiments with bifenthrin as the sole carbon source were performed in 250-mL Erlenmeyer flasks containing 50 mL sterile MSM with 50 mg·L−1 bifenthrin. Co-metabolism experiments were carried out in the same medium supplemented with 1% (w/v) glucose [39]. The seed suspension was aseptically inoculated into the MSM and incubated for 7 days at pH 7.2 at 32°C with shaking at 150 rpm. Each treatment was set in triplicate with non-inoculated samples as control. Samples (30-mL) were withdrawn periodically from the cultures to examine growth by recording the optical density (OD) value at 600 nm using spectrophotometer and to measure the bifenthrin concentration by HPLC as described previously [34], [40].
Biodegradation experiments by the yeast with different concentrations of bifenthrin (100–600 mg·L−1) and with various pyrethroids including bifenthrin, cyfluthrin, deltamethrin, fenvalerate, cypermethrin, and fenpropathrin (50 mg·L−1) were carried out in MSM supplemented with 1% glucose for 5 days at pH 7.2 at 32°C.
Identification of bifenthrin metabolites
The metabolites of bifenthrin in cell-free filtrates of the yeast cultures grown in MSM containing 50 mg·L−1 of bifenthrin were detected by gas chromatopraphy-mass spectrometry (GC-MS) (Agilent, USA). The cell-free filtrates were collected at 2, 4, 6, and 8 days, respectively. After acidification to pH 2 with 2 M HCl, the cultures were extracted with ethyl acetate and supernatant was dehydrated, dried and re-dissolved in methanol [28]. After filtration with 0.45 µm membrance (Millipore, USA), the samples were analyzed by GC-MS according to Zhang et al. [40].
Biodegradation of bifenthrin in soil
Soil samples were taken from a depth of 5–20 cm over a 25 m2 area at grass-covered field, sieved to 2 mm and stored at 4°C in the dark for 14 days, before use [24]. The physicochemical properties of the soil were (grams per kilogram of dry weight): organic matter, 10.5; total N, 0.5; total P, 0.4; total K, 18.2; and pH, 6.9. The soil has a sandy loam texture (sand 65.0%, silt 28.0%, clay 7.0%). The soil intended for our bioremediation studies has not been used for agricultural purposes during the past 5 years, located in Guangzhou, Southern China. To investigate the removal potential of bifenthrin by the yeast in soils, 200 g of soil were placed in 500 mL-Erlenmeyer flask and their water contents were adjusted to 40% of water-holding capacity [39]. Soil moisture content was maintained at a constant level throughout the experiment by addition of distilled water when necessary. Bifenthrin was added to a final concentration of 50 mg·kg−1 of soil in acetone solution. After mixing and solvent evaporation the microbial suspensions were inoculated into soils (in triplicate) by drip irrigation to give the final concentration of 1.0×107 CFU·g−1 of soil and incubated at 32°C. The inoculum was thoroughly mixed under sterile conditions. In addition, sterilized soil (autoclaving at 121°C for 60 min) was also used to compare the removal efficiency of bifenthrin in soils [41], [42]. Triplicate samples of non-inoculated soils were kept as control. After 2, 4, 6, 8, and 10 days, soil samples (20 g) were removed for analysis to determine the residual concentration of bifenthrin. The extracting and analytical methods were analogous to those used in the liquid medium.
Chemical analysis
The pyrethroid residues were determined on an Agilent 1100 HPLC system equipped with a Hypersil ODS2 C18 reversed phase column (4.6 nm×250 mm, 5 µm) with array detection from 190–400 nm (total scan) based on retention time and peak area of the pure standard [29]. In brief, 30-mL of cell-free medium was extracted using 60 mL of acetone/petroleum ether (1∶1, v/v) in an ultrasonic bath. After partitioning, the supernatants were passed through a 0.22 mm polytetrafluoroethylene (PTFE) membrane filter (Millipore, USA), and the filtrates were concentrated using rotary evaporator (Heidolph, Germany). Residues were dissolved in acetone and analyzed by HPLC. A mixture of acetonitrile and water (85∶15, v/v) was used as the mobile phase at a flow rate of 1.0 mL·min−1. The injection volume was 10 µL.
The metabolites of bifenthrin were identified on an Agilent 6890N/5975 GC-MS system equipped with auto-sampler, an on-column, split/splitless capillary injection system, and with HP-5MS capillary column (30.0 m×250 µm×0.25 µm) with array detection from 30–500 nm (total scan). The operating conditions were as follows: the column was held at 160°C for 5 min, ramped at 10°C·min−1 to 200°C (first ramp), held at 200°C for 1 min, ramped at 10°C·min−1 to 280°C (second ramp), and then held at at 280°C for 8 min [40]. The temperatures corresponding to transfer line and the ion trap were 280°C and 230°C, respectively, and the ionization energy was 70 eV. The injection volume was 1.0 µL with splitless sampling at 250°C. Helium was used as a carrier gas at a flow rate of 1.5 mL·min−1.
Statistical analysis
Results were assessed by analysis of variance (ANOVA) and statistical analyses were performed on three replicates of data obtained from each treatment. The significance (p<0.05) of differences was treated statistically by one-, two-, or three-way ANOVA and evaluated by post hoc comparison of means using lowest significant differences (LSD) using SAS software packages.
Ethics Statement
No specific permits were required for the described field studies. No specific permissions were required for these locations. We confirm that the location is not privately-owned or protected in any way. We confirm that the field studies did not involve endangered or protected species.
Results
Isolation and identification of the bifenthrin-degrading strain ZS-02
Isolates were able to grow with bifenthrin as the sole carbon source identified one strain, designated ZS-02 and completely metabolized 50 mg·L−1 of bifenthrin within 8 days. This strain was an obligate aerobe (oxidase- and catalase-positive). Colonies were small, ivory, round, and with the entire margin when grown on YPD agar plates for 5 days. Cells were oval, and 3–4 µm in length and 2–3 µm in width (Fig. 1). It was positive in utilization of glucose, sucrose, trehalose, maltose, glycerol, xylose, raffinose, sorbierite, methyl-α-D-glucopyranoside, and cellobiose. It was negative in utilization of arabinose, adonitol, xylitol, galactose, inositol, 2-keto-D-gluconate, lactose, melezitose, and N-acetyl-D-glucosamine (Table 2).
