

Abstract
We studied the effects of dietary cholesterol intake on lipid and lipoprotein levels in healthy young women (n = 13) who were otherwise eating an American Heart Association (AHA) diet. The study used a randomized, three-way crossover design to determine the effects of 0, 1, or 3 eggs added per day (dietary cholesterol range, 108 to 667 mg/d). Each of the three diets was eaten for 8 weeks, with a washout period between diets. Three fasting blood samples were obtained during the last 3 weeks of each diet period to observe changes in fasting plasma lipid levels associated with the menstrual cycle. We also obtained blood just before and 4 and 8 hours after the subjects ingested a standard high-fat formula. During the menstrual cycle, total cholesterol and LDL cholesterol levels fell by 0.051 mmol/L (1.99 mg/dL) and 0.064 mmol/L (2.48 mg/dL) per week, respectively. HDL cholesterol concentrations increased by 0.060 mmol/L (2.3 mg/dL) per week during the first half of the cycle and then fell by 0.050 mmol/L (1.94 mg/dL) per week during the second half. Therefore, all statistical analyses were performed on values adjusted to midcycle. Total fasting cholesterol concentrations increased by 0.073 mmol/L (2.81 mg/dL) per 100 mg dietary cholesterol added to the diet per day (P=.001). LDL cholesterol increased by 0.054 mmol/L (2.08 mg/dL) per 100 mg/d dietary cholesterol (P=.003); this accounted for about 75% of the rise in total cholesterol. HDL cholesterol concentrations increased by 0.015 mmol/L (0.57 mg/dL) per 100 mg/d dietary cholesterol (P<.04). There was a wide range of responses among the women. Plasma apoB levels increased significantly, 0.93 mg/dL per 100 mg/d dietary cholesterol (P=.025), whereas apoA-I levels tended to rise (1.35 mg/dL per 100 mg/d, P=.056). Increases in dietary cholesterol did not produce any observable effects on fasting plasma cholesteryl ester transfer protein levels and had no effect on the response to a standard high-fat formula. Although menstrual-cycle changes in plasma total, LDL, and HDL cholesterol levels were observed, the effects of the diets were similar in the follicular and luteal phases of the menstrual cycle. Additionally, despite changes associated with the menstrual cycle, within-subject variation in plasma total cholesterol was actually smaller in this study than in our study of young men.
Keywords: plasma lipids, dietary fatty acids, menstrual cycle, lipoproteins
In a recent study in healthy young men of the effects of increases in dietary cholesterol on plasma lipids and lipoproteins, we demonstrated that plasma total cholesterol levels increased by 0.038 mmol/L (1.47 mg/dL) for each additional 100 mg cholesterol added to the diet per day.1 This response was associated with LDL cholesterol (LDL-C); HDL cholesterol (HDL-C) tended to increase, but the response was small and did not reach statistical significance. Response to dietary cholesterol varied widely in our study group, and this variability did not appear to be related to apolipoprotein (apo) E genotype or levels of cholesteryl ester transfer protein (CETP). The mean responses to dietary cholesterol that we observed in these young healthy men were smaller than those reported by Keys et al2 and Hegsted et al3 but were consistent with other studies with a background diet low in total and saturated fat.4,5
Women were not included in our study or in the studies of several others.2–4 Zanni et al6 studied the effects of dietary cholesterol in only women, but in that investigation diets were fed for only 15 days.6 Both men and women have been included in several more recent studies that focused on the effects of dietary cholesterol,7–9 but only in one9 were the data analyzed by the sex of the subjects. Because of the paucity of data and increasing awareness of the importance of atherosclerotic cardiovascular disease in women, we conducted a dose-response study of dietary cholesterol in a group of healthy young women who were otherwise eating a diet recommended by the American Heart Association (AHA) and the National Cholesterol Education Program (NCEP).10 We measured fasting plasma lipid and lipoprotein levels during the three diets and determined the effects of increases in dietary cholesterol on the metabolism of chylomicrons and chylomicron remnants.
Methods
Subjects
Eighteen subjects were recruited from a population of 100 healthy female medical and dental students, aged 22 to 31 years, who had their blood cholesterol levels tested at the Health Sciences campus of Columbia University in New York City. Cholesterol levels were screened with an LDX lipid analyzer (Cholestech Corp) to obtain nonfasting, finger-stick blood cholesterol levels. Students with cholesterol concentrations between the 25th and 80th percentiles for age and sex were recruited. Individuals who were interested in participating in the study were invited to return after a 12-hour fast for the determination of plasma total cholesterol, triglyceride, and HDL-C levels from a venous blood sample.
The coordinating dietitian interviewed all the women before they were enrolled regarding daily dietary intake, ethanol consumption, and smoking habits. Individuals with extreme dietary habits or significant food intolerances were excluded. The investigators stipulated that participants had to be willing to comply with limitations on alcohol intake to no more than two liquor drinks or five beers per week. We also excluded volunteers who smoked or who followed vigorous exercise regimens. All the women were in good health; none had serious medical problems or were taking medications that might affect plasma lipid levels. None was taking oral contraceptives.
The experimental protocol was reviewed and approved by the Institutional Review Board at Columbia University. Informed consent was obtained from all participants before screening and again before enrollment in the study. Students did not receive monetary compensation for their participation in the study.
Protocol
This study was designed to determine the response of healthy young women to several levels of dietary cholesterol added to the Step 1 diet recommended by the AHA and the NCEP. A three-way crossover design was used, in which the subjects ate three separate diets, each for 8 weeks. In between each diet period, the subjects were on ad libitum diets for periods ranging from 4 to 6 weeks. The women were randomly assigned lo different diet sequences; all possible diet sequences were used. During the final 3 weeks of each diet period, blood samples were taken once per week for determination of lipids and lipoproteins, apoB and apoA-I, and CETP. All fasting samples were obtained between 8 and 9 AM after a 12-hour overnight fast. Buffy coats were isolated from these samples for determination of apoE genotypes. During the week of the middle third of each subject’s menstrual cycle, blood samples were obtained just before and 4 and 8 hours after ingestion of a standard high-fat formula that contained 53 g fat, 60 000 U vitamin A per meter squared of body surface, and 300 mg cholesterol. The subjects did not eat any other foods during the 8-hour period. This test was performed 1 or 2 days after the fasting blood sample was taken during the same week. The fat-formula lest at the end of each diet period allowed us to compare the long-term effects of different levels of dietary cholesterol on chylomicron and chylomicron remnant metabolism.
