

Abstract
Wolbachia are a group of intracellular inherited endosymbiontic bacteria infecting a wide range of insects. In this study the infection status of Wolbachia (Rickettsiales: Rickettsiaceae) was measured in the Asiatic rice leafroller, Cnaphalocrocis medinalis (Guenée) (Lepidoptera: Pyralidae), from twenty locations in China by sequencing wsp, ftsZ and 16S rDNA genes. The results showed high infection rates of Wolbachia in C. medinalis populations. Wolbachia was detected in all geographically separate populations; the average infection rate was ∼ 62.5%, and the highest rates were 90% in Wenzhou and Yangzhou populations. The Wolbachia detected in different C. medinalis populations were 100% identical to each other when wsp, ftsZ, and 16S rDNA sequences were compared, with all sequences belonging to the Wolbachia B supergroup. Based on wsp, ftsZ and 16S rDNA sequences of Wolbachia, three phylogenetic trees of similar pattern emerged. This analysis indicated the possibility of inter-species and intra-species horizontal transmission of Wolbachia in different arthropods in related geographical regions. The migration route of C. medinalis in mainland China was also discussed since large differentiation had been found between the wsp sequences of Chinese and Thai populations.
Keywords: infection rate, phylogenetic relationship
Introduction
The genus Wolbachia (Rickettsiales: Rickettsiaceae) is a group of intracellular gram-negative and vertically inherited endosymbiontic bacteria that belong to the order Rickettsiales in the α-subdivision of the class Proteobacteria (Werren 1997). Numerous surveys indicate that Wolbachia can infect a wide range of arthropods and filarial nematodes. Recent surveys indicate that 20% to 76% of examined insects harbor Wolbachia (Hilgenboecker et al. 2008), as well as many arachnids, terrestrial crustaceans, and mites, making this group one of the most widespread obligate bacterial endosymbionts ever described (Cordaux et al. 2001; Gotoh et al. 2003; Rowley et al. 2004).
Wolbachia play important roles in ecology, evolution, and reproductive regulation in their hosts (Werren 1997), and have been considered as a potent evolutionary force. In nematodes, Wolbachia appear to play a mutualistic role in development and reproduction (Bandi et al. 1999; Langworthy et al. 2000; Bandi et al. 2001; Casiraghi et al. 2002). In arthropods, Wolbachia often obligatorily live inside the cytoplasm in reproductive tissues and are associated with a number of different reproductive phenotypes in its hosts, such as cytoplasmic incompatibility (Shoemaker et al. 1999; Hurst and Werren 2001; Bordenstein and Wernegreen 2004; Jaenike et al. 2006), feminization, parthenogenesis inducing, male killing, and modifying fecundity (Hurst et al. 1999; Stouthamer et al. 1999; Baldo et al. 2005). These reproductive symptoms are regarded as selfish strategies of the symbionts whereby the frequency of female offspring increased (O'Neill et al. 1997; Werren 1997; Stouthamer et al. 1999; Haine 2008; Werren et al. 2008). Some Wolbachia strains also reduce host fitness by reducing fecundity or modifying growth rates (Chu et al. 2005), although the mechanism is not well understood.
Wolbachia cannot be cultured outside their hosts, so detection of infection has been based largely on amplification of Wolbachia DNA using allele-specific polymerase chain reactions (PCR). To date, wsp, 16S rDNA, ftsZ, groEL, coxA, fbpA, hcpA, gatB, dnaA and gltA genes have all been characterized and used for phylogenetic studies. Wolbachia strains are usually clustered into eight divergent clades based on these genes, which are described as supergroups A–H (O'Neill et al. 1992; Bandi et al. 1998; Zhou et al. 1998; Schulenburg et al. 2000; Werren and Windsor 2000; Lo et al. 2002; Bordenstein and Rosengaus 2005; Casiraghi et al. 2005) and IK (Gorham et al. 2003; Casiraghi et al. 2005; Ros et al. 2009), which were added later.
The Asiatic rice leafroller, Cnaphalocrocis medinalis (Guenée) (Lepidoptera: Pyralidae), is a migratory rice pest with 1–11 generations depending on its geographical distribution in rice-planting regions worldwide. C. medinalis also distributes widely in rice production areas from north to south in China, from Heilongjiang province and Inner Mongolia autonomous region to Taiwan and Hainan, excluding the Xinjiang and Ningxia autonomous region. In recent decades, C. medinalis has caused serious decreases in rice yields in China most notably due its outbreak from 2003–2005 (Liu et al. 2008).
In this study, the infection status of the endosymbiont Wolbachia in C. medinalis from twenty different regions of China was determined, and the genetic differentiation between the Wolbachia strains from Thailand and China were also analyzed. Study of the infection status and transmission mechanism of Wolbachia has been considered to be very helpful in utilization of insect natural enemies for pest control.
