

Abstract
Collective decision-making plays a central part in the lives of many social animals. Two important factors that influence collective decision-making are information uncertainty and conflicting preferences. Here, I bring together, and briefly review, basic models relating to animal collective decision-making in situations with information uncertainty and in situations with conflicting preferences between group members. The intention is to give an overview about the different types of modelling approaches that have been employed and the questions that they address and raise. Despite the use of a wide range of different modelling techniques, results show a coherent picture, as follows. Relatively simple cognitive mechanisms can lead to effective information pooling. Groups often face a trade-off between decision accuracy and speed, but appropriate fine-tuning of behavioural parameters could achieve high accuracy while maintaining reasonable speed. The right balance of interdependence and independence between animals is crucial for maintaining group cohesion and achieving high decision accuracy. In conflict situations, a high degree of decision-sharing between individuals is predicted, as well as transient leadership and leadership according to needs and physiological status. Animals often face crucial trade-offs between maintaining group cohesion and influencing the decision outcome in their own favour. Despite the great progress that has been made, there remains one big gap in our knowledge: how do animals make collective decisions in situations when information uncertainty and conflict of interest operate simultaneously?
Keywords: collective decisions, game-theory models, self-organizing systems
1. Introduction
Collective decision-making plays a central part in the lives of many social animals. Groups often decide collectively about vital issues, e.g. when or where to nest, forage, migrate, shelter or rest [1–5]. Decision outcomes can be crucial for the survival and fitness of individuals in the group and also for group cohesion [6–9]. Thus, successful group living requires the ability to make good collective decisions. Good decisions require good information, which is not always accessible. Good collective decisions additionally require that different, and often conflicting, needs/preferences of group members are reconciled. Therefore, two important factors that influence collective decision-making and decision outcomes are information uncertainty and conflicting preferences.
Usually, decisions are made under uncertainty: that is, individuals lack certain information about at least some aspects of the matter under consideration [4,10,11]. As a consequence, they might make a bad decision. Sharing decision-making in collective decisions can help, because several decision-makers can pool their personal information, and also eliminate individual errors [4,10–15]. Consequently, the risk of making a mistake and settling on a bad option often decreases with the number of decision-makers [11]. This well-known phenomenon is exploited by betting agents, Internet search engines and stock markets [16]. It also plays an important role in collective decision-making in social animals. For example, by sharing decisions widely, swarming honeybees and emigrating ants choose the best available new nest sites [1,5]; shoals of fish successfully avoid predators [17,18]; and flocks of birds find suitable migration routes [12,13,19].
Sharing a decision, however, does not universally lead to a better choice. If the validity of information held by potential decision-makers varies widely, it can be better to go along with the opinion of the one best-informed ‘expert’ and ignore the opinions of all other individuals [20–23]. For example, groups of elephants, primates and dolphins often follow the most experienced group member [24–26].
In addition to involving information uncertainty, most collective decisions, in practice, also involve conflicting ‘preferences’: that is, individual stakeholders in the decision often disagree as to what would constitute desirable decision outcomes in principle [27,28]. For example, in foraging decisions, males and females often prefer different types of forage [29,30]; hungry individuals often prefer different group activities and destinations from tired group members [24,27,31–33]; and young or vulnerable animals often prefer safer areas than older, less vulnerable group members [34,35].
Conflicting preferences are principally different from disagreements caused by uncertain information. This is an important point and is best illustrated by giving a simple human example to the reader, as follows. You might disagree with your partner about which TV channel to choose, either because you like a different TV series from her (conflicting preferences) or because you both like the same show but you think it runs on channel 1 while she thinks it runs on channel 2 (uncertain information). In the first case, at least one of you will have to forgo their own favourite TV programme whatever the collective choice of channel, while in the second case, at least in principle, you could make a choice that pleases both of you. Therefore, during the collective decision-making process, you might behave differently in the first (conflict) situation from the way you behave in the last (uncertainty) situation.
Conflicting preferences are ubiquitous in animal collective decisions, and their survival, fitness and welfare implications can be considerable [36–39]. Conflicts can be momentous and shape the social organization of a species [30,37].
In the following, I briefly review some basic models relating to animal collective decision-making in situations with information uncertainty or in situations with conflicting preferences between group members. This article is not intended to exhaustively review all collective animal decision-making models. Instead, its intention is to give an overview about the different basic types of modelling approaches that have been employed and the questions that they address and raise. Modelling approaches include specific analytical models, quorum models, self-organizing system models, further simulation models, game-theory models and other evolutionary simulation models. The two main questions that animal collective decision-making models address are: (i) How do animals pool personal information efficiently in order to make accurate decisions? and (ii) What individual decision-making strategies evolve in conflict situations and which individuals control the decision outcome?
(Please note that the parameter definitions for each section are separate and the same letters can be used for different parameters in different sections and subsections.)
2. Information uncertainty: how do animals pool information efficiently and make accurate collective decisions?
In 1785, the Marquis de Condorcet suggested that a jury with n (n > 2) members can make more accurate decisions than a solitary decision-maker. His argument was that if every jury member has an equal probability p to be correct which is larger than chance (i.e. p > 0.5), then the probability that a majority (i.e. (n + 1)/2 or more) of jury members decide correctly is
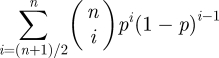 |
(for simplicity it is assumed that n is odd). This probability increases with n and is always larger than the probability p that a single individual is correct. Thus, in situations with information uncertainty, it often pays to share decisions (e.g. by majority vote), since several decision-makers contribute information and also eliminate individual errors [21].
