

Abstract
Background
Tarebia granifera (Lamarck, 1822) is originally from South-East Asia, but has been introduced and become invasive in many tropical and subtropical parts of the world. In South Africa, T. granifera is rapidly invading an increasing number of coastal lakes and estuaries, often reaching very high population densities and dominating shallow water benthic invertebrate assemblages. An assessment of the feeding dynamics of T. granifera has raised questions about potential ecological impacts, specifically in terms of its dietary overlap with native gastropods.
Methodology/Principal Findings
A stable isotope mixing model was used together with gut content analysis to estimate the diet of T. granifera and native gastropod populations in three different coastal lakes. Population density, available biomass of food and salinity were measured along transects placed over T. granifera patches. An index of isotopic (stable isotopes) dietary overlap (IDO, %) aided in interpreting interactions between gastropods. The diet of T. granifera was variable, including contributions from microphytobenthos, filamentous algae (Cladophora sp.), detritus and sedimentary organic matter. IDO was significant (>60%) between T. granifera and each of the following gastropods: Haminoea natalensis (Krauss, 1848), Bulinus natalensis (Küster, 1841) and Melanoides tuberculata (Müller, 1774). However, food did not appear to be limiting. Salinity influenced gastropod spatial overlap. Tarebia granifera may only displace native gastropods, such as Assiminea cf. ovata (Krauss, 1848), under salinity conditions below 20. Ecosystem-level impacts are also discussed.
Conclusion/Significance
The generalist diet of T. granifera may certainly contribute to its successful establishment. However, although competition for resources may take place under certain salinity conditions and if food is limiting, there appear to be other mechanisms at work, through which T. granifera displaces native gastropods. Complementary stable isotope and gut content analysis can provide helpful ecological insights, contributing to monitoring efforts and guiding further invasive species research.
Introduction
Alien invasive species (AIS) can cause disruptions to ecosystems. However, quantifying their impacts is problematic due to the complexity of ecological interactions [1], [2]. Stable isotope analysis is a powerful quantitative tool for detecting and tracking changes in trophic structure and ecosystem processes (energy flows) caused by abiotic and biotic interactions [3]. The combination of stable isotopes with other techniques is very useful for assessing interactions between several organisms and contributes towards a better understanding of how an ecosystem can be affected by native and non-native species [4], [5]. Stable isotope analysis provides a time-integrated view of ecological processes, whereas gut content analysis reveals snapshots of feeding activity [6]. Stable isotope and gut content analyses are complementary, provide insight into diets and trophic dynamics in ecosystems [7], [8] and have successfully been used in several studies addressing the impacts of AIS on aquatic ecosystems [8]–[12]. Although comparable studies on gastropods are lacking, numerous studies have been published on invasive gastropods such as Pomacea canaliculata (Lamarck, 1822) (prosobranch, Ampullariidae) [13]–[15] and Potamopyrgus antipodarum (Gray, 1843) (prosobranch, Hydrobiidae) [16], [17], some of which involved stable isotope techniques [18], [19].
Tarebia granifera (Lamarck, 1822) (prosobranch, Thiaridae) is a non-native invasive gastropod originally from South-East Asia. This AIS is reported from many tropical and sub-tropical areas of North and South America and Africa [20]–[24]. Tarebia granifera was accidentally introduced in South Africa, probably in the 1990s via the aquarium trade [24]. This is reported as one of the most recent introductions of non-native gastropod into South African natural environments, where it has quickly become invasive and widespread, particularly in the KwaZulu-Natal and Mpumalanga provinces [24], [25]. Tarebia granifera is parthenogenetic and ovoviviparous, giving birth to live juveniles and often reaching population densities of over 1000 ind. m−2 [26]. Tarebia granifera has been successfully used as a biological control of schistosomiasis in the Caribbean, since it is able to displace host native gastropods [27]–[29]. In South Africa, T. granifera is rapidly invading an increasing number of coastal lakes and estuaries, so there is concern about potential ecological disturbances due to the displacement of native invertebrates [24], [25], [30]. Tarebia granifera can have a very high feeding impact and may out-compete native gastropods for food resources [31]. However, the mechanisms through which T. granifera may compete with native invertebrates are not yet clear, and questions about its diet still need to be addressed [31].
This study aims to: (1) use gut content and stable isotope analyses to estimate the diets of T. granifera and other dominant gastropod populations; and (2) estimate dietary niche overlap between invasive T. granifera and native gastropods. The framework of this field-based study also assesses spatial overlap between gastropods as well as availability of food in the form of benthic microalgae. The potential of T. granifera to affect trophic dynamics, and compete with native gastropods for food resources, is discussed in light of the findings.
Materials and Methods
Ethics Statement
All necessary permits were obtained from the iSimangaliso Wetland Park Authority for the described field studies at each location, under a Research Agreement for the project titled “Climate Change and the Management of KZN estuaries: St Lucia Estuary”.
Study site
Sampling was conducted at three different coastal lakes which were all invaded by Tarebia granifera over the last decade (Fig. 1).
Figure 1. Study area.

The Kosi Lakes, Lake Sibaya and the St. Lucia Estuary are Ramsar Wetlands of International Importance within the iSimangaliso Wetland Park, a UNESCO World Heritage Site in Maputaland, northern KwaZulu-Natal, South Africa. Sites sampled are marked as dots on the map.
The St. Lucia Estuary was sampled in June 2007, October 2009 and February 2010. It is the largest estuarine lake in Africa, with a surface area of about 325 km2 and average depth of 0.9 m [32]. The sample site, Catalina Bay, is a large limestone flat covered by shallow water and is located on the eastern shores of South Lake (28°13′S, 032°29′E). Vegetation found by the shore included Cyperus laevigatus, Juncus kraussii and Salicornia spp.. Throughout the sampling period, Catalina Bay experienced significant changes in salinity and water level. These were underlined by an unusual mouth breaching event, caused by extreme wave action in the Indian Ocean during March 2007, which resulted in six months of open mouth conditions [33]. Prior to this, the estuary had been completely isolated from the ocean for almost five years.
Tarebia granifera has persisted in the St. Lucia Estuary at least since 2005, but its invasion is restricted to freshwater seepage areas along the eastern shores of the South Lake, where it tends to dominate benthic assemblages. Shallow water benthic assemblages are otherwise dominated by the native gastropods Assiminea cf. ovata and Haminoea natalensis. Assiminea cf. ovata (Krauss, 1848) (previously known as A. bifasciata) (prosobranch, Assimineidae) is a small (shell height ≈5 mm) native gastropod with a very wide salinity tolerance that can be found in river mouths, estuaries and lagoons on the east coast of South Africa and Mozambique [34]. Assiminea cf. ovata is thus well adapted to the hydrological dynamics of the St. Lucia Estuary [35], [36]. Haminoea natalensis (Krauss, 1848) (opistobranch, Haminoeidae) occurs from Port St Johns (Eastern Cape) to southern Mozambique [37] and periodically dominates the benthic assemblage along the shallow saline shores of South Lake (pers. obs.).
Lake Sibaya's south-east basin (27°25′S, 032°41′E) was sampled in November 2009 (Fig. 1). This freshwater lake has a surface area of 60 to 77 km2 and an average depth of 13 m [38]. It was connected to the ocean in the past and has a number of originally marine and estuarine species uniquely adapted to freshwater conditions [39]. The profile of the littoral zone is steep and, at the time of the study, the level of the lake was low according to previous records [40]. The substrate was composed of white sand and vegetation at the water edge included Phragmites sp., Typha latifolia and Cyperus sp.. Submerged macrophytes, such as Potamogeton spp. were dominant close to shore. Tarebia granifera dominated the benthic community in the relatively small shallow terrace, where shelter from wave action was provided by vegetation. It is unclear when T. granifera was introduced, but its invasion has spread at least along the entire eastern shallow shores of Lake Sibaya. The native Bulinus natalensis (Küster, 1841) (pulmonate, Planorbidae) was mostly found on submerged macrophytes. Bulinus natalensis occurs on the lowlands of KwaZulu-Natal, South Africa [41].