Materials and Methods
Insect sample collection
In July and October 2009, the samples of C. medinalis larvae were collected from 20 paddy fields in 15 provinces of China (Table 2). When sampling, C. medinalis 1st–4th instar larvae were collected, placed in 95% ethanol, marked with the location and time, and taken back to the laboratory for further analysis.
Table 2.
Cnaphalocrocis medinalis samples collected in China for Wolbachia detection.

DNA extraction
To extract DNA, the entire body was used if the larva was < 1 mm in length, while only the abdomen was used if larva size was ∼ 1–5 mm. DNA was extracted according to the description of Ahmed et al. (2009). Briefly, samples were washed several times by double distilled water and put into 1.5 ml centrifuge tubes with DNA extraction buffer (100 mmol/L Tris-HCl, pH 8.0, 50 mmol/L NaCl, 50 mmol/L EDTA, 1% SDS, 0.15 mmol/L Spermine, 0.5 mmol/L Spermidine) and proteinase K (20 mg/mL). Samples were homogenated and digested at 56° C for three hours. The homogenate was mixed afterwards with an equal volume of phenol for 10 minutes and centrifuged at 12,000 rpm for four minutes. The centrifugation was repeated twice, and chloroform (isoamyl alcohol 24:1) was used instead of phenol for the last repetition. The supernatant was precipitated overnight at -20° C and then centrifuged at 12,000 rpm for 20 minutes to sediment the DNA pellet. The pellet was then allowed to dry at room temperature. The dried DNA pellet was re-suspended in 30 µl of TE buffer (10 mM Tris-HCl, 1 mM EDTA, pH 8.0) and kept at 4° C until use for PCR.
Twenty larvae from each C. medinalis population were selected randomly for DNA extraction and further detection.
PCR amplification
Three pairs of primers were used to amplify the wsp, ftsZ and 16S rDNA fragments of different C. medinalis samples by PCR, according to the methods of Braig et al. (1998), Werren et al. (1995), and West et al. (1998). Polymerase chain reactions were done in 20 µl reaction volumes containing: 2 µl 10× PCR buffer (TakKara Bio, www.takarabio.com), 2 µl 25 mM MgCl2, 1.5 µl dNTPs (10 mM each), 1.0 µl forward primer, 20 µl reverse primer, and 1.2 units of Taq DNA polymerase (Takara). To achieve the final volume of 20 µl, double distilled H2O was added. For Wolbachia wsp gene amplification, the primers were wsp-F: 5′ TGGTCCAATAAGTGAGAGAAAC 3′ and wsp-R: 5′AAAAATTAAACGCTACTCCA 3′. The Wolbachia ftsZ PCR primers were ftsZ-F: 5′TACTGACTGTTGGAGTTGTAACTAAGC CGT 3′ and ftsZ-R: 5′-TGCCAG TTGAAGAAACTCTAACTC 3′; both the wsp and ftsZ primers can amplify a 0.6 kb DNA fragment (Zhou et al. 1998; Jeyaprakash and Hoy 2000). The primers for Wolbachia 16S rDNA PCR amplification were 16S-F: 5′ TTGTAGCCTGCTATGGTATAACT 3′ and 16S-R: 5′ GAATAGGAGTTTTCATGT 3′, amplifying a 0.9 kb DNA fragment (O'Neill et al. 1992).
The PCR amplification program of wsp and 16S rDNA primers included an initial denaturation at 94° C for three minutes followed by 35 cycles with a denaturation step at 94° C for one minute, annealing at 55° C for one minute, extension at 72° C for two minutes, and final extension at 72° C for 10 minutes. The ftsZ PCR amplification program was done according to three linked profiles. First, one cycle of denaturation at 94° C for three minutes followed by 10 cycles of denaturation at 94° C for 10 seconds, annealing at 65° C for 30 seconds, and extension at 68° C for one minute. This was followed by 25 cycles, each cycle with a denaturation step at 94° C for 10 seconds, annealing at 65° C for 30 seconds, extension at 68° C for one minute, plus an additional 20 seconds for each consecutive cycle. All PCR amplifications were done in a Thermal Cycler 48 (Bioer Technology, www.bioer.com.cn).
PCR Product Detection, Cloning, Sequencing and Analysis.
The amplified PCR products were electrophoresized on 1.0% agarose gel with water as a negative control, then cloned, screened, and two-way sequenced on an ABI PRISMTM 3730XL Automated DNA Sequencer (Applied Biosystems, www.appliedbiosystems.com). Six positive clones were sequenced per insect.
The wsp, ftsZ, and 16S rDNA sequences of Wolbachia from different C. medinalis populations were first blasted in NCBI, then analyzed and aligned with DNAStar (www.dnastar.com) and Clustal X1.83 (www.clustal.org). Some related Wolbachia wsp, ftsZ, and 16S rDNA sequences in other insects such as whitefly Bemisia tabaci and butterfly Pieris rapae were downloaded as references (Table 1). ANOVA was used to identify and compare differences among populations and all data was analyzed using DPS software (Tang and Feng 2002). Phylogenetic trees were constructed using Mega 4.0 software (MEGA, www.megasoftware.net) with maximum parsimony and neighbor joining methods (maximum likelihood model). Bootstrap analysis was done with 1000 replications, and bootstrap values were calculated using a 50% majority rule.