Figure 1. Morphological characteristics of strain ZS-02 under scanning electron microscope (10,000×).

Table 2. Morphological and physio-biochemical characteristics of Candida pelliculosa strain ZS-02.
| Characteristics | Results | Characteristics | Results |
| Colonies | small, ivory, round, with entire margin | Cells | oval, 3–4 µm in length and 2–3 µm in width |
| Oxidase | + | Catalase | + |
| Glucose | + | Sucrose | + |
| Trehalose | + | Maltose | + |
| Glycerol | + | Xylose | + |
| Raffinose | + | Sorbierite | + |
| Methyl-α-D- glucopyranoside | + | N-acetyl-D- glucosamine | − |
| Cellobiose | + | Xylitol | − |
| Arabinose | − | Adonitol | − |
| Galactose | − | Inositol | − |
| Lactose | − | Melezitose | − |
| 2-Keto-D-gluconate | − |
+, tested positive/utilized as substrate; −, tested negative/not utilized as substrate.
PCR amplification of 18S rDNA gene from strain ZS-02, a single fragment of 1,452 bp, GenBank accession number JN700989, was obtained and completely sequenced. Phylogenetic analysis (Fig. 2) based on the 18S rDNA sequences revealed that strain ZS-02 belonged to Candida group and was closely related to Candida sp. BG02-6-9-4 (99%). The isolate was further classified as Candida pelliculosa by API 20C AUX system (bioMérieux, France), with very good identification (99.9%). Thus, strain ZS-02 was identified as C. pelliculosa based on morphological, physio-biochemical characteristics, API 20C AUX test, and 18S rDNA gene analysis. This is the first report of pyrethroid-degrading species from the genus Candida.
Figure 2. Phylogenetic tree based on the 18S rRNA sequence of strain ZS-02 and related strains.

Numbers in parentheses represent the sequences accession number in GenBank. Numbers at the nodes indicate bootstrap values from the neighborhood-joining analysis of 1,000 resampled data sets. Bar represents sequence divergence.
Optimization of the bifenthrin-degrading conditions by strain ZS-02
The Box-Behnken design was applied to analyze the main and interactive effects of important variables including temperature (X 1), pH (X 2), and inoculum (X 3) based on earlier single-factor experiments. The experimental design matrix and the response of dependent variable for bifenthrin degradation are presented in Table 1. Subsequently, the data from Table 1 were processed by response surface regression procedure of SAS software and results were fitted with the second-order polynomial model equation (Eq.(2)):
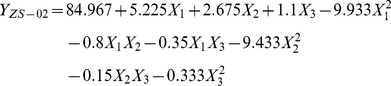 |
(2) |
where Y ZS-02 is the predicted bifenthrin degradation (%) by strain ZS-02; X1, X2, and X3 are the coded values for the temperature, pH, and inoculum, respectively.
An R 2 of 0.9770 indicated that approximately 98% of the variability in response could be covered by the model, demonstrating that predicted values of the model were in perfect agreement with the experimental values. The results of the regression parameter estimate revealed that linear and square terms of temperature (X 1) and pH (X 2) values showed significant effects (p<0.05) on the bifenthrin degradation by strain ZS-02, but the linear and square terms of inoculum (X 3) and the interaction terms were insignificant (p>0.05).
With the value of inoculum (the non-significant variable) fixed at OD600 nm = 0.3, the three-dimensional response surface was plotted to directly display the effects of temperature and pH on the bifenthrin degradation by strain ZS-02 by day 5 (Fig. 3). As shown in Fig. 3, the plot of bifenthrin biodegradation had a theoretical maximum value of 86.56% at the stationary point. At the stationary point, the optimum levels for the two variables of X1 and X2 were found to be 0.23 and 0.12 in terms of the coded units, that is, temperature 32.3°C and pH 7.2, respectively. So the optimal conditions for bifenthrin degradation by strain ZS-02 were determined to be 32.3°C, pH 7.2, and inoculum at OD600 nm = 0.3.
Figure 3. Response surface plot showing the effects of temperature and pH on bifenthrin degradation by strain ZS-02 with inoculum at OD600 = 0.3.

Effect of extra carbon source on the growth of strain ZS-02 and degradation of bifenthrin in mineral salt medium (MSM)
Growth of strain ZS-02 monitored by optical density (OD600 nm) and degradation kinetics of bifenthrin in MSM is shown in Fig. 4. After incubation for 7 days, 92.5% of the 50 mg·L−1 bifenthrin initially added to the medium was degraded by strain ZS-02, while in the same period disappearance rate of bifenthrin in MSM supplemented with 1% glucose was significantly higher (p<0.05) and reached 98.9%. Obviously, addition of carbon source enhanced the degradation of bifenthrin by this strain. No significant change in bifenthrin concentration was observed in non-inoculated cultures. Bifenthrin removal was associated with a concomitant increase of cell density. Similar to degradation, growth of strain ZS-02 was strongly stimulated in the presence of glucose, and was the most effective within 3 days of incubation whereas OD600 nm of the culture was significantly increased from 0.3 to 1.5. In contrast, the OD600 nm of the culture without additional of carbon source was significantly lower (p<0.05) and ranged from 0.3 to 1.0 after 3 days of incubation.
Figure 4. Cell growth of strain ZS-02 and degradation kinetics of bifenthrin during biodegradation studies.

○, degradation kinetics in MSM supplemented with bifenthrin as the sole carbon source; □, degradation kinetics in MSM supplemented with 1% glucose as an additional source of carbon; □, non-inoculated control (degradation); •, cell growth in MSM supplemented with bifenthrin as the sole carbon source; ▴, cell growth in MSM supplemented with 1% glucose as an additional source of carbon; ▪; non-inoculated control (growth). Values are the means of three replicates with standard deviation.
