

Abstract
Social networks affect in such a fundamental way the dynamics of the population they support that the global, population-wide behavior that one observes often bears no relation to the individual processes it stems from. Up to now, linking the global networked dynamics to such individual mechanisms has remained elusive. Here we study the evolution of cooperation in networked populations and let individuals interact via a 2-person Prisoner's Dilemma – a characteristic defection dominant social dilemma of cooperation. We show how homogeneous networks transform a Prisoner's Dilemma into a population-wide evolutionary dynamics that promotes the coexistence between cooperators and defectors, while heterogeneous networks promote their coordination. To this end, we define a dynamic variable that allows us to track the self-organization of cooperators when co-evolving with defectors in networked populations. Using the same variable, we show how the global dynamics — and effective dilemma — co-evolves with the motifs of cooperators in the population, the overall emergence of cooperation depending sensitively on this co-evolution.
Introduction
Dynamical processes involving populations of individuals constitute paradigmatic examples of complex systems. From epidemic outbreaks to opinion formation and behavioral dynamics [1]–[9], the impact of the underlying web of ties in the overall behavior of the population is well known. In this context, Evolutionary Games [10], [11] provide one of the most sophisticated examples of complex dynamics in which the role of the underlying network topology proves ubiquitous. For instance, when cooperation is modeled as a prisoner's dilemma game, cooperation may emerge (or not) depending on how the population is networked [12]–[37].
Up to now, it has been hard to characterize in detail the global dynamics by which local self-regarding actions lead to a collective cooperative scenario, relating it to the network topology. Indeed, most network studies have been focused on the analysis of the evolutionary outcome of cooperation [14] — either by means of the numerical analysis of steady states [12], [16], [18], [19], [24], [25], [29], [32], [34], [35], [38]–[40] or by means of the analytical determination of the conditions for fixation in the population or by means of the determination of positive inclusive fitness effects for particular homogeneous network interaction structures and low intensities of selection [17], [20], [22] — without characterizing the self-organization process by which one of the strategies outcompetes the other. Here we show how networked individuals, engaging in a prisoner's dilemma (PD) of cooperation, give rise to a global, population wide, behavioral dynamics which deviates strongly from the original PD, depending sensitively on the underlying network of contacts: Homogeneous networks promote a coexistence dynamics between cooperators and defectors — akin to the Chicken or Snowdrift game [11], [38], [41]–[43] — whereas heterogeneous networks, from broad scale to scale-free [4], [44], favor the coordination between them, similar to the Stag-hunt game [45].
To this end we define a time-dependent variable — that we call the average gradient of selection (AGoS) — and use it to track the self-organization of cooperators when co-evolving with defectors under network reciprocity. Similar to existing analytical approaches [43], [46], the AGoS is able to provide a measure of the change in time of the frequency of cooperative traits under selection. The AGoS can be computed for arbitrary intensity of selection (see Methods), arbitrary population structure and arbitrary game parameterization. We further prove that the global games are not fixed: they change in time, co-evolving with the motifs of cooperators in the population. The evolutionary outcome of such a self-organization process will depend sensitively on this co-evolution, which can be followed using a time-dependent AGoS.
Dynamical Model
Let us consider pairwise interactions between individuals who can behave either as a Cooperator (C) or Defector (D). Whenever cheated by a D, a C receives a payoff S (the sucker's payoff), while the D receives T (temptation to defect). Mutual cooperation provides R (reward) to each player, while mutual defection provides P (punishment). One obtains the prisoner's dilemma (PD) — the most famous metaphor of cooperation — whenever T>R>P>S. We formalize the dilemma in terms of a single parameter B (benefit) by defining T = B>1, R = 1, S = 1-B and P = 0. The results remain quantitatively unaltered if one adopts the more popular parameterization T = b, R = b–c, S = −c and P = 0.
In the framework of evolutionary game theory, we adopt a stochastic update rule of social learning, where in each time step a random individual i imitates the strategy of a randomly selected neighbor j with a probability that increases with the fitness difference between them [14], [24], [47]–[50] (see Methods). In the limit of well-mixed populations of size N
[10], [11], the frequency j/N of Cs will increase (decrease) in time depending on whether the gradient of selection [51], [52]
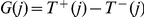 is positive (negative), where
is positive (negative), where 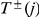 [47] represent the probabilities to increase and decrease the number of Cs in the population by one. For the PD,
[47] represent the probabilities to increase and decrease the number of Cs in the population by one. For the PD, 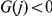 for all j and, as a result, cooperation will most probably die out. The same scenario is obtained when N→∞, where we recover the scenarios described by the famous replicator dynamics [11], [47]–[50]. The elegance of this result (despite the doomsday scenario for Cs) is best appreciated when we realize that the population ends up adopting the Nash-equilibrium of a PD game interaction between two individuals: everybody defects. Consequently, there is no difference in the outcome of the game, from an individual or from a (collective) population wide perspective, a feature that, as discussed below, will not remain true in structured populations.
for all j and, as a result, cooperation will most probably die out. The same scenario is obtained when N→∞, where we recover the scenarios described by the famous replicator dynamics [11], [47]–[50]. The elegance of this result (despite the doomsday scenario for Cs) is best appreciated when we realize that the population ends up adopting the Nash-equilibrium of a PD game interaction between two individuals: everybody defects. Consequently, there is no difference in the outcome of the game, from an individual or from a (collective) population wide perspective, a feature that, as discussed below, will not remain true in structured populations.