Table 1.
Reference sequences of Wolbachia used in the phylogenetic analyses.

Some reference sequences of Wolbachia from different insects as well as mites were downloaded from GenBank (Table 1) for Wolbachia phylogenetic analysis. Three reference sequences belonging to the Wolbachia A supergroup were used as the outgroup in the phylogenetic trees of wsp, ftsZ, and 16S rDNA; Ostrinia furnacalis (EU294311), Thecodiplosis japonensis (AF220605), and Drosophila simulans (AY227742).
Results
Wolbachia detection in C. medinalis by PCR
The infection of Wolbachia in different C. medinalis populations was detected by PCR with wsp, ftsZ, and 16S rDNA genes. Results showed that in total, 250 of 400 individual larvae from 20 populations were detected to be positive. The wsp and ftsZ PCR products were ∼ 600 bp (Figures 1 and 2) while the 16S rDNA PCR products of Wolbachia from C. medinalis were ∼ 900 bp (Figure 3). The align results with DNAStar software indicated that all the wsp sequences of Wolbachia from 250 C. medinalis individuals were 100% identical to each other. Similar results were obtained from the ftsZ sequences and 16S rDNA sequences.
Figure 1.
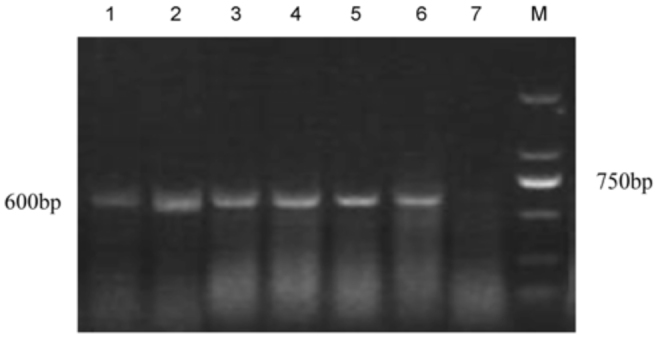
Electrophoresis of PCR products of Wolbachia wsp gene from Cnaphalocrocis medinalis by general primers. M: Molecular size standards, lane 7: negative control, lanes 1–6: different regions of C. medinalis (corresponding population number 1 to 6 in Table 2). High quality figures are available online.
Figure 2.
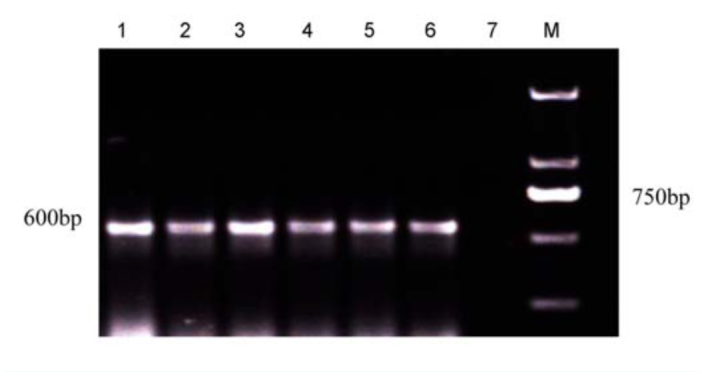
Electrophoresis of PCR products of Wolbachia ftsZ gene from Cnaphalocrocis medinalis by special primers. M: Molecular size standards, lane 7: negative control, lanes 1–6: different populations of Cnaphalocrocis medinalis (corresponding population code 1 to 6 in Table 2). High quality figures are available online.
Figure 3.
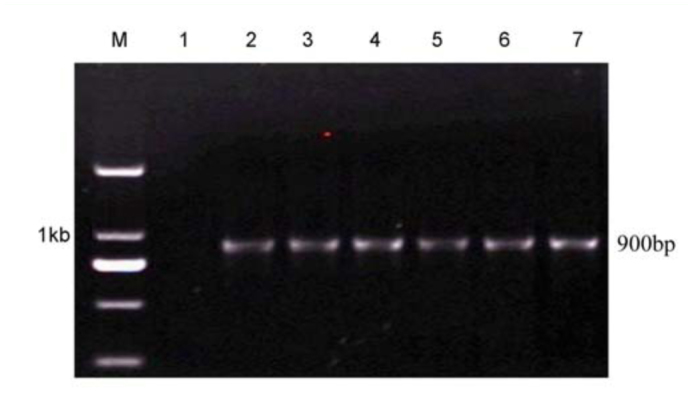
Electrophoresis of PCR products of Wolbachia 16S rDNA gene from Cnaphalocrocis medinalis by special primers. M: Molecular size standards, lane 1: negative control, lanes 2–7: different populations of C. medinalis (corresponding population code 1 to 6 in Table 2). High quality figures are available online.