Biodegradation of bifenthrin with different initial concentrations
Strain ZS-02 grew on bifenthrin up to the concentration, as high as 600 mg·L−1. As shown in Fig. 5a, the lag phase was extended at higher bifenthrin concentration. Strain ZS-02 completely degraded bifenthrin at concentration of 100 mg·L−1 within 5 days. At concentrations of 200, 300, and 400 mg·L−1, the degradation rates reached 97.1%, 95.8%, and 91.3% after 5 days of incubation, respectively. However, only 87.6% and 81.4% degradation were achieved at higher initial concentrations of 500 and 600 mg·L−1, respectively.
Figure 5. Biodegradation of bifenthrin with different initial concentrations.

(a) Degradation kinetics of bifenthrin at different initial concentrations by strain ZS-02. •, 100 mg·L−1; □, 200 mg·L−1; ▴, 300 mg·L−1; □, 400 mg·L−1; ▪, 500 mg·L−1; □, 600 mg·L−1. Values are the means of three replicates with standard deviation. (b) Relationship between specific degradation rate and initial bifenthrin concentration by strain ZS-02.
The decrease in the specific bifenthrin degradation rate with an increase in the initial bifenthrin concentration implied that bifenthrin acts as an inhibitor to strain ZS-02. Therefore, the substrate inhibition model (Eq.(3)) adapted from Wang et al. [43] was used to explain the degradation kinetics of bifenthrin by strain ZS-02.
| (3) |
where q max is the maximum specific bifenthrin degradation rate (day−1), K i is the substrate inhibition constant (mg·L−1), K s is the half-saturation constant (mg·L−1), S is the substrate concentration (mg·L−1), and S m is a critical inhibitor concentration of the substrate which decreases degradation.
The kinetic parameters of strain ZS-02 estimated from non-linear regression using matrix laboratory (MATLAB) software (Version 7.8) were qmax = 1.7015 day−1 and K s = 86.2259 mg·L−1. The inhibitory effect of bifenthrin was considered to occur in a linear manner at K i = 187.2340 mg·L−1. The S m was determined to be 127.06 mg·L−1. The relationship between q and S is shown in Fig. 5b. The value of R 2 was 0.9516 demonstrating that the experimental data were well correlated with the model. As indicated in Fig. 5b, when S were lower than 127.06 mg·L−1, q gradually increased. At higher concentrations, inhibition by bifenthrin became prominent.
Biodegradation of various pyrethroids by strain ZS-02
The abilities of strain ZS-02 to degrade different pyrethroids are shown in Fig. 6. The strain was capable of metabolizing all of the pyrethroids tested. Bifenthrin was the most preferred substrate, with a degradation rate of 95.1% within 5 days of incubation. Cyfluthrin, deltamethrin, fenvalerate, cypermethrin, and fenpropathrin were degraded slower than bifenthrin, with degradation rates of 94.8%, 93.4%, 92.6%, 87.7%, and 51.3%, respectively.
Figure 6. Degradation of various pyrethroids by strain ZS-02 by day 5.

Values are the means of three replicates with standard deviation. Different letters indicate significant differences (P<0.05, LSD test).
Identification of bifenthrin metabolites
To illuminate the pathway of bifenthrin degradation by strain ZS-02, bifenthrin metabolic products in cell-free filtrates were extracted and confirmed by GC-MS. The degradation products identified on the basis of mass spectrum were matched with authentic standard compounds from the National Institute of Standards and Technology (NIST, USA) library database.
The GC-MS analysis revealed the presence of five degradation products, as summarized in Table 3. The peak at retention time of 16.391 min corresponded to bifenthrin standard. This peak decreased over time and completely disappeared after 8 days. Bifenthrin [A] disappeared concomitantly with formation of five degradation products (Fig. S1). The five compounds corresponded with those of authentic cyclopropanecarboxylic acid [B], 2-methyl-3-biphenylyl methanol [C], 4-trifluoromethoxy phenol [D], 2-chloro-6-fluoro benzylalcohol [E], and 3,5-dimethoxy phenol [F]. The retention times of these compounds were 9.561, 9.508, 8.785, 11.327, and 12.674 min, respectively. However, the peaks corresponding to these compounds were transient and they disappeared finally. Eventually, no persistent accumulative product was detected.
Table 3. Chromatographic properties of metabolites of bifenthrin degraded by strain ZS-02.
| Code | Retention time (min) | m/z | Compounds |
| A | 16.391 | 458 | Bifenthrin |
| B | 9.561 | 195 | Cyclopropanecarboxylic acid |
| C | 9.508 | 282 | 2-Methyl-3-biphenylyl methanol |
| D | 8.785 | 430 | 4-Trifluoromethoxy phenol |
| E | 11.327 | 282 | 2-Chloro-6-fluoro benzylalcohol |
| F | 12.674 | 281 | 3,5-Dimethoxy phenol |
On the basis of the metabolic products formed, we proposed the degradation pathway for bifenthrin by strain ZS-02 (Fig. 7). In other words, the parent bifenthrin [A] was first metabolized by hydrolysis of the carboxylester linkage to produce cyclopropanecarboxylic acid [B] and 2-methyl-3-biphenylyl methanol [C]. Subsequently, 2-methyl-3-biphenylyl methanol [C] was further transformed by biphenyl cleavage, resulting in formation of 4-trifluoromethoxy phenol [D], 2-chloro-6-fluoro benzylalcohol [E], and 3,5-dimethoxy phenol [F]. Finally, all these metabolites were not detected in the culture medium after 8 days of incubation. These results indicated that the added bifenthrin (50 mg·L−1) was completely degraded by strain ZS-02 without any accumulative metabolites at the end of reaction.
Figure 7. Proposed pathway for the degradation of bifenthrin by strain ZS-02.