In principle, Condorcet's ‘jury theorem’ also applies to animals [4,12,13,21,22]. Therefore, several animal models have investigated when and how animals pool information through decision-sharing. Most of these models are particularly concerned with mechanisms, since on first sight the relatively limited cognitive abilities of animals relative to humans appear to put more constraints on information pooling than is the case in human juries.
2.1. The quorum models
Empirical data suggest that animals are capable of taking advantage of the jury theorem effect: groups of eusocial insects, fish and birds often collectively make more accurate decisions than they would do individually [1,5,12,17–19,40,41]. However, it is not immediately obvious how animals achieve such increased collective accuracy. One important and influential suggestion has been that animals use a quorum response [7,9,11,42–44], as follows.
In a quorum response, the likelihood that an individual animal decides in favour of a particular option (e.g. to move in a particular direction or to stay in a particular patch) increases with the number of other animals which have already decided in favour of that option (e.g. which are moving in that direction, or which are present in a given patch). Moreover, this increase is usually not linear but step-like. That is, the probability of an animal to choose a particular option increases steeply once a threshold ‘quorum’ of other animals has chosen that option. Deneubourg & Goss [43] and Beckers et al. [44] suggested the following basic function to describe such quorum responses mathematically:
whereby p is the probability that an animal will choose a particular option, A is the number of animals which have already chosen the option, B is the threshold quorum at which the response steeply increases and m (m ≥ 2) determines the steepness of the response (figure 1). This kind of quorum response was used to satisfactorily describe observed collective behaviours in several systems, including foraging ants, moving fish shoals and sheltering cockroaches [7,17,42]. Thus, it appears that even cognitively simple animals can, and do, use quorum responses.
Figure 1.

Example of a quorum response. Grey line shows the quorum response (with m = 5, B = 10); black symbols are simulated data assuming a simple step response by animals (i.e. p = 0 for A < 10 and p = 1 for A > 10) and that animals estimate the size of A with a normal-distributed error (with μ = 0 and σ = 2; note that the error was bounded so that the estimate cannot become negative).
However, the underlying cognitive abilities that are required are not entirely clear. In order to implement the above quorum response function, animals have to not only estimate the number of other individuals A relative to the threshold quorum B, but also weigh this estimate in a nonlinear fashion by exponent m. Even a human would find this a difficult task. It might, thus, be that animals are doing something slightly simpler that closely resembles the above response function. For example, animals might respond in a simple all-or-none fashion. That is, if the number of other animals choosing an option appears to be smaller than a certain threshold (i.e. A < B), they do not choose the option (i.e. p = 0), while if the number appears to be larger than the threshold (i.e. A > B), they choose the option (i.e. p = 1). Since animals are likely to make a certain amount of errors when estimating the number A of other animals choosing an option, their observable (noisy) all-or-none response can resemble the form of the suggested quorum function (figure 1). However, this would imply that the parameter m in the observed response is controlled by the standard deviation of the animal's estimation errors, and animals would, thus, have little scope for adjusting the parameter value profitably to given decision situations (see below).
Sumpter & Pratt [11] investigated whether a quorum response could principally lead to high collective decision accuracy. For this purpose, the authors assumed that animals made a decision between two mutually exclusive options A and B. Further, they assumed that option A was objectively the better (i.e. correct) option but that individual animals had noisy information about the quality of options. As a consequence, animals had only a slightly higher spontaneous probability of choosing option A (pA) than of choosing option B (pB; pA > pB). The authors suggested that under such circumstances animals might benefit from paying attention to the behaviour of conspecifics. They assumed that an animal's probability ProbA(t) of committing to option A at time step t depends on the number of animals A(t) that are already committed to option A, namely in the manner of a quorum response,
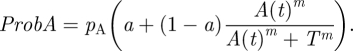 |
Here, T is the threshold quorum at which the response is steepest, m determines the relative steepness of the quorum response and a (a ≤ 1) determines how much an animal is influenced by the choices of other animals (relative to relying on its own personal judgement). The probability ProbB(t) of selecting option B at time step t is calculated in a similar manner. An uncommitted animal can also decide at time step t not yet to commit to any option. The simulated decision-making process begins with n uncommitted animals. At each time step t, uncommitted animals randomly ‘encounter’ a decision option with a fixed probability. They then decide to commit to this option (or not) according to the relevant probability ProbA(t) or ProbB(t), respectively. Once an individual is committed to one option it stays committed. The simulation ends when all animals have committed to an option.
Sumpter & Pratt [11] reported that a steep quorum response (i.e. large m), a medium quorum threshold (i.e. n/4 < T < n/2) and a high reliance on the judgement of others (i.e. a ≈ 0) led to the most accurate decisions (in terms of the proportion of animals that were committed to the better option A at the end of the simulations). Moreover, the proportion of animals committing to option A was considerably higher than it would have been if each animal had decided independently. Thus, a quorum response led indeed to higher collective decision accuracy than did solitary decision-making. However, this higher accuracy came at the price of a slower overall decision speed.
Based on their results, Sumpter & Pratt suggested that animals could fine-tune the parameters of their quorum response (m, a and T) according to their requirements with respect to decision accuracy and speed. In particular, steep responses (i.e. large m) allow animals to make accurate decisions rapidly. Empirical evidence supports the notion that animals adjust parameters a and T according to requirements [45]. However, it might be that animals have relatively little influence on parameter m. This is because the steepness of their response might be mainly limited by their cognitive ability to estimate A(t) error-free (see above and figure 1).
