

Abstract
The mycotoxin deoxynivalenol (DON) causes serious problems worldwide in the production of crops such as wheat and barley because of its toxicity toward humans and livestock. A bacterial culture capable of degrading DON was obtained from soil samples collected in wheat fields using an enrichment culture procedure. The isolated bacterium, designated strain WSN05-2, completely removed 1,000 μg/mL of DON from the culture medium after incubation for 10 days. On the basis of phylogenetic studies, WSN05-2 was classified as a bacterium belonging to the genus Nocardioides. WSN05-2 showed significant growth in culture medium with DON as the sole carbon source. High-performance liquid chromatography analysis indicated the presence of a major initial metabolite of DON in the culture supernatant. The metabolite was identified as 3-epi-deoxynivalenol (3-epi-DON) by mass spectrometry and 1H and 13C nuclear magnetic resonance analysis. The amount of DON on wheat grain was reduced by about 90% at 7 days after inoculation with WSN05-2. This is the first report of a Nocardioides sp. strain able to degrade DON and of the yet unknown 3-epi-DON as an intermediate in the degradation of DON by a microorganism.
Keywords: Fusarium graminearum, Trichothecenes, Deoxynivalenol, Mycotoxin degradation, Nocardioides
Introduction
The trichothecene mycotoxin deoxynivalenol (3α,7α,15-trihydroxy-12,13-epoxytrichothec-9-en-8-one, DON; Fig. 1) is a secondary metabolite produced by many species of the genus Fusarium (Mirocha et al. 1976; Ramakrishna et al. 1989; Schoental 1985). This mycotoxin occurs predominantly in grains such as wheat, barley, oats, rye, and maize, and less often in rice, sorghum, and triticale (Goswami and Kistler 2004; Goswami et al. 2006; Yoshizawa and Jin 1995), during growth or storage in high-moisture environments. The main toxic effect of DON and other trichothecene mycotoxins at the cellular level in both humans and domesticated animals is the inhibition of protein synthesis by binding to the ribosome (Ehrlich and Daigle 1987; Middlebrook and Leatherman 1989a, b; Rotter et al. 1996). The trichothecenes have also been reported to have immunosuppressive effects (Berek et al. 2001; Bondy and Pestka 2000; Pestka et al. 2004). The 12,13-epoxide group is essential for the toxic activity of trichothecenes (Eriksen et al. 2004). Although Fusarium head blight is suppressed by fungicides and by the use of resistant varieties, these measures do not reliably reduce the DON levels to below the legal limit. The concern about DON contamination is reinforced by the high resistance of the toxin to heat (Lauren and Smith 2001; Yumbe-Guevara et al. 2003). In addition, attempts to develop a decontamination strategy based on gamma irradiation have been unsuccessful (O’Neill et al. 1993).
Fig. 1.

Structure of deoxynivalenol (3α,7α,15-trihydroxy-12,13-epoxytrichothec-9-en-8-one)
The utilization of DON-degrading bacteria can be a promising approach for in situ detoxification. The exploration and characterization of novel bacteria that degrade DON will lead to a better understanding of the diversity in DON-degrading bacteria and the way in which they degrade DON in the environment. It may also provide us with more specific choices of the strains and the procedures to be used for the in situ detoxification of DON.
To date, several microbial consortia and bacterial isolates that degrade DON have been reported and their degradation products have been identified (Karlovsky 1999). It has been reported that DON is converted to de-epoxy DON under some anaerobic conditions such as those of the rumen or intestine (He et al. 1992; King et al. 1984; Swanson et al. 1987). Indeed, an anaerobic bacterial isolate, Eubacterium sp. strain BBSH797, has been shown to de-epoxydize DON (Fuchs et al. 2002). Recently, some bacterial strains isolated from chicken large intestines have been reported to de-epoxidize DON, but the details of culturing, isolation, and identification of those strains have yet to be published (Young et al. 2007). For the application of DON-degrading bacteria for detoxification in wheat fields, the bacterial candidates must have the capability of degrading DON under aerobic conditions; however, information about aerobic bacteria able to degrade DON has been limited. Shima et al. (1997) isolated Gram-negative bacterial strain E3-39 from a soil sample, which aerobically metabolized DON into 3-oxo-4-deoxynivalenol (3-keto-DON). This strain represents the only example of aerobic DON-degrading bacteria, and its oxidation of DON to 3-keto-DON is the only aerobic bacterial catabolism of DON known to date. Volkl et al. (2004) constructed a microbial consortia that aerobically degrades DON in which DON was also converted to 3-keto-DON.
Here, we report a novel Gram-positive bacterium Nocardioides sp. strain WSN05-2 which utilizes DON as the sole source of carbon and energy under aerobic conditions. This strain degrades DON to produce a novel intermediate, 3-epi-DON.