Biodegradation of bifenthrin in soil
The potential of strain ZS-02 to eliminate bifenthrin in contaminated soils was investigated under controlled environmental conditions (temperature and soil humidity). The degradation kinetics are shown in Fig. 8. In the non-sterilized soil introduced with strain ZS-02 (1.0×107 CFU·g−1 of soil), bifenthrin degradation started rapidly as utilization was observed at the beginning of incubation, apparently there was no lag phase and 75.1% of the 50 mg·kg−1 bifenthrin initially added to the soil was removed after 10 days of treatment; whereas in control with indigenous soil microorganisms, this activity decreased only by 8.4%. In case of sterilized soil inoculated with strain ZS-02, 64.7% of added bifenthrin was eliminated within 10 days; while a non-inoculated control showed that the total bifenthrin content decreased only by 3.4%. Obviously, strain ZS-02 could efficiently eliminate bifenthrin in contaminated soils as compared to the non-inoculated control. In addition, the disappearance rate of bifethrin in the non-sterilized soil was higher (p<0.05) than that in the sterilized soil.
Figure 8. Degradation kinetics of bifenthrin in different contaminated soils by strain ZS-02.

□, non-inoculated control (sterilized soil); ○, non-inoculated control with indigenous microbial community (non-sterilized soil); ▴, sterilized soil introduced with strain ZS-02; •, non-sterilized soil introduced with strain ZS-02. Values are the means of three replicates with standard deviation.
To confirm the effects on degradation of bifenthrin in different soils by strain ZS-02, the first-order kinetic model (Eq.(4)) adapted from Cycoń et al. [39] was used to describe the rate of total bifenthrin reduction.
| (4) |
where C 0 is the initial concentration of bifenthrin at time zero, C t is the concentration of bifenthrin at time t, k and t are the degradation rate constant (day−1) and degradation period in days, respectively.
The half-life (t 1/2) for bifenthrin in various soils was calculated using the general formula as expressed in Eq.(5).
| (5) |
where k is the rate constant (day−1).
The kinetic parameters for all runs calculated from the equations are tabulated in Table 4. Degradation kinetics showed that the process corresponds to first-order kinetics with R 2 ranging from 0.9492 to 0.9860, demonstrating that the experimental data were well correlated with the model. The kinetic constant (k) for the non-sterilized soil with strain ZS-02 was 0.1411 day−1, while the k for the sterilized soil was 0.1096 day−1. In the sterilized soil, longer half-lives (t 1/2) of 6.3 days were observed, as compared to the non-sterilized soil with a t 1/2 of 4.9 days. These results revealed that the bifenthrin removal efficiency by strain ZS-02 could be enhanced by the indigenous soil microorganisms. Furthermore, the t 1/2 for bifenthrin in various soils introduced with strain ZS-02 (t 1/2 = 4.9–6.3 days) were significantly shortened (p<0.05) as compared to that in the non-inoculated control soils (t 1/2 = 78.8–130.7 days).
Table 4. Kinetic parameters of degradation of bifenthrin in soils by strain ZS-02.
| Soil treatments | Regression equation | k (day−1) | t 1/2 (days) | R 2 |
| nSS+bifenthrin | Ct = 50.1846e−0.0088t | 0.0088 | 78.8 | 0.9799 |
| SS+bifenthrin | Ct = 50.0841e−0.0053t | 0.0053 | 130.7 | 0.9806 |
| nSS+bifenthrin+ZS-02 | Ct = 51.1033e−0.1411t | 0.1411 | 4.9 | 0.9860 |
| SS+bifenthrin+ZS-02 | Ct = 52.8446e−0.1096t | 0.1096 | 6.3 | 0.9492 |
Each figure in the table represents the mean of three replicates. All figures were significantly different at p<0.05. nSS: non-sterilized soil with indigenous soil microbes; SS: sterilized soil without inoculum; +: introduced with; Ct: residual concentration of bifenthrin (mg·kg−1); t: degradation period (days). R 2: correlation coefficient.
Discussion
Bifenthrin, a broad-spectrum type I synthetic pyrethroid insecticide, has been heavily used throughout the world for pest control both in agricultural and urban environment during the past two decades, resulting in widespread contamination [2], [3], [5], [8], [16]. Bioremediation involves the use of living microorganisms to degrade and detoxify pollutants, and is generally considered to be the most significant process determining the fate and behavior of xenobiotics in the environment [44]. Although several microorganisms capable of degrading pyrethroids have been isolated from diverse geographic locations [26]–[30], yet no report on the isolation of bifenthrin-degrading strain is available. A possible reason was attributed to that this chemical is persistent and refractory to microbial degradation [45]. Additionally, the existing papers lack the information involved in the biodegradation of pyrethroids by yeast. Yeast possesses the biochemical and ecological capacity to degrade environmental organic chemicals, either by chemical modification or by influencing chemical bioavailability [46]. However, the potential use for yeast in bioremediation of pyrethroids has not received the attention it deserves. This is the first report to our knowledge about bioremediation of pyrethroids by yeast.
In the present study, the screening of bifenthrin-degrading yeast by the method of enrichment procedure from active sludge affected by pyrethroids allowed us to select some potential isolates with a high survivability in the environment and maximal degrading activity towards bifenthrin. It was generally considered that the conditions for environmental microorganisms enrichment and screening are crucial in the selection of isolates not only with the desired degrading enzyme systems but having specific regulation of the degradation pathways as well [47]. One most active yeast, designated ZS-02, able to completely metabolize bifenthrin was identified as Candida pelliculosa based on morphological, physio-biochemical characteristics, API 20C AUX test, and 18S rDNA gene analysis. Previous research has shown that the potential pyrethroid-degrading microorganisms were mostly from genera Bacillus and Pseudomonas [26], [27], [38], while C. pelliculosa strain ZS-02 appeared to be a new species that was found highly effective in degrading bifenthrin and several other pyrethroids. Members of this genus are ubiquitous in the environment and some isolates have antagonistic effect on pathogen due to their production of bioactive secondary metabolites [48]. However, there is limited information concerning the use of these isolates to degrade and detoxify pollutants. The current results expand their use.