Results
The previous analysis assumes finite yet structureless populations, a feature which is seldom observed in practice, with strong implications in many natural phenomena. A homogeneous network of size N represents the simplest case of a structured population, where all individuals engage in the same number of games k with their first neighbors, also imitating their behavior. Let us consider a homogeneous random network — also called a regular random graph — in which all links are randomly connected, while all nodes have each the same number of links [4], [53], [54]. In this case, individuals with the same strategy no longer share the same fitness: fitness becomes context-dependent. The same happens to 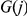 , becoming hard to define it analytically. Consequently, we define the AGoS — denoting it by
, becoming hard to define it analytically. Consequently, we define the AGoS — denoting it by 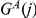 — as the average i) over all possible transitions taking place in every node of the network throughout evolution, and ii) over a large number of networked evolutions (see Methods). This AGoS, which must be computed numerically, becomes therefore network dependent but context independent, as it recovers its population averaged, or mean-field, character. Hence, the AGoS may constitute a powerful tool to understand dynamical processes at a population-wide scale stemming from individually defined, but often seemingly unrelated, rules.
— as the average i) over all possible transitions taking place in every node of the network throughout evolution, and ii) over a large number of networked evolutions (see Methods). This AGoS, which must be computed numerically, becomes therefore network dependent but context independent, as it recovers its population averaged, or mean-field, character. Hence, the AGoS may constitute a powerful tool to understand dynamical processes at a population-wide scale stemming from individually defined, but often seemingly unrelated, rules.
The results for 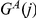 on homogeneous networks of size N = 103, k = 4 and different values of B are shown in Fig. 1a. Unlike well-mixed populations where cooperation has no chance, homogeneous networks can sustain cooperation [12], [14], [15], [24], [54]. The shape of
on homogeneous networks of size N = 103, k = 4 and different values of B are shown in Fig. 1a. Unlike well-mixed populations where cooperation has no chance, homogeneous networks can sustain cooperation [12], [14], [15], [24], [54]. The shape of 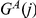 no longer pictures a defection dominance dilemma typical of a PD, but a gradient of selection similar to what one observes under co-existence dilemmas in well-mixed populations [43]. In other words, even though every individual engages in a PD, from a global, population-wide perspective, homogeneous networks are able to create an emerging collective dynamics promoting the co-existence between Cs and Ds. As we show below, the emergence of an unanticipated global (macroscopic) dynamics from a distinct individual (microscopic) dynamics pervades throughout all evolutionary dynamical processes in structured populations studied here. The co-existence point xR (see Fig. 1a) is associated with the internal root (
no longer pictures a defection dominance dilemma typical of a PD, but a gradient of selection similar to what one observes under co-existence dilemmas in well-mixed populations [43]. In other words, even though every individual engages in a PD, from a global, population-wide perspective, homogeneous networks are able to create an emerging collective dynamics promoting the co-existence between Cs and Ds. As we show below, the emergence of an unanticipated global (macroscopic) dynamics from a distinct individual (microscopic) dynamics pervades throughout all evolutionary dynamical processes in structured populations studied here. The co-existence point xR (see Fig. 1a) is associated with the internal root (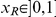 ) of
) of 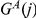 — inexistent in well-mixed populations — whose location decreases with increasing B. Together with xR one obtains a coordination root (
— inexistent in well-mixed populations — whose location decreases with increasing B. Together with xR one obtains a coordination root (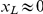 , see Fig. 1a) of
, see Fig. 1a) of 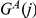 since, in the absence of cooperative partners, Cs will always be disadvantageous. However, the impact of
since, in the absence of cooperative partners, Cs will always be disadvantageous. However, the impact of 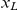 is minor, as shown in Fig. 1b (see discussion below). Remarkably, this characterization remains valid for other types of homogeneous networks, such as lattices and regular rings (as well as for other possible mechanisms of strategy update) whereas differences in the positions of the stable root (xR) in of
is minor, as shown in Fig. 1b (see discussion below). Remarkably, this characterization remains valid for other types of homogeneous networks, such as lattices and regular rings (as well as for other possible mechanisms of strategy update) whereas differences in the positions of the stable root (xR) in of 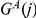 and their dependence on B correlate perfectly with results obtained previously [14], [18], [24], [54]–[58], where steady-states of evolution were analyzed for such structures (see Text S1).
and their dependence on B correlate perfectly with results obtained previously [14], [18], [24], [54]–[58], where steady-states of evolution were analyzed for such structures (see Text S1).
Figure 1. Time-independent AGoS.

(a) We plot GA(j) for a population of players interacting via a PD in a homogeneous random network, for two values of the benefit B. Globally, GA(j) indicates that the population evolves towards a co-existence scenario. (b) Stationary distributions showing the pervasiveness of each fraction j/N in time. In line with the AGoS in a), the population spends most of the time in the vicinity of the stable-like root xR of GA(j). When j/N≈0, Cs become disadvantageous, giving rise to an unstable-like root xL of GA(j) which, however, plays a minor role as shown (N = 103, <k> = 4 and β = 1.0). Homogeneous random networks were obtained by repeatedly swapping the ends of pairs of randomly chosen links of a regular lattice [54].