Materials and methods
Media
The mineral medium (MM) used for the enrichment procedure is that of Kirimura et al. (1999) with the following modifications. The medium contained (per liter) 1.6 g of Na2HPO4, 1 g of KH2PO4, 0.5 g of MgSO4⋅7H2O, 0.5 g of NaNO3, 0.5 g of (NH4)2SO4, 0.025 g of CaCl2⋅2H2O, 2 mL of trace metal solution (1.5 g of FeCl2⋅4H2O, 0.190 g of CoCl2⋅6H2O, 0.1 g of MnCl2⋅4H2O, 0.07 g of ZnCl2, 0.062 g of H3BO3, 0.036 g of Na2MoO4⋅2H2O, 0.024 g of NiCl2⋅6H2O, 0.017 g of CuCl2⋅2H2O, and 0.01 g of AlK(SO4)2⋅12H2O per liter), 1 mL of vitamin solution (2 mg of biotin, 2 mg of folic acid, 5 mg of thiamine-HCl, 5 mg of riboflavin, 10 mg of pyridoxine-HCl, 50 mg of cyanocobalamin, 5 mg of niacin, 5 mg of Ca-pantothenate, 5 mg of p-aminobenzoate, and 5 mg of thiolactic acid per liter), and the indicated amount of DON (10–1,000 μg/mL). The nutrient-rich potato semisynthetic (PS) medium lacking DON, used for maintaining pure cultures, consisted of 300 g potato extract, 5 g of peptone, 20 g of sucrose, 2 g of Na2HPO4⋅12H2O, and 0.5 g of Ca(NO3)2 per liter.
Trichothecenes
DON was purchased from Wako Pure Chemicals (Osaka, Japan) and adjusted to 10 g/L in sterile distilled water.
Environmental samples
Soil samples were collected from the environment including wheat field soil of the National Institute for Agro-Environmental Sciences, Tsukuba, Ibaraki, Japan, for the isolation of microorganisms capable of transforming DON.
Enrichment of bacterial cultures for DON degradation
Soil samples of approximately 0.5 g were suspended in 2.5 mL of MM supplemented with 100 μg/mL of DON, and the cultures were incubated at 28°C for 7 days with shaking. Then, 100 μL of these cultures was added to 2.5 mL of MM containing 100 μg/mL DON, followed by 7 days of incubation with shaking at 28°C. This procedure was repeated two more times. Culture samples showing bacterial growth, as indicated by turbidity, were selected, and the concentrations of DON in the culture media were monitored by high-performance liquid chromatography (HPLC) as described below. One culture sample was found to decrease the DON concentration, and the DON-degrading bacteria were isolated from this sample.
Isolation of a pure strain from DON-degrading bacterial communities
The cultures obtained following the enrichment culture procedure were serially diluted in sterile distilled water, and 0.1-mL aliquots were spread onto nutrient agar medium (NA; Difco, Grand Island, NY, USA) and 100-fold diluted NA. The resulting plates were incubated at 28°C for 5 days. Randomly selected bacterial colonies were analyzed for DON-degrading ability using HPLC. A bacterial strain (WSN05-2) that degraded DON was selected and stored at −20°C until use. WSN05-2 strain is stored (in liquid nitrogen vapor at −165°C by lyophilization or deep freezing at −80°C) at the Microorganisms Section (MAFF) of the NIAS Genebank, National Institute of Agrobiological Sciences, Tsukuba, Ibaraki, Japan, as MAFF311580.
Taxonomic characterization of the isolated bacterium
The strain WSN05-2 was tested by Gram staining and its cell morphology examined. The 16S ribosomal RNA gene (rDNA) of the isolate was also analyzed. Template DNA for a polymerase chain reaction (PCR) was prepared using a DNA extraction kit (ISOPLANT II; Wako Pure Chemicals) according to the manufacturer’s instructions. The primers designed to specifically amplify the 16S rDNA region, 10f (5′-AGTTTGATCCTGGCTCAG-3′) and 1541r (5′-AAGGAGGTGATCCAGCCG-3′), were synthesized by Espec Oligo Service Corp. (Ibaraki, Japan). The reaction mixture contained Premix Taq (Ex Taq Version; Takara Bio Inc., Shiga, Japan), 10 pmol of each primer, and template DNA. PCR was performed using a GeneAmp PCR System 9700 (Applied Biosystems, Foster City, CA, USA) with the following program: 94°C for 3 min; 30 cycles of 95°C for 1 min, 55°C for 1 min, and 72°C for 1 min 30 s; 72°C for 10 min; and a 4°C soak. The PCR products were purified using a QIAquick PCR Purification Kit (Qiagen K. K., Tokyo, Japan). DNA sequencing was conducted using an ABI 3100 Prism DNA Sequencer (Applied Biosystems). Sequencing reactions were performed using the forward primer 10f, the reverse primer 1541r, and a one half volume reaction mixture of the BigDye Terminator v. 3.1 Ready Reaction Cycle Sequencing Kit (Applied Biosystems), containing 1 pmol primer, BigDye Terminator v. 3.1 5× sequencing buffer (Applied Biosystems), and purified PCR product. The sequencing yielded approximately 1,214 bp of useful 16S rDNA sequence. Sequences similar to the 16S rDNA of WSN05-2 were identified using BLAST (National Center for Biotechnology Information, http://www.ncbi.nlm.nih.gov). WSN05-2 strain (accession no. MAFF311580) are deposited at the Microorganisms Section (MAFF) of the NIAS Genebank, National Institute of Agrobiological Sciences, Tsukuba, Ibaraki, Japan.