Previous studies have shown that temperature and pH are two important factors which strongly influence the degradation ability of microorganisms able to degrade xenobiotic compounds [39], [49], [50]. Our results indicated that strain ZS-02 engaged in degrading bifenthrin over a wide range of temperatures (20–40°C) and pH (5–9) particularly at low pH. This is an important feature of a microorganism to be employed for bioremediation of variable environments [49]. Furthermore, the optimal conditions for bifenthrin degradation by strain ZS-02 were determined to be 32.3°C and pH 7.2 via response surface methodology (RSM). RSM is an efficient statistical technique that has been successfully applied to optimize degradation conditions for various microorganisms [30], [32], [33], [38], [51]. It is a faster and less expensive method for gathering research result than the conventional practice of one factor optimization [51]. Moreover, RSM can distinguish interaction effects from the effects of individual factors through appropriate construction of the process model [30], [38]. In our studies, a mathematical model was developed, and this model could be effectively used to predict and optimize the bifenthrin degradation conditions by strain ZS-02 within the limits of chosen factors.
C. pelliculosa strain ZS-02 utilized bifenthrin as the sole carbon for growth as well as co-metabolized it in the presence of extra carbon source, thus suggesting adaptation to oligotrophic and eutrophic conditions. However, this observation did not quite agree with Sørensen et al. [52] who reported isoproturon-degrading strain Sphingomonas sp. SRS2 was unable to grow on rich media, thus only adaptation to oligotrophic conditions. Furthermore, the growth of strain ZS-02 was stimulated and degradation of bifenthrin significantly enhanced in the presence of glucose in our studies (Fig. 4). This might be because of the cometabolism with additional carbon source lead to enhanced degradation of bifenthrin by strain ZS-02. Similar accelerated degradation in the presence of extra carbon source was observed by Anwar et al. [50] and Cycoń et al. [39] for different microorganisms degrading organophosphates.
It was noteworthy that this particular strain could tolerate and efficiently degrade bifenthrin up to the concentration, as high as 600 mg·L−1. However, the specific bifenthrin degradation rate decreased with an increase in the initial bifenthrin concentration (Fig. 5b). These findings indicated that increased bifenthrin concentration had a marked effect on degradation performance of strain ZS-02, but did not lead to complete inhibition. These results proved that strain ZS-02 was responsible for bifenthrin degradation. Another important feature which is worth mentioning is that this strain was capable of degrading a variety of of pyrethroids including bifenthrin, cyfluthrin, deltamethrin, fenvalerate, cypermethrin, and fenpropathrin, demonstrating that the pyrethroid hydrolase involved in degradation may have broad-spectrum substrate specificity. Bifenthrin was degraded faster than other pyrethroids tested, suggesting that presence of the cyano group caused relative reduction in the hydrolysis rate, due to either stabilization of the ester bond or toxic effect of the substituent group. Another possible reason for degradation reduction could be attributed to the fact that substitution of the biphenyl for a 3-phenoxybenzyl hinders the pyrethroid hydrolase-substrate interaction in strain ZS-02. However, the result contrasts with previous findings of Wang et al. [31] who reported that replacement of the 3-phenoxybenzyl with a biphenyl decreased the pyrethroid hydrolysis rate in Ochrobactrum tritici strain pyd-1. Additionally, cyfluthrin, deltamethrin, fenvalerate, and cypermethrin were degraded at much faster rates than fenpropathrin, indicating that with fluoro, bromo, chloroben, or chloro group on the chrysanthemic acid greatly improved the degradation efficiencies. It might be because of that these substituent groups strongly promoted the hydrolase activity. Therefore, we could conclude that the degradation efficiencies of strain ZS-02 depend on the molecular structure of the pyrethroids. Similar result has been proposed by Wang et al. [31], [45] and Guo et al. [29] for various microorganisms degrading pyrethroids.
It was generally suggested that ester hydrolysis by carboxylesterases was the main degradation pathway of pyrethroids in a multitude of species, from mammals, insects to microorganisms [28], [30], [38], [45], [53]. In our studies, the degradation pathway of bifenthrin by strain ZS-02 was investigated by metabolite identification, and the proposed pathway is shown in Fig. 7. It was evident from the results that strain ZS-02 first degraded bifenthrin by hydrolysis of ester linkage to produce cyclopropanecarboxylic acid and 2-methyl-3-biphenylyl methanol, leading to loss of its insecticide activity. Subsequently, 2-methyl-3-biphenylyl methanol was further transformed by biphenyl cleavage to form 4-trifluoromethoxy phenol, 2-chloro-6-fluoro benzylalcohol, and 3,5-dimethoxy phenol, resulting in its detoxification. Finally, all these degradation products were not detected by GC-MS after 8 days of incubation. These results indicated that the 50 mg·L−1 bifenthrin initially added to the medium was completely degraded by strain ZS-02 without any accumulative metabolites at the end of reaction. To the best of our knowledge, this is the first report of a novel pathway of degradation of bifenthrin by hydrolysis of ester linkage and cleavage of biphenyl in a microorganism. The bifenthrin metabolic pathway by ZS-02, however, appears to be similar to the initial step of cypermethrin degradation by Micrococcus sp. strain CPN 1, in which cypermthrin was converted to an acid and an alcohol by hydrolysis [28].
The potential of strain ZS-02 to eliminate bifenthrin in contaminated soils was evaluated to find out whether such strain is suitable for bioremediation purposes. In these experiments soils were inoculated with 1.0×107 CFU·g−1 of soil and this inoculum density was found to efficiently remove bifenthrin from the environment. As shown in Fig. 8, the introduced strain ZS-02 quickly adapted to the environment and rapidly degraded bifenthrin at the beginning of incubation without any apparent lag phase. After 10 days of incubation, 75.1% and 64.7% of the added bifenthrin (50 mg·kg−1) was eliminated in the non-sterilized and sterilized soils, respectively. The t 1/2 for bifenthrin was remarkably reduced by 63.9 and 124.4 days as compared to the control, implying that strain ZS-02 may have great potential to eliminate bifenthrin in contaminated soils. Furthermore, the removal efficiency of bifenthrin in the non-sterilized soil was significantly higher (p<0.05) than that in the sterilized soil, indicating that the indigenous soil microorganisms strongly promoted the ability of strain ZS-02 to eliminate bifenthrin. Enhancement of degradation could be explained by this reason that the introduced strain ZS-02 and soil microorganisms may have a synergistic effect on removal of pollutants. These results are consistent with previous observation [41], [42].