2.2. The effective leadership model
Groups that make collective decisions about communal movement directions can often be very large. For example, flocks of starlings, shoals of fish and swarms of insects can consist of hundreds to thousands of individual animals [46]. In such groups, quorum responses, such as those described above, could not be implemented globally. Apart from overstretching any cognitive and sensory abilities of individuals, any global quorum responses would be far too slow for effective motion. Instead, it is generally accepted that movement decisions in large groups are based on local self-organizing interactions between neighbouring individuals that result in global cohesive and synchronized group movements [47–51]. However, it is not immediately obvious how information-based decisions are made and mediated in such groups.
Couzin et al. [10] asked how information about profitable movement directions is transferred and pooled in large self-organizing groups (of up to 200 animals). They developed a simulation model in which some group members have pertinent information, such as knowledge about the location of a food source [25] or of a migration route [12,19], while others do not. Animals move in continuous, two-dimensional space and at any one time have a position vector Ci(t), a movement direction vector Ei(t) and a speed vi(t). All animals move simultaneously at discrete time steps. The authors make some basic assumptions about the local movement rules that individuals use. In particular, they assume that the highest priority of each individual is to avoid collision by moving away from neighbours that come too close. If there is no immediate danger of collision, the animals desire to move towards, and align movement direction with, local neighbours (animals within a ‘social attraction range’, R) in order to maintain group cohesion. This desired ‘direction of social attraction’ Si(t) is calculated for each individual at each time step, using the relative positions and movement directions of neighbouring individuals. Those individuals that have pertinent information also have a desire to move in the direction suggested by this information (the ‘profitable direction’) Di(t). If Si(t) and Di(t) differ (which they usually do), those informed individuals have to compromise between their desired direction of social attraction and the profitable direction. Their overall desired movement direction then becomes: Mi(t) = (1 − ω)Si(t) + ω Di(t), whereby ω (0 < ω < 1) determines whether an animal moves mainly in the direction of social attraction or in the profitable direction. At each time step, individuals implement their resulting desired direction (i.e. Si(t) for uninformed individuals, and Mi(t) for informed individuals), subject to some error and a maximum permitted turning angle per time.
Couzin et al.'s [10] model shows that, in large self-organizing groups, information can be transferred efficiently without signalling and when group members do not even know which individuals have information. Moreover, the larger the group, the smaller the proportion of informed individuals needed to guide the group. A group can achieve high accuracy already with a small proportion of informed individuals (figure 2). This model is a prime example of how animals can pool the personal information of all group members efficiently without requiring any sophisticated cognitive abilities. The accuracy predictions of Couzin et al.'s model are similar to Condorcet's jury theorem once a minimum number of animals within the group hold information (figure 2). Dyer et al. [14,52] have done some first empirical investigation of Couzin et al.'s model, using humans as subjects and by recreating the model situation. Their observations were in agreement with model predictions.
Figure 2.

Accuracy with which the self-organized group moves in the profitable direction as a function of the number of informed animals within the group. The black solid lines in both graphs show the simulated accuracy in self-organized movements by groups of size (a) n = 200 and (b) n = 10 (after Couzin et al. [10]; note that their measure of accuracy has been rescaled to render possible comparisons with Condorcet accuracy). The grey lines give Condorcet accuracies for juries with the same number of jurors as there were informed animals in the group (note that the slight oscillations arise because juries with odd and even numbers of jurors are included), whereby the dashed grey lines assume an individual juror accuracy equivalent to the effective accuracy of a group in which only one individual is informed ((a) p = 0.53; (b) p = 0.68; lower boundary of comparable Condorcet jury accuracies) and the solid grey lines assume an individual juror accuracy that best fits the simulated group accuracy when there are many informed individuals ((a,b) p = 0.83 in both cases; upper boundary of comparable Condorcet jury accuracies). If the number of informed individuals within a group is very low, the decision accuracy of the self-organized group is poor (solid black line). However, decision accuracy increases much more steeply with the number of informed individuals than does the accuracy in comparable Condorcet juries. Therefore, already from a relatively moderate number of informed individuals onwards (6+ in groups of 200; 2+ in groups of 10), the self-organized groups make decisions that are similar in accuracy to those of Condorcet juries, which consist of relatively well-informed individuals.
In the effective leadership model, there is a trade-off between decision accuracy and maintaining group cohesion (mediated by the parameter ω). That is, the larger the ω, the higher the accuracy with which the group moves in the profitable direction, but also the larger the risk that the group splits. Another potential trade-off could be between decision accuracy and speed. Couzin et al. [10] do not investigate trade-offs between decision accuracy and speed, as does the quorum response model by Sumpter & Pratt [11]. Codling et al. [15] developed a related model that looked specifically at decision speed (i.e. the average time taken to reach a particular spatial target). In this model, all individuals had directional information that was uncertain. The authors found that usually decision speed increased with group size (i.e. the number of informed individuals), unless group size was so large that the necessity of frequent collision avoidance hampered navigation. Since accuracy also increases with the number of informed individuals, there might not be the necessity to trade-off decision speed and accuracy in conflict-free, self-organized movement decisions.
2.3. The independence–interdependence model
Condorcet's jury theorem (see §2.2) applies when individuals have noisy but independent information about what is the best decision option. If there are interdependencies between decision-makers, this can facilitate information pooling but also amplify individual errors [4,21,39,53]. The models discussed so far have not explicitly looked at this problem.