Fig. 1a shows that, as we change focus from an individual to a population wide perspective, one witnesses the emergence of an effective game transformation, as evidenced by 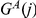 , which brings along important consequences: For instance, the fixation time — the time required for cooperators to invade the entire population —becomes much larger in homogeneous networks when compared to well-mixed populations, as the population spends a large period of time in the vicinity of the xR, mainly when selection is strong (see Methods). This, in turn, is responsible for computer simulations to spend arbitrary amounts of time in the same configuration, even when, in the absence of mutations (as is the case here), the only absorbing states are associated with monomorphic configurations of the population, that is, with configurations comprising cooperators-only or defectors-only. The stationary distribution (Fig. 1b), which represents the pervasiveness of each fraction of Cs in time, confirms the scenario portrayed by
, which brings along important consequences: For instance, the fixation time — the time required for cooperators to invade the entire population —becomes much larger in homogeneous networks when compared to well-mixed populations, as the population spends a large period of time in the vicinity of the xR, mainly when selection is strong (see Methods). This, in turn, is responsible for computer simulations to spend arbitrary amounts of time in the same configuration, even when, in the absence of mutations (as is the case here), the only absorbing states are associated with monomorphic configurations of the population, that is, with configurations comprising cooperators-only or defectors-only. The stationary distribution (Fig. 1b), which represents the pervasiveness of each fraction of Cs in time, confirms the scenario portrayed by 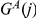 in Fig. 1a, stressing the similarities with the evolutionary dynamics in (finite) well-mixed populations under co-existence dilemmas [50], [59], [60], and putting in evidence the marked difference between individual preferences and the population-wide dynamics.
in Fig. 1a, stressing the similarities with the evolutionary dynamics in (finite) well-mixed populations under co-existence dilemmas [50], [59], [60], and putting in evidence the marked difference between individual preferences and the population-wide dynamics.
In Fig. 1, our analysis was limited to 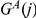 that is, we averaged over the entire time span of all runs. However, the AGoS itself evolves in time —
that is, we averaged over the entire time span of all runs. However, the AGoS itself evolves in time — 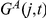 — as detailed in Methods. Let us explore this time dependence of the AGoS. If, at the beginning of each simulated evolution, Cs and Ds are randomly spread in the network, the occurrence of clusters of the same strategy will not occur in general. Hence, for the PD we have that
— as detailed in Methods. Let us explore this time dependence of the AGoS. If, at the beginning of each simulated evolution, Cs and Ds are randomly spread in the network, the occurrence of clusters of the same strategy will not occur in general. Hence, for the PD we have that 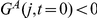 in general. As populations evolve, Cs (Ds) breed Cs (Ds) in their neighborhood, promoting the assortment of strategies, with implications both on the fitness of each player and on the shape (and sign) of
in general. As populations evolve, Cs (Ds) breed Cs (Ds) in their neighborhood, promoting the assortment of strategies, with implications both on the fitness of each player and on the shape (and sign) of 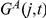 . In Fig. 2a we plot
. In Fig. 2a we plot 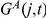 for three particular generations, whereas Fig. 2b portrays the time evolution of the internal roots of
for three particular generations, whereas Fig. 2b portrays the time evolution of the internal roots of 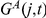 , on which we superimposed two evolutionary runs starting with 50% of Cs randomly placed in the population. As
, on which we superimposed two evolutionary runs starting with 50% of Cs randomly placed in the population. As 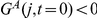 , the fraction of cooperators will start decreasing (Fig. 2a). However, with time, strategy assortment leads to the emergence of a co-existence root, toward which the fraction cooperators converges. The ensuing coexistence between Cs and Ds, entirely described by the shape of
, the fraction of cooperators will start decreasing (Fig. 2a). However, with time, strategy assortment leads to the emergence of a co-existence root, toward which the fraction cooperators converges. The ensuing coexistence between Cs and Ds, entirely described by the shape of 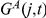 , steams from the self-organization of Cs and Ds in the network, defining a global dynamics which is impossible to foresee solely from the nature of the local (PD) interactions.
, steams from the self-organization of Cs and Ds in the network, defining a global dynamics which is impossible to foresee solely from the nature of the local (PD) interactions.
Figure 2. Time-dependent AGoS.

(a) We plot GA(j,t) for three different instants of evolutionary time. Each line provides a snapshot for a given moment, portraying the emergence of a population-wide (time-dependent) co-existence-like dilemma stemming from an individual (time-independent) defection dominant dilemma (PD). (b) The circles show the position of the different interior roots of GA(j,t), whereas the solid (dark blue) line and (light blue) crosses show two independent evolutionary runs starting from 50% of Cs and Ds randomly placed in the networked population. Open (full) circles stand for unstable, xL (stable, xR) roots of GA(j,t) (B = 1.01, N = 103, <k> = 4 and β = 10.0).
It is now generally accepted, however, that homogeneous networks provide a simplified picture of real interaction networks [5], [44], [61], [62]. Most social structures share a marked heterogeneity, where a few nodes exhibit a large number of connections, whereas most nodes comprise just a few. The fingerprint of this heterogeneity is provided by the associated degree distributions, which exhibit a broad-scale shape, often resembling a power-law [4], [44], [61]. In the following we use 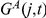 to understand how heterogeneity shifts the internal roots in Fig. 1 to the right, thereby transforming a co-existence scenario into a coordination one. To this end, we compute
to understand how heterogeneity shifts the internal roots in Fig. 1 to the right, thereby transforming a co-existence scenario into a coordination one. To this end, we compute 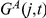 employing scale-free (SF) networks of Barabási and Albert (BA) (see Methods) [61].
employing scale-free (SF) networks of Barabási and Albert (BA) (see Methods) [61].