In conclusion, the C. pelliculosa strain ZS-02 isolated in the present study appeared to be highly efficient in degrading bifenthrin in different contaminated soil and water resources. Degradation of bifenthrin occurred at 20–40°C and pH 5–9. This is an important feature of a microorganism to be employed for bioremediation of variable environments. The yeast utilized bifenthrin as the sole carbon for growth as well as co-metabolized it in the presence of extra carbon source, thus suggesting adaptation to oligotrophic and eutrophic conditions. Another important feature which is worth mentioning is that this strain was proficient in degrading a variety of pyrethroids, indicating that the yeast could be ideal for remediation of soil and water contaminated with pyrethroids. Moreover, the yeast harbors the metabolic pathway for the detoxification of bifenthrin. This is the first report of a novel pathway of degradation of bifenthrin by hydrolysis of ester linkage and cleavage of biphenyl in a microorganism, which we propose plays an important role in the bifenthrin biogeocycle. Finally, strain ZS-02 is the first described yeast capable of degrading bifenthrin and several other pyrethroids.
Supporting Information
GC-MS spectra of metabolites produced from bifenthrin by Candida pelliculosa strain strain ZS-02. (a) Bifenthrin; (b) Cyclopropanecarboxylic acid; (c) 2-Methyl-3-biphenylyl methanol; (d) 4-Trifluoromethoxy phenol; (e) 2-Chloro-6-fluoro benzylalcohol; (f) 3,5-Dimethoxy phenol.
(TIF)
Acknowledgments
The authors would like to thank Dr. Muhammad Rizwan-ul-haq for his helpful comments on the manuscript.
Footnotes
Competing Interests: The authors have declared that no competing interests exist.
Funding: The authors gratefully acknowledge the grants from the National Natural Science Foundation (No. 30871660) and Project of Scientific Technological Planning of Guangdong Province (No. 2009B020310005), People's Republic of China. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Laffin B, Chavez M, Pine M. The pyrethroid metabolites 3-phenoxybenzoic acid and 3-phenoxybenzyl alcohol do not exhibit estrogenic activity in the MCF-7 human breast carcinoma cell line or Sprague-Dawley rats. Toxicology. 2010;267:39–44. doi: 10.1016/j.tox.2009.10.003. [DOI] [PubMed] [Google Scholar]
- 2.Hintzen EP, Lydy MJ, Belden JB. Occurrence and potential toxicity of pyrethroids and other insecticides in bed sediments of urban streams in central Texas. Environ Pollut. 2009;157:110–116. doi: 10.1016/j.envpol.2008.07.023. [DOI] [PubMed] [Google Scholar]
- 3.Weston DP, Holmes RW, Lydy MJ. Residential runoff as a source of pyrethroid pesticides to urban creeks. Environ Pollut. 2009;157:287–294. doi: 10.1016/j.envpol.2008.06.037. [DOI] [PubMed] [Google Scholar]
- 4.Wang LM, Liu WP, Yang CX, Pan ZY, Gan JY, et al. Enantioselectivity in estrogenic potential and uptake of bifenthrin. Environ Sci Technol. 2007;41:6124–6128. doi: 10.1021/es070220d. [DOI] [PubMed] [Google Scholar]
- 5.Liu J, Yang Y, Yang Y, Zhang Y, Liu WP. Disrupting effects of bifenthrin on ovulatory gene expression and prostaglandin synthesis in rat ovarian granulosa cells. Toxicology. 2011;282:47–55. doi: 10.1016/j.tox.2011.01.007. [DOI] [PubMed] [Google Scholar]
- 6.Holmes RW, Anderson BS, Phillips BM, Hunt JW, Crane DB, et al. Statewide investigation of the role of pyrethroid pesticides in sediment toxicity in California's urban waterways. Environ Sci Technol. 2008;42:7003–7009. doi: 10.1021/es801346g. [DOI] [PubMed] [Google Scholar]
- 7.Riederer AM, Hunter RE, Hayden SW, Ryan PB. Pyrethroid and organophosphorus pesticides in composite diet samples from Atlanta, USA adults. Environ Sci Technol. 2010;44:483–490. doi: 10.1021/es902479h. [DOI] [PubMed] [Google Scholar]
- 8.Weston DP, Asbell AM, Hecht SA, Scholz NL, Lydy MJ. Pyrethroid insecticides in urban salmon streams of the Pacific Northwest. Environ Pollut. 2011;159:3051–3056. doi: 10.1016/j.envpol.2011.04.008. [DOI] [PubMed] [Google Scholar]
- 9.Tewary DK, Kumar V, Ravindranath SD, Shanker A. Dissipation behavior of bifenthrin residues in tea and its brew. Food Control. 2005;16:231–237. [Google Scholar]
- 10.Mohapatra S, Ahuja AK, Sharma D. Persistence of bifenthrin residues on mango (Mangifera indica) fruit. Pestic Res J. 2007;19:110–112. [Google Scholar]
- 11.Shafer TJ, Meyer DA, Crofton KM. Developmental neurotoxicity of pyrethroid insecticides: critical review and future research needs. Environ Health Persp. 2005;113:123–136. doi: 10.1289/ehp.7254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Wolansky MJ, Harrill JA. Neurobehavioral toxicology of pyrethroid insecticides in adult animals: a critical review. Neurotoxicol Teratol. 2008;30:55–78. doi: 10.1016/j.ntt.2007.10.005. [DOI] [PubMed] [Google Scholar]
- 13.Dutta R, Das N. Immunomodulation of serum complement (C3) and macrophages by synthetic pyrethroid fenvalerate: In vitro study. Toxicology. 2011;285:126–132. doi: 10.1016/j.tox.2011.04.012. [DOI] [PubMed] [Google Scholar]
- 14.Jin YX, Zheng SS, Fu ZW. Embryonic exposure to cypermethrin induces apoptosis and immunotoxicity in zebrafish (Danio rerio). Fish Shellfish Immunol. 2011;30:1049–1054. doi: 10.1016/j.fsi.2011.02.001. [DOI] [PubMed] [Google Scholar]
- 15.Perry MJ, Venners SA, Barr DB, Xu XP. Environmental pyrethroid and organophosphorus insecticide exposures and sperm concentration. Reprod Toxicol. 2007;23:113–118. doi: 10.1016/j.reprotox.2006.08.005. [DOI] [PubMed] [Google Scholar]
- 16.Wang C, Chen F, Zhang Q, Fang Z. Chronic toxicity and cytotoxicity of synthetic pyrethroid insecticide cis-bifenthrin. J Environ Sci. 2009;21:1710–1715. doi: 10.1016/s1001-0742(08)62477-8. [DOI] [PubMed] [Google Scholar]
- 17.Ansari AR, Rahman S, Kaur M, Anjum S, Raisuddin S. In vivo cytogenetic and oxidative stress-inducing effects of cypermethrin in freshwater fish, Channa punctata Bloch. Ecotoxicol Environ Saf. 2011;74:150–156. doi: 10.1016/j.ecoenv.2010.08.036. [DOI] [PubMed] [Google Scholar]
- 18.US EPA. Endocrine Disruptor Screening Program (EDSP). 2009. US EPA,Washington, DC.