The infection rate of Wolbachia in different C. medinalis populations
The infection rates of Wolbachia in the twenty C. medinalis populations collected from different geographical regions varied significantly (F = 2.3750, p < 0.01, df = 19), ranging from 40–90% with an average of 62.5% (Table 2). The highest infection rates were found in the C. medinalis Yangzhou and Wenzhou populations (90%), while the lowest rates were recorded in Ya' an, Changsha, and Tianjin populations (40%).
Phylogenetic analysis of the Wolbachia wsp, ftsZ, and 16S rDNA sequences
The phylogenetic analysis results of the Wolbachia wsp sequences from C. medinalis and other 21 reference sequences are shown in Figure 4a. There were two major branches in the phylogenetic tree based on Wolbachia wsp sequences (HQ336507). The first branch clustered Wolbachia sequences from the C. medialis Chinese population and the Scirpophaga incertulas (AF481197) Thailand population, containing 11 Wolbachia wsp genes of insects and mites from Japan, China, Korea, and Thailand, into a single clade. The second branch included six Wolbachia populations from five species of insects including Sitophilus zeamais (AB469362), Cadra cautella (AB469186), Neriene limbatinella (EU916191), Lycaena phlaeas (AB094377), and Smaragdina semiaurantiaca (GU236976-7). This second branch, while comprised of insects from different taxonomic families and orders, served as a sister branch with Paederus fuscipes (EU916190) and the first branch. However, one reference sequence of Wolbachia wsp gene from C. medialis Thai population has been found to be clustered in the periphery except near outgroup in the tree; the homology of the Wolbachia wsp sequences from C. medialis Chinese and Thai populations was only 83.2%.
Figure 4.

The phylogenetic trees of Wolbachia detected in Cnaphalocrocis medinalis and other insects and mites based on their wsp, ftsZ, 16S rDNA sequences. Trees inferred from maximum parsimony and neighbor joining methods (maximum likelihood model) using MEGA 4.0 program were similar though less resolved (data not shown.) The sequences of Ostrinia furnacalis (EU2943 11) and Thecodiplosis japonensis (AF220605), were used as outgroups in Fig. 4A and B, respectively. Additionally, the sequence of Drosophila simulons (AY227742) was used in Fig.4C. High quality figures are available online.
Figure 4b showed the phylogenetic analysis results based on the ftsZ gene sequnces of Wolabchia in C. medinalis and other hosts. Result revealed that the ftsZ sequence (HQ336508) in C. medinalis samples shared 100% identity with those sequences from Acraea encedon (AJ271199), Hypolimnas bolina (AB167399), Phyllonorycter quinnata (AJ005887) and Parornix devoniella (AJ005888). Four reference sequences from four species of lady beetles (EU627750, EU27753-5) clustered into one clade, showing 100% identity to each other and the lowest homology to the sequence from C. medinalis. The homology of all sequences of the Wolbachia ftsZ gene from the rice leaf roller and lady beetles was close to identical at 98.9%.
The phylogenetic tree of 16S rDNA sequences of Wolbachia was shown in Figure 4c. Similar to the wsp and ftsZ tree, no regular pattern was found in this tree. The 16S rDNA sequences (HQ336509) of Wolbachia from C. medinalis was first clustered into a subclade with Bactrocera dorsalis (DQ098949), sharing 99% identity, then subsequently was clustered into higher clades with other sequences from different hosts in various families and orders.
Discussion
Since first discovered in Culex pipiens (Hertig 1936), Wolbachia have been described as a widespread and common insect bacteria all over Neotropical (Borm et al. 2003), Palaearctic (West et al. 1998) and Nearctic regions (Werren and Windsor 2000). Samples of insect species from these three regions have almost the same Wolbachia infection rate (20%). Over 50% of a set of Southeast Asian ant species tested positive for Wolbachia (Wenseleers et al. 1998), Jeyaprakash and Hoy (2000) found over 76% of the samples of insect species were infected with Wolbachia, and in the current study, the average infection rate of Wolbachia in the Asiatic leafroller in China was 62.5%, ranging from 40 to 90% depending on their geographical distributions. Such findings support that Wolbachia is wide spread in numerous arthropods.