Kosi Lakes' Lake Nhlange (26°57′S, 032°49′E) was sampled in November 2008 (Fig. 1). The lake has a surface area of 30.7 to 37 km2 and an average depth of 7.2 m [42]. Lake Nhlange experiences freshwater conditions, despite being part of a system that is connected to the Indian Ocean [42]. The substrate was composed of clear white sand and marginal vegetation included Phragmites sp, Cyperus sp. and Juncus kraussii. Submerged macrophytes Ceratophyllum demersum and Nymphaea spp. were found at the water's edge. The benthic assemblage in the sampling area was once again dominated by T. granifera, with a few Melanoides tuberculata individuals also found occasionally. Tarebia granifera was found at boat launching areas where it was noticed only in recent years. Melanoides tuberculata (Müller, 1774) (prosobranch, Thiaridae) is composed of a great variety of lineages or morphs which are conchologically and genetically distinct [22]. Melanoides tuberculata has a worldwide natural distribution, including tropical Africa, Asia and Oceania. An alien morph of M. tuberculata from south-east Asia has invaded Africa [43]–[45]. Although it is assumed that the M. tuberculata in this study is native, this remains to be confirmed.
Transects
Tarebia granifera populations tended to be concentrated in dense patches. To assess spatial overlap between T. granifera and other gastropods, 4 point transects were placed parallel to the shore line and across patches of T. granifera. Transects were selected where T. granifera overlapped with a single native gastropod species. Transect lengths varied depending on the area covered by the T. granifera patch. Points were placed at even spaces so that points 1 and 4 were at the edge of the T. granifera patch and points 2 and 3 were within the patch. Samples were collected in triplicate with a Zabalocki-type Ekman grab (sampling area 0.0236 m2) and fixed in 10% formalin. In the laboratory, gastropods were counted and densities (ind. m−2) were calculated. Shell heights (SH) were measured with Vernier calipers. Salinity was measured with a YSI 6920 multiprobe.
To estimate microphytobenthos (MPB) biomass, duplicate sediment cores were collected (upper cm only) with a Perspex corer (internal diameter: 20 mm) at every point along transects. Chlorophyll a and phaeopigments were extracted in 90% acetone at 4°C over a 48 h period and then measured using a 10-AU Turner Designs fluorometer fitted with a narrow-band, non-acidification system [46]. Chlorophyll a and phaeopigment concentrations are added and reported as pigment concentration (mg. m−2) [47].
Gastropod gut content analysis
Twenty individuals from each species were randomly collected along each transect (see above) and immediately preserved in 10% formalin to prevent further digestion [48]. The digestive tract was later removed and the gut contents were extracted with a fine pipette under a dissecting microscope (40× magnification). Gut contents were then viewed under an inverted microscope (400× magnification) and classified based on gross morphology [49]. The following classes were used: microalgae, detritus, filamentous algae, and sand particles.
Stable isotope samples
Gastropods and their potential food sources were collected and stored separately, first in a cooler box and later in a freezer at −20°C, before being processed and analysed.
Gastropods were collected at random along each transect (see above). Muscle tissue from the foot of individual snails was dissected and pooled to create representative composites. Five replicates were prepared for each species, containing either 2 or 10 individuals each (depending on size).
Potential types of food sources for gastropods were sampled in triplicate along each transect. Reeds, sedges, grasses and macrophytes as well as filamentous algae (Cladophora sp.) and detritus (DTR) were collected and thoroughly cleaned in distilled water to remove sediment particles and organisms. These samples were then independently homogenized and freeze dried with a mortar and pestle and liquid nitrogen. Sediment cores were collected with a Perspex corer (diameter 7 cm). The upper cm layer of sediment was suspended in a 2 L container of filtered water and stirred, so that microphytobenthos (MPB) stayed in suspension while sediment and sedimentary organic matter (SOM) settled to the bottom. The sediment was rinsed thoroughly with distilled water, treated with 1 M hydrochloric acid (HCl) for 24 h to remove carbonates, rinsed with distilled water, and then homogenized and freeze-dried as described above. The supernatant containing MPB was collected on a pre-combusted GF/F filter. MPB samples were treated with excess 2% HCl over 2 h, rinsed with distilled water, dried in an air-circulating oven at 60°C for 24 h and wrapped in tin foil prior to being sent for isotopic analysis. All other samples were weighted to appropriate quantities for analysis (5×8 mm tin capsules with approximately 1 mg sample or 20 mg sample in Eppendorf microcentrifuge tubes in the case of sediment).
Stable isotope analysis
Carbon and nitrogen stable isotope ratios and C/N values were measured at the Stable Light Isotope Unit of the Department of Archaeology, University of Cape Town, South Africa. A Flash EA 1112 series elemental analyzer (Thermo Finnigan, Italy) was used with a Delta Plus XP IRMS (isotope ratio mass spectrometer) (Thermo electron, Germany) and a Conflo III gas control unit (Thermo Finnigan, Germany). Isotopic ratios were expressed as δ values (‰) relative to the Vienna PeeDee Belemnite standard for carbon and to atmospheric N2 standard for nitrogen according to:
Where X is 13C or 15N and R is the corresponding ratio of 13C/12C or 15N/14N. A lipid normalization procedure was not followed [50], since a mathematical normalization (e.g. [51]) did not significantly affect the major outcomes of these analyses.
Highest average trophic position (TP) of gastropods was calculated with the following equation (adapted in [4]):
where δ15Nconsumer is the average δ15N signature measured directly from a gastropod species, δ15Nbase is the average δ15N signature of the most δ15N depleted food source, and Δn is the assumed average enrichment in δ15N per trophic level (Δn = 2).
Data analysis
The Bayesian isotopic mixing model SIAR v 4.0 (Stable Isotope Analysis in R, [52]) was used to generate probability function distributions, showing the most feasible solutions to the contribution of different types of potential food sources to the diet of gastropods, with 95%, 75% and 25% credible intervals. Fractionation correction values of 0.4‰ δ13C and 2‰ δ15N were assumed [53]. Preliminary analyses were run with the aim to determine the appropriate number of sources in the mixing models [54], [55]. Based on results from gut content analyses and mixing model trial runs, a narrower set of sources was selected for each gastropod species, which provided a better resolution of results [56].Sources that were not found in the gut content analysis and/or had minor contributions (<5%) in the trial mixing models were omitted from the final mixing models. Isotopic dietary overlap (IDO, %) between species j and k was then calculated with the following equation based on Schoener's Index [57], [58]:
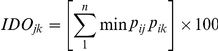 |
where p is the SIAR mean contribution of source i resulting from the final mixing models run for subjects j and k respectively, and n is the total number of different resources used by both j and k. IDO values have an absolute limit of 100%, which indicates complete overlap, and values exceeding 60% were considered to indicate significant dietary overlap [57].
Normality and equality of variance of stable isotope data were confirmed, and analysis of variance (ANOVA) was used for comparisons between locations and times. Pearson correlation was used to assess relationships between availability of food, gastropod abundance and diet. A t-test was used to compare trophic position between T. granifera and native gastropods. A linear regression was used to assess the relationship between gut content and stable isotope results. Analyses were done with the statistical package SPSS version 19 for Windows.
Results
Spatial overlap and resource availability
Tarebia granifera exhibited a patchy distribution and tended to be concentrated in shallow water (depth <1 m). Transects revealed spatial overlap between T. granifera and Haminoea natalensis (Fig. 2A), Assiminea cf. ovata (Fig. 3A and 4A), Bulinus natalensis (Fig. 5A) and Melanoides tuberculata (Fig. 6A) at respective sampling sites in the three lakes of Maputaland (Fig. 1). Tarebia granifera was found together with native gastropods in a total of 10 of the 20 transect points sampled. Of the 10 transect points where these gastropods were found together, 7 were at the edge of the T. granifera patch (transect points 1 and 4). Haminoea natalensis was the only native gastropod not to be found in the middle (transect points 2 and 3) of a T. granifera patch.
Figure 2. Gastropods, food sources and diets in Catalina Bay, 2007.