- 19.Baskaran S, Kookana RS, Naidu R. Degradation of bifenthrin, chlorpyrifos and imidacloprid in soil and bedding materials at termiticidal application rates. Pestic Sci. 1999;55:1222–1228. [Google Scholar]
- 20.Laskowski DA. Physical and chemical properties of pyrethroids. Rev Environ Contam T. 2002;174:49–170. doi: 10.1007/978-1-4757-4260-2_3. [DOI] [PubMed] [Google Scholar]
- 21.Lee S, Gan JY, Kim JS, Kabashima JN, Crowley DE. Microbial transformation of pyrethroid insecticides in aqueous and sediment phases. Environ Toxicol Chem. 2004;23:1–6. doi: 10.1897/03-114. [DOI] [PubMed] [Google Scholar]
- 22.Mohapatra S, Ahuja AK. Effect of moisture and soil type on the degradation of bifenthrin in soil. Pestic Res J. 2009;19:110–112. [Google Scholar]
- 23.World Health Organization (WHO) 2009. The WHO Recommended Classification of Pesticides by Hazard and Guidelinesto Classification 2009, WHO, Geneva.
- 24.Fenlon AK, Jones CK, Semple TK. The effect of soil:water ratios on the induction of isoproturon, cypermethrin and diazinon mineralisation. Chemosphere. 2011;82:163–168. doi: 10.1016/j.chemosphere.2010.10.027. [DOI] [PubMed] [Google Scholar]
- 25.Singh BK, Walker A. Microbial degradation of organophosphorus compounds. FEMS Microbiol Rev. 2006;30:428–471. doi: 10.1111/j.1574-6976.2006.00018.x. [DOI] [PubMed] [Google Scholar]
- 26.Maloeny SE, Maule A, Smith ARW. Purification and preliminary characterization of permethrinase from a pyrethroid-transforming strain of Bacillus cereus. Appl Environ Microbiol. 1993;59:2007–2013. doi: 10.1128/aem.59.7.2007-2013.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Grant RJ, Daniell TJ, Betts WB. Isolation and identification of synthetic pyrethroid-degrading bacteria. J Appl Microbiol. 2002;92:534–540. doi: 10.1046/j.1365-2672.2002.01558.x. [DOI] [PubMed] [Google Scholar]
- 28.Tallur PN, Megadi VB, Ninnekar HZ. Biodegradation of cypermethrin by Micrococcus sp. strain CPN 1. Biodegradation. 2008;28:77–82. doi: 10.1007/s10532-007-9116-8. [DOI] [PubMed] [Google Scholar]
- 29.Guo P, Wang BZ, Hang BJ, Li L, Ali SW, et al. Pyrethroid- degrading Sphingobium sp. JZ-2 and the purification and characterization of a novel pyrethroid hydrolase. Int Biodeter Biodegr. 2009;63:1107–1112. [Google Scholar]
- 30.Zhang C, Jia L, Wang SH, Qu J, Xu LL, et al. Biodegradation of beta-cypermethrin by two Serratia spp. with different cell surface hydrophobicity. Bioresoure Technol. 2010;101:3423–3429. doi: 10.1016/j.biortech.2009.12.083. [DOI] [PubMed] [Google Scholar]
- 31.Wang BZ, Ma Y, Zhou WY, Zheng JW, He J, et al. Biodegradation of synthetic pyrethroids by Ochrobactrum tritici strain pyd-1. World J Microbiol Biotechnol. 2011;27:2315–2324. [Google Scholar]
- 32.Chen SH, Hu QB, Hu MY, Luo JJ, Weng QF, et al. Isolation and characterization of a fungus able to degrade pyrethroids and 3-phenoxybenzaldehyde. Bioresource Technol. 2011;102:8110–8116. doi: 10.1016/j.biortech.2011.06.055. [DOI] [PubMed] [Google Scholar]
- 33.Chen SH, Lai KP, Li YN, Hu MY, Zhang YB, et al. Biodegradation of deltamethrin and its hydrolysis product 3-phenoxybenzaldehyde by a newly isolated Streptomyces aureus strain HP-S-01. Appl Microbiol Biotechnol. 2011;90:1471–1483. doi: 10.1007/s00253-011-3136-3. [DOI] [PubMed] [Google Scholar]
- 34.Chen SH, Hu MY, Liu JJ, Zhong GH, Yang L, et al. Biodegradation of beta-cypermethrin and 3-phenoxybenzoic acid by a novel Ochrobactrum lupini DG-S-01. J Hazard Mater. 2011;187:433–440. doi: 10.1016/j.jhazmat.2011.01.049. [DOI] [PubMed] [Google Scholar]
- 35.Barnett JA, Payne RW, Yarrow D. Yeasts: Characteristics and Identification, 3rd ed. 2000. Cambridge University Press, Cambridge, UK.