Wolbachia are mostly transmitted through egg cytoplasm of the hosts from parents to offspring, but several studies revealed that they did not have a consistent relationship between Wolbachia and hosts in phylogenetic trees (Schilthuizen and Stouthamer 1998; Zhou et al. 1998). It is supposed that there may be horizontal transmission between different hosts, including intra-species and inter-species transmission (Heath et al. 1999). This phenomenon has proven very common in spiders (Rowley et al. 2004) as well as between some arthropods and their parasitoids (Jeyaprakash and Hoy 2000), though the mechanism of this horizontal transmission is still not clear. For example, the horizontal transmission of Wolbachia has been found in a parastic wasp, Nasonia giraulti, and their blowfly hosts Protocalliphora sp., in some drosophilid parasitoids and their hosts, as well as in the parasitoid Leptopilina boulardi and their fly host D. simulans (Werren et al. 1995; Heath et al. 1999; Vavre et al. 1999). In our study, no regular pattern was found in any of the three phylogenetic trees for the genetic relationship between C. medinalis and other insects or mites; the Wolbachia sequences from C. medinalis clustered into one clade with different insects or mites in different trees. This suggests two conclusions. First, while there was no direct evidence to verify the horizontal transmission of Wolbachia between their hosts, the pattern of transmission of Wolbachia was not limited within host species and by geographical locations. Second, as the Wolbachia from C. medinalis Chinese population was clustered into one clade with different insects or mites in different trees, it was not possible to determine the transmission among arthropods in the field by comparing one phylogenetic tree using single genes.
In the current study, all the Wolbachia sequences from the different geographical populations were 100% identical to each other, which suggested that no evolutionary differentiation had occurred in China. This may have been due to the migration of C. medinalis since it is a migratory pest. For example, during population expansion, the Wolbachia—infected males and females could have spread widely in China, thus increasing the opportunities of Wolbachia being transmitted through a broader range by mating between or among individuals from the same or different regions. For example, Huigens et al. (2000 and 2004) found frequent horizontal transmission from infected to uninfected wasp larvae that shared a common food source. Frequent horizontal transmission occurred between infected and uninfected Trichogramma kaykai, Trichogramma deion, Trichogramma pretiosurm, Trichogramma atopovirilia when eggs were laid in their common host Apodemia mormodeserti. The transferred Wolbachia were then vertically transmitted to the new host's offspring. However, this aspect needs further research to reveal this complex mechanism.
Kittayapong et al. (2003) used Long PCR and long wsp primers to investigate the infection status of Wolbachia in C. medinalis collected from rice fields in 29 provinces of Thailand. They showed that the average infection rate in the C. medinalis Thai populations was 48.8%, which was much lower than the infection rate in our study. This difference is possibly related to sample size. Similar to those found in China, all the detected Wolbachia sequences using wsp gene in 29 provinces of Thailand were 100% identical and belong to the Wolbachia B supergroup. However, large differentiation between the Wolbachia wsp sequences from C. medinalis Chinese and Thai populaions was found; sequence homology was 83.2%. It is thought that the C. medinalis population in China may have come from Southeast Asia and entered into China by the first northward mass migration during early March to April. Additionally, C. medinalis populations from different regions of China may be those migrants from Thailand or their offspring. Thus, Wolbachia in C. medinalis Chinese and Thai populations should be the same strain or quite close to each other (Chang et al. 1980; Gao et al. 2008). However, the results in our study showed large differences between the Wolbachia wsp sequences in the C. medinalis Chinese and Thai populations. We speculate that either the first northward mass migration of C. medinalis was not from the direction of Thailand, or that Wolbachia in Chinese populations may have displaced the strain of Wolbachia in Thailand populations when C. medinalis migrated to China from Thailand. Further investigation in this area is needed.
In summary, the infection status of Wolbachia in C. medinalis Chinese populations was investigated and the average infection rate of Wolbachia was found to be 62.5%. The phylogenetic trees based on the wsp, fstZ, and 16S rDNA revealed the possibility of interand intra-species horizontal transmission of Wolbachia in different arthropods. In view of the biological roles of Wolbachia in their hosts, especially for host reproduction such as cytoplasmic incompatibility and male killing, further studies on how to make good use of the transmission patterns of Wolbachia to enhance the biological control of pests deserves more emphasis (Gong and Shen 2002; Tsai et al. 2002; Miao et al. 2004; Zabalou et al. 2004; Pfarr and Hoerauf, 2005; Ruan and Liu 2005).
Acknowledgements
This work was supported by Special Fund for Agro-Scientific Research in the Public Interest of China (200903051) and National Basic Research and Development Program (973) of China (2006CB1020002).