Fig. 3a shows 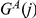 for BA networks of N = 103 nodes and an average degree k = 4, whereas the circles in Fig. 3b portray the time evolution of the internal roots of
for BA networks of N = 103 nodes and an average degree k = 4, whereas the circles in Fig. 3b portray the time evolution of the internal roots of 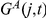 . Heterogeneous networks lead to a global dynamics dominated by a coordination threshold, originating the appearance of two basins of attraction split by an unstable root xL of
. Heterogeneous networks lead to a global dynamics dominated by a coordination threshold, originating the appearance of two basins of attraction split by an unstable root xL of 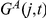 , analogous to the evolutionary dynamics under 2-person and N-person Stag-hunt dilemmas in unstructured populations [45], [51], [63], [64]. This unstable root represents the critical fraction of Cs above which they are able to assort, thereby invading highly connected nodes, rendering cooperation an advantageous strategy, as Cs acquire a higher probability of being imitated than Ds. On SF networks the requirement to reach the hubs, which ensures the formation of cooperative star-like clusters [39], [52], makes invasion harder for isolated Cs. This moves the unstable root located close to j/N≈0 in homogeneous networks (see Fig. 1) to higher fractions of Cs. Yet, once this coordination is overcome, Cs benefit from the strong influence of hubs to rapidly spread in the population, eventually leading to fixation. As a result, the stable internal root which characterizes
, analogous to the evolutionary dynamics under 2-person and N-person Stag-hunt dilemmas in unstructured populations [45], [51], [63], [64]. This unstable root represents the critical fraction of Cs above which they are able to assort, thereby invading highly connected nodes, rendering cooperation an advantageous strategy, as Cs acquire a higher probability of being imitated than Ds. On SF networks the requirement to reach the hubs, which ensures the formation of cooperative star-like clusters [39], [52], makes invasion harder for isolated Cs. This moves the unstable root located close to j/N≈0 in homogeneous networks (see Fig. 1) to higher fractions of Cs. Yet, once this coordination is overcome, Cs benefit from the strong influence of hubs to rapidly spread in the population, eventually leading to fixation. As a result, the stable internal root which characterizes 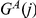 in homogeneous networks collapses into the vicinity of j = N on heterogeneous structures, promoting the evolution towards fully cooperative populations. Naturally, the location of the unstable root of
in homogeneous networks collapses into the vicinity of j = N on heterogeneous structures, promoting the evolution towards fully cooperative populations. Naturally, the location of the unstable root of 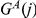 is an increasing function of B (see Fig. 3a). It is noteworthy that our results remain qualitatively valid for other update rules, such as the discrete analogue of the replicator dynamics on graphs, used in many references, e.g., [16], [27], [38], [65]. In fact, the AGoS is capable of identifying particular features of such dynamics: For instance, the partially deterministic nature of such update rule may lead to evolutionary deadlocks in heterogeneous (scale-free) networks, creating stationary states close to full cooperation [16], [27]. In such situation, the AGoS will reflect the occurrence of these stationary configurations by shifting to the left-hand side the stable (xR) equilibrium, which may no longer coincide with j = N, remaining, however, in its vicinity.
is an increasing function of B (see Fig. 3a). It is noteworthy that our results remain qualitatively valid for other update rules, such as the discrete analogue of the replicator dynamics on graphs, used in many references, e.g., [16], [27], [38], [65]. In fact, the AGoS is capable of identifying particular features of such dynamics: For instance, the partially deterministic nature of such update rule may lead to evolutionary deadlocks in heterogeneous (scale-free) networks, creating stationary states close to full cooperation [16], [27]. In such situation, the AGoS will reflect the occurrence of these stationary configurations by shifting to the left-hand side the stable (xR) equilibrium, which may no longer coincide with j = N, remaining, however, in its vicinity.
Figure 3. AGoS on BA networks.

(a) Starting from a defection dominant PD played at an individual level, a coordination dynamics emerges at a global, population-wide scale, for the three values of B depicted. (b) Evolution of the unstable root xL of GA(j,t) (open circles), exhibiting the time-dependence of the global dynamics; solid (dark blue) line and (light blue) crosses show two independent evolutionary runs starting from 50% of Cs and Ds randomly placed. The ultimate fate of Cs in each run depends on whether the population composition crosses over the time-dependent value xL of GA(j,t) thereby overcoming the dynamical coordination barrier during evolution. (B = 1.25, N = 103, <k> = 4 and β = 0.1). BA networks were obtained combining growth and preferential attachment, following the model proposed by Barabási and Albert [61].
The existence of a coordination barrier for Cs in heterogeneous networks, which must first occupy the hubs before outcompeting Ds, leads to an intricate interplay between the time-dependent decline of xL (see Fig. 3b) and the global fraction of Cs. In Fig. 3b we show, with full lines, two evolutions in BA networks (for the same value of B = 1.25): One, which fixates in full cooperation and another, which fixates in full defection. Whenever the fraction of cooperators j/N remains sizeable for long enough, xL will eventually decrease to values satisfying j/N>xL, such that the global coordination barrier is overcome and the population will fixate into full cooperation (light blue line). Otherwise, j/N may remain always below xL with the population fixating into full defection (dark blue line). Clearly, heterogeneous networks lead to the emergence of a global coordination barrier and associated basins of attraction that evolve in time, in a way which is well described by the time-dependent AGoS.