Extraction and analysis of trichothecenes
A 100-μL aliquot of a fresh culture sample was mixed with 400 μL of ethyl acetate and 100 μL of salting out solution (water saturated with NaCl). After vigorous shaking for 10 s on a vortex mixer, the resultant organic solvent layer was collected. This procedure was repeated three times and the collected organic solvent layer was dried by evaporation. The residue was resuspended in 100 μL of methanol and DON was quantitated by reversed-phase HPLC. The HPLC system (Waters, Milford, MA, USA) consisted of a 600E pump, 2996 photodiode array detector, Shodex Silica C18M 4E column (4.6-mm ID × 250 mm; Showa Denko K. K., Tokyo, Japan), and Empower 2 software. The mobile phase contained methanol and water (15:85, v/v) at a flow rate of 1.0 mL/min. Measurements of DON for routine assay were performed in wavelength at 220 nm. When DON and its metabolites (compound A and compound B) were simultaneously measured, a wavelength of 240 nm was used. The column was heated to 40°C.
Assay of activity for the degradation of DON in wheat grain
The isolated bacterium, strain WSN05-2, was shaken in PS medium at 28°C for 2 days. Bacterial cells were recovered by centrifugation at 6000×g for 10 min and washed twice with 50 mM potassium phosphate buffer (pH 7.0). The cells were suspended in MM medium containing DON (100 μg/mL) and grown at 28°C for 30 h. The resultant bacterial cells were washed twice with potassium phosphate buffer (pH 7.0) and suspended in sterile distilled water. Two milliliters of DON solution (1,000 μg/mL) was added to 100 wheat grains (cv. Norin no. 61), mixed thoroughly, and air-dried naturally for 1 h at 25°C in a Petri dish. Sixty DON-treated grains were inoculated with 600 μL (10 μL/kernel) of bacterial suspension (OD610 = 4.0) in a Petri dish (9-cm diameter) and incubated in a moist chamber for 2 days. Three inoculated grains were selected randomly, air-dried for 30 s with an air dryer, and ground into flour with a mixer (A10S11, IKA-Labortechnik, Staufen, Germany). The flour was mixed with 1 mL of 85% acetonitrile and shaken for 1 h. The supernatant was recovered after centrifugation at 10,000×g for 5 min. The sediment was mixed with 600 μL of 85% acetonitrile, treated in the same way, and the supernatant was recovered. The volume of the combined supernatants was reduced to 100 μL by evaporation. The solution was mixed with 600 μL of saturated salt solution and extracted twice with 700 μL of ethyl acetate. The organic phases were combined, ethyl acetate was removed by evaporation, and the residue was dissolved in 500 μL of 50% methanol for HPLC analysis.
Isolation and characterization of a DON metabolite, 3-epi-DON
A 20-mg sample of DON was incubated in MM (20 mL) with the strain WSN05-2 at 28°C for 96 h. The culture supernatant was concentrated by freeze drying and then extracted with methanol. DON transformation products were isolated and purified by preparative HPLC fractionation using an Inertsil ODS-3 column (20-mm ID × 250 mm; GL Sciences Inc., Tokyo, Japan) in the mobile phase containing methanol and water (15:85, v/v) at a flow rate of 8.0 mL/min. Evaporation in vacuo yielded 4.5 mg of pure compound. Mass spectrometry (MS) and 1H, 13C, and 2D nuclear magnetic resonance (NMR) spectroscopy were used for structural characterization. NMR spectra were recorded on an Alpha 600 spectrometer (JEOL, Tokyo, Japan) using standard JEOL software (Alpha Data System) and Inova 600 (Varian, Palo Alto, CA, USA) equipped with a TH5 probe at 30°C, with the sample dissolved in deuterated chloroform. Chemical shifts were determined with respect to the residual chloroform and quoted with respect to tetramethylsilane. Coupling constants were expressed in hertz and multiplicities were indicated with the standard symbols: 2D NMR experiments, 1H-1H-COSY, HSQC, HMBC, and NOESY. High-resolution mass spectra were obtained with a JMS-GCmate II gas chromatograph/mass spectrometer (JEOL) in the direct-inlet electron impact ionization (DI-EI) mode. The optical rotations were determined using a P-1020 spectropolarimeter (JASCO, Tokyo, Japan).