(A) Four-point 6 m transect showing gastropod densities and microphytobenthic (MPB) biomass (as chl-a concentration). (B) Carbon and nitrogen stable isotope signatures of gastropods (number in brackets: trophic position) and their potential food sources; such as detritus (DTR) and sedimentary organic matter (SOM). SIAR boxplots show the proportional contribution (%) of different food sources to the diet of (C) Tarebia granifera and (D) Haminoea natalensis. Samples were collected in a freshwater seepage area of Catalina Bay in June 2007. Salinity ranged from 16 to 32. The water level of the South Lake rose due to the March 2007 mouth breach, after which the freshwater ponds associated with seepage areas along the eastern shores were flooded by seawater.
Figure 3. Gastropods, food sources and diets in Catalina Bay, 2009.

(A) Four-point 6 m transect showing gastropod densities and microphytobenthic (MPB) biomass (as chl-a concentration). (B) Carbon and nitrogen stable isotope signatures of gastropods (number in brackets: trophic position) and their potential food sources, such as detritus (DTR) and sedimentary organic matter (SOM). SIAR boxplots show the proportional contribution (%) of different sources to the diet of (C) Tarebia granifera, (D) Assiminea cf. ovata. Samples were collected in a freshwater seepage area of Catalina Bay in October 2009. Salinity ranged from 1 to 10. The water level at South Lake was once again low, thereby allowing the formation of fresh and brackish water ponds along the eastern shores.
Figure 4. Gastropods, food sources and diets in Catalina Bay, 2010.

(A) Four-point 10 m transect showing gastropod densities and microphytobenthic (MPB) biomass (as chl-a concentration). (B) Carbon and nitrogen stable isotope signatures of gastropods (number in brackets: trophic position) and their potential food sources, such as detritus (DTR) and sedimentary organic matter (SOM). SIAR boxplots show the proportional contribution (%) of different sources to the diet of (C) Tarebia granifera (shell height ≈13 mm), (D) T. granifera juveniles (shell height ≤5 mm) and (E) Assiminea cf. ovata. Samples were collected in a freshwater seepage area of Catalina Bay in February 2010. Salinity was 0.15–0.8. Although the salinity of the South Lake ranged between 47 and 56, its water level continued to decrease and larger freshwater ponds were formed in seepage areas along the eastern shores.
Figure 5. Gastropods, food types and diets in Lake Sibaya, 2009.

(A) Four-point 3 m transect showing gastropod densities and microphytobenthic (MPB) biomass (as chl-a concentration). (B) Carbon and nitrogen stable isotope signatures of gastropods (number in brackets: trophic position) and their potential food sources, such as detritus (DTR) and sedimentary organic matter (SOM). SIAR boxplots show the proportional contribution (%) of different sources to the diet of (C) Tarebia granifera and (D) Bulinus natalensis. Samples were collected in November 2009. This lake has no connection to the sea and contains pure freshwater.
Figure 6. Gastropods, food sources and diets in Lake Nhlange, 2008.

(A) Four-point 3 m transect showing gastropod densities and microphytobenthic (MPB) biomass (as chl-a concentration). (B) Carbon and nitrogen stable isotope signatures of gastropods (number in brackets: trophic position) and their potential food sources, such as detritus (DTR) and sedimentary organic matter (SOM). SIAR boxplots show the proportional contribution (%) of different sources to the diet of (C) Tarebia granifera and (D) Melanoides tuberculata. Samples were collected at Lake Nhlange in Kosi Bay, in November 2008. At the time, the site was dominated by freshwater conditions.
Gastropod densities as well as available microphytobenthic (MPB) biomass varied within transects (Fig. 2A– 6A; Table S1).Generally, native gastropods had lower population densities when compared to T. granifera (Table S1). However, Assiminea cf. ovata was found in densities greater than T. granifera at Catalina Bay in 2009 (Table S1). MPB biomass was available as a food source to gastropods at all sites, although its biomass was variable (Table S1). MPB biomass was not significantly correlated with gastropod densities (Table 1), despite being positively correlated with gastropod MPB diets (Table 1). Native gastropod MPB diet was strongly positively correlated with T. granifera MPB diet; however, it was negatively correlated with T. granifera density (Table 1).
Table 1. Correlations between microphytobenthic biomass and gastropod densities and diets.
| MPB | T. granifera density | Native density | T. granifera MPB diet | Native MPB diet | ||
| MPB | r | 1 | −0.211 | −0.021 | 0.615 | 0.49 |
| p | ns | ns | <0.001 | <0.05 | ||
| T. granifera density | r | −0.211 | 1 | −0.232 | 0.036 | −0.330 |
| p | ns | ns | ns | <0.05 | ||
| Native density | r | −0.021 | −0.232 | 1 | −0.114 | 0.354 |
| p | ns | ns | ns | <0.05 | ||
| T. granifera MPB diet | r | 0.615 | 0.036 | −0.114 | 1 | 0.649 |
| p | <0.001 | ns | ns | <0.001 | ||
| Native MPB diet | r | 0.409 | −0.330 | 0.354 | 0.649 | 1 |
| p | <0.05 | <0.05 | <0.05 | <0.001 |
Pearson correlations (r is the correlation coefficient; n = 60 for every case) between microphytobenthic (MPB) biomass, Tarebia granifera, native gastropod densities and corresponding contribution of MPB to their diets. Diets were estimated by a stable isotope mixing model. Samples were collected at three coastal lakes in Maputaland. Native gastropods were Haminoea natalensis, Assiminea cf. ovata, Bulinus natalensis and Melanoides tuberculata.
Gastropod diets
Gut content analysis revealed that microalgae and detritus were ingested by all gastropods (Table 2). Filamentous algae were also found in the guts of most gastropods (Table 2): their occurrence was 89.17% in T. granifera and 92% in native gastropods. Sand particles were only found in the guts of H. natalensis, B. natalensis, and in smaller amounts in T. granifera adults (shell height ≥10 mm) and M. tuberculata (Table 2). Distinctive remains of meiofauna were not found in gut contents. Macrophytes and fringing vegetation such as reeds, sedges and grasses, were excluded as potential food sources as their presence was not detected in the gut.
Table 2. Gastropod gut content analyses in three coastal lakes of Maputaland.
| Gut contents | |||||
| Location and year | Species | Microalgae | Detritus | Filamentous algae | Sand particles |
| CB 2007 | Tarebia granifera | 100 | 100 | 90 | 25 |
| Haminoea natalensis | 100 | 100 | 100 | 90 | |
| CB 2009 | T. granifera | 100 | 100 | 75 | 15 |
| Assiminea cf. ovata | 100 | 100 | 90 | 0 | |
| CB 2010 | T. granifera | 100 | 100 | 100 | 30 |
| T. granifera juveniles | 100 | 100 | 100 | 0 | |
| A. cf. ovata | 100 | 100 | 100 | 0 | |
| LS 2009 | T. granifera | 100 | 100 | 80 | 5 |
| Bulinus natalensis | 100 | 100 | 85 | 100 | |
| LN 2008 | T. granifera | 100 | 100 | 90 | 10 |
| Melanoides tuberculata | 100 | 100 | 85 | 15 | |
Gut contents were classified based on gross morphology. Data are presented as percentage occurrence in the guts of gastropods (n = 20). Samples were collected at Catalina Bay (CB) in the St. Lucia Estuary, Lake Sibaya (LS) and Lake Nhlange (LN) in Kosi Lakes.
Statistical analyses revealed that T. granifera δ13C and δ15N signatures differed significantly between locations and between times at Catalina Bay (Table 3, Fig. 2B– 6B). The δ13C signature of food source types also differed significantly between locations and between times (Table 4, Fig. 2B– 6B). The δ15N signature of food source types did not differ significantly between locations (Table 4), although it differed significantly over time at Catalina Bay (Table 4). There was no significant difference between the trophic position of T. granifera and that of native gastropods (t-test, df = 8, t = 0.406, p>0.05).