Discussion
To establish the link between individual and collective behavior constitutes, undeniably, one of the main goals of the analysis of any complex multi-particle or multi-component system [66]. Here we establish such a link showing how it depends on the underlying network topology. Our study shows that behavioral dynamics of individuals facing a cooperation dilemma in social networks can be understood as though the network structure is absent but individuals face a different dilemma: The structural organization of a population of self-regarding individuals helps circumventing the Nash equilibrium of a cooperation dilemma by creating a new dynamical system that can be globally characterized by two internal fixed points, xL (unstable) and xR (stable). While a single defector will be always advantageous (creating an unstable fixed point at x = 1.0), a single cooperator will be always disadvantageous (prompting a stable equilibrium at x = 0.0). As cooperators assort into stable clusters, they may also become advantageous above a certain critical fraction of cooperators (k/N>xL, associated with a critical cluster size) and below another critical fraction of cooperators xR, above which defectors will be able to ripe again the benefits of exploiting the many surrounding cooperators. Whereas in homogeneous networks the stable equilibria dictate the overall dynamics — as in co-existence dilemmas — heterogeneous networks create a global dynamics mainly dominated by the unstable equilibria, creating a coordination problem.
Strictly speaking, such a dynamical system resulting from individuals interacting (locally) via a two-person game, cannot be mapped onto a two-person evolutionary game in a well-mixed population, since the latter can only comply with a maximum of one internal fixed point [43]. On the contrary, such dynamics resembles that from, e.g., N-person dilemmas [67], [68] in the presence of coordination thresholds [51],[64],[69]. It is as if the global dynamics of a 2-person dilemma in structured populations can be properly described as a time-dependent N-person dilemma, in which the coordination or co-existence features emerge from the population structure itself, with different network topologies emphasizing differently this co-existence/coordination dichotomy.
It is worth emphasizing that the approach developed here in the context of the two-person PD may be useful ― and immediately applicable ― in understanding the evolutionary dynamics of other game interactions, as well as in understanding other aspects of human sociality that extend beyond cooperation. From human behaviors and ideas, to diseases spreading or to individual preferences, most have been modeled as a person-to-person spreading process embedded in a social network [5], [8], [62]. In such frameworks, the identification and categorization of the global, population-wide dynamics which emerges from the apparently unrelated nature of the local interactions may enable one to anticipate the emergent outcomes of such complex biological and social systems.
Methods
Evolution is modelled via a stochastic birth-death process [47], [70], [71]. Each individual x adopts the strategy of a randomly selected neighbour y with probability given by the Fermi function 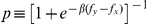 [14], [47], where fx (fy) stands for the accumulate payoff of x (y) and
[14], [47], where fx (fy) stands for the accumulate payoff of x (y) and 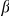 controls the intensity of selection. In structured populations, the difference of the probabilities to increase and decrease the number of Cs (
controls the intensity of selection. In structured populations, the difference of the probabilities to increase and decrease the number of Cs (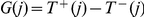 ) becomes context dependent, but can be computed numerically. For each individual i we compute the probability of changing behavior at time t,
) becomes context dependent, but can be computed numerically. For each individual i we compute the probability of changing behavior at time t, 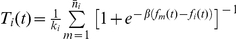 , where
, where 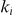 stands for the degree of node i and
stands for the degree of node i and 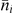 for the number of neighbors of i having a strategy different from that of i. The time-dependent
AGoS at a given time t of simulation p, where we have j
Cs in the population of size N, is defined as
for the number of neighbors of i having a strategy different from that of i. The time-dependent
AGoS at a given time t of simulation p, where we have j
Cs in the population of size N, is defined as 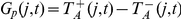 , where
, where 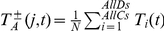 . For a given network type, we run Ω = 2×107 simulations (using 103 randomly generated networks) starting from all possible initial fractions j/N of cooperators. Each configuration of the population is defined here by the fraction j/N of cooperators. Evolutions run for Λ = 105 time steps. Hence, the overall, time-independent, AGoS is given by the average
. For a given network type, we run Ω = 2×107 simulations (using 103 randomly generated networks) starting from all possible initial fractions j/N of cooperators. Each configuration of the population is defined here by the fraction j/N of cooperators. Evolutions run for Λ = 105 time steps. Hence, the overall, time-independent, AGoS is given by the average 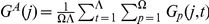 over all simulations and time-steps. The time-dependent gradients
over all simulations and time-steps. The time-dependent gradients 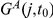 for a particular generation t
0 (and corresponding roots shown in Fig. 2 and 3b) were computed averaging over the configurations occurring during N previous time-steps (1 generation):
for a particular generation t
0 (and corresponding roots shown in Fig. 2 and 3b) were computed averaging over the configurations occurring during N previous time-steps (1 generation): 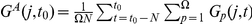 . The stationary distributions pictured in Fig. 1b were obtained computing the fraction of time the population spent in each overall configuration j/N. In some specific limits — in particular, for weak selection or well-mixed populations — our numerical approach will provide results analogous to those obtained with other methods (see for instance [20], [22], [43], [46], [47], [50], [60], [71]–[74]).