On Monday through Friday, lunches and dinners were eaten under supervision in the Columbia University–Bard Hall student dining facility. Monday breakfast was prepared and served by the Principal Investigator and the coordinating dietitian. All other breakfasts and evening snacks were packaged and distributed at the preceding dinner. Breakfasts and lunches for the weekend were packaged and distributed on Friday nights. Step 1 dietary guidelines, which stress low cholesterol and fat intakes, were given for two self-selected weekend dinners. Participants kept records of their weekend food consumption, and these records were reviewed by the dietitian every Monday at breakfast. These food records indicated that, in general, the self-selected weekend meals met the Step 1 dietary guidelines. Compliance with the diet was assessed and monitored daily by use of a self-administered form at each meal, by direct supervision in the dining hall, and by review of coded meal trays. The dietary staff and subjects met on a regular basis. The protocol was conducted in a double-blind fashion; only the dietary staff and the statisticians knew of the subjects’ dietary assignments.
Diets
All study meals were prepared with fresh ingredients in accordance with computer-analyzed recipes and menu plans. Nutrient analyses of the composition of the research diets were done with the Nutrition Data System program from the University of Minnesota (version 2.3). The Step 1 diet contained 55% of calories from carbohydrates, 15% from protein, and 30% from fat with 9% saturated, 14% monounsaturated, and 7% polyunsaturated fatty acids. The baseline content of cholesterol in all of the diets was 125 mg/d. The diets differed in their total cholesterol content, provided as 0, 1, and 3 grade A eggs per day. The eggs were added by weight (average wt, 50 g). All diets were similar in appearance. Daily calorie requirements were estimated with data on each subject’s height, weight, diet history, and physical activity pattern. The subjects were assigned to one of three caloric groups (1600, 1900, or 2200 kcal/d). Subjects were weighed every Monday before breakfast. Caloric intake level was adjusted either by shifting the subject to another caloric level or by providing low- or no-cholesterol snacks that met Step I dietary guidelines. The use of snacks allowed for flexibility in daily caloric intake without affecting diet composition.
A 2-week menu cycle was used throughout the study. The women were served a variety of foods, including beef, pork, poultry, fish, dairy products, fruits, vegetables, grains and grain products, legumes, and desserts. Grade A large eggs were used, which, according to US Department of Agriculture (USDA) Handbook 8, contain 215 mg cholesterol each. The number of eggs consumed was disguised within the overall diet by maintaining the egg bulk with egg substitutes (Eggbeaters, Fleischmann Inc). The fatty acid content remained the same despite increases in egg consumption. The eggs (and egg substitute) were served daily at lunch and dinner (except on Saturday and Sunday, when they were provided only with lunch). The basal level of cholesterol also was distributed between the lunch and dinner meals.
Food samples were prepared for compositional analyses at the end of each dietary period by use of USDA guidelines. Homogenates were prepared from a week’s meals from each of the three research diets. Six composite samples (two 1-week composites for each study diet) were sent to Hazelton Laboratories America for analyses of protein, carbohydrate, total fat, individual fatty acids, and cholesterol.
Laboratory
Sampling
Blood samples were drawn into tubes containing EDTA (1.0 mg/mL) for plasma or into empty tubes for serum. The samples were placed immediately on ice and centrifuged at 2000 rpm for 20 minutes at 4°C within 1 hour of sampling. Plasma and serum samples were stored in multiple aliquots at −70°C after the addition of aprotinin (Trasylol, FBA Pharmaceuticals) and azide. Samples for retinyl ester determination were protected from light and stored under nitrogen at −70°C.
Plasma Lipids
Cholesterol and triglyceride concentrations were measured by enzymatic methods with a Hitachi 705 automated spectrophotometer. HDL-C was measured by the same enzymatic method after precipitation of apoB-containing lipoproteins with magnesium and dextran by use of reagents supplied by Sigma Chemical Co.11 LDL-C was estimated by the Lipid Research Clinics method.12 These measurements were performed with frozen plasma samples after the study was completed. Our laboratory participates in the quality control program for lipid determination administered by the Centers for Disease Control and Prevention, Atlanta, Ga. The interassay coefficients of variation (CVs) were less than 3% for both cholesterol and triglyceride determinations.
Apoprotein Assays
Serum apoB and apoA-I levels were measured by immunonephelometry with a Beckman nephelometer. All samples from an individual were analyzed in the same assay. The interassay and intra-assay CVs were less than 5% for both assays. Plasma CETP levels were determined by a specific radioimmunoassay as previously described.13 The interassay and intra-assay CVs were less than 9%.
ApoE Genotyping
ApoE genotyping was performed by polymerase chain reaction (PCR) with the HhAI restriction enzyme.14 Briefly, leukocyte DNA was amplified by PCR with specifically synthesized oligonucleotide primers and Taq polymerase. The amplified apoE products were then digested with 5 U HhAI enzyme at 37°C for 4 hours, and the digest was subjected to electrophoresis on a 12% nondenaturing polyacrylamide gel for 3 hours at a constant current of 10 mA. The gels were treated with ethidium bromide for 10 to 15 minutes, and the DNA fragments were visualized by UV illumination. DNA fragments of known size were used as markers.
Retinyl Ester Determination
Plasma retinol and retinyl palmitate levels were measured by reversed-phase high-performance liquid chromatography with a procedure similar to that described by Bieri et al.15 This method uses an internal standard technique for the calculation of retinol and retinyl ester levels. The within-assay and between-assay CVs for retinol and retinyl ester determinations were less than 7%.
Statistical Analysis
Menstrual cycle effects on total cholesterol, LDL-C, and HDL-C were observed. For total and LDL-C, an ANOVA was done to estimate a common menstrual cycle-related slope for all subjects (which allowed each woman’s level on each diet to be different). For HDL-C, the ANOVA estimated two common slopes, one for the first 14 days of the menstrual cycle and a second for the remainder of the menstrual cycle. For all three responses, the ANOVA also provided levels adjusted to the middle of the menstrual cycle. All statistical analyses of the lipid responses (slopes) were done with the adjusted values. Each response variable was analyzed for seasonal and carryover effects by repeated-measures ANOVA, with each subject’s period as the repeated-measure factor and diet and preceding diet as separate effects. No carryover or seasonal effects were found.
Dose response to dietary cholesterol was analyzed by linear regression. For each subject, the response variable (eg, total cholesterol) at the three levels of dietary cholesterol was fitted by a straight-line function of dietary cholesterol, and a slope was determined. The adequacy of each linear model was determined by testing the significance of a quadratic term. After the linear model was demonstrated as adequate, the mean of the slopes of all subjects was determined and tested to see whether it was significantly different from zero (one-sample t test). With no missing data, this is equivalent to fitting a single slope to all subjects while allowing each subject to have a different y intercept. We also tested the model that Keys et al2 suggested, in which the response to dietary cholesterol is linear with the square root of dietary cholesterol rather than the absolute values.