List et al. [4] developed an agent-based model of a decision process based on nest-site selection by swarming honeybees [1] that looked specifically at the relationship between independence and interdependence of judgements in collective animal decisions. The authors assume that n animals participate in the decision-making process between k mutually exclusive decision options that differ in quality. Animals can signal to each other in favour of an option, and by observing the signals of other animals. The model simulates the decision-making process in discrete time steps Δt, as follows.
The authors argue that the probability of an animal starting to signal at time t + Δt in favour of option j is Pj(t + Δt) = (1 − λ)πj + λ fj(t), whereby πj is the probability that the animal discovers information about, and signals in favour of, option j independently of others, fj(t) is the proportion of others that signal in favour of option j and λ (0 ≤ λ≤ 1) is the degree to which an animal depends on the behaviour of others (i.e. its ‘interdependence’). The time period for which an animal signals in favour of an option depends on its assessment of the option's quality. With probability μ, an animal relies on its own independent assessment of the option's quality; with probability 1 − μ, it copies the assessment (i.e. signalling periods) of others. Thus, parameter μ determines the independence of quality assessment. The simulation ends when the group has reached consensus in favour of an option (using a range of consensus criteria).
List et al. [4] reported that a high interdependence λ between individuals with respect to considering particular options, but also a high independence μ of animals with respect to quality assessment, is essential for decision accuracy (i.e. for reaching a consensus in favour of the highest quality option). The authors concluded that, without interdependence, the rapid convergence to a consensus would be undermined and there would not be a ‘snowballing’ of attention to the highest quality option. On the other hand, without independence, a consensus would still emerge, but it would no longer robustly be in favour of the highest quality option. Instead, options that accidentally receive some initial signalling through random fluctuations are likely to become the final consensus choice. The authors suggested that only when independence and interdependence are combined in the right manner can animals achieve high collective accuracy. Empirical evidence on swarming honeybees supports the model predictions [1,40,54].
3. Conflicting preferences: who controls the collective decision outcome and why?
When the preferences of stakeholders in a decision are conflicting, there can also be conflicts about which animals contribute to the collective decision outcome in the role of ‘decision-makers’. This is because the decision-makers can influence the collective decision outcome in favour of their own personal preference and thereby affect the fitness of all stakeholders. Thus, the focus of relevant models is often on who influences decision outcomes and why, and the models usually concentrate on evolutionary questions rather than on mechanisms.
Theoretical research on animal collective decisions in conflict situations is relatively recent. Thus, the topics covered by published models are, as yet, relatively patchy, which renders it difficult to give a coherent thread through this section. I have arranged the models ranging approximately from less to more complex systems, starting with a group-level model (§3.1); followed by pair-coordination models (§3.2); models of decisions in relatively small groups with global interactions(§3.3); and, finally, self-organized system models of collective decisions in large groups with local interactions (§3.4). This arrangement also reflects the historical development of the research area.
3.1. The group-level model
One of the first models that investigated animal collective decision-making in conflict situations examined decisions about the timing of group activities [28]. In many social animals, group members have to synchronize group activities (e.g. resting and foraging) in order to maintain group cohesion. However, the optimal timing of group activities can differ widely between individuals depending on their sex, age, size and physiological state, and suboptimal timing can involve costs to individuals [27,29,30,32,33,36,37,55–58]. Therefore, there are often conflicts of interest about the timing of group activities.
Conradt & Roper [28] developed a model to ask how a group of animals should make a decision about communal activity timing in such conflict situations. They argued that, in principle, decision-making could range from ‘shared’ decisions (i.e. all group members are decision-makers) to ‘dictatorial’ (unshared) decisions which are made by one particular individual (the dictator) and the rest of the group abides by its decision. They then proceed to ask what the costs to a decision-making group are if the decision was either shared or dictatorial (figure 3). The model suggested that, for most biologically relevant assumptions about the shape of the fitness cost function, the expected net costs are lower to a group that makes shared decisions than to a group with a dictator. Conradt & Roper [28], therefore, suggested that shared decision-making should be widespread in social animals.
Figure 3.

Illustration of a collective consensus decision about the timing of a group activity change. Time runs from left to right on the axis. A group of size n starts a group activity at time t = 0. Each individual group member has its own personal optimal time ti for changing activity. The group changes activity collectively at one time. If the decision about this time is made unshared by a single dictator (e.g. the dominant), the group changes group activity at the time that is optimal to the dictator (tdictator). If the decision is made shared (by a ‘majority vote’), the group changes activity when the majority of animals prefer to do so (i.e. at time t(n + 1)/2). The ‘consensus cost’ to an individual (which arises if the individual changes activity at a time that is different from its personal optimal time) increases with the difference between its optimal time and the actual time at which the group (including the individual) changes activity (indicated here by arrows for individual 2). The expected net costs to a group in a dictatorial decision are: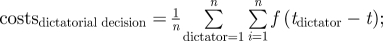 and in a shared decision:
and in a shared decision: 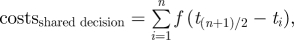 whereby f(Δt) are the fitness costs to an individual of changing activity at a time that is different by Δt from its own optimal time.
whereby f(Δt) are the fitness costs to an individual of changing activity at a time that is different by Δt from its own optimal time.
In support of the predictions, empirical studies have reported shared decision-making, or shared (i.e. transient) leadership, in conflict situations in a wide range of animals, including birds, primates, bats, carnivores and ungulates [6,19,28,59–70]. However, unshared (dictatorial) decision-making, or hierarchical decision-making, has also been observed in several species, including dolphins, elephants, primates and birds [24–26,71,72].