. The stationary distributions pictured in Fig. 1b were obtained computing the fraction of time the population spent in each overall configuration j/N. In some specific limits — in particular, for weak selection or well-mixed populations — our numerical approach will provide results analogous to those obtained with other methods (see for instance [20], [22], [43], [46], [47], [50], [60], [71]–[74]).
Homogeneous random networks were obtained by repeatedly swapping the ends of pairs of randomly chosen links of a regular network [54]. BA networks were obtained combining growth and preferential attachment, following the model proposed by Barabási and Albert [4], [61]. All networks used have N = 103 nodes and an average degree k = 4.
Supporting Information
Supporting Text (containing one additional figure) on the analysis of the evolutionary steady states in homogeneous networks.
(PDF)
Footnotes
Competing Interests: The authors have declared that no competing interests exist.
Funding: Financial support from FCT-Portugal is gratefully acknowledged. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Granovetter M. The strength of weak ties. Am J Sociol. 1973;78:1360. [Google Scholar]
- 2.Watts D. A twenty-first century science. Nature. 2007;445(7127):489. doi: 10.1038/445489a. [DOI] [PubMed] [Google Scholar]
- 3.Lloyd AL, May RM. How viruses spread among computers and people. Science. 2001;292:1316–1317. doi: 10.1126/science.1061076. [DOI] [PubMed] [Google Scholar]
- 4.Dorogovtsev SN. Lectures on Complex Networks. Oxford: Oxford University Press; 2010. [Google Scholar]
- 5.Fowler JH, Christakis NA. Cooperative behavior cascades in human social networks. Proc Natl Acad Sci U S A. 2010;107(12):5334–5338. doi: 10.1073/pnas.0913149107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Centola D. The Spread of Behavior in an Online Social Network Experiment. Science. 2010;329:1194. doi: 10.1126/science.1185231. [DOI] [PubMed] [Google Scholar]
- 7.Lazer D, Pentland A, Adamic L, Aral S, Barabasi AL, et al. Computational social science. Science. 2009;323(5915):721–723. doi: 10.1126/science.1167742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Barrat A, Barthelemy M, Vespignani A. Dynamical processes in complex networks. Cambridge: Cambridge University Press; 2008. [Google Scholar]
- 9.Onnela JP, Reed-Tsochas F. Spontaneous emergence of social influence in online systems. Proc Natl Acad Sci U S A. 2010;107(43):18375–18380. doi: 10.1073/pnas.0914572107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Maynard-Smith J. Evolution and the Theory of Games. Cambridge: Cambridge University Press; 1982. [Google Scholar]
- 11.Sigmund K. The Calculus of Selfishness. Princeton: Princeton University Press; 2010. [Google Scholar]
- 12.Nowak MA, May RM. Evolutionary games and spatial chaos. Nature. 1992;359:826–829. [Google Scholar]
- 13.Nakamaru M, Matsuda H, Iwasa Y. The evolution of cooperation in a lattice-structured population. Journal of Theoretical Biology. 1997;184(1):65–81. doi: 10.1006/jtbi.1996.0243. [DOI] [PubMed] [Google Scholar]
- 14.Szabó G, Toke C. Evolutionary prisoner's dilemma game on a square lattice. Phys Rev E. 1998;58:69–73. [Google Scholar]
- 15.Hauert C. Effects of space in 2×2 games. Int J Bifurcation Chaos. 2002;12:1531–1548. [Google Scholar]
- 16.Santos FC, Pacheco JM. Scale-free networks provide a unifying framework for the emergence of cooperation. Phys Rev Lett. 2005;95(9):098104. doi: 10.1103/PhysRevLett.95.098104. [DOI] [PubMed] [Google Scholar]
- 17.Lieberman E, Hauert C, Nowak MA. Evolutionary dynamics on graphs. Nature. 2005;433(7023):312–316. doi: 10.1038/nature03204. [DOI] [PubMed] [Google Scholar]
- 18.Santos FC, Pacheco JM, Lenaerts T. Evolutionary dynamics of social dilemmas in structured heterogeneous populations. Proc Natl Acad Sci U S A. 2006;103(9):3490–3494. doi: 10.1073/pnas.0508201103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Santos FC, Rodrigues JF, Pacheco JM. Graph topology plays a determinant role in the evolution of cooperation. Proc Biol Sci. 2006;273(1582):51–55. doi: 10.1098/rspb.2005.3272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ohtsuki H, Hauert C, Lieberman E, Nowak MA. A simple rule for the evolution of cooperation on graphs and social networks. Nature. 2006;441(7092):502–505. doi: 10.1038/nature04605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ohtsuki H, Nowak MA. The replicator equation on graphs. Journal of Theoretical Biology. 2006;243(1):86–97. doi: 10.1016/j.jtbi.2006.06.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Taylor PD, Day T, Wild G. Evolution of cooperation in a finite homogeneous graph. Nature. 2007;447:469–472. doi: 10.1038/nature05784. [DOI] [PubMed] [Google Scholar]