The post–fat formula response was calculated as the mean of the values determined after the meal, corrected for the baseline value. The responses during the different diets were analyzed by repeated-measures ANOVA.
Results
Table 1 depicts the mean characteristics and baseline lipids of the 13 subjects who completed the study. The subjects were young, nonobese women who had normal plasma lipid levels on a free-living diet. Body weight remained constant throughout the study. Two of the original 18 women withdrew from the study after the first diet period, and three more withdrew after the second diet period. Four women withdrew, unable to come for meals regularly because of schedule changes. One subject was forced to withdraw because of a new, unrelated health problem.
Table 1.
Subject Characteristics
| Characteristic | Mean±SD | Range |
|---|---|---|
| Age, y | 23.5±1.9 | 21–28 |
| Weight, kg | 55.5±6.0 | 47–65 |
| BMI, kg/m2 | 20.8±1.5 | 18.9–23.0 |
| Total cholesterol, mmol/L | 4.06±0.33 | 3.59–4.70 |
| Triglycerides, mmol/L | 0.86±0.23 | 0.61–1.30 |
| LDL cholesterol, mmol/L | 2.38±0.33 | 1.78–2.85 |
| HDL cholesterol, mmol/L | 1.51±0.27 | 1.16–1.83 |
BMI indicates body mass index. Plasma lipid values were determined from a single blood sample obtained at the time of recruitment for the study before subjects began the diets. To convert millimoles per liter cholesterol to milligrams per deciliter, multiply by 38.67. To convert millimoles per liter triglycerides to milligrams per deciliter, multiply by 88.57.
The analyzed and calculated compositions of the three diets are presented in Table 2. The compositions of the diets, excluding cholesterol, were essentially the same in all three periods. As we had planned, the quantities of total fat, carbohydrate, and protein as well as the proportions of fat as saturated, monounsaturated, and polyunsaturated fatty acids met the guidelines of the Step 1 diet. The total fat content of each diet was slightly lower than the goal of 30%. The reduction in total fat was the result of slightly lower levels of saturated and monounsaturated fatty acids compared with calculated values. As expected, the amount of cholesterol in each diet varied according to the number of eggs present. On average, each egg increased the analyzed cholesterol content of the diet by about 177 mg, which was essentially the same as we found in our previous study in men.1 This was approximately 15% less cholesterol, by analysis, than was expected from calculations based on USDA Food Composition Tables. The difference between the calculated and the analytic values might have resulted from a systematic error in the USDA tables or in our method of compositing samples for analysis. Our calculated responses to increases in dietary cholesterol (based on each additional 100 mg/d) would, however, have been 15% lower if we had used the accepted value for egg cholesterol content (215 mg per egg yolk) rather than the measured values.
Table 2.
Composition of the Diets
| Eggs, No. | 0 | 1 | 3 |
|---|---|---|---|
| Cholesterol, mg/d* | 108 (125) | 277 (340) | 667 (770) |
| Total fat, %† | 28.0 (30) | 28.0 (30) | 28.4 (30) |
| Saturated | 8.3 (9) | 8.2 (9) | 8.4 (9) |
| Monounsaturated | 12.7 (14) | 12.6 (14) | 13.0 (14) |
| Polyunsaturated | 7.1 (7) | 7.2 (7) | 7.2 (7) |
| Carbohydrate, %† | 56.8 (55) | 55.8 (55) | 57.2 (55) |
| Protein, %† | 15.2 (15) | 15.1 (15) | 15.4 (15) |
Food samples were prepared and analyzed as described in “Methods.” Analyzed values are followed in parentheses by calculated values obtained with the Nutrition Data System, version 2.3, from the University of Minnesota.
Calculated amounts of dietary cholesterol in the one-egg and three-egg diets were based on the assumption that one egg contained 215 mg cholesterol.
% indicates percent total caloric intake.
Analysis of each woman’s three weekly lipid values during each period relative to her menstrual cycle indicated that total cholesterol decreased during the cycle (Fig 1). Overall, total cholesterol and LDL-C fell by 0.051 mmol/L per week (1.99 mg/dL, P<.02) and 0.064 mmol/L per week (2.48 mg/dL, P<.001), respectively, during each menstrual cycle. We also observed that HDL-C levels were affected by the menstrual cycle; values increased by 0.060 mmol/L per week (2.3 mg/dL, P<.001) during the first half of the cycle and decreased by 0.050 mmol/L per week (1.94 mg/dL, P<.01) during the second half. Triglyceride concentrations did not appear to be affected by the menstrual cycle. The changes in total cholesterol, LDL-C, and HDL-C levels observed during the menstrual cycle were not, however, affected by the level of dietary cholesterol.
Fig 1.

Scatterplots showing plasma total cholesterol (top), LDL cholesterol (middle), and HDL cholesterol (bottom) concentrations during the menstrual cycle. Blood was obtained for plasma lipid determination from all women studied once a week during the final 3 weeks of each diet period. Data are plotted as deviations from each subject’s mean for one diet period; data from all three diet periods are plotted for each woman. Because menstrual cycle effects were similar during all three diets, data are combined in one plot. Total and LDL cholesterol levels fell by 0.051 mmol/L (P<.02) and 0.064 mmol/L per week (P<.001), respectively, during the menstrual cycle. HDL cholesterol concentrations rose by 0.060 mmol/L per week (P<.001) and fell by 0.050 mmol/L per week (P<.01) during the first and second halves of the cycle, respectively. Triglyceride levels were unaffected by the menstrual cycle (data not shown). To convert millimoles per liter cholesterol to milligrams per deciliter, multiply by 38.67.
The dose responses of fasting plasma total cholesterol, LDL-C, and HDL-C concentrations (adjusted to mid-cycle) to increasing dietary cholesterol are depicted in Fig 2. There were statistically significant linear increases in plasma total cholesterol, LDL-C, and HDL-C with increases in dietary cholesterol intake. Plasma total cholesterol increased by 0.073 mmol/L (2.81 mg/dL) for each additional 100 mg/d dietary cholesterol (P=.001; 95% confidence interval [CI], 0.035 to 0.110 mmol/L per 100 mg/d dietary cholesterol). About 75% of the rise in total plasma cholesterol was accounted for by the response in LDL-C, which increased by 0.054 mmol/L (2.08 mg/dL) per 100 mg/d dietary cholesterol (P=.003; 95% CI, 0.022 to 0.085 mmol/L).
Fig 2.