Nucleotide sequence
The 16S rDNA sequence of the strain WSN05-2 has been deposited in the DDBJ under accession no. AB282680.
Statistical analysis
Statistical analysis of data was carried out using JMP software (version 5.01J; SAS Institute Japan, Tokyo, Japan). Tukey’s test was used for the comparison of means.
Results
Isolation of DON-transforming strain WSN05-2 and its taxonomic characterization
A bacterial community enrichment culture strategy with MM containing DON (100 μg/mL) as the only carbon and energy source was used to isolate DON-catabolizing bacteria from soil samples. Among ten environmental soil samples, bacterial growth was apparent after three passages in MM with DON only for one sample collected from a wheat field. When the bacteria in MM were plated on NA by the dilution method, at least four different bacterial colonies grew on the agar plate over 5 days at 25°C, but not all of these bacterial isolates displayed DON degradation. The bacterial strain with DON-degrading activity was recovered with 1/100 NA when it formed a colony of <1 mm in diameter over 5 days at 25°C, although its growth was slower than that of the other bacteria. The strain was designated WSN05-2. WSN05-2 is a Gram-positive, non-spore-forming, coccal-shaped bacterium. The sequence of its 16S rDNA revealed that WSN05-2 is closely related to the Nocardioides sp. strain AN3 (AB183711), which belongs to the genus Nocardioides, as illustrated in the phylogenetic tree in Fig. 2. On the basis of these characteristics, WSN05-2 was inferred to be a member of the genus Nocardioides.
Fig. 2.

Phylogenetic tree based on 16S rRNA gene sequences of the strain WSN05-2 (AB282680) and related microorganisms. The bar indicates 1% sequence divergence
DON-degrading ability of WSN05-2
The DON degradation rate and growth curve of WSN05-2 were examined using concentrations of DON ranging from 10 to 1,000 μg/mL (Fig. 3). The growth of WSN05-2 was directly proportional to the amount of DON in the medium. The final OD values at 140 h after treatment as an indicator of bacterial growth increased with DON concentration. HPLC analysis revealed two metabolites (Fig. 4) that appeared during the growth of WSN05-2 in culture medium containing 1,000 μg/mL DON, but neither component was detected after 10 days of incubation. Furthermore, NMR analysis of a residue obtained from ethyl acetate extracts of culture supernatants after 10 days of incubation did not reveal any signal typical of trichothecenes, indicating that the trichothecene skeleton had been completely destroyed.
Fig. 3.

Degradation of DON by the strain WSN05-2 in MM containing DON at 1,000 μg/mL (filled circle, open circle), 100 μg/mL (filled triangle, open triangle), or 10 μg/mL (filled square, open square) and growth of the strain in these media. Filled symbols DON concentration, Open symbols growth of WSN05-2
Fig. 4.

HPLC elution profile of extracted DON-degrading culture medium. The strain WSN05-2 was grown for 50 h in MM containing 1,000 μg/mL DON
Identification of the DON metabolite 3-epi-DON
Strain WSN05-2 was cultured for 2 days in MM containing 1,000 μg/mL DON and the culture was concentrated by freeze drying and extracted with methanol. Analytical HPLC analysis indicated the appearance of two major biotransformation products (retention times of 9.9 min for compound A and 5.1 min for compound B). Compound A was fully characterized by 1H and 13C NMR spectroscopy (Table 1) and MS analyses. All NMR assignments were confirmed by 1H-1H and 13C-1H 2D correlated spectroscopy. Comparison with the NMR data for DON (Table 1) facilitated the structural identification. The 13C and DEPT spectra indicated the presence of the carbon skeleton of trichothecene in compound A. The molecular mass derived from the observation of a molecular ion peak at m/z 296.12862 in DI-EI-MS and the NMR data indicated a molecular formula of C15H20O6, 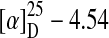 (c 0.25, methanol). The two doublets of proton signals with a small geminal coupling constant (4.5 Hz) at 3.16 and 3.11 ppm was indicative of the presence of a 12,13-epoxide group, but comparison of the chemical shifts of DON and compound A showed the absence of spin–spin interaction between 2-H and 3-H in compound A (Table 1). 1H-1H COSY results, which showed no correlation signal between 2-H and 3-H (Fig. 5), also supported this result. Additionally, the HSQC experiment unambiguously revealed the overlapping signals of 15-Ha, 15-Hb (δH = 3.72, 3.80, d, 12.0; C-15, δc = 62.1 ppm) and 2-H (δH = 3.70, s; C-2, δc = 83.8 ppm). In contrast, a correlation between H-3 and H-11 was observed in a NOESY analysis of compound A (Fig. 5). Structural analysis of compound A calculated the dihedral angle of 2-H and 3-H at 90.6°, indicating no or a very small 3J-coupling constant between 2-H and 3-H. These results are consistent with those of the NMR analysis. These results indicate that an epimer is produced by bacterial transformation. Therefore, the first transformation of DON by the strain WSN05-2 is an epimerization of C-3, and compound A is 3-epi-DON. Compound B had the maximum spectrum at around 240 nm and DON and 3-epi-DON had those at around 220 nm (data not shown), suggesting that the production of compound B from 3-epi-DON or DON involves the structural change that results in the difference in absorbance spectrum, although compound B has not yet been structurally determined.