3.2. Pair-coordination models
3.2.1. The leader–follower model
One of the limitations of the group-level model is that it does not look at the problem of collective decision-making from an individual-level point of view. Rands et al. [33] developed a model that goes a step further and investigate collective decision-making about activity synchronization as a dynamic game between two individual animals which are each trying to maximize their own individual survival. In this model, each of the two individuals chooses between resting or foraging during a series of consecutive periods. Foraging is necessary for nutrition uptake, and if the energy reserves of an individual drop below a critical threshold it dies. Thus, foraging offers survival benefits. However, those benefits decrease with the nutritional state of the animal. For an animal close to the critical energy reserve threshold, foraging has large survival benefits, while for an animal with large energy reserves, foraging has only relatively small additional benefits. Foraging also has survival costs. Predation risk is always higher during foraging than during resting. However, this risk is ameliorated when two individuals forage together (e.g. owing to an increase in vigilance; [73]). To summarize, an animal has to balance increased predation risk against its need to replenish reserves when deciding whether to forage or rest, and there are advantages of synchronizing foraging bouts with the other animal. Conflicts of preferences arise because the optimal timings for foraging and resting are likely to differ between the two animals, depending on their individual nutritional states.
Rands et al.'s [33] model defined a strategy π(s,z) that specifies an individual's probability of foraging in any given time step as a function of its own state s and the state z of its partner. They then looked for the evolutionarily stable strategy π* that maximizes an individual's long-term chances of survival, assuming that its partner adopts the same strategy, using an iterated damped best-response procedure.
Rands et al.'s model predicts that the equilibrium behaviour of both individuals is highly synchronized (figure 4), that differences in the energy reserves of the two players develop spontaneously and that the individual with lower reserves emerges as the leader that determines when the pair should forage, while the individual with the higher reserves becomes a follower. This is because an individual with relatively low reserves needs to forage whatever the other individual chooses to do. On the other hand, for an individual with relatively high reserves, it can be advantageous to start to forage when the other individual forages in order to decrease predation risk, rather than to wait until its own reserves have fallen too close to the critical threshold and then, potentially, to have to forage alone. Further, the spontaneously arising differences in energy reserves between the two animals are likely to persist over time. This is because the low-reserve leader will only forage until it has replenished its reserves sufficiently above the critical threshold. On the other hand, the high-reserve follower is likely to forage for longer than it would have done on its own and thereby to boost its reserves further above the critical threshold. As a consequence, an individual is likely to remain the lower reserve partner, and thus the leader, over an extended period. Leadership can, thus, appear to be intransient.
Figure 4.

Predictions of the behaviour of two foragers as a function of their energy reserves according to the leader–follower model (after Rands et al. [33]).
Although Rands et al.'s model predicts (relatively intransient) leaders and followers in collective decisions that involve conflicting preferences, this result does not per se contradict the prediction of shared decision-making derived in the group-level model above. In particular, in a shared decision, the group changes activity when the majority (i.e. (n + 1)/2 animals) prefer to do so. In a group of two animals (i.e. n = 2), it is (n + 1)/2 = 1.5. Thus, in a shared decision, a group of two can use 1 or 2 as the ‘cut-off’ point. That is, the group should always move from resting to foraging either when the first individual prefers to do so or when the second does, and it should do so independently of the individuals' identities (or dominance rank). In Rands et al.'s model, the group follows the first individual that wants to forage, regardless of its identity. Thus, the decision is strictly speaking shared.
Rands et al.'s prediction of reserve-dependent leader behaviour is supported by empirical studies on fish, insects and ungulates [31,35,57,74,75]. Sueur et al. [70] developed Rands et al.'s [33] idea further in a dynamic model based on empirical data of needs of group members in larger groups in primates. They reported similar results, namely that the individual with the lowest reserves emerges as a leader of the group and that this leadership can be fairly consistent owing to uneven needs of group members.
3.2.2. The pair-synchronization model
Dostálková & Špinka [27] also developed an individual-level model of activity synchronization (here, of departure timing) for pairs of two animals. This model makes even simpler assumptions than the follower–leader model by Rands et al. [33]. Similarly to the group-level model, Dostálková & Špinka [27] assumed that the two animals A and B gain by synchronizing their departure but differ in their optimal timing (TA and TB, respectively), and that each animal incurs synchronization costs if it departs at a time that is not optimal to itself, whereby costs can differ between individuals departing too early or too late. Thus, there is a conflict of preferences with respect to departure timing.
The authors argued that, in order to be able to synchronize departure, an animal has to be prepared to depart during a time window around its optimal time. The strategy (RA,WA) defines this time window as (TA − RA to TA + WA) for animal A. If animal B departs before TA − RA, animal A will not depart. If animal B departs within animal A's time window, then animal A departs together with animal B. If animal B has not departed before time TA + WA, animal A departs at time TA + WA. Animal B then either follows or stays behind, according to its own time window (TB − RB to TB + WB). Thus, depending on its strategy (RA,WA), animal A might pay an early or late departure cost and might receive grouping benefit. Dostálková & Špinka [27] looked for the best strategy (R*,W*) at which the net expected gains to an individual are maximal.
The model predicts for the majority of biologically relevant parameter values that the best strategy for an animal is to be ready to join the other individual in foraging before its own optimal time (i.e. R* > 0), but not necessarily to wait beyond that optimal time (i.e. W* = 0) unless waiting is cheap (figure 5). Waiting is usually disadvantageous because both animals might be waiting for each other long after both their optimal times have elapsed.