Line graphs showing responses of plasma total (top), LDL (middle), and HDL (bottom) cholesterol concentrations to increases in dietary cholesterol intake. Individual points are mean±SD for the 13 women on each diet. The regression line is the mean of the individual regression lines for each of the 13 subjects. Plasma total, LDL, and HDL cholesterol levels increased by 0.073 mmol/L (P=.001), 0.054 mmol/L (P=.003), and 0.015 mmol/L (P<.04), respectively, for each additional 100 mg/d of dietary cholesterol. To convert millimoles per liter to milligrams per deciliter, multiply by 38.67. Chol indicates cholesterol. Cholesterol values are mean±SD.
HDL-C increased significantly, by 0.015 mmol/L (0.57 mg/dL) per 100 mg/d dietary cholesterol (P<.04; 95% CI, 0.001 to 0.029 mmol/L per 100 mg/d dietary cholesterol). No significant response could be demonstrated by linear regression analysis for plasma triglyceride concentrations (data not shown). The mean±SD for each lipid on each of the three diets is presented in Table 3. The values in Table 3 are adjusted for menstrual cycle variations.
Table 3.
Mean Plasma Lipids at Each Level of Dietary Cholesterol
| Dietary Cholesterol, mg/d* | Plasma Lipids, mmol/L
|
|||
|---|---|---|---|---|
| Total C | LDL-C | Triglycerides | HDL-C | |
| 108 | 4.11 (0.31) | 2.18 (0.34) | 1.02 (0.17) | 1.51 (0.27) |
| 277 | 4.27 (0.39) | 2.28 (0.42) | 1.06 (0.21) | 1.55 (0.29) |
| 677 | 4.52 (0.42) | 2.48 (0.39) | 1.05 (0.16) | 1.60 (0.26) |
C indicates cholesterol. Values are mean (±SD) for all 13 subjects during each of the three diets. To convert millimoles per liter cholesterol to milligrams per deciliter, multiply by 38.67. To convert millimoles per liter triglycerides to milligrams per deciliter, multiply by 88.57.
Measured amounts of dietary cholesterol on each of the three diets.
When we fitted the total cholesterol, LDL-C, and HDL-C responses with the square-root function used by Keys et al2 (which fits a curvilinear response), the fits were as good as with the linear models but no better. The estimated slopes for response were 0.027 mmol/L per square root of milligrams dietary cholesterol per day for total cholesterol (95% CI, 0.013 to 0.041), 0.019 mmol/L per square root of milligrams dietary cholesterol per day for LDL-C (95% CI, 0.008 to 0.031), and 0.0054 mmol/L per square root of milligrams dietary cholesterol per day for HDL-C (95% CI, 0.002 to 0.011). The root-mean-squared error for total cholesterol (a measure of the variability in the data after the linear model was fitted) was 0.148 mmol/L with absolute levels and 0.142 mmol/L with the square roots of dietary cholesterol; for LDL-C, 0.155 and 0.150 mmol/L, respectively; and for HDL-C, 0.084 and 0.083 mmol/L, respectively. Thus, both models fit our data equally well.
There was a wide distribution of individual dose responses for total cholesterol, LDL-C, and HDL-C (Fig 3). To facilitate visual inspection of the data, the total cholesterol, LDL-C, and HDL-C responses of each subject were adjusted to values corresponding to the mean concentrations for all subjects on the zero-egg diet. The slopes describing the response of total plasma cholesterol to increasing dietary cholesterol ranged from −0.020 to +0.176 mmol/L per 100 mg/d dietary cholesterol, whereas those describing the LDL-C response ranged from −0.041 to +0.140 mmol/L per 100 mg/d dietary cholesterol. HDL-C also exhibited a wide range of responses to dietary changes; individual responses varied from −0.022 to +0.047 mmol/L per 100 mg/d dietary cholesterol.
Fig 3.
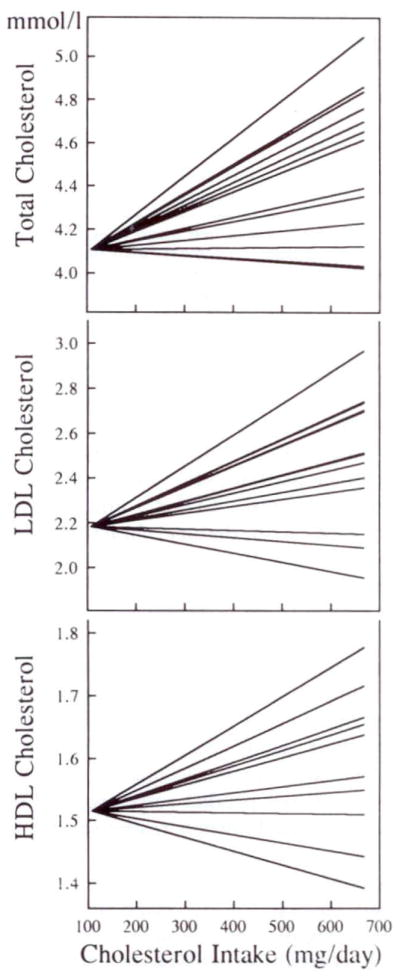
Graphs showing individual dose-response data for total (top), LDL (middle), and HDL (bottom) cholesterol levels to increases in dietary cholesterol. Each line is derived from regression analysis of each subject’s cholesterol levels on the three diets. To depict the range of responses more clearly, all individual responses have been adjusted to an initial level for total, LDL, and HDL cholesterol that represents the mean level of each variable for the entire group on the zero-egg diet. The results indicate that there was a wide range of mostly positive responses to dietary cholesterol.
Since we observed each woman over 3 consecutive weeks on each diet, at least one observation was made in each half of the menstrual cycle per diet. We analyzed data from each half of the cycle separately with no adjustment for menstrual cycle variation. The slopes of plasma total cholesterol, LDL-C, and HDL-C in response to increases in dietary cholesterol were very similar in both halves of the menstrual cycle (Table 4), and triglyceride values were unaffected by diet in both halves of the cycle (data not shown).
Table 4.
Effect of the Menstrual Cycle on Cholesterol Responses at Each Level of Dietary Cholesterol
| Cholesterol | Slope
|
Difference | |
|---|---|---|---|
| First Half | Second Half | ||
| Total | 0.064 ± 0.079 (P<.006) | 0.073 ± 0.059 (P<.001) | 0.008 ± 0.053 (P=NS) |
| LDL | 0.048 ± 0.067 (P<.015) | 0.055 ± 0.051 (P<.003) | 0.007 ± 0.055 (P=NS) |
| HDL | 0.017 ± 0.020 (P<.02) | 0.015 ± 0.028 (P<.04) | −0.002 ± 0.024 (P=NS) |
Each woman’s level of dietary cholesterol from the first and second halves of her menstrual cycle during each diet were analyzed separately to generate dose responses to increases in dietary cholesterol. Slopes are in millimoles per liter per 100 mg dietary cholesterol per day. Values are mean ± SD. Difference is the mean ± SD of the individual differences between each woman’s slopes during the first and the second halves of her menstrual cycle.