References
- Ahmed MZ, Shatters RG, Ren SX, Jin GH, Mandour NS, Qiu BL. Genetic distinctions among the Mediterranean and Chinese populations of Bemisia tabaci Q biotype and their endosymbiont Wolbachia populations. Journal of Applied Entomology. 2009;133:733–741. [Google Scholar]
- Baldo L, Lo N, Werren JH. Mosaic nature of the Wolbachia surface protein. Journal of Bacteriology. 2005;187:5406–5418. doi: 10.1128/JB.187.15.5406-5418.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bandi C, Anderson TJ, Genchi C, Blaxter ML. Phylogeny of Wolbachia in filarial nematodes. Proceedings of the Royal Society of London Series B. 1998;265:2407–2413. doi: 10.1098/rspb.1998.0591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bandi C, McCall JW, Genchi C, Corona S, Venco L, Sacchi L. Effects of tetracycline on the filarial worms Brugia pahangi and Dirofilaria immitis and their bacterial endosymbionts Wolbachia. International Journal of Parasitology. 1999;29:357–364. doi: 10.1016/s0020-7519(98)00200-8. [DOI] [PubMed] [Google Scholar]
- Bandi C, Trees AJ, Brattig NW. Wolbachia in Filarial nematodes: evolutionary aspects and implications for the pathogenesis and treatment of filarial diseases. Veterinary Parasitology. 2001;98:215–238. doi: 10.1016/s0304-4017(01)00432-0. [DOI] [PubMed] [Google Scholar]
- Bordenstein SR, Rosengaus RB. Discovery of a novel Wolbachia supergroup in Isoptera. Current Microbiology. 2005;51:393–398. doi: 10.1007/s00284-005-0084-0. [DOI] [PubMed] [Google Scholar]
- Bordenstein SR, Wernegreen JJ. Bacteriophage Xux in endosymbionts (Wolbachia): infection frequency, lateral transfer, and recombination rates. Molecular Biology and Evolution. 2004;21:1981–1991. doi: 10.1093/molbev/msh211. [DOI] [PubMed] [Google Scholar]
- Borm SV, Wenseleers T, Billen J, Boomsma JJ. Clone and sequencing of wsp encoding gene fragments reveals a diversity of coinfecting Wolbachia strains in Acromyrmex leafcutter ants. Molecular Phylogenetics and Evolution. 2003;26:102–109. doi: 10.1016/s1055-7903(02)00298-1. [DOI] [PubMed] [Google Scholar]
- Braig HR, Zhou W, Dobson SL, O'Neill SL. Cloning and characterization of a gene encoding the major surface protein of the bacterial endosymbiont Wolbachia. Journal of Bacteriology. 1998;180:2373–2378. doi: 10.1128/jb.180.9.2373-2378.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casiraghi M, Bordenstein SR, Baldo L, Lo N, Beninati T, Wernegreen JJ, Werren JH, Bandi C. Phylogeny of Wolbachia pipientis based on gltA, groEL and ftsZ gene sequences: clustering of arthropod and nematode symbionts in the F supergroup, and evidence for further diversity in the Wolbachia tree. Microbiology. 2005;151:4015–4022. doi: 10.1099/mic.0.28313-0. [DOI] [PubMed] [Google Scholar]
- Casiraghi M, McCall JW, Simoncini L, Kramer LH, Sacchi L, Genchi C, Werren JH, Bandi C. Tetracycline treatment and sex-ratio distortion: a role for Wolbachia in the moulting of filarial nematodes? International Journal of Parasitology. 2002;32:1457–1468. doi: 10.1016/s0020-7519(02)00158-3. [DOI] [PubMed] [Google Scholar]
- Chang SS, Lo ZC, Keng CG, Li GZ, Chen, Xl W. Studies on the migration of rice leaf roller Cnaphalocrocis medinalis Guenee. Acta Entomologica Sinica. 1980;23:130–140. [Google Scholar]
- Chu D, Zhang YJ, Bi YP, Fu HB. Wolbachia endosymbionts and their effects on the fitness of the arthropod hosts. Acta Microbiol Sinica. 2005;45:817–820. [PubMed] [Google Scholar]
- Cordaux R, Michel-Salzat A, Bouchon D. Wolbachia infection in crustaceans: novel hosts and potential routes for horizontal transmission. Journal of Evolutionary Biology. 2001;14:237–243. [Google Scholar]
- Gao YB, Chen X, Bao YX, Yang RM, Liu TL, Zhai BP. Dynamic analysis on the migration of the rice leaf roller Cnaphalocrocis medinalis (Lepidoptera: Pyralidae) by Doppler Insect Monitoring Radar and numerical simulation. Acta Ecologica Sinica. 2008;28:5238–5237. [Google Scholar]
- Gong P, Shen ZR. Wolbachia endosymbionts and their manipulation of reproduction of arthropod hosts. Acta Entomologica Sinica. 2002;45:241–252. [Google Scholar]
- Gorham CH, Fang QQ, Durden LA. Wolbachia endosymbionts in fleas (Siphonaptera). Journal of Parasitology. 2003;89:283–289. doi: 10.1645/0022-3395(2003)089[0283:WEIFS]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- Gotoh T, Noda H, Hong XY. Wolbachia distribution and cytoplasmic incompatibility based on a survey of 42 spider mite species (Acari: Tetranychidae) in Japan. Heredity. 2003;91:208–216. doi: 10.1038/sj.hdy.6800329. [DOI] [PubMed] [Google Scholar]