- 23.Taylor PD, Day T, Wild G. From inclusive fitness to fixation probability in homogeneous structured populations. Journal of Theoretical Biology. 2007;249(1):101–110. doi: 10.1016/j.jtbi.2007.07.006. [DOI] [PubMed] [Google Scholar]
- 24.Szabó G, Fáth G. Evolutionary games on graphs. Phys Rep. 2007;446(4–6):97–216. [Google Scholar]
- 25.Gómez-Gardeñes J, Campillo M, Floría LM, Moreno Y. Dynamical Organization of Cooperation in Complex Topologies. Phys Rev Lett. 2007;98:108103. doi: 10.1103/PhysRevLett.98.108103. [DOI] [PubMed] [Google Scholar]
- 26.Ohtsuki H, Nowak MA, Pacheco JM. Breaking the symmetry between interaction and replacement in evolutionary dynamics on graphs. Phys Rev Lett. 2007;98(10):108106. doi: 10.1103/PhysRevLett.98.108106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Gómes-Gardeñes J, Poncela J, Floría LM, Moreno Y. Natural selection of cooperation and degree hierarchy in heterogeneous populations. J Theo Bio. 2008;253(2):296–301. doi: 10.1016/j.jtbi.2008.03.007. [DOI] [PubMed] [Google Scholar]
- 28.Poncela J, Gómes-Gardeñes J, Floría LM, Moreno Y. Robustness of cooperation in the evolutionary prisoner's dilemma on complex networks. New J Phys. 2007;9:187. [Google Scholar]
- 29.Masuda N. Participation costs dismiss the advantage of heterogeneous networks in evolution of cooperation. Proceedings of the Royal Society B: Biological Sciences. 2007;274(1620):1815–1821. doi: 10.1098/rspb.2007.0294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Lehmann L, Keller L, Sumpter D. The evolution of helping and harming on graphs: the return of the inclusive fitness effect. Journal of evolutionary biology. 2007;20(6):2284–2295. doi: 10.1111/j.1420-9101.2007.01414.x. [DOI] [PubMed] [Google Scholar]
- 31.Szolnoki A, Perc M, Danku Z. Towards effective payoffs in the prisoner's dilemma game on scale-free networks. Physica A. 2008;387:2075–2082. [Google Scholar]
- 32.Vukov J, Szabó G, Szolnoki A. Evolutionary prisoner's dilemma game on Newman-Watts networks. Physical Review E. 2008;77(2):026109. doi: 10.1103/PhysRevE.77.026109. [DOI] [PubMed] [Google Scholar]
- 33.Perc M. Evolution of cooperation on scale-free networks subject to error and attack. New Journal of Physics. 2009;11:033027. [Google Scholar]
- 34.Floría L, Gracia-Lázaro C, Gómez-Gardeñes J, Moreno Y. Social network reciprocity as a phase transition in evolutionary cooperation. Physical Review E. 2009;79(2):026106. doi: 10.1103/PhysRevE.79.026106. [DOI] [PubMed] [Google Scholar]
- 35.Perc M, Szolnoki A. Coevolutionary games–A mini review. BioSystems. 2010;99(2):109–125. doi: 10.1016/j.biosystems.2009.10.003. [DOI] [PubMed] [Google Scholar]
- 36.Perc M, Wang Z. Heterogeneous aspirations promote cooperation in the prisoner's dilemma game. PLoS One. 2010;5(12):e15117. doi: 10.1371/journal.pone.0015117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Marshall JAR. Ultimate causes and the evolution of altruism. Behavioral Ecology and Sociobiology. 2010:1–10. [Google Scholar]
- 38.Hauert C, Doebeli M. Spatial structure often inhibits the evolution of cooperation in the snowdrift game. Nature. 2004;428(6983):643–646. doi: 10.1038/nature02360. [DOI] [PubMed] [Google Scholar]
- 39.Santos FC, Santos MD, Pacheco JM. Social diversity promotes the emergence of cooperation in public goods games. Nature. 2008;454(7201):213–216. doi: 10.1038/nature06940. [DOI] [PubMed] [Google Scholar]
- 40.Szabó G, Vukov J, Szolnoki A. Phase diagrams for an evolutionary prisoner's dilemma game on two-dimensional lattices. Physical Review E. 2005;72(4):047107. doi: 10.1103/PhysRevE.72.047107. [DOI] [PubMed] [Google Scholar]
- 41.Smith JM, Parker GA. The logic of asymmetric contests. Animal Behaviour. 1976;24(1):159–175. [Google Scholar]
- 42.Gintis H. Game Theory Evolving. Princeton, Princeton University Press; 2000. [Google Scholar]
- 43.Hofbauer J, Sigmund K. Evolutionary Games and Population Dynamics. Cambridge, UK: Cambridge Univ. Press; 1998. [Google Scholar]
- 44.Amaral LA, Scala A, Barthelemy M, Stanley HE. Classes of small-world networks. Proc Natl Acad Sci U S A. 2000;97(21):11149–11152. doi: 10.1073/pnas.200327197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Skyrms B. 2004. The Stag Hunt and the Evolution of Social Structure: Cambridge University Press.