A search for baseline and study variables that might predict an individual subject’s response indicated that no significant relations existed between response and baseline lipids at the time of recruitment when the subjects were eating their usual, free-living diets. Participant characteristics, such as body mass index and apoE genotype, also did not predict response, and no significant relations existed between response and lipid concentrations during the zero-egg diet.
Fasting serum apoB levels increased with consumption of dietary cholesterol; the mean slope of the rise in apoB was 0.93 mg/dL per 100 mg/d dietary cholesterol (P=.025). A correlation existed (r=.80, P<.001) between individual slopes for LDL-C and serum apoB slopes (Fig 4). ApoA-I concentrations also tended to rise as egg intake increased from 0 to 1 to 3 eggs per day. The mean slope of the change in apoA-I concentration was 1.35 mg/dL per 100 mg/d cholesterol (P=.056), and the slopes for apoA-I levels correlated with the slopes for HDL-C (r=.80, P<.001).
Fig 4.
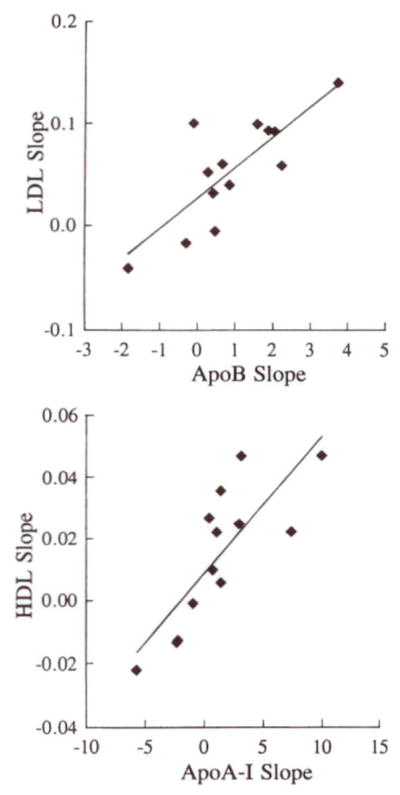
Scatterplots showing correlations between LDL cholesterol and apolipoprotein (apo) B slopes (top, R=.80) and between HDL cholesterol and apoA-l slopes (bottom, R=.80). Each subject’s change in lipoprotein cholesterol between the 0- and 3-egg diets was related to the change in the appropriate apolipoprotein. The results suggest that increases in LDL and possibly HDL cholesterol levels were associated with increases in the number of particles in each lipoprotein class.
We did not find a significant dose response in CETP to increases in dietary cholesterol intake. The mean values for CETP were 2.49±0.75, 2.36±0.78, and 2.56±0.73 μg/mL on the 0-, 1-, and 3-egg diets, respectively.
When the women consumed a standard liquid formula containing 53 g fat and 60 000 U vitamin A per meter squared, the areas above baseline (defined as the fasting plasma level that day) for triglycerides and retinyl palmitate did not differ for the three diets (Fig 5). The integrated 8-hour responses of total cholesterol, LDL-C, and HDL-C were small, as expected after ingestion of 300 mg cholesterol (Table 5). In response to ingestion of the large triglyceride load, LDL-C and HDL-C levels generally fell, whereas the total cholesterol responses (which would include VLDL cholesterol [not measured] in addition to LDL-C and HDL-C) hovered around zero. The mean responses for total cholesterol, LDL-C, and HDL-C, corrected for fasting levels, were not different for the three diets. However, the absolute levels of total cholesterol, LDL-C, and HDL-C during the 8 hours were consistent with the differences in fasting levels for the three diets (Fig 6).
Fig 5.

Graphs showing post–fat formula plasma lipid responses to increasing dietary cholesterol. Retinyl palmitate (top) and total plasma triglyceride (bottom) levels were measured just before and 4 and 8 hours after a standard high-fat formula providing 53 g fat per meter squared, 60 000 U vitamin A per meter squared, and 300 mg cholesterol. These tests were performed on all 13 subjects on each of the three diets. Increases in dietary cholesterol intake had no effect on the response to this high-fat meal after correcting for fasting levels.
Table 5.
Post-Fat Formula Cholesterol and Triglyceride Responses at Each Level of Dietary Cholesterol
| Dietary Cholesterol, mg/d* | Cholesterol Level, mmol/L
|
||
|---|---|---|---|
| Total | LDL | HDL | |
| 108 | −0.01 (0.17) | −0.26 (0.16) | −0.04 (0.06) |
| 277 | 0.02 (0.19) | −0.24 (0.24) | −0.04 (0.09) |
| 677 | 0.06 (0.18) | −0.24 (0.18) | 0.00 (0.08) |
Post–fat formula (4 and 8 hours after fat load) responses were calculated as the mean of the values obtained during each test corrected for baseline (premeal) value. All 13 subjects had a post–fat formula test during each of the three diets. Data are presented as mean (± SD)for 13 subjects on each diet. Negative values indicate a decrease in a level during the test. None of the responses on any diet was significantly different from any other response. To convert millimoles per liter cholesterol to milligrams per deciliter, multiply by 38.67.
Measured amounts of dietary cholesterol during each of the three diets.
Fig 6.

Graphs showing post–fat formula plasma lipid responses to increases in dietary cholesterol. Total plasma (top), LDL (middle), and HDL (bottom) cholesterol levels were measured just before and 4 and 8 hours after subjects were fed a standard high-fat formula providing 53 g fat per meter squared, 60 000 U vitamin A per meter squared, and 300 mg cholesterol. This test was performed on all 13 subjects during each of the three diets. Lines represent the diets containing 0, 1, and 3 eggs per day, as shown in Fig 5. Although increases in dietary cholesterol intake had no effect on the response to this high-fat meal, after correcting for fasting levels (see Table 4), the absolute difference in the levels of cholesterol on the different egg intakes can be seen.
In our small group of subjects, eight women were either ε3/2 or ε3/3 and five women were ε4/3 or ε4/4. We did not see differences in the response of any lipid or lipoprotein fraction between these two subgroups of subjects.