Table 3. Statistical analyses of stable isotope signatures of Tarebia granifera.
| Dependant variable | Source | df | MS | F | p |
| δ13C | Location | 2 | 136.126 | 118.7 | <0.001 |
| Error | 22 | 1.147 | |||
| Time | 2 | 4.324 | 34.538 | <0.001 | |
| Error | 12 | 0.125 | |||
| δ15N | Location | 2 | 4.052 | 5.07 | <0.05 |
| Error | 22 | 0.799 | |||
| Time | 2 | 5.074 | 44.337 | <0.001 | |
| Error | 12 | 0.114 |
ANOVA for δ13C and δ15N of T. granifera from Catalina Bay, Lake Sibaya and Lake Nhlange (different locations in Maputaland), and from 2007, 2009 and 2010 at Catalina Bay (different times).
Table 4. Statistical analyses of stable isotope signatures of gastropod food source types.
| Dependant variable | Source | df | MS | F | p |
| δ13C | Food×Location | 6 | 17.079 | 4.381 | <0.05 |
| Error | 48 | 3.899 | |||
| Food×Time | 6 | 16.138 | 5.519 | <0.05 | |
| Error | 24 | 2.924 | |||
| δ15N | Food×Location | 6 | 4.659 | 1.477 | ns |
| Error | 48 | 3.155 | |||
| Food×Time | 6 | 8.881 | 6.356 | <0.001 | |
| Error | 24 | 1.397 |
Two-way ANOVA for δ13C and δ15N, including microphytobenthos, detritus, Cladophora sp. and sedimentary organic matter (different food source types) collected at Catalina Bay, Lake Sibaya and Lake Nhlange (different locations in Maputaland), and collected in 2007, 2009 and 2010 at Catalina Bay (different times).
Stable isotope mixing model results indicated the largest proportional contributions of food sources to the diets of gastropods (Fig. 2C– 6C, Table S1). At Catalina Bay, in 2007 microphytobenthos (MPB) made the greatest contribution to the diet of T. granifera (13–74% of total); in 2009 it was MPB (2–53%), detritus (4–46%) and sedimentary organic matter (SOM) (0–47%); and in 2010 Cladophora sp. (0–61%) and SOM (0–58%). At Lake Sibaya, MPB (4–54%) and SOM (1–55%) made the greatest contribution to the diet of T. granifera; but at Lake Nhlange it was MPB (0–57%) and detritus (0–505) (Fig. 2C– 6C, Table S1). Cladophora sp. (43–93%) made the greatest contribution to the diet of T. granifera juveniles at Catalina Bay in 2010 (Fig. 4D, Table S1 ). Haminoea natalensis fed on MPB (29–95%) (Fig. 2D, Table S1). Assiminea cf. ovata fed on MPB in 2009 (25–93%) and Cladophora sp. in 2010 (53–98%) (Fig. 3D, Fig. 4E, Table S1). At Lake Sibaya, the native B. natalensis appeared to feed on a combination of all available food sources (Fig. 5D, Table S1). At Lake Nhlange, M. tuberculata diet consisted for the most part of MPB (0–55%), detritus (DTR) (0–50%) and SOM (4–63%) (Fig. 6D, Table S1).
Assuming that the microalgae found in the gut content analysis correspond to the mixing model MPB food source, detritus to DTR and filamentous algae to Cladophora sp.; there was a significant correlation between gut contents and diet estimated from stable isotope mixing models (n = 31, r2 = 0.69, p = 0.139).
Isotopic (stable isotope) dietary overlap (IDO) was significant (over 60%) between T. granifera and native gastropods in most cases (Table 5). The IDO between T. granifera adults and A. cf. ovata was not significant (below 60%) at Catalina Bay in 2010 (Table 5). The IDO between T. granifera adults and juveniles was also not significant (Table 5). However, the IDO of 95% between T. granifera juveniles (shell height ≤5 mm) and A. cf. ovata was the highest recorded in the study (Table 5).
Table 5. Isotopic (stable isotopes) dietary overlap between gastropods in three coastal lakes of Maputaland.
| Gastropod species | Location and year | Salinity | IDO (%) |
| Tarebia granifera a and Haminoea natalensis | CB 2007 | 16–32 | 75 |
| T. granifera and Assiminea cf. ovata | CB 2009 | 1–10 | 62 |
| T. granifera and A. cf. ovata | CB 2010 | 0.15–0.8 | 50 |
| T. granifera juvenilesa and A. cf. ovata | CB 2010 | 0.15–0.8 | 95 |
| T. granifera and T. granifera juveniles | CB 2010 | 0.15–0.8 | 49 |
| T. granifera and Bulinus natalensis | LS 2009 | 0.18–0.45 | 87 |
| T. granifera and Melanoides tuberculata | LN 2008 | 0.91–1.45 | 85 |
T. granifera = shell height ≥10 mm; T. granifera juveniles = shell height ≤5 mm; all other gastropods were adults.
Locations are Catalina Bay (CB) in the St. Lucia Estuary, Lake Sibaya (LS) and Lake Nhlange (LN) in Kosi Lakes. Isotopic dietary overlap (IDO, %) values over 60% indicate significant overlap and a value of 100% indicates absolute overlap.
Discussion
According to the competitive exclusion principle, species with identical ecological niche compete with each other for resources and, if these are limiting, there is a tendency towards the exclusion of the weaker competitor or the development of resource partitioning or niche differentiation [59]–[61]. Although recent research indicates that competition involving invasive species only rarely causes local extinction [62], there is still a need to better understand the mechanisms of competitive interactions and how the ecosystem is affected.
This study focuses only on situations where spatial overlap existed between the invasive gastropod Tarebia granifera and another single native gastropod species. Yet the spatial distribution of T. granifera can simultaneously overlap with that of several species at a time [26]. However, this study does not address the greater complexity of interactions between multiple gastropod species in the same area.
Tarebia granifera invaded the shallow waters (less than 2 m depth) of Kosi Bay's Lake Nhlange, Lake Sibaya and the eastern shores of the South Lake of the St. Lucia Estuary (Fig. 1). The T. granifera populations tend to form dense patches (often >1000 ind. m−2) of varying dimensions. Tarebia granifera patches such as the ones sampled in this study are often found in very shallow embayments with clear sandy substrate where particulate organic matter settles or where there is freshwater seepage. In contrast, the native species in this study have much lower population densities and their distribution is more extensive and much less patchy [26]. E.g. Assiminea cf. natalensis at the St. Lucia Estuary is a historically dominant species found throughout the system [35]. Tarebia granifera also appears to be displacing native gastropods from certain areas in the iSimangaliso Wetland Park (Fig. 1), whereas the native species do not appear to be mutually exclusive.
Tarebia granifera has previously been classified as both detritivore and grazer [31], [63]. This study shows that T. granifera has a generalist diet which includes a wide variety of food sources. Tarebia granifera is established in all major coastal lakes of Maputaland (Fig. 1), where in particular cases its diet was either largely composed of a single or a combination of general food sources (Fig. 2C– 6C, Table S1). Native gastropods A. cf. ovata and Haminoea natalensis tended to favor a single food source in Catalina Bay, whereas Bulinus natalensis and Melanoides tuberculata had a more varied diet in Lake Sibaya and Lake Nhlange respectively. Microphytobenthos (MPB) featured in the diet of all gastropods and its proportional contribution was always positively correlated with its available biomass in the environment (Table 1). Although T. granifera in high densities had the potential to consume MPB until it became limiting [31], there were other food sources available to gastropods, such as detritus (DTR), the filamentous algae Cladophora sp. and sedimentary organic matter (SOM) (Table S1).