Figure 5.

Best mutual strategies of individuals in pair synchronizations depending on the synchronization costs relative to grouping benefits (horizontal axis) and whether the costs are higher for departing too early or too late (vertical axis; after Dostálková & Špinka [27]). If the synchronization costs are large relative to grouping benefits, full synchronization might no longer be the best strategy. If departing too early is less costly than departing too late, animals should be ready to depart before their optimal time but not wait beyond it. If departing too late is less costly than departing too early, animals should be ready to depart before their optimal time but also to wait beyond it.
Thus, the pair-synchronization model predicts, like the leader–follower model, that a group of two should change activity synchronously when the first individual does so, and that the decision is, strictly speaking (see above), a shared decision. This ‘follow the first’ should happen even if there is no difference in energy reserves between the two individuals. Instead, it is simply a consequence of temporal asymmetry in information. That is, an animal can gather information from its partner's behaviour about the time when its partner's time window has elapsed (i.e. time T + W), but not when the window starts (i.e. time T − R). It is this asymmetry in information which renders waiting disadvantageous.
3.3. Models of collective decisions in relatively small groups with global interactions
The leader–follower model and the pair-synchronization model investigate collective decisions in groups of two animals. However, collective decisions taken by two individuals can differ from decisions in larger groups [38]. In particular, shared (majority) decision-making is difficult to define in groups of two (see above), and the evolution of leader/follower roles is likely to be principally different from that in larger groups [76,77]. Thus, to investigate collective decision-making in conflict situations further, models of groups with more than two individuals are required. In this subsection, models are presented of collective decisions in groups which are larger than two but still small enough so that all group members can, in principle, interact with all other group members (i.e. global interactions are possible).
3.3.1. Activity synchronization between three animals
Conradt & Roper [78] developed a model for groups of three animals about decisions on the timing of activity changes that require departure. The model assumes that each animal has its own optimal time ti to depart which differs between animals (see §2.1 and figure 3). An animal gains grouping benefits in a cohesive group depending on group size (GB2 or GB3, respectively; [73]). Cohesion requires departure synchronization with others. However, departing earlier or later than is optimal incurs a ‘synchronization cost’ (SCearly or SClate, respectively). Thus, there is an incentive to synchronize departure but also a conflict of preferences. Conradt & Roper [78] examined the evolutionarily stable sets of strategies which animals A, B and C are likely to adopt in those circumstances, as follows.
The authors argued that each individual animal has the choice between two behaviours. An animal can either play ‘INSIST’ on its own preference and depart at its own optimal time, or it can play ‘GIVE-IN’ and do what at least one other individual does. The authors, therefore, defined an animal's strategy ri as the probability that the animal plays INSIST (whereby 1 − ri is the probability that it plays GIVE-IN). Thus, the set of strategies for animals A, B and C is given by (rA, rB, rC). Synchronization of departure and group coherence (and, thus, the related costs and benefits to individuals) depend on the set of strategies (rA, rB, rC). For example, if all three animals play INSIST, they will all depart at their own optimal times and become solitary. Conradt & Roper [78] assumed that each animal tries to maximize its own individual net gains and looked for the resulting stable set of strategies (rA*, rB*, rC*).
The model predicts that shared and dictatorial decision-making (by a dominant individual) are both evolutionarily stable strategies (ESSs) in groups of three (figure 6), and which strategy evolves depends on the population's starting position and the parameter values for costs and benefits. The model further predicts that shared decision-making is particularly likely to evolve when groups are heterogeneous in composition (i.e. tA, tB and tC differ widely); when synchronization costs (SCs) are large; when alternative decision outcomes differ widely in potential costs (i.e. SCearly and SClate differ); when grouping benefits (GBs) are small; or when groups are close to, or above, optimal size (i.e. GB2 > GB3). Empirical data to test these predictions are still scarce.
Figure 6.

Phenotypic evolution of decision-making about activity synchronization in groups of three (after Conradt & Roper [78]). Each trilinear coordinate system (TCS) represents populations consisting of three different phenotypes. Different TCSs are shown closely together to indicate the overall dynamics. Arrows show the directions in which the system evolves. Broken arrows indicate where the system evolves from one TCS to another. For clarity, in shaded areas, the direction of evolution is not shown, and the dynamics shown are not exhaustive (e.g. substituting ‘sub-majority’ for ‘super-majority’ phenotypes leads to similar dynamics).
3.3.2. Synchronization of movement destination between three animals
So far, the models that investigate collective decisions involving conflicts have looked at decisions about activity synchronization. Another important area for collective decision-making in animals, in which conflicts arise, is decisions about movement destinations [2]. In order to stay cohesive and gain grouping benefits, individuals have to agree group movement destinations [10,46,47,73]. However, the optimal destination often differs between individuals [3,24,29,30,37,79]. Conradt & Roper [80] developed a model of decisions about movement destination in groups of three animals, as follows.
The model assumes that animals have to decide between two possible movement destinations (e.g. two foraging patches), and that two animals prefer one of the destinations (majority-type animals) while the third animal prefers the other destination (minority-type animal). As in the last model, an animal in a cohesive group gains grouping benefits (GB2 or GB3, respectively), whereby cohesiveness requires moving to the same destination. On the other hand, if an animal moves to a non-preferred destination, it incurs a cost (‘consensus cost’). Thus, while it is advantageous for animals to move together to one destination, there is a conflict of interest between majority-type and minority-type animals as to where to move to.