- Haine ER. Symbiont-mediated protection. Proceedings of the Royal Society of London Series B. 2008;275:353–361. doi: 10.1098/rspb.2007.1211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heath BD, Butcher RD, Whitfield WG, Hubbard SF. Horizontal transfer of Wolbachia between phylogenetically distant insect species by a naturally occurring mechanism. Current Biology. 1999;9:313–316. doi: 10.1016/s0960-9822(99)80139-0. [DOI] [PubMed] [Google Scholar]
- Hertig M. The rickettsia, Wolbachia pipientis (gen.et sp.n.) and associated inclusions of the mosquite Culex pipiens. Parasitology. 1936;28:453–486. [Google Scholar]
- Hilgenboecker K, Hammerstein P, Schlattmann P, Telschow A, Werren JH. How many species are infected with Wolbachia? — a statistical analysis of current data. FEMS Microbiology Letters. 2008;281:215–222. doi: 10.1111/j.1574-6968.2008.01110.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hurst GDD, Jiggins FM, Von der Schulenburg JHG, Bertrand D, West SA, Goriachev II, Zakharov IA, Werren JH, Stouthamer R, Majerus MEN. Male killing Wolbachia in two species of insects. Proceedings of the Royal Society of London Series B. 1999;266:735–740. [Google Scholar]
- Hurst GDD, Werren JH. The role of selWsh genetic elements in eukaryotic evolution. Nature Reviews Genetics. 2001;2:597–606. doi: 10.1038/35084545. [DOI] [PubMed] [Google Scholar]
- Huigens ME, Almeida RP, Boons PAH, Luck RF, Stouthamer R. Natural interspeciecific and intraspecific horizontal transfer of parthenogenesis-inducing Wolbachia in Trichogramma wsaps. Proceedings of the Royal Society of London Series B. 2004;271:509–515. doi: 10.1098/rspb.2003.2640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huigens ME, Luck RF, Klaassen RHG, Maas MFPM, Timmermans MJTN, Stouthamer R. Infectious parthenogenesis. Nature. 2000;405:178–189. doi: 10.1038/35012066. [DOI] [PubMed] [Google Scholar]
- Jaenike J, Dyer KA, Cornish C, Minhas MS. Asymmetrical reinforcement and Wolbachia infection in Drosophila. PLoS Biology. 2006;4(10):e325. doi: 10.1371/journal.pbio.0040325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeyaprakash A, Hoy MA. Long PCR improves Wolbachia DNA amplification: wsp sequences found in 76% of sixty-three arthropod species. Insect Molecular Biology. 2000;9:393–405. doi: 10.1046/j.1365-2583.2000.00203.x. [DOI] [PubMed] [Google Scholar]
- Kittayapong P, Jamnongluk W, Thipaksorn A, Milne JR, Sindhusake C. Wolbachia infection complexity among insects in the tropical rice-field community. Molecular Ecology. 2003;12:1049–1060. doi: 10.1046/j.1365-294x.2003.01793.x. [DOI] [PubMed] [Google Scholar]
- Langworthy NG, Renz A, Mackenstedt U, Henkleduhrsen K, De Bronsvoort MB, Tanya VN, Donnelly MJ, Trees AJ. Macrofilaricidal activity of tetracycline against the filarial nematode Onchocerca ochengi: elimination of Wolbachia precedes worm death and suggests a dependent relationship. Proceedings of the Royal Society of London Series B. 2000;267:1063–1069. doi: 10.1098/rspb.2000.1110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu Y, Wang JQ, Feng XD, Jiang XH. Analysis on the occurring of Cnaphalocrocis medinalis in 2007 and forecasting its occuring trends in 2008. China Plant Protection. 2008;28:33–35. [Google Scholar]
- Lo N, Casiraghi M, Salati E, Bazzocchi C, Bandi C. How many Wolbachia supergroups exist? Molecular Biology and Evolution. 2002;19:341–346. doi: 10.1093/oxfordjournals.molbev.a004087. [DOI] [PubMed] [Google Scholar]
- Miao H, Hong XY, Xie L, Xue XF. Sequencing and sequence analysis of the wsp gene of Wolbachia in Tetranychus cinnabarinus (Acari: Tetranychidae). Acta Entomologica Sinica. 2004;47:738–743. [Google Scholar]
- O'Neill SL, Giordano R, Colbert AM, Karr TL, Robertson HM. 16S rRNA phylogenetic analysis of the bacterial endosymbionts associated with cytoplasmic incompatibility in insects. Proceedings of the National Academy of Science USA. 1992;89:2699–2702. doi: 10.1073/pnas.89.7.2699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Neill SL, Hoffmann AA, Werren JH. Influential Passengers: inherited microorganisms and arthropod reproduction. Oxford University Press; 1997. [Google Scholar]