- 36.Sambrook J, Russell DW. Molecular Cloning: A Laboratory Manual, 3rd ed. 2001. Cold Spring Harbor Laboratory, NY.
- 37.Tamura K, Dudley J, Nei M. MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol Biol Evol. 2007;24:1596–1599. doi: 10.1093/molbev/msm092. [DOI] [PubMed] [Google Scholar]
- 38.Zhang C, Wang SH, Yan YC. Isomerization and biodegradation of beta-cypermethrin by Pseudomonas aeruginosa CH7 with biosurfactant production. Bioresource Technol. 2011;102:7139–7146. doi: 10.1016/j.biortech.2011.03.086. [DOI] [PubMed] [Google Scholar]
- 39.Cycoń M, Wojcik M, Piotrowska-Seget Z. Biodegradation of the organophosphorus insecticide diazinon by Serratia sp. and Pseudomonas sp. and their use in bioremediation of contaminated soil. Chemosphere. 2009;76:494–501. doi: 10.1016/j.chemosphere.2009.03.023. [DOI] [PubMed] [Google Scholar]
- 40.Zhang SB, Yin LB, Liu Y, Zhang DY, Luo XW, et al. Cometabolic biotransformation of fenpropathrin by Clostridium species strain ZP3. Biodegradation. 2011;22:869–875. doi: 10.1007/s10532-010-9444-y. [DOI] [PubMed] [Google Scholar]
- 41.Chen SH, Yang L, Hu MY, Liu JJ. Biodegradation of fenvalerate and 3- phenoxybenzoic acid by a novel Stenotrophomonas sp. strain ZS-S-01 and its use in bioremediation of contaminated soils. Appl Microbiol Biotechnol. 2011;90:755–767. doi: 10.1007/s00253-010-3035-z. [DOI] [PubMed] [Google Scholar]
- 42.Chen SH, Geng P, Xiao Y, Hu MY. Bioremediation of β-cypermethrin and 3-phenoxybenzaldehyde contaminated soils using Streptomyces aureus HP-S-01. Appl Microbiol Biotechnol. 2011 doi: 10.1007/s00253-011-3640-5. in press. [DOI] [PubMed] [Google Scholar]
- 43.Wang YQ, Zhang JS, Zhou JT, Zhang ZP. Biodegradation of 4-aminobenzenesulfonate by a novel Pannonibacter sp. W1 isolated from activated sludge. J Hazard Mater. 2009;169:1163–1167. doi: 10.1016/j.jhazmat.2009.04.002. [DOI] [PubMed] [Google Scholar]
- 44.Singh BK. Organophosphorus-degrading bacteria: ecology and industrial applications. Nature Rev Microbiol. 2009;7:156–163. doi: 10.1038/nrmicro2050. [DOI] [PubMed] [Google Scholar]
- 45.Wang BZ, Guo P, Hang BJ, Li L, He J, et al. Cloning of a novel pyrethroid-hadrolyzing carboxylesterase gene from Sphingobium sp. strain JZ-1 and characteization of the gene product. Appl Environ Microbiol. 2009;75:5496–5500. doi: 10.1128/AEM.01298-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Harms H, Schlosser D, Wick LY. Untapped potential: exploiting fungi in bioremediation of hazardous chemicals. Nature Rev Microbiol. 2011;9:177–192. doi: 10.1038/nrmicro2519. [DOI] [PubMed] [Google Scholar]
- 47.Xu GM, Zheng W, Li YY, Wang SH, Zhang JS, et al. Biodegradation of chlorpyrifos and 3,5,6-trichloro-2-pyridinol by a newly isolated Paracoccus sp. strain TRP. Int Biodeter Biodegr. 2008;62:51–56. [Google Scholar]
- 48.Bello GD, Monaco C, Rollan MC, Lampugnani G, Arteta N, et al. Biocontrol of postharvest grey mould on tomato by yeasts. J Phytopathology. 2008;156:257–263. [Google Scholar]
- 49.Singh BK, Walker A, Denis J, Wight DJ. Bioremedial potential of fenamiphos and chlorpyrifos degrading isolates: influence of different environmental conditions. Soil Biol Biochem. 2006;38:2682–2693. [Google Scholar]
- 50.Anwar S, Liaquat F, Khan QM, Khalid ZM, Iqbal S. Biodegradation of chlorpyrifos and its hydrolysis product 3,5,6-trichloro-2-pyridinolby Bacillus pumilus strain C2A1. J Hazard Mater. 2009;168:400–405. doi: 10.1016/j.jhazmat.2009.02.059. [DOI] [PubMed] [Google Scholar]
- 51.Dritsa V, Rigas F, Doulia D, Avramides EJ, Hatzianestis I. Optimization of culture conditions for the biodegradation of lindane by the polypore fungus Ganoderma australe. Water Air Soil Pollut. 2009;204:19–27. [Google Scholar]
- 52.Sørensen SR, Ronen Z, Aamand J. Isolation from agricultural soil and characterization of a Sphingomonas sp. able to mineralize the phenylurea herbicide isoproturon. Appl Environ Microbiol. 2001;67:5403–5409. doi: 10.1128/AEM.67.12.5403-5409.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Heidari R, Devonshire AL, Campbell BE, Dorrian SJ, Oakeshott JG, et al. Hydrolysis of pyrethroids by carboxylesterases from Lucilia cuprina and Drosophila melanogaster with active sites modified by in vitro mutagenesis. Insect Biochem Mol Biol. 2005;35:597–609. doi: 10.1016/j.ibmb.2005.02.018. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
GC-MS spectra of metabolites produced from bifenthrin by Candida pelliculosa strain strain ZS-02. (a) Bifenthrin; (b) Cyclopropanecarboxylic acid; (c) 2-Methyl-3-biphenylyl methanol; (d) 4-Trifluoromethoxy phenol; (e) 2-Chloro-6-fluoro benzylalcohol; (f) 3,5-Dimethoxy phenol.
(TIF)