(c 0.25, methanol). The two doublets of proton signals with a small geminal coupling constant (4.5 Hz) at 3.16 and 3.11 ppm was indicative of the presence of a 12,13-epoxide group, but comparison of the chemical shifts of DON and compound A showed the absence of spin–spin interaction between 2-H and 3-H in compound A (Table 1). 1H-1H COSY results, which showed no correlation signal between 2-H and 3-H (Fig. 5), also supported this result. Additionally, the HSQC experiment unambiguously revealed the overlapping signals of 15-Ha, 15-Hb (δH = 3.72, 3.80, d, 12.0; C-15, δc = 62.1 ppm) and 2-H (δH = 3.70, s; C-2, δc = 83.8 ppm). In contrast, a correlation between H-3 and H-11 was observed in a NOESY analysis of compound A (Fig. 5). Structural analysis of compound A calculated the dihedral angle of 2-H and 3-H at 90.6°, indicating no or a very small 3J-coupling constant between 2-H and 3-H. These results are consistent with those of the NMR analysis. These results indicate that an epimer is produced by bacterial transformation. Therefore, the first transformation of DON by the strain WSN05-2 is an epimerization of C-3, and compound A is 3-epi-DON. Compound B had the maximum spectrum at around 240 nm and DON and 3-epi-DON had those at around 220 nm (data not shown), suggesting that the production of compound B from 3-epi-DON or DON involves the structural change that results in the difference in absorbance spectrum, although compound B has not yet been structurally determined.
Table 1.
1H and 13C NMR data for DON and compound A (in CDCl3 at 30°C)
| Position | 1H-NMR δH, ppm (mult., J in Hz) | 13C-NMR δC (ppm) | ||
|---|---|---|---|---|
| DON | Compound A | DON | Compound A | |
| 2 | 3.6 (d, 4.5) | 3.70 (s) | 80.8 | 83.8 |
| 3 | 4.5 (ddd, 4.5, 4.5, 11.0) | 4.33 (dd, 3.0, 7.5) | 69.2 | 72.5 |
| 4 | 2.23 (dd, 4.0, 15.0) | 2.93 (dd, 7.5, 15.5) | 43.4 | 46.1 |
| 2.10 (dd, 11.0, 15.0) | 1.60 (dd, 3.0, 15.5) | |||
| 5 | – | – | 46.5 | 46.2 |
| 6 | – | – | 52.6 | 51.8 |
| 7 | 4.90 (br. s) | 4.75 (s) | 74.1 | 74.0 |
| 8 | – | – | 199.6 | 199.9 |
| 9 | – | – | 136.6 | 136.3 |
| 10 | 6.55 (d, 6.0) | 6.53 (dd, 1.5, 6.0) | 137.7 | 138.1 |
| 11 | 4.57 (d, 56.0) | 4.19 (d, 6.0) | 70.5 | 70.0 |
| 12 | – | – | 64.3 | 65.3 |
| 13 | 3.35 (d, 4.0) | 3.16 (d, 4.5) | 47.8 | 46.2 |
| 3.22 (d, 4.0) | 3.11 (d, 4.5) | |||
| 14 | 1.33 (3H, s) | 1.22 (3H, s) | 13.8 | 14.1 |
| 15 | 3.90 (d, 12.0) | 3.80 (d, 12.0) | 62.0 | 62.1 |
| 3.74 (d, 12.0) | 3.72 (d, 12.0) | |||
| 16 | 1.90 (br. s) | 1.87 (3H, s) | 15.2 | 15.3 |
Fig. 5.

COSY and selective NOESY correlations of DON (a) and compound A, a DON metabolite (3-epi-DON) (b)
Depletion of DON and the accumulation of metabolites
We investigated the time-dependent change of DON, compound A (3-epi-DON), and compound B by WSN05-2 in MM medium containing 1,000 μg/L DON (Fig. 6). After 50 h of incubation, accumulation of compound A and compound B was initiated. Compound A was most accumulated after 100 h and disappeared after 200 h. On the other hand, accumulation of compound B became maximum after 150 h, and compound B disappeared after 225 h. Compound B remained detectable by HPLC for several hours after the disappearance of 3-epi-DON. This result inferred that compound B was derived from 3-epi-DON, and we performed an additional feeding experiment using 3-epi-DON as the substrate to demonstrate this hypothesis. When purified 3-epi-DON (100 μg/mL) was added to the MM medium instead of DON, accumulation of compound B by WSN05-2 was observed. This result indicated that compound B was a lower intermediate of 3-epi-DON (data not shown).