Discussion
The results of this study indicate that healthy young women, like their male counterparts,1 respond to higher dietary cholesterol intakes by modestly increasing their plasma total cholesterol levels in a linear fashion. However, there are important differences between our previous observations in young men and those we now present for young, healthy women. First, the slope of the mean response of total plasma cholesterol in the women was almost twice what we observed in the men: 0.073 versus 0.038 mmol/L per 100 mg/d dietary cholesterol. Second, unlike the men, in whom the rise in total cholesterol was accounted for almost completely by a rise in LDL-C (0.036 mmol/L per 100 mg/d dietary cholesterol), LDL-C accounted for only 75% of the rise in total cholesterol in the women (0.054 mmol/L per 100 mg/d dietary cholesterol). Although not statistically significant, the mean LDL-C slope in the women was about 50% greater than that in the men,1 which suggests a greater response to dietary cholesterol in young women. This apparent difference, however, may derive from the difference in caloric intake between the two groups; the men consumed almost 40% more calories (average caloric intake in the women was 1800 kcal/d versus 2450 kcal/d in the men). If the LDL-C slopes we observed are expressed as millimoles per liter per 100 milligrams dietary cholesterol per 1000 kcal/d, the mean slope is approximately 0.097 for the women and about 0.088 for the men. Thus, after adjusting for caloric intake, which in turn adjusts for weight, we see no clear difference between the responses of the young men and women that we studied. HDL-C rose significantly in our group of women (0.015 mmol/L per 100 mg/d dietary cholesterol) and accounted for the rest of the rise in total cholesterol levels. In contrast, we did not see a consistently significant effect of increases in dietary cholesterol on HDL-C concentrations in healthy young men.1
Most published studies of the effects of dietary cholesterol on plasma lipids have involved only men. Zanni et al6 investigated the effect of increasing dietary cholesterol in nine women. That study used four diet periods of 15 days each to examine the effects of 130 and 875 mg cholesterol on the background of 31%-fat diets with a ratio of polyunsaturated to saturated fat (P/S) of either 0.64 or 2.14. During diets with both lower and higher P/S ratios, LDL-C increased approximately 2 mg/dL for each additional 100 mg/d dietary cholesterol. Thus, Zanni et al6 observed responses to increasing dietary cholesterol similar to those we saw in the present study with a background P/S ratio of 0.87. HDL-C responses did appear to be affected by the P/S ratio in the study by Zanni et al.6 There was no change in HDL-C associated with the addition of 745 mg cholesterol to the diet in the presence of a P/S ratio of 0.64; in contrast, increases in dietary cholesterol on the background of a P/S ratio of 2.14 resulted in a rise in HDL-C of about 1.5 mg/dL per 100 mg/d dietary cholesterol. The larger HDL response to increased dietary cholesterol during the high P/S diet was most likely a concomitant of the lower HDL-C concentrations associated with the very large quantity of dietary polyunsaturated fatty acids used by Zanni et al.6 Our diets, with a P/S ratio of about 0.87, appear to have elicited HDL responses between those seen by Zanni et al.6
Several studies in which dietary cholesterol differed significantly between test diets included both men and women,7–9 but separate analyses were performed for men and women only in the study by Clifton and Nestel.9 They compared the effects of increasing both dietary fat and cholesterol on plasma lipids and lipoproteins in 25 women and 26 men; their results provide support for our findings. When 31 g fat (mostly saturated) and 650 mg cholesterol were added to a low-fat, low-cholesterol diet for 2 weeks, total plasma cholesterol increased by 0.056 and 0.060 mmol/L per 100 mg/d dietary cholesterol in the women and men, respectively. However, when 14 women under the age of 50 years were compared with 13 men of similar age, the respective increases in total cholesterol levels were 0.056 and 0.033 mmol/L per 100 mg/d dietary cholesterol. Thus, among the younger subjects, the response among women exceeded that among men.9 These results are similar to our findings in the present report and in our previous study of young men,1 although we noted a larger response among our female subjects. Also in accordance with our findings, Clifton and Nestel9 demonstrated a greater increase in HDL-C (particularly in HDL2) in women versus men. This finding was consistent in both the age groups they analyzed. Furthermore, their finding of a rise in HDL-C of 0.024 mmol/L per 100 mg/d dietary cholesterol in younger women is very close to our slope of 0.017 in the present report.
In the young women we studied, the slope of the rise in total cholesterol was considerably smaller than those reported by Hegsted et al3 or Mattson et al.16 Although our present results for the response of total cholesterol are similar to those reported by Keys et al,2 LDL-C accounted for only two thirds of the increase in total cholesterol in the women we studied. Since Keys et al2 studied only men, in whom increases in LDL-C account for essentially all the rise in total cholesterol,1 we believe that the LDL-C response among the women in our study was reduced compared with the likely LDL-C response among the men studied by Keys et al.2 Thus our present results are consistent with our previous observations in young men,1 in whom responses to dietary cholesterol were less than previously reported.2,3,16 We believe that the differences between those previous results and ours derive from the lower-fat, lower-saturated fat content of our diet. This hypothesis is supported by the results of the Faribault second study,4 which demonstrated a clear inverse relation between dietary saturated fat content and response to dietary cholesterol. In a smaller metabolic study, Schonfeld et al5 also demonstrated a strong interaction between the P/S ratio of the diet and response to dietary cholesterol. Therefore, although some investigators have not observed interactions between dietary fatty acid composition and response to cholesterol,6,17,18 we believe that the relatively low-fat, low–saturated fat content of our Step 1 background diet is the reason for the differences between our results and those of Keys et al,2 Hegsted et al,3 and Mattson et al.16
In the present study, as in our previous report on young men,1 we found that increases in LDL-C correlated with increases in serum apoB concentrations, indicating that the number of LDL particles had increased. Increases in LDL particle number can result from both increases in LDL production and decreases in receptor-mediated uptake of LDL; both of these effects have been demonstrated in studies of the effects of dietary cholesterol.19,20 Indeed, the variability of response to dietary cholesterol is likely to derive from interindividual differences in LDL-C production and clearance.20,21 We also demonstrated that the rise in HDL-C observed in our female subjects as dietary cholesterol increased was associated with a rise in serum apoA-I concentrations. Zanni et al6 also reported an increase in apoA-I concentrations among women on a diet with a high P/S ratio and high in cholesterol.