- 46.Price GR. Selection and covariance. 520Nature. 1970;227(5257) doi: 10.1038/227520a0. [DOI] [PubMed] [Google Scholar]
- 47.Traulsen A, Nowak MA, Pacheco JM. Stochastic dynamics of invasion and fixation. Phys Rev E. 2006;74(1 Pt 1):011909. doi: 10.1103/PhysRevE.74.011909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Traulsen A, Pacheco JM, Nowak MA. Pairwise comparison and selection temperature in evolutionary game dynamics. J Theor Biol. 2007;246(3):522–529. doi: 10.1016/j.jtbi.2007.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Traulsen A, Nowak MA, Pacheco JM. Stochastic payoff evaluation increases the temperature of selection. J Theor Biol. 2007;244(2):349–356. doi: 10.1016/j.jtbi.2006.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Traulsen A, Pacheco JM, Imhof LA. Stochasticity and evolutionary stability. Phys Rev E. 2006;74(2 Pt 1):021905. doi: 10.1103/PhysRevE.74.021905. [DOI] [PubMed] [Google Scholar]
- 51.Pacheco JM, Santos FC, Souza MO, Skyrms B. Evolutionary dynamics of collective action in N-person stag hunt dilemmas. Proc Biol Sci. 2009;276(1655):315–321. doi: 10.1098/rspb.2008.1126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Pacheco JM, Pinheiro FL, Santos FC. Population Structure Induces a Symmetry Breaking Favoring the Emergence of Cooperation. PLoS Comput Biol. 2009;5(12):e1000596. doi: 10.1371/journal.pcbi.1000596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Bollobás B. 2001. Random graphs: Cambridge Univ Press.
- 54.Santos FC, Rodrigues JF, Pacheco JM. Epidemic spreading and cooperation dynamics on homogeneous small-world networks. Phys Rev E. 2005;72(5 Pt 2):056128. doi: 10.1103/PhysRevE.72.056128. [DOI] [PubMed] [Google Scholar]
- 55.Hauert C, Szabó G. Game theory and physics. Am J Phys. 2005;73(5):405–414. [Google Scholar]
- 56.Perc M. Coherence resonance in a spatial prisoner's dilemma game. New J Phys. 2006;8:22. [Google Scholar]
- 57.Perc M, Szolnoki A. Social diversity and promotion of cooperation in the spatial prisoner's dilemma game. Phys Rev E. 2008;77(1 Pt 1):011904. doi: 10.1103/PhysRevE.77.011904. [DOI] [PubMed] [Google Scholar]
- 58.Szabó G, Szolnoki A. Cooperation in spatial prisonerís dilemma with two types of players for increasing number of neighbors. Phys Rev E. 2009;79(1):016106. doi: 10.1103/PhysRevE.79.016106. [DOI] [PubMed] [Google Scholar]
- 59.Antal T, Scheuring I. Fixation of strategies for an evolutionary game in finite populations. Bull Math Biol. 2006;68:1923–1944. doi: 10.1007/s11538-006-9061-4. [DOI] [PubMed] [Google Scholar]
- 60.Chalub FACC, Souza MO. From discrete to continuous evolution models: a unifying approach to drift-diffusion and replicator dynamics. Theoretical Population Biology. 2009;76(4):268–277. doi: 10.1016/j.tpb.2009.08.006. [DOI] [PubMed] [Google Scholar]
- 61.Barabási AL, Albert R. Emergence of scaling in random networks. Science. 1999;286(5439):509–512. doi: 10.1126/science.286.5439.509. [DOI] [PubMed] [Google Scholar]
- 62.Christakis NA, Fowler JH. The collective dynamics of smoking in a large social network. N Engl J Med. 2008;358(21):2249–2258. doi: 10.1056/NEJMsa0706154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Skyrms B. The Stag Hunt Proceedings and Addresses of the American Philosophical Association. 2001;75(2):31–41. [Google Scholar]
- 64.Santos FC, Pacheco JM. Risk of collective failure provides an escape from the tragedy of the commons. Proc Natl Acad Sci U S A. 2011;108(26):10421. doi: 10.1073/pnas.1015648108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Santos FC, Pacheco JM. A new route to the evolution of cooperation. J Evolution Biol. 2006;19:726–733. doi: 10.1111/j.1420-9101.2005.01063.x. [DOI] [PubMed] [Google Scholar]
- 66.Schelling TC. Micromotives and Macrobehavior. New York: Norton; 1978. [Google Scholar]
- 67.Kollock P. Social Dilemmas: The anatomy of cooperation. Annu Rev Sociol. 1998;24:183–214. [Google Scholar]
- 68.Gokhale C, Traulsen A. Evolutionary games in the multiverse. Proc Natl Acad Sci U S A. 2010;107(12):5500. doi: 10.1073/pnas.0912214107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Souza MO, Pacheco JM, Santos FC. Evolution of Cooperation under N-person Snowdrift Games. J Theo Bio. 2009;260(4):581–588. doi: 10.1016/j.jtbi.2009.07.010. [DOI] [PubMed] [Google Scholar]
- 70.Karlin S, Taylor HMA. A first course in Stochastic Processes. London: Academic; 1975. [Google Scholar]
- 71.Nowak MA, Sasaki A, Taylor C, Fudenberg D. Emergence of cooperation and evolutionary stability in finite populations. Nature. 2004;428(6983):646–650. doi: 10.1038/nature02414. [DOI] [PubMed] [Google Scholar]
- 72.Page KM, Nowak MA. Unifying evolutionary dynamics. Journal of Theoretical Biology. 2002;219(1):93–98. [PubMed] [Google Scholar]
- 73.Traulsen A, Hauert C. Stochastic evolutionary game dynamics. Schuster H-G, editor. Reviews of Nonlinear Dynamics and Complexity Wiley-VCH 2009 [Google Scholar]
- 74.Chalub FACC, Souza MO. Continuous Models for Genetic Evolution in Large Populations. Dynamics, Games and Science I. 2011:239–242. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supporting Text (containing one additional figure) on the analysis of the evolutionary steady states in homogeneous networks.
(PDF)