Autochthonous primary production (represented by MPB, Cladophora sp. and SOM) contributed on average 78% of gastropod diets. δ13C signatures were significantly more enriched at Catalina Bay, when compared with Lake Nhlange and Lake Sibaya, which is expected because of the influence of salt water at that site [64]. Detritus (DTR) is the only food source that could represent an allochthonous contribution in the form of terrestrial plant matter, and it only contributed on average up to 20% of gastropod diets. Although terrestrial primary production can support secondary production of aquatic mollusks (since most species, including T. granifera, have the ability to feed on [65] and digest terrestrial plant matter via cellulase activity [66]), aquatic primary production was more important for the gastropods at the coastal lakes in this study. Animal contributions, in the form of decaying matter which would not be easily recognized in a gut content analysis, can also be represented in the DTR signature which would explain its wide variation in δ13C. Decaying animal matter and/or meiofaunal organisms can make a small but significant contribution to the diet of T. granifera. Melanoides tuberculata, which in this study exhibited a diet identical to that of T. granifera, is capable of assimilating animal matter [53]. This opens the possibility that T. granifera may also feed on dead gastropods, which would be beneficial to its survival at very high densities and under unfavorable environmental conditions. Detritus and sedimentary organic matter may account for the high organic carbon content (up to 60%) not accounted for by the gut fluorescence technique in T. granifera [31]. However, although as many samples as possible were collected and broadly classified, due to logistical limitations not all potential food sources may have been considered in this study.
Assiminea cf. ovata has an affinity for MPB, as do other assimineid species [67]. However, the increase in abundance of T. granifera at St Lucia from 2009 to 2010 may have influenced a dietary shift in A. cf. ovata, which started to feed less on MPB and more on Cladophora sp. (Fig. 3D, 4E and Table S1). This is not surprising, as dietary shifts in native species tend to be caused by non-native species, as they disrupt trophic connections [1]. If resource partitioning between T. granifera and A. cf. ovata took place, it was effective in reducing IDO from 62% in 2009 to 50% in 2010. However, it is very likely that dietary shifts occured in response to changing environmental conditions, which affected food availability and quality at Catalina Bay.
Due to its remarkable salinity tolerance, T. granifera managed to survive the unusual 2007 mouth breaching event when seawater entered the St. Lucia Estuary and flooded the freshwater seepage areas of Catalina Bay (Fig. 1) [30]. Assiminea cf. ovata was abundant in most of the seepage areas only in 2008, while the salinity level was high (around 30) [26]. When fresh water conditions became re-established in the seepage areas in 2009, T. granifera increased in numbers and spread until 2010, when it once again dominated benthic assemblages whilst evidently displacing A. cf. ovata [26]. Along the eastern shores of South Lake (Fig. 1), A. cf. ovata continued to be abundant in areas with high salinity and was even found in a few freshwater seepage areas where T. granifera was not present (e.g. Dead Tree Bay: 28°9′2.83″S, 32°27′53.76″E). At least over the last six years, A. cf. ovata and H. natalensis have consistently been found together in shallow-water habitats on the western shores of South Lake (Fig. 1), where there are fewer freshwater seepage areas, with only one being found to harbor T. granifera, i.e. Makakatana Bay (28°14′10.87″S, 32°25′11.61″E). In contrast to T. granifera, H. natalensis does not appear to displace A. c.f. ovata.
Salinity is indeed one of the most important factors influencing the distribution of gastropods in coastal lakes of Maputaland [26]. Although there was a significant IDO between T. granifera and H. natalensis, there was very little spatial overlap between the two, since H. natalensis preferred the saltier conditions outside the seepage area. High salinity conditions were also associated with A. cf. ovata [26]. When salinity dropped below 20, T. granifera expanded its range and density. Assiminea cf. ovata was recorded in the St. Lucia Estuary at low salinities, but only when T. granifera was either absent or present in relatively low densities. This suggests that T. granifera may displace A. cf. ovata only under specific salinity conditions. This also supports the theory that vulnerable native species can resist invasion by non-native species under certain conditions [68].
Interestingly, at Catalina Bay in 2010, it was observed that both A. cf. ovata and T. granifera juveniles tended to concentrate in extremely shallow areas (depth <0.1 metre), whereas T. granifera adults tended to concentrate in adjacent deeper waters (depth >0.1 metre). In this environment, where water level fluctuation is commonplace, it is conceiveable that smaller snails would continue to feed in extremely shallow areas, whereas larger snails tend to move away and/or burrow to avoid dessication. This behavior reduces the potential for competitive interactions involving T. granifera adults and can be interpreted in terms of spatial niche differentiation. However, the spatial overlap between A. cf. ovata and T. granifera juveniles remains unchanged and becomes effectively greater than that between A. cf. ovata and T. granifera adults. Tarebia granifera juveniles are often numerous, have fast growth rates and may even grow faster and larger in the presence of other snails [24], [31], [69]. They are voracious feeders and may have greater impacts on food stocks than adults [31], [70]. It is therefore suggested that T. granifera juveniles played a role in the displacement of A. cf. ovata. A high degree of spatial overlap between T. granifera adults and juveniles has been recorded [26], however, there is also evidence of an ontogenetic dietary shift in T. granifera, since SOM was only used by adults and IDO was not significant (Table 5, Table S1, Fig. 4C–D). Resource partitioning certainly minimizes potential competition between T. granifera adults and juveniles.
Food resource use within populations at the microhabitat scale can be a significant factor in gastropod interactions [71]. Some of the large variations in isotopic signatures recorded for gastropods in this study may be due to individual variation, which in turn may be primarily affected by habitat heterogeneity and resource availability [71]. This adds to the complexity of addressing dietary overlap and potential competition for food resources between gastropods.
Tarebia granifera and B. natalensis appear to have very similar and overlapping diets (Table S1, Table 5). Thiarids and sympatric pulmonates can have similar diets [72]. The lack of competitive exclusion may be explained by selective ingestion and behavioral differences [73], as well as selection of different micro-habitats, such as macrophyte fronds where B. natalensis positions itself close to the water surface [74], thus avoiding physical interactions with T. granifera which tend to stay on the substrate. However, in the Caribbean, T. granifera is known to displace Biomphalaria glabrata (Say, 1818) (pulmonate, Planorbidae), which like B. natalensis is associated with submerged vegetation rather than the substrate. Further empirical studies are thus needed to address interactions between T. granifera and B. natalensis which may involve chemical cues rather than competition for food [27], [75].
Tarebia granifera has been widely reported to out-compete M. tuberculata for food resources and space and there is concern that other native thiarids, such as Thiara amarula (Linnaeus, 1758) may also be negatively affected [24]. As expected, the diets of these gastropods are similar and IDO is significant (Table 5, Table S1), suggesting that T. granifera and M. tuberculata can compete for resources. Yet, there is spatial overlap between these species in Lake Nhlange. This apparent competitive co-existence [76] may be explained by the generalist diet of both species together with adequate availability of resources.
At Catalina Bay and Lake Nhlange, there was no indication that food resources were limiting. The lowest available MPB biomass was recorded on the eastern shores of Lake Sibaya in a sheltered bay (table S1). A larger proportion of the eastern shore of Lake Sibaya is comprised of exposed sandy terraces that are even poorer in nutrients in comparison to sheltered bays, which tend to accumulate organic deposits [77]. Tarebia granifera is currently the only recorded gastropod species dominating the benthic assemblage in open terraces of Lake Sibaya [26]. The unusually small size of the T. granifera individuals at Lake Sibaya's open terraces may be explained by lower food availability and quality [26]. This suggests that food resources are limiting in large areas of Lake Sibaya and this may have played a role in the apparent displacement of the historically abundant native gastropods M. tuberculata and Bellamya capillata (prosobranch, Viviparidae) [34], [78], by T. granifera. However, both M. tuberculata and B. capillata may still persist in parts of Lake Sibaya that have not been surveyed and particularly at depths greater that 5 m where less T. granifera can be found.
In the St. Lucia Estuary, there appears to be a relatively high concentration of dissolved nutrients despite the drought conditions [79], [80]. Nutrient input is likely to come from fecal matter of ungulates, waterfowl and particularly hippopotami that frequent freshwater seepage areas and play a part in shaping them [81], [82]. Nutrient input and concentrations are also affected by water level fluctuations which can be extremely variable at Catalina Bay. This may explain the δ15N signature variation and enrichment observed at Catalina Bay in 2010. Nutrient enrichment certainly led to the increase in abundance of Cladophora sp., which featured prominently in gastropod diets at Catalina Bay only in 2010. Nutrient enrichment can have overriding effects on several system processes [83], initially favoring both native and non-native species. However, T. granifera appear to have a greater and faster growth per unit resource consumption, coupled with earlier onset of reproduction, which contributes to their invasion success [84], [85].