Fig. 6.

Degradation of DON by WSN05-2 in MM containing 1,000 μg/mL of DON. Filled circle peak area of DON, Triangle peak area of compound A, Square peak area of compound B, Open circle growth of WSN05-2
Degradation of DON on wheat grain by WSN05-2
The amount of DON on wheat grains inoculated with the WSN05-2 strain was reduced by about 60% after 3 days, whereas no significant change was observed on the control grains. After 7 days, the reduction of DON levels on grains reached about 90% (Fig. 7).
Fig. 7.

Decrease of DON on DON-inoculated wheat grains by strain WSN05-2. The amount of DON was analyzed by HPLC after 0, 3, and 7 days. Different letters indicate significant differences (p = 0.05), as determined by a Tukey test
Discussion
Bacterial strain WSN05-2 isolated from a soil sample transformed DON into two products, 3-epi-DON (compound A) and compound B, of unknown structure (Figs. 5 and 6). Both products disappeared from culture supernatants after a prolonged incubation, indicating that they are not dead-end products but rather intermediates in the degradation of DON by WSN05-2. To date, the reported bacterial degradation pathways for DON have been limited to two types: the oxidation of DON into 3-keto-DON under aerobic conditions (Shima et al. 1997; Volkl et al. 2004) and the reductive de-epoxydation into de-epoxy DON under anaerobic conditions (Eriksen et al. 2004; Fuchs et al. 2002). Very recently, the aerobic fungus Aspergillus tubingensis NJA-1 has been demonstrated to degrade DON with the appearance of an unidentified metabolite in the culture medium, which was postulated as the hydrolyzed product of DON (He et al. 2008). Therefore, this is the first study showing that 3-epi-DON is an intermediate in the degradation pathway of DON.
The possibility exists that the toxicity of 3-epi-DON is as high as that of DON because the 12,13-epoxide group, which is assumed to be essential for toxic activity, remains intact in this transformation product. A synthetically derived epimer of T-2 (type A trichothecene mycotoxin) has been reported to bind to the same ribosomal site as authentic T-2, but apparently with a lower affinity (Middlebrook and Leatherman 1989a, b). The ribosomal binding and immunosuppressive properties of 3-epi-DON may provide valuable information on the structural basis of toxicity of trichothecenes.
Based on a phylogenetic rDNA analysis, the DON degrading strainWSN05-2 belongs to the genus Nocardioides (Fig. 2). Recent reports on the strains belonging to this genus have revealed that Nocardioides contains many bacteria that degrade xenobiotic compounds such as atrazine (Piutti et al. 2003; Satsuma 2006), crude oil (Schippers et al. 2005), carbazole (Inoue et al. 2005, 2006), and dibenzofuran (Kubota et al. 2005). The metabolic versatility of these bacteria suggests that the genus Nocardioides has evolved some mechanisms to adapt to uncommon carbon sources. To our knowledge, WSN05-2 is currently the only available bacterial isolate able to utilize DON as the sole carbon and energy source (Fig. 3), whereas the other DON-degrading bacteria reported so far require other substrates for growth (Fuchs et al. 2002; Shima et al. 1997; Volkl et al. 2004).
In addition to WSN05-2, which originated from soil in a wheat field, we isolated similar strains belonging to the genus Nocardioides from wheat field soil and wheat leaves (data not shown). When strain WSN05-2 was inoculated onto wheat grains treated with DON, the amount of DON on wheat grains decreased by about 90% in 7 days (Fig. 7). From these results, we speculate that the Nocardioides strain WSN05-2 might be associated with wheat plants and exerts more or less its DON-degrading activity in wheat. However, little is known about where the DON exists in the DON-infested wheat grain. For the effective application of DON-degrading bacteria, the dynamics of DON and DON-degrading ability of the strain WSN05-2 in the Fusarium-infected wheat grain should be studied in detail. The isolation and characterization of more DON-degrading bacteria and the analysis of the degradation pathways of DON may provide more options and better theoretical strategies for in situ detoxification. The analysis of the second major intermediate of DON degradation by WSN05-2 (compound B) by NMR is in progress to elucidate the complete catabolic pathway of DON by this strain.