We did not see an effect of increases in dietary cholesterol on plasma CETP levels. In our previous study, CETP levels appeared to increase modestly at the highest level of dietary cholesterol (four eggs per day)1; in the present study, subjects ingested a maximum of three eggs per day. Martin et al22 reported increases in CETP levels in young men eating high-cholesterol diets. Our results suggest either a sex difference in the CETP response to dietary cholesterol or a dose response; we fed lower levels of cholesterol in the present study compared with both our previous work in men1 and the study by Martin et al.22
Numerous studies in animals fed large quantities of dietary cholesterol suggest that chylomicron and/or chylomicron remnant metabolism can be affected. Plasma responses of triglyceride (a measure of chylomicron metabolism) and retinyl palmitate (a measure of chylomicron remnant metabolism) to a high-fat meal did not differ during the three diets in the present study. This result is consistent with our previous findings in young men.1 Our results suggest that diets with as many as three to four eggs per day do not significantly alter postprandial lipoprotein metabolism. These results are compatible with the studies by Weintraub et al23 and Eriksson et al24 in which marked differences in LDL-C receptor function did not affect chylomicron removal. As in our study of young men, plasma levels of total cholesterol, LDL-C, and HDL-C in the women throughout the 8-hour post–fat load period paralleled their fasting levels on the three diets containing various numbers of eggs.
Women have often been excluded from metabolic studies of lipids and lipoproteins because of the effects of the menstrual cycle. Our finding of a decline in total cholesterol and LDL-C over the course of the menstrual cycle is consistent with previous studies by Kim and Kalkhoff,25 who reported values for both parameters, and Jones et al,26 Lussier-Cacan et al,27 Tangney et al,28 and De Leon et al,29 who reported only total cholesterol concentrations. Lebech and Kjoer,30 Woods et al,31 and Karpanou et al32 did not, however, observe changes in total and LDL-C during the menstrual cycle in healthy women. HDL levels appeared to be stable throughout the menstrual cycle in most of the studies in which they were determined.23,30–33 In contrast, we observed a rise in HDL-C during the follicular phase and a decline during the luteal phase of the menstrual cycle. This biphasic pattern of HDL-C change is consistent with the rise and fall of plasma estrogen levels during the menstrual cycle. Triglyceride levels appeared to increase at the time of ovulation in several studies25,27,28,31; we did not observe such a trend. Additionally, like Berlin et al,33 who observed similar reductions in total cholesterol and LDL-C during the menstrual cycle in women eating four different diets, we found no interaction between diet and menstrual cycle.
Despite changes in total cholesterol, LDL-C, and HDL-C concentrations associated with the menstrual cycle, we found that the within-subject variation was smaller in the women we studied than in the men.1 The root-mean-squared error for total cholesterol was 0.148 mmol/L in the women and 0.257 mmol/L in the men.1 This difference could be seen even if the data for the women were not adjusted for menstrual cycle variation. Similar differences for within-subject variation between women and men were seen for LDL-C and HDL-C. Finally, we found that the slopes of response of total cholesterol, LDL-C, and HDL-C to increasing dietary cholesterol were not different in the first versus the second half of the menstrual cycle. Thus our results and those from most previous studies indicate that menstruating women should be included in studies of the effects of diet on plasma lipids and lipoproteins.
Acknowledgments
This work was supported by grants HL-36000, HL-21006, and RR-645 from the National Institutes of Health and a grant from the American Egg Board. Support was also provided by Fleischmann Inc. The authors wish to thank Colleen Ngai, Nelson Fontanez, Jeffrey Jones, and Minnie Myers for their excellent technical assistance; Colleen Johnson, Roberta Hole-man, and the diet staff for their outstanding work in preparing and serving meals; and Ludmila Karameros for assistance with the manuscript. The assistance of the Irving Center for Clinical Research is greatly appreciated.
References
- 1.Ginsberg UN, Karmally W, Siddiqui M, Holleran S, Tall AR, Rumsey SC, Deckelbaum RJ, Blaner WS, Ramakrishnan R. A dose-response study of the effects of dietary cholesterol on fasting and postprandial lipid and lipoprotein metabolism in healthy young men. Arterioscler Thromb. 1994;14:576–586. doi: 10.1161/01.atv.14.4.576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Keys A, Anderson JT, Grande F. Serum cholesterol response to changes in the diet. II: the effect of cholesterol in the diet. Metabolism. 1965;14:759–765. doi: 10.1016/0026-0495(65)90002-8. [DOI] [PubMed] [Google Scholar]
- 3.Hegsted DM, McGandy RB, Myers ML, Stare FJ. Quantitative effects of dietary fat on serum cholesterol in man. Am J Clin Nutr. 1965;17:281–295. doi: 10.1093/ajcn/17.5.281. [DOI] [PubMed] [Google Scholar]
- 4.National Diet-Heart Study Research Group. Faribault second study. National Diet-Heart Study final report. Circulation. 1968;37(suppl I):I-260–I-274. [PubMed] [Google Scholar]
- 5.Schonfeld G, Patsch W, Rudel LL, Nelson C, Epstein M, Olson RE. Effects of dietary cholesterol and fatty acids on plasma lipoproteins. J Clin Invest. 1982;69:1072–1079. doi: 10.1172/JCI110542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Zanni EE, Zannis VI, Blum CB, Herbert PN, Breslow JL. Effect of egg cholesterol and dietary fats on plasma lipids, lipoproteins, and apoproteins of normal women consuming natural diets. J Lipid Res. 1987;28:518–527. [PubMed] [Google Scholar]
- 7.Edington JD, Geekie M, Carter R, Benfield L, Ball M, Mann J. Serum lipid response to dietary cholesterol in subjects fed a low-fat, high-fiber diet. Am J Clin Nutr. 1989;50:58–62. doi: 10.1093/ajcn/50.1.58. [DOI] [PubMed] [Google Scholar]