Although in the same trophic level, and thus probably performing a similar ecosystem function, T. granifera differs from the native benthic invertebrates of Maputaland in many ways. Besides having different physiology, behavior and life history, T. granifera can also differ from other species in terms of its elemental turnover rate [86] and biomass accrual. Tarebia granifera populations attain extremely high densities due to their high parthenogenetic reproduction rate. Large amounts of carbon are therefore sequestered by T. granifera. The average size of individuals within gastropod species is naturally a factor in determining the biomass of the different populations. When compared to native gastropod species, such as B. capillata at Lake Sibaya, the estimated per capita biomass of T. granifera can be on average ten times lower (unpublished data). At Catalina Bay, the average per capita biomass of T. granifera is about double that of A. cf. ovata (unpublished data). However, the biomass of T. granifera populations is often much greater than that of potentially vulnerable native gastropods in the same trophic level. Therefore, the invasion of sites such as Catalina Bay by T. granifera results in the sequestration of a lot more carbon than usual in benthic primary consumers. This might not be an issue in terms of reducing the availability of nutrients to native gastropods, since there seem to be a great abundance of nutrient resources at Catalina Bay. At sites like Lake Sibaya, the invasion of T. granifera may not result in the sequestration of such high magnitudes of carbon, but because nutrients can be limiting, T. granifera can at least affect their availability to native gastropods and interfere with vital nutrient cycling dynamics in complex ways [31], [87]–[89].This study did not focus on higher trophic level interactions. Indeed, there is a general lack of multiple trophic level studies that may reveal more complex ecological responses [90]. However, snails like T. granifera do not seem to be successfully predated upon by native species in iSimangaliso, although they can be attacked by birds, fish and invertebrates, such as crabs and leeches. Trematode parasites are also absent in T. granifera [26]. Predators and parasites could regulate populations of alien gastropods [45]. The lack of successful predation and parasitism is thus likely to facilitate the establishment and spread of T. granifera in South Africa.
Supporting Information
Available microphytobenthos biomass and gastropod densities and diets in three coastal lakes of Maputaland. The contributions of microphytobenthos (MPB), detritus (DTR), Cladophora sp. and sedimentary organic matter (SOM) to the diets of gastropods were calculated with the SIAR (stable isotope analysis in R) mixing model using Carbon (δ13C) and Nitrogen (δ15N) signatures. Gastropod shell height (range) was recorded. Available MPB biomass is presented as pigment concentration.
(DOC)
Acknowledgments
We are grateful to the iSimangaliso Park Authority and Ezemvelo KZN Wildlife for supporting this project. Special thanks go to R. Taylor, C. Fox and R. Kyle for their invaluable assistance and logistical support.
Footnotes
Competing Interests: The authors have declared that no competing interests exist.
Funding: Funding was provided by the National Research Foundation, South Africa, Marine and Coastal Management (DEAT-MCM) and the World Wide Fund for Nature (WWF). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Vander Zanden MJ, Casselman JM, Rasmussen JB. Stable isotope evidence for the food web consequences of species invasions in lakes. Nature. 1999;401:464–467. [Google Scholar]
- 2.Lockwood JL, Hoopes MF, Marchetti MP. Invasion Ecology. Malden, U.S.A.: Blackwell Publishing; 2007. [Google Scholar]
- 3.Dawson TE, Siegwolf RTW. Stable isotopes as indicators of ecological change. U.S.A.: Elsevier; 2007. [Google Scholar]
- 4.Post DM. Using stable isotopes to estimate trophic position: models, methos, and assumptions. Ecology. 2002;83:703–718. [Google Scholar]
- 5.Caut S, Roemer GW, Donlan CJ, Courchamp F. Coupling stable isotopes with bioenergetics to estimate interspecific interactions. Ecol Appl. 2006;16:1893–1900. doi: 10.1890/1051-0761(2006)016[1893:csiwbt]2.0.co;2. [DOI] [PubMed] [Google Scholar]
- 6.Gearing JN. The study of diet and trophic relationships through natural abundance 13C. In: Coleman DC, Fry B, editors. San Diego, California: Academic Press, Inc; 1991. [Google Scholar]
- 7.Peterson BJ. Stable isotopes as tracers of organic matter input and transfer in benthic food webs: A review. Acta Oecol. 1999;20:479–487. [Google Scholar]
- 8.Clarke LR, Vidergar DT, Bennett DH. Stable isotopes and gut content show diet overlap among native and introduced piscivores in a large oligotrophic lake. Ecol Freshw Fish. 2005;14:267–277. [Google Scholar]
- 9.Rudnick D, Resh V. Stable isotopes, mesocosms and gut content analysis demonstrate trophic differences in two invasive decapod crustacea. Freshwater Biol. 2005;50:1323–1336. [Google Scholar]
- 10.Caut S, Angulo E, Courchamp F. Dietary shift of an invasive predator: rats, seabirds and sea turtles. J Appl Ecol. 2008;45:428–437. doi: 10.1111/j.1365-2664.2007.01438.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zambrano L, Valiente E, Vander Zanden M. Food web overlap among native axolotl (Ambystoma mexicanum) and two exotic fishes: carp (Cyprinus carpio) and tilapia (Oreochromis niloticus) in Xochimilco, Mexico City. Biol Invasions. 2010;12:3061–3069. [Google Scholar]
- 12.Zambrano L, Valiente E, Zanden MJV. Stable isotope variation of a highly heterogeneous shallow freshwater system. Hydrobiologia. 2010;646:327–336. [Google Scholar]
- 13.Carlsson NOL. Invasive golden apple snails are threatening natural ecosystems in Southeast Asia. In: Joshi RC, Sebastian LS, editors. Global Advances in Ecology and Management of Golden Apple Snails. Nueva Ecija: Philippines Rice Research Institute; 2006. pp. 61–72. [Google Scholar]
- 14.Fang L, Wong PK, Lin L, Lan C, Qiu JW. Impact of invasive apple snails in Hong Kong on wetland macrophytes, nutrients, phytoplankton and filamentous algae. Freshwater Biol. 2010;55:1191–1204. [Google Scholar]
- 15.Wong P, Kwong K, Qiu J-W. Complex interactions among fish, snails and macrophytes: implications for biological control of an invasive snail. Biol Invasions. 2009;11:2223–2232. [Google Scholar]
- 16.Schreiber ESG, Lake PS, Quinn GP. Facilitation of native stream fauna by an invading species? Experimental investigations of the interaction of the snail, Potamopyrgus antipodarum (Hydrobiidae) with native benthic fauna. Biol Invasions. 2002;4:317–325. [Google Scholar]
- 17.Kerans BL, Dybdahl MF, Gangloff MM, Jannot JE. Potamopyrgus antipodarum: distribution, density, and effects on native macroinvertebrate assemblages in the Greater Yellowstone Ecosystem. J N Am Benthol Soc. 2005;24:123–138. [Google Scholar]
- 18.Aberle N, Hillebrand H, Grey J, Wiltshire KH. Selectivity and competitive interactions between two benthic invertebrate grazers (Asellus aquaticus and Potamopyrgus antipodarum): an experimental study using 13C- and 15N-labelled diatoms. Freshwater Biol. 2005;50:369–379. [Google Scholar]
- 19.Brenneis VEF, Sih A, De Rivera CE. Coexistence in the intertidal: interactions between the non-indigenous New Zealand mud snail Potamopyrgus antipodarum and the native estuarine isopod Gnorimosphaeroma insulare. Oikos. 2010;119:1755–1764. [Google Scholar]
- 20.Abbott RT. A study of an intermediate snail host (Thiara granifera) of the oriental lung fluke (Paragonimus). Proc U S Nat Mus. 1952;102:72–116. [Google Scholar]
- 21.Chaniotis BN, Butler JM, Ferguson F, Jobin WR. Bionomics of Tarebia granifera (Gastropoda:Thiaridae) in Puerto Rico, an Asian vector of Paragonimiasis westermani. Caribb J Sci. 1980;16:81–89. [Google Scholar]
- 22.Pointer JP, Facon B, Jarne P, David P. Thiarid snails, invading gastropods of tropical freshwater habitats. Xenophora. 2003;104:14–20. [Google Scholar]