References
- Berek L, Petri IB, Mesterhazy A, Teren J, Molnar J. Effects of mycotoxins on human immune functions in vitro. Toxicol in Vitro. 2001;15:25–30. doi: 10.1016/S0887-2333(00)00055-2. [DOI] [PubMed] [Google Scholar]
- Bondy GS, Pestka JJ. Immunomodulation by fungal toxins. J Toxicol Environ Health B Crit Rev. 2000;3:109–143. doi: 10.1080/109374000281113. [DOI] [PubMed] [Google Scholar]
- Ehrlich KC, Daigle KW. Protein-synthesis inhibition by 8-oxo-12,13-epoxytrichothecenes. Biochim Biophys Acta. 1987;923:206–213. doi: 10.1016/0304-4165(87)90005-5. [DOI] [PubMed] [Google Scholar]
- Eriksen GS, Pettersson H, Lundh T. Comparative cytotoxicity of deoxynivalenol, nivalenol, their acetylated derivatives and de-epoxy metabolites. Food Chem Toxicol. 2004;42:619–624. doi: 10.1016/j.fct.2003.11.006. [DOI] [PubMed] [Google Scholar]
- Fuchs E, Binder EM, Heidler D, Krska R. Structural characterization of metabolites after the microbial degradation of type A trichothecenes by the bacterial strain BBSH 797. Food Addit Contam. 2002;19:379–386. doi: 10.1080/02652030110091154. [DOI] [PubMed] [Google Scholar]
- Goswami RS, Kistler HC. Heading for disaster: Fusarium graminearum on cereal crops. Mol Plant Pathol. 2004;5:515–525. doi: 10.1111/j.1364-3703.2004.00252.x. [DOI] [PubMed] [Google Scholar]
- Goswami RS, Xu JR, Trail F, Hilburn K, Kistler HC. Genomic analysis of host–pathogen interaction between Fusarium graminearum and wheat during early stages of disease development. Microbiology. 2006;152:1877–1890. doi: 10.1099/mic.0.28750-0. [DOI] [PubMed] [Google Scholar]
- He P, Young LG, Forsberg C. Microbial transformation of deoxynivalenol (vomitoxin) Appl Environ Microbiol. 1992;58:3857–3863. doi: 10.1128/aem.58.12.3857-3863.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- He C, Fan Y, Liu G, Zhang H. Isolation and identification of a strain of Aspergillus tubingensis with deoxynivalenol biotransformation capability. Int J Mol Sci. 2008;9:2366–2375. doi: 10.3390/ijms9122366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inoue K, Habe H, Yamane H, Omori T, Nojiri H. Diversity of carbazole-degrading bacteria having the car gene cluster: isolation of a novel Gram-positive carbazole-degrading bacterium. FEMS Microbiol Lett. 2005;245:145–153. doi: 10.1016/j.femsle.2005.03.009. [DOI] [PubMed] [Google Scholar]
- Inoue K, Habe H, Yamane H, Nojiri H. Characterization of novel carbazole catabolism genes from gram-positive carbazole degrader Nocardioides aromaticivorans IC177. Appl Environ Microbiol. 2006;72:3321–3329. doi: 10.1128/AEM.72.5.3321-3329.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karlovsky P. Biological detoxification of fungal toxins and its use in plant breeding, feed and food production. Nat Toxins. 1999;7:1–23. doi: 10.1002/(SICI)1522-7189(199902)7:1<1::AID-NT37>3.0.CO;2-9. [DOI] [PubMed] [Google Scholar]
- King RR, McQueen RE, Levessue D, Greenhalgh R. Transformation of deoxynivalenol (vomitoxin) by rumen microorganisms. J Agric Food Chem. 1984;32:1181–1183. doi: 10.1021/jf00125a061. [DOI] [Google Scholar]
- Kirimura K, Nakagawa H, Tsuji K, Matsuda K, Kurane R, Usami S. Selective and continuous degradation of carbazole contained in petroleum oil by resting cells of Sphingomonas sp. CDH-7. Biosci Biotechnol Biochem. 1999;63:1563–1568. doi: 10.1271/bbb.63.1563. [DOI] [PubMed] [Google Scholar]
- Kubota M, Kawahara K, Sekiya K, Uchida T, Hattori Y, Futamata H, Hiraishi A. Nocardioides aromaticivorans sp. nov., a dibenzofuran-degrading bacterium isolated from dioxin-polluted environments. Syst Appl Microbiol. 2005;28:165–174. doi: 10.1016/j.syapm.2004.10.002. [DOI] [PubMed] [Google Scholar]
- Lauren DR, Smith WA. Stability of the Fusarium mycotoxins nivalenol, deoxynivalenol and zearalenone in ground maize under typical cooking environments. Food Addit Contam. 2001;18:1011–1016. doi: 10.1080/02652030110052283. [DOI] [PubMed] [Google Scholar]
- Middlebrook JL, Leatherman DL. Binding of T-2 toxin to eukaryotic cell ribosomes. Biochem Pharmacol. 1989;38:3103–3110. doi: 10.1016/0006-2952(89)90021-X. [DOI] [PubMed] [Google Scholar]