- 8.Edington J, Geekie M, Carter R, Benfield L, Fisher K, Ball M, Mann J. Effect of dietary cholesterol on plasma cholesterol concentration in subjects following reduced fat, high fibre diet. Br Med J. 1987;294:333–336. doi: 10.1136/bmj.294.6568.333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Clifton PM, Nestel PJ. Influence of gender, body mass index, and age on response of plasma lipids to dietary fat plus cholesterol. Arterioscler Thromb. 1992;12:955–962. doi: 10.1161/01.atv.12.8.955. [DOI] [PubMed] [Google Scholar]
- 10.The Expert Panel. Report of the National Cholesterol Education Program Expert Panel on detection, evaluation, and treatment of high blood cholesterol in adults. Arch Intern Med. 1988;148:36–69. [PubMed] [Google Scholar]
- 11.Warnick GR, Nguyen T, Albers AA. Comparison of improved precipitation methods for quantification of high-density lipoprotein cholesterol. Clin Chem. 1985;31:217–222. [PubMed] [Google Scholar]
- 12.Lipid Research Clinic Program. Lipid and Lipoprotein Analysis. DHEW Publication No. 9NIH0. Vol. 1. Washington, DC: National Institutes of Health; 1974. Manual of Laboratory Operations; pp. 75–628. [Google Scholar]
- 13.Marcel YL, McPherson R, Hogue M, Czarnecka H, Zawadzki Z, Weech PK, Whitlock ME, Tall AR, Milne RW. Distribution and concentration of cholesteryl ester transfer protein in plasma of normolipemic subjects. J Clin Invest. 1990;85:10–17. doi: 10.1172/JCI114397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hixson J, Vernier D. Restriction isotyping of human apolipoprotein E by gene amplification and cleavage with HhAI. J Lipid Res. 1991;31:545–548. [PubMed] [Google Scholar]
- 15.Bieri JG, Tolliver TJ, Catignani GL. Simultaneous determination of a-tocopherol and retinol in plasma or red cells by high-pressure liquid chromatography. Am J Clin Nutr. 1979;32:2143–2149. doi: 10.1093/ajcn/32.10.2143. [DOI] [PubMed] [Google Scholar]
- 16.Mattson FH, Erickson BA, Kligman AM. Effects of dietary cholesterol on serum cholesterol in man. Am J Clin Nutr. 1972;25:589–594. doi: 10.1093/ajcn/25.6.589. [DOI] [PubMed] [Google Scholar]
- 17.McNamara DJ, Kolb R, Parker TS, Batwin H, Samuel P, Brown CD, Ahrens EH., Jr Heterogeneity of cholesterol homeostasis in man. J Clin Invest. 1987;79:1729–1739. doi: 10.1172/JCI113013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kestin M, Clifton PM, Rouse IL, Nestel PJ. Effect of dietary cholesterol in normolipidemic subjects is not modified by nature and amount of dietary fat. Am J Clin Nutr. 1989;50:528–532. doi: 10.1093/ajcn/50.3.528. [DOI] [PubMed] [Google Scholar]
- 19.Packard CJ, McKinney L, Carr K, Shepherd J. Cholesterol feeding increases low density lipoprotein synthesis. J Clin Invest. 1983;72:45–51. doi: 10.1172/JCI110983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Mistry P, Miller NE, Laker M, Hazzard WR, Lewis B. Individual variation in the effects of dietary cholesterol on plasma lipoproteins and cellular cholesterol homeostasis in man. J Clin Invest. 1981;67:493–502. doi: 10.1172/JCI110058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ginsberg H, Le N-A, Mays C, Gibson J, Brown WV. Lipoprotein metabolism in non-responders to increased dietary cholesterol. Arteriosclerosis. 1981;1:463–470. doi: 10.1161/01.atv.1.6.463. [DOI] [PubMed] [Google Scholar]
- 22.Martin LJ, Connelly PW, Nancoo D, Wood N, Zhang ZJ, Maguire G, Quinet E, Tall AR, Marcel YL, McPherson R. Cholesteryl ester transfer protein and high density lipoprotein responses to cholesterol feeding in men: relationship to apolipoprotein E genotype. J Lipid Res. 1993;34:437–446. [PubMed] [Google Scholar]
- 23.Weintraub MS, Eisenberg S, Breslow JL. Different patterns of postprandial lipoprotein metabolism in normal, type IIa, type III, and type IV hyperlipoproteinemic individuals: effects of treatment with cholestyramine and gemfibrozil. J Clin Invest. 1987;79:1110–1119. doi: 10.1172/JCI112926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Eriksson M, Angelin B, Henriksson P, Ericsson S, Vitols S, Berglund L. Metabolism of lipoprotein remnants in humans. Arterioscler Thromb. 1991;11:827–837. doi: 10.1161/01.atv.11.4.827. [DOI] [PubMed] [Google Scholar]
- 25.Kim H-J, Kalkhoff RK. Changes in lipoprotein composition during the menstrual cycle. Metabolism. 1979;28:663–668. doi: 10.1016/0026-0495(79)90020-9. [DOI] [PubMed] [Google Scholar]
- 26.Jones DY, Judd JT, Taylor PR, Campbell WS, Nair PP. Menstrual cycle effect on plasma lipids. Metabolism. 1988;37:1–2. doi: 10.1016/0026-0495(88)90020-0. [DOI] [PubMed] [Google Scholar]
- 27.Lussier-Cacan S, Xhignesse M, Desmarais J-L, Davignon J, Kafrissen ME, Chapdelaine A. Cyclic fluctuations in human serum lipid and apolipoprotein levels during the normal menstrual cycle: comparison with changes occurring during oral contraceptive therapy. Metabolism. 1991;40:849–854. doi: 10.1016/0026-0495(91)90014-n. [DOI] [PubMed] [Google Scholar]
- 28.Tangney C, Brownie C, Wu S-M. Impact of menstrual periodicity on serum lipid levels and estimates of dietary intakes. J Am Coll Nutr. 1991;10:107–113. doi: 10.1080/07315724.1991.10718133. [DOI] [PubMed] [Google Scholar]
- 29.De Leon RG, Austin KL, Richards L, Guerrero F. Lipid and hormonal profile of Panamanian women during the menstrual cycle. Int J Gynecol Obstet. 1992;39:219–226. doi: 10.1016/0020-7292(92)90660-b. [DOI] [PubMed] [Google Scholar]
- 30.Lebech A-M, Kjoer A. Lipid metabolism and coagulation during the normal menstrual cycle. Horm Metab Res. 1989;21:445–448. doi: 10.1055/s-2007-1009258. [DOI] [PubMed] [Google Scholar]
- 31.Woods M, Schaefer EJ, Morrill A, Goldin BR, Longcope C, Dwyer D, Gorbach SL. Effect of menstrual cycle phase on plasma lipids. J Clin Endocrinol Metab. 1987;65:321–323. doi: 10.1210/jcem-65-2-321. [DOI] [PubMed] [Google Scholar]
- 32.Karpanou EA, Vyssoulis GP, Georgoudi DG, Toutouza MG, Toutousaz PK. Disparate serum lipid changes between normotensive and hypertensive women during the menstrual cycle. Am J Cardiol. 1992;70:111–113. doi: 10.1016/0002-9149(92)91402-p. [DOI] [PubMed] [Google Scholar]
- 33.Berlin E, Judd JT, Nair PP, Jones DY, Taylor PR. Dietary fat and hormonal influences on lipoprotein fluidity and composition in premenopausal women. Atherosclerosis. 1991;86:95–110. doi: 10.1016/0021-9150(91)90206-i. [DOI] [PubMed] [Google Scholar]