- Pfarr KM, Hoerauf AM. The annotated genome of Wolbachia from the filarial nematode Brugia malayi: what it means for progress in antifilarial medicine. PLoS Medicine. 2005;2(4):e110. doi: 10.1371/journal.pmed.0020110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ros VI, Fleming VM, Feil EJ, Breeuwer JA. How Diverse is the Genus Wolbachia? Multiple-Gene Sequencing Reveals a Putatively New Wolbachia Supergroup Recovered from Spider Mites (Acari: Tetranychidae). Applied and Environmental Microbiology. 2009;4:1036–1043. doi: 10.1128/AEM.01109-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rowley SM, Raven RJ, McGraw EA. Wolbachia pipientis in Australian spiders. Current Microbiology. 2004;49:208–214. doi: 10.1007/s00284-004-4346-z. [DOI] [PubMed] [Google Scholar]
- Ruan YM, Liu SS. Detection and phylogenetic analysis of prokaryotic endosymbionts in Bemisia tabaci. Acta Entomologica Sinica. 2005;48:859–865. [Google Scholar]
- Schilthuizen M, Stouthamer R. Distribution of Wolbachia among the guild associated with the parthenogenetic gall wasp Diplolepis rosae. Heredity. 1998;81:270–274. [Google Scholar]
- Schulenburg JH, Hurst GD, Huigens TM, Van Meer MM, Jiggins FM, Majerus ME. Molecular evolution and phylogenetic utility of Wolbachia ftsZ and wsp gene sequences with special reference to the origin of malekilling. Molecular Biology and Evolution. 2000;17:584–600. doi: 10.1093/oxfordjournals.molbev.a026338. [DOI] [PubMed] [Google Scholar]
- Shoemaker DD, Katju V, Jaenike J. Wolbachia and the evolution of reproductive isolation between Drosophila recens and Drosophila subquinaria. Evolution. 1999;53:1157–1164. doi: 10.1111/j.1558-5646.1999.tb04529.x. [DOI] [PubMed] [Google Scholar]
- Stouthamer R, Breeuwer JA, Hurst GDD. Wolbachia pipientis: Microbial manipulator of arthropod reproduction. Annual Review of Microbiology. 1999;53:71–102. doi: 10.1146/annurev.micro.53.1.71. [DOI] [PubMed] [Google Scholar]
- Tang QY, Feng MG. DPS data processing system for practical statistics. Science Press; 2002. [Google Scholar]
- Tsai KH, Wu WJ, Chen WJ. Biological Characteristics of the Endosymbiont Wolbachia. Formosan. Entomology. 2002;22:1–18. [Google Scholar]
- Vavre F, Fleury F, Lepetit D, Fouillet P, Bouletreau M. Phylogenetic evidence for horizontal transmission of Wolbachia in host-parasitoid associations. Molecular Biology and Evolution. 1999;16:1711–1723. doi: 10.1093/oxfordjournals.molbev.a026084. [DOI] [PubMed] [Google Scholar]
- Wenseleers T, Ito F, Van Borm S, Huybrechts R, Volckaert F, Billen J. Widespread occurrence of the microorganism Wolbachia in ants. Proceedings of the Royal Society of London Series B. 1998;265:1447–1452. doi: 10.1098/rspb.1998.0456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Werren JH. Biology of Wolbachia. Annual Review of Entomology. 1997;42:587–609. doi: 10.1146/annurev.ento.42.1.587. [DOI] [PubMed] [Google Scholar]
- Werren JH, Baldo L, Clark ME. Wolbachia: master manipulators of invertebrate biology. Nature Reviews Microbiology. 2008;6:741–751. doi: 10.1038/nrmicro1969. [DOI] [PubMed] [Google Scholar]
- Werren JH, Windsor DM. Wolbachia infection frequencies in insects: evidence of a global equilibrium? Proceedings of the Royal Society of London Series B. 2000;267:1277–1285. doi: 10.1098/rspb.2000.1139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Werren JH, Zhang W, Guo LR. Evolution and phylogeny of Wolbachia: reproductive parasites of arthropods. Proceedings of the Royal Society of London Series B. 1995;261:55–71. doi: 10.1098/rspb.1995.0117. [DOI] [PubMed] [Google Scholar]
- West SA, Cook JM, Werren JH, Godfray HC. Wolbachia in two insect host parasitoid communities. Molecular Ecology. 1998;7:1457–1465. doi: 10.1046/j.1365-294x.1998.00467.x. [DOI] [PubMed] [Google Scholar]
- Zabalou S, Riegler M, Theodorakopoulou M, Stauffer C, Savakis C, Bourtzis K. Wolbachia- induced cytoplasmic incompatibility as a means for insect pest population control. Proceedings of the National Academy of Science USA. 2004;101:15042–15045. doi: 10.1073/pnas.0403853101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou W, Rousset F, O'Neill SL. Phylogeny and PCR-based classification of Wolbachia strains using wsp gene sequences. Proceedings of the Royal Society of London Series B. 1998;265:509–515. doi: 10.1098/rspb.1998.0324. [DOI] [PMC free article] [PubMed] [Google Scholar]