- 23.Ben-Ami F. First report of the invasive freshwater snail Tarebia granifera (Lamarck, 1816) (Gastropoda: Thiaridae) from Israel. Nautilus. 2006;120:156–161. [Google Scholar]
- 24.Appleton CC, Forbes AT, Demetriades NT. The occurrence, bionomics and potential impacts of the invasive freshwater snail Tarebia granifera (Lamarck, 1822) (Gastropoda: Thiaridae) in South Africa. Zool Med Leiden. 2009;83:525–536. [Google Scholar]
- 25.Appleton CC. Alien and invasive freshwater gastropoda in South Africa. Afr J Aquat Sci. 2003;28:69–81. [Google Scholar]
- 26.Miranda NAF, Perissinotto R, Appleton CC. Population structure of an invasive parthenogenetic gastropod in coastal lakes and estuaries of Northern KwaZulu-Natal, South Africa. PLoS ONE. 2011;6:e24337. doi: 10.1371/journal.pone.0024337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Perez JG, Vargas M, Malek EA. Displacement of Biomphalaria glabrata by Thiara granifera under Natural Conditions in the Dominican-Republic. Mem I Oswaldo Cruz. 1991;86:341–347. doi: 10.1590/s0074-02761991000300008. [DOI] [PubMed] [Google Scholar]
- 28.Pointier J. Invading freshwater gastropods: some conflicting aspects for public health. Malacologia. 1999;41:403–411. [Google Scholar]
- 29.Pointier JP, Jourdane J. Biological control of the snail hosts of schistosomiasis in areas of low transmission: the example of the Caribbean area. Acta Tropica. 2000;77:53–60. doi: 10.1016/s0001-706x(00)00123-6. [DOI] [PubMed] [Google Scholar]
- 30.Miranda NAF, Perissinotto R, Appleton CC. Salinity and temperature tolerance of the invasive freshwater gastropod Tarebia granifera. S Afr J Sci. 2010;106:55–61. [Google Scholar]
- 31.Miranda NAF, Perissinotto R, Appleton CC. Feeding dynamics of the invasive gastropod Tarebia granifera in coastal and estuarine lakes of northern KwaZulu Natal, South Africa. Estuarine Coastal Shelf Sci. 2011;91:442–449. [Google Scholar]
- 32.Taylor R, Adams JB, Haldorsen S. Primary habitats of the St Lucia Estuarine System, South Africa, and their responses to mouth management. Afr J Aquat Sci. 2006;31:31–41. [Google Scholar]
- 33.Bate GC, Taylor RH. Sediment salt-load in the St Lucia Estuary during the severe drought of 2002–2006. Environ Geol. 2007;55:1089–1098. [Google Scholar]
- 34.Appleton CC. Mollusca. In: Day JH, de Moor I, editors. Guides to the freshwater invertebrtes of Southern Africa. Gezina: Water Research Commission; 2002. pp. 42–141. [Google Scholar]
- 35.Boltt RE. The Benthos of Some Southern African Lakes Part V: The Recovery of the Benthic Fauna of St Lucia Lake Following a Period of Excessively High Salinity. Trans R Soc S Afr. 1975;41:295–323. [Google Scholar]
- 36.MacKay F, Cyrus D, Russell KL. Macrobenthic invertebrate responses to prolonged drought in South Africa's largest estuarine lake complex. Estuarine Coastal Shelf Sci. 2010;86:553–567. [Google Scholar]
- 37.Macnae W. Tectibranch Molluscs from Southern Africa. Ann Natal Mus. 1962;15:183–199. [Google Scholar]
- 38.Bruton MN. An outline of the ecology of Lake Sibaya, with emphasis on the vertebrate community. In: Bruton MN, Cooper KH, editors. Studies on the ecology of Maputaland. Grahamstown: Rhodes University; 1980. pp. 382–407. [Google Scholar]
- 39.Bruton MN. Lake Sibaya - a land-locked estuary. Scientiae. 1975;16:18–28. [Google Scholar]
- 40.Allanson BR. The physico-chemical limnology of Lake Sibaya. In: Allanson BR, editor. Lake Sibaya. Hague: Dr. W. Junk Publishers; 1979. pp. 42–74. [Google Scholar]
- 41.de Kock KN, Wolmarans CT. Distribution and habitats of Bulinus natalensis and its role as intermediate host of economically important helminth parasites in South Africa. Afri J Aquat Sci. 2006;31:63–69. [Google Scholar]
- 42.Begg GW. The Kosi system: aspects of its biology, management and research. In: Bruton MN, Cooper KH, editors. Studies on the ecology of Maputaland. Grahamstown: Rhodes University; 1980. [Google Scholar]
- 43.Genner MJ, Michel E, Erpenbeck D, De Voogd N, Witte F, et al. Camouflaged invasion of Lake Malawi by an Oriental gastropod. Mol Ecol. 2004;13:2135–2141. doi: 10.1111/j.1365-294X.2004.02222.x. [DOI] [PubMed] [Google Scholar]
- 44.Genner MJ, Todd JA, Michel E, Erpenbeck D, Jimoh A, et al. Amassing diversity in an ancient lake: evolution of a morphologically diverse parthenogenetic gastropod assemblage in Lake Malawi. Mol Ecol. 2007;16:517–530. doi: 10.1111/j.1365-294X.2006.03171.x. [DOI] [PubMed] [Google Scholar]
- 45.Genner MJ, Michel E, Todd JA. Resistance of an invasive gastropod to an indigenous trematode parasite in Lake Malawi. Biol Invasions. 2007;10:41–49. [Google Scholar]
- 46.Welschmeyer NA. Fluorometric analysis of chlorophyll a in the presence of chlorophyll b and pheopigments. Lymnol Oceanogr. 1994;39:1985–1992. [Google Scholar]
- 47.Strickland JDH, Parsons TR. A practical handbook for sewater analysis. Bulletin Fisheries Research Board of Canada. 1968;167:1–311. [Google Scholar]
- 48.Malaquias MAE, Condinho S, Cervera JL, Sprung M. Diet and feeding biology of Haminoea orbygniana (Mollusca: Gastropoda: Cephalaspidea). J Mar Biol Assoc U K. 2004;84:767–772. [Google Scholar]
- 49.Alfaro AC. Diet of Littoraria scabra, while vertically migrating on mangrove trees: gut content, fatty acid, and stable isotope analyses. Estuarine Coastal Shelf Sci. 2008;79:718–726. [Google Scholar]
- 50.Kiljunen M, Grey J, Sinisalo T, Harrod C, Immonen H, et al. A revised model for lipid-normalizing δ13C values from aquatic organisms, with implications for isotope mixing models. J Appl Ecol. 2006;43:1213–1222. [Google Scholar]
- 51.Post D, Layman C, Arrington D, Takimoto G, Quattrochi J, et al. Getting to the fat of the matter: models, methods and assumptions for dealing with lipids in stable isotope analyses. Oecologia. 2007;152:179–189. doi: 10.1007/s00442-006-0630-x. [DOI] [PubMed] [Google Scholar]
- 52.Parnell AC, Inger R, Bearhop S, Jackson AL. Source partitioning using stable isotopes: coping with too much variation. PLoS ONE. 2010;5:e9672. doi: 10.1371/journal.pone.0009672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Coat S, Monti D, Bouchon C, Lepoint G. Trophic relationships in a tropical stream food web assessed by stable isotope analysis. Freshwater Biol. 2009;54:1028–1041. [Google Scholar]
- 54.Ben-David M, Schell DM. Mixing models in analyses of diet using multiple stable isotopes: a response. Oecologia. 2001;127:180–184. doi: 10.1007/s004420000570. [DOI] [PubMed] [Google Scholar]
- 55.Phillips DL, Newsome SD, Gregg JW. Combining sources in stable isotope mixing models: alternative methods. Oecologia. 2005;144:520–527. doi: 10.1007/s00442-004-1816-8. [DOI] [PubMed] [Google Scholar]
- 56.Mantel SK, Salas M, Dudgeon D. Foodweb structure in a tropical Asian forest stream. J N Am Benthol Soc. 2004;23:728–755. [Google Scholar]
- 57.Schoener TW. Theory of feeding strategies. Annu Rev Ecol Syst. 1971;2:369–404. [Google Scholar]
- 58.Krebs CJ. Ecological methodology. New York: HarperCollins Publishers; 1989. [Google Scholar]
- 59.Hardin G. The competitive exclusion principle. Science. 1960;131:1292–1297. doi: 10.1126/science.131.3409.1292. [DOI] [PubMed] [Google Scholar]
- 60.Schoener TW. Resource partitioning in ecological communities. Science. 1974;185:27–39. doi: 10.1126/science.185.4145.27. [DOI] [PubMed] [Google Scholar]
- 61.Wiens JA. Fat times, lean times and competition among predators. Trends Ecol Evol. 1993;8:348–349. doi: 10.1016/0169-5347(93)90216-C. [DOI] [PubMed] [Google Scholar]
- 62.Sax DF, Stachowicz JJ, Brown JH, Bruno JF, Dawson MN, et al. Ecological and evolutionary insights from species invasions. Trends Ecol Evol. 2007;22:465–471. doi: 10.1016/j.tree.2007.06.009. [DOI] [PubMed] [Google Scholar]
- 63.Larned ST, Chong CT, Punewai N. Detrital fruit processing in a Hawaiian stream ecosystem. Biotropica. 2001;33:241–248. [Google Scholar]
- 64.Fry B, Sherr EB. 13C measurements as indicators of carbon flow in marine and freshwater ecosystems. Contrib Mar Sci. 1984;27:13–47. [Google Scholar]
- 65.Appleton CC. 2005. Invasive freshwater invertebrates in South Africa.