The model consists of two parts. The first part simulates the movements of all three animals until they reach one of the two destinations, depending on their behavioural strategies. The outcome is then translated into expected individual net gains, depending on grouping benefits, consensus costs and the behavioural strategies of the three animals. The second, game-theory, part determines the evolutionarily stable sets of strategies based on those individual net gains. Details are as follows.
The movement simulations are based on Couzin et al.'s [10] model (see §2.2). Since each animal now has a preferred destination, each animal compromises between its attraction to other animals (‘direction of social attraction’: vector Si(t)) and the direction towards its preferred destination (vector Di(t)): Mi(t)= (1 − ωi) · Si(t) + ωi · Di(t) (see §2.2 for further details). Here, ωi (0 < ωi < 1) is the degree of assertiveness that the animal employs. That is, if an animal is highly assertive (ωi ≈ 1), it mainly moves in its preferred direction Di(t). If it is little assertive (ωi ≈ 0), it moves mainly in the direction of social attraction Si(t). The simulation ends when all animals have reached one of the two destinations and the resulting individual net gains are calculated for different sets of strategies by the three animals (ωA, ωB and ωC). These expected individual net gains are then used in the game-theory part of the model to determine evolutionarily stable sets of strategies (ω*A, ω*B and ω*C).
The model predicts, like the last one, that shared and dictatorial decision-making are both ESSs in groups of three (figure 7). In particular, when dictatorial decisions evolve, it is often that animal, the ‘dictator’, which would otherwise face the highest consensus costs. This result is similar to those of Rands et al. [33].
Figure 7.

Schematic of the predictions following from the group-of-three model on synchronisation of movement destinations (after Conradt & Roper 2009 [80]). The area above the dotted line refers to groups that are above optimal group size; the area below the dotted line refers to groups of suboptimal group size.
Further, the model predicts that shared decision-making is particularly likely to evolve when conflicts are low relative to grouping benefits. This prediction is the opposite of the predictions of the last model that investigated decisions about activity synchronization. In the past model, decision-sharing was particularly likely to evolve when consensus costs were high relative to grouping benefits. The reason for this apparent contradiction lies in the different nature of decisions about activity synchronization (with continuous cost functions) and decisions about movement destinations (with disjunct cost functions; [81]). This is best illustrated by an example, as follows.
Imagine making a decision with a friend about going to a restaurant. Assume that you prefer to go at 19.00 h, and your friend at 20.00 h. If you compromise and agree to go at 19.30 h, neither of you is too much inconvenienced. You should be particularly ready to compromise (and share the decision), if synchronization costs are high and grouping benefits are small, because by such a compromise you can stop the group from splitting.
Now assume further that you prefer to go to a Chinese restaurant in the north of town and your friend prefers to go to an Indian restaurant in the east of town. This time you cannot compromise in the same manner as before, because in the northeast of town there are no restaurants but there is a petrol station, which is of no use to either you or your friend. Such circumstances usually require ‘hard’ decisions, of an ‘either/or’ kind, to be made. Here, if grouping benefits are large relative to consensus costs, no group member wishes to risk group fragmentation. Therefore, a not-to-high level of assertiveness is advantageous. This results in shared decision-making. On the other hand, if grouping benefits are low relative to consensus costs, risking group fragmentation rather than moving towards a less preferred target becomes an evolutionary option. It will then benefit some individuals to be highly assertive. In response, selection might favour other individuals to be unassertive in order to avoid group fragmentation. The result is dictatorial (unshared) decision-making.
3.3.3. Leaders in groups of up to 10
Johnstone & Manica [76] investigated the evolution of intrinsic leadership strategies in groups with up to 10 animals. The authors assumed that group members made a large number of consecutive decisions together, whereby each individual had a different preference from any other group member. Animals had an incentive to stay together, and an animal's decision gains increased proportionally to the number of other group members that settled on the same decision option as itself. However, the factor of this increase was higher if the animal settled together with others on its preferred option than when it settled on any other option.
The authors assumed that each animal could adopt an individual strategy λi, whereby, with probability λi, the animal always chooses its own preferred choice, and, with probability 1 − λi, it always copies the most recent choice of a randomly chosen group member. Thus, λi is an animal's intrinsic propensity to leadership. In order to find the evolutionarily stable sets of strategies, the authors used numerical simulations in which the increase/decrease of frequencies of different strategies depended on respective individual gains.
Johnstone & Manica [76] found for nearly all group sizes and conflict conditions that stable dimorphisms of extreme leaders (i.e. λ ≈ 1) and followers (i.e. λ ≈ 0) evolved. Only in populations with very small group size, and at low to moderate conflict level, did polymorphisms evolve. The mainly observed stable dimorphisms were maintained by frequency-dependent selection, since leaders did relatively well in groups that consisted mainly of followers, but poorly when leader-type frequency was high. The proportion of leaders in the population increased with the degree of conflict, while the degree of coordination decreased with the degree of conflict. Thus, the authors suggest that leader and follower types evolve spontaneously in populations even in the absence of physiological differences between individuals (compare with Rands et al. [33]). This differs somewhat from the group-of-three models above, which suggest that shared decision-makers can invade frequency-dependent selected leader–follower societies without asymmetries. However, Johnstone & Manica's [76] model does not allow for strategies that are likely to lead to shared decision-making (such as copy the majority). It remains to be seen whether and how such strategies would modify the model's predictions.