- Middlebrook JL, Leatherman DL. Specific association of T-2 toxin with mammalian cells. Biochem Pharmacol. 1989;38:3093–3102. doi: 10.1016/0006-2952(89)90020-8. [DOI] [PubMed] [Google Scholar]
- Mirocha CJ, Pathre SV, Schauerhamer B, Christensen CM. Natural occurrence of Fusarium toxins in feedstuff. Appl Environ Microbiol. 1976;32:553–556. doi: 10.1128/aem.32.4.553-556.1976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Neill K, Damoglou AP, Patterson MF. The stability of deoxynivalenol and 3-acetyl deoxynivalenol to gamma-irradiation. Food Addit Contam. 1993;10:209–215. doi: 10.1080/02652039309374143. [DOI] [PubMed] [Google Scholar]
- Pestka JJ, Zhou HR, Moon Y, Chung YJ. Cellular and molecular mechanisms for immune modulation by deoxynivalenol and other trichothecenes: unraveling a paradox. Toxicol Lett. 2004;153:61–73. doi: 10.1016/j.toxlet.2004.04.023. [DOI] [PubMed] [Google Scholar]
- Piutti S, Semon E, Landry D, Hartmann A, Dousset S, Lichtfouse E, Topp E, Soulas G, Martin-Laurent F. Isolation and characterisation of Nocardioides sp. SP12, an atrazine-degrading bacterial strain possessing the gene trzN from bulk- and maize rhizosphere soil. FEMS Microbiol Lett. 2003;221:111–117. doi: 10.1016/S0378-1097(03)00168-X. [DOI] [PubMed] [Google Scholar]
- Ramakrishna Y, Bhat RV, Ravindranath V. Production of deoxynivalenol by Fusarium isolates from samples of wheat associated with a human mycotoxicosis outbreak and from sorghum cultivars. Appl Environ Microbiol. 1989;55:2619–2620. doi: 10.1128/aem.55.10.2619-2620.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rotter BA, Prelusky DB, Pestka JJ. Toxicology of deoxynivalenol (vomitoxin) J Toxicol Environ Health. 1996;48:1–34. doi: 10.1080/009841096161447. [DOI] [PubMed] [Google Scholar]
- Satsuma K. Characterisation of new strains of atrazine-degrading Nocardioides sp. isolated from Japanese riverbed sediment using naturally derived river ecosystem. Pest Manag Sci. 2006;62:340–349. doi: 10.1002/ps.1172. [DOI] [PubMed] [Google Scholar]
- Schippers A, Schumann P, Sproer C. Nocardioides oleivorans sp. nov., a novel crude-oil-degrading bacterium. Int J Syst Evol Microbiol. 2005;55:1501–1504. doi: 10.1099/ijs.0.63500-0. [DOI] [PubMed] [Google Scholar]
- Schoental R. Trichothecenes, zearalenone, and other carcinogenic metabolites of Fusarium and related microfungi. Adv Cancer Res. 1985;45:217–290. doi: 10.1016/S0065-230X(08)60270-5. [DOI] [PubMed] [Google Scholar]
- Shima J, Takase S, Takahashi Y, Iwai Y, Fujimoto H, Yamazaki M, Ochi K. Novel detoxification of the trichothecene mycotoxin deoxynivalenol by a soil bacterium isolated by enrichment culture. Appl Environ Microbiol. 1997;63:3825–3830. doi: 10.1128/aem.63.10.3825-3830.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swanson SP, Nicoletti J, Rood HD, Jr, Buck WB, Cote LM, Yoshizawa T. Metabolism of three trichothecene mycotoxins, T-2 toxin, diacetoxyscirpenol and deoxynivalenol, by bovine rumen microorganisms. J Chromatogr. 1987;414:335–342. doi: 10.1016/0378-4347(87)80058-0. [DOI] [PubMed] [Google Scholar]
- Volkl A, Vogler B, Schollenberger M, Karlovsky P. Microbial detoxification of mycotoxin deoxynivalenol. J Basic Microbiol. 2004;44:147–156. doi: 10.1002/jobm.200310353. [DOI] [PubMed] [Google Scholar]
- Yoshizawa T, Jin YZ. Natural occurrence of acetylated derivatives of deoxynivalenol and nivalenol in wheat and barley in Japan. Food Addit Contam. 1995;12:689–694. doi: 10.1080/02652039509374358. [DOI] [PubMed] [Google Scholar]
- Young JC, Zhou T, Yu H, Zhu H, Gong J. Degradation of trichothecene mycotoxins by chicken intestinal microbes. Food Chem Toxicol. 2007;46:136–143. doi: 10.1016/j.fct.2006.07.028. [DOI] [PubMed] [Google Scholar]
- Yumbe-Guevara BE, Imoto T, Yoshizawa T. Effects of heating procedures on deoxynivalenol, nivalenol and zearalenone levels in naturally contaminated barley and wheat. Food Addit Contam. 2003;20:1132–1140. doi: 10.1080/02652030310001620432. [DOI] [PubMed] [Google Scholar]