- 66.Antonio ES, Kasai A, Ueno M, Kurikawa Y, Tsuchiya K, et al. Consumption of terrestrial organic matter by estuarine molluscs determined by analysis of their stable isotopes and cellulase activity. Estuarine Coastal Shelf Sci. 2010;86:401–407. [Google Scholar]
- 67.Kurata K, Minami H, Kikuchi E. Stable isotope analysis of food sources for salt marsh snails. Mar Ecol Prog Ser. 2001;223:167–177. [Google Scholar]
- 68.Grosholz E. Ecological and evolutionary consequences of coastal invasions. Trends Ecol Evol. 2002;17:22–27. [Google Scholar]
- 69.Kawata M, Ishigami H. The growth of juvenile snails in water conditioned by snails of a different species. Oecologia. 1992;91:245–248. doi: 10.1007/BF00317791. [DOI] [PubMed] [Google Scholar]
- 70.Pape E, Muthumbi A, Kamanu CP, Vanreusel A. Size-dependent distribution and feeding habits of Terebralia palustris in mangrove habitats of Gazi Bay, Kenya. Estuarine Coastal Shelf Sci. 2008;76:797–808. [Google Scholar]
- 71.Doi H, Yurlova NI, Kikuchi E, Shikano S, Yadrenkina EN, et al. Stable isotopes indicate individual level trophic diversity in the freshwater gastropod Lymnaea stagnalis. J Mollus Stud. 2010;76:384–338. [Google Scholar]
- 72.Madsen H. Food selection by freshwater snails in the Gezira irrigation canals, Sudan. Hydrobiologia. 1992;228:203–217. [Google Scholar]
- 73.Barnese LE, Lowe RL, Hunter RD. Comparative grazing efficiency of pulmonate and prosobranch snails. J N Am Benthol Soc. 1990;9:35–44. [Google Scholar]
- 74.Seuffert ME, Martin PR. Dependence on aerial respiration and its influence on microdistribution in the invasive freshwater snail Pomacea canaliculata (Caenogastropoda, Ampullariidae). Biol Invasions. 2010;12:1695–1708. [Google Scholar]
- 75.Gomez JD, Vargas M, Malek EA. Biological control of Biomphalaria glabrata by Thiara granifera under Laboratory Conditions. Trop Med Parasitol. 1990;41:43–45. [PubMed] [Google Scholar]
- 76.Gabler H-M, Amundsen P-A. Feeding strategies, resource utilisation and potential mechanisms for competitive coexistence of Atlantic salmon and alpine bullhead in a sub-Arctic river. Aquat Ecol. 2010;44:325–336. [Google Scholar]
- 77.Bruton MN, Boltt RE. Aspects of the biology of Tilapia mossambica Peters (Pisces: Cichlidae) in a natural freshwater lake (Lake Sibaya, South Africa). J Fish Biol. 1975;7:423–445. [Google Scholar]
- 78.Hart RC. The invertebrate comunities: zooplankton, zoobenthos and littoral fauna. In: Allanson BR, editor. Lake Sibaya. Hague: Dr. W. Junk Publishers; 1979. pp. 108–161. [Google Scholar]
- 79.Humphries P, Baldwin DS. Drought and aquatic ecosystems: an introduction. Oxford, ROYAUME-UNI: Blackwell Science; 2003. pp. 1141–1146. [Google Scholar]
- 80.Perissinotto R, Pillay D, Bate G. Microalgal biomass in the St Lucia Estuary during the 2004 to 2007 drought period. Mar Ecol Prog Ser. 2010;405:147–161. [Google Scholar]
- 81.Kilham P. The effect of hippopotamuses on potassium and phosphate ion concentrations in an African lake. Am Midland Nat. 1982;108:202–205. [Google Scholar]
- 82.McCarthy TS, Ellery WN, Bloem A. Some observations on the geomorphological impact of hippopotamus (Hippopotamus amphibius L.) in the Okavango Delta, Botswana. Afr J Ecol. 1998;36:44–56. [Google Scholar]
- 83.Wojdak JM. Relative strength of top-down, bottom-up, and consumer species richness effects on pond ecosystems. Ecol Monogr. 2005;75:489–504. [Google Scholar]
- 84.Brown KM. Resource overlap and competition in pond snails: an experimental analysis. Ecology. 1982;63:412–422. [Google Scholar]
- 85.Winsome T, Epstein L, Hendrix PF, Horwath WR. Competitive interactions between native and exotic earthworm species as influenced by habitat quality in a California grassland. Appl Soil Ecol. 2006;32:38–53. [Google Scholar]
- 86.McIntyre PB, Flecker AS. Rapid turnover of tissue nitrogen of primary consumers in tropical freshwaters. Oecologia. 2006;148:12–21. doi: 10.1007/s00442-005-0354-3. [DOI] [PubMed] [Google Scholar]
- 87.Pillay D, Branch GM, Steyn A. Complex effects of the gastropod Assiminea globulus on benthic community structure in a marine-dominated lagoon. J Exp Mar Biol Ecol. 2009;380:47–52. [Google Scholar]
- 88.López-López E, Sedeño-Díaz JE, Vega PT, Oliveros E. Invasive mollusks Tarebia granifera Lamarck, 1822 and Corbicula fluminea Müller, 1774 in the Tuxpam and Tecolutla rivers, Mexico: spatial and seasonal distribution patterns. Aquat Invasions. 2009;4:435–450. [Google Scholar]
- 89.Reise K, Olenin S, Thieltges D. Are aliens threatening European aquatic coastal ecosystems? Helgoland Mar Res. 2006;60:77–83. [Google Scholar]
- 90.Hooper DU, Chapin FS, Ewel JJ, Hector A, Inchausti P, et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol Monogr. 2005;75:3–35. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Available microphytobenthos biomass and gastropod densities and diets in three coastal lakes of Maputaland. The contributions of microphytobenthos (MPB), detritus (DTR), Cladophora sp. and sedimentary organic matter (SOM) to the diets of gastropods were calculated with the SIAR (stable isotope analysis in R) mixing model using Carbon (δ13C) and Nitrogen (δ15N) signatures. Gastropod shell height (range) was recorded. Available MPB biomass is presented as pigment concentration.
(DOC)