3.4. Self-organized system models of collective decisions in large groups with local interactions
3.4.1. The leading-according-to-need model
Modelling collective decision-making is a complex problem, because the possibilities of interactions between animals increase exponentially with the number of animals. So far, the discussed models of collective decision-making in conflict situations have concentrated on relatively small groups of maximally 10 animals. In the following, I will introduce two models that deal with larger groups.
Conradt et al. [82] investigated collective decisions between two mutually exclusive movement destinations in large self-organizing groups (of up to 100 animals). The authors assumed that some group members prefer one of the destinations (and gain more benefits if they actually arrive at this destination), while the other group members prefer the other destination (and gain more benefits if they arrive at that destination). Thus, there is a conflict of interest between group members with respect to movement destination. Moreover, animals gain grouping benefits if the group remains cohesive [73].
Conradt et al.'s [82] model is closely based on Couzin et al.'s [10] model, with some important differences. First, Conradt et al. [82] introduced conflict between individuals into the movement decision by assuming different benefits to different animals from different decision outcomes. Second, they assumed that all individuals have knowledge about their preferred option. Third, they looked at decisions between movement destinations rather than movement directions. Finally, they allowed individuals to modify their individual behaviour (i.e. the parameters of their local movement rules) according to their personal interests, as follows.
As in a previous model (§3.3.2), an animal's overall desired movement direction at any one time is a compromise between its ‘direction of social attraction’ and its preferred direction: Mi(t)= (1 − ωi) Si(t) + ωi Di(t); and ωi (0 < ωi < 1) determines whether an animal mainly moves in the direction of social attraction or in its personally preferred direction (see §3.4.2. for further details). Conradt et al. [82] termed the parameter ωi the ‘degree of assertiveness’ of individual i. Note that individual animals can vary in their parameter ωi. The end of the simulations is determined by all animals having reached one of the two possible destinations.
Conradt et al.'s [82] model makes some interesting predictions for conflict resolutions in large self-organizing groups. Even in large groups, individual animals can increase their influence on collective decisions by strategically changing simple behavioural parameters, in particular their assertiveness, movement speed and social attraction range. However, they do so at the expense of an increased risk of group fragmentation and a decrease in movement efficiency (i.e. decision speed). The authors argue that the resulting trade-offs faced by each animal render it likely that group movements are led by those animals for which reaching a particular destination is either most crucial or group cohesion is least important. They term this phenomenon ‘leading according to need’ and ‘leading according to social indifference', respectively. Both kinds of leading can occur in the absence of knowledge of or communication about the needs of other group members and without the assumption of altruistic cooperation. Leading according to need is a similar prediction to that by Rands et al.'s [33] that the animal with the greatest need to forage emerges as the leader in groups of two (§3.2). Empirical studies on shoals of fish and herds of ungulates support these predictions [31,35,73].
3.4.2. The flock landing model
Another self-organizing system simulation model that looks at decisions in large groups (here, up to greater than 1000 animals) is a model by Bhattacharya & Vicsek [83]. They investigated the collective landing decisions of bird flocks (i.e. decisions about collective activity changes). The model assumes that different birds within a flock have different times ti when they prefer to land, depending on their personal energy reserves. Those times are normally distributed around a mean landing time Tmean with a standard deviation σ. Thus, there is a conflict of interest with respect to the timing of the landing. However, birds also try to maintain group cohesion.
The model by Bhattacharya & Vicsek [83] is based on an influential motion model of self-propelled particles by Vicsek et al. [49]. Birds move within a flock in discrete time steps within a continuous three-dimensional space above a flat landing surface. The authors decouple the horizontal and vertical movements in the model. On the horizontal plane, flying birds move in the average direction of motion of all neighbouring animals within a horizontal social radius R (that can also include already landed birds). Group cohesion is maintained by an attraction of animals back towards the group's horizontal centre of mass if they stray too far from it.
The vertical movement of a flying bird depends on whether the bird is in an internal state of cruising or landing at the time. If the bird is in a state of cruising, its vertical movements follow a random walk around a normal flight height z0 that is bounded between z0 + c and z0 − c (where z0 ≫c). If the bird is in a state of landing, it decreases its height by v per time step. If a bird reaches a height of 0, it lands. Once a bird has landed, it stays landed and motionless.
Flying birds can change from an internal state of cruising to landing but also back from landing to cruising. At any one time, a flying bird's internal state depends on its own preferred landing time ti and the motivation to land of its neighbours within its horizontal social radius, whereby a factor J determines how much the bird adjusts its internal state to the behaviour of others versus its own preference. For example, if J is large, then the bird's internal state is mainly influenced by its neighbours and little by its own optimal time.
Bhattacharya & Vicsek [83] reported that the synchronization of landing times by birds in the flock increases with the interdependence factor J, as expected. Remarkable is that the mean actual landing time of the flock also increases with the interdependence of birds, and well beyond the mean optimal landing time Tmean (figure 8). That is, the stronger birds are influenced in their landing decisions by the behaviour of their neighbours, the later the flock lands, and well beyond the time when it would have been optimal for the majority of flock members. The reason for this appears to be a delay between the time of a bird's change in personal preference (ti) and the time when this change in personal preference starts to communicate itself to other birds (through actual downward movements of the bird). The more each bird is influenced by the behaviour of other birds, the longer this time delay is. This phenomenon is related to Dostálková & Špinka's [27] observation that a temporal asymmetry in information often makes pairs of animals change activities later than is good for either animal (§3.2.2). Therefore, intermediate values of interdependence J between animals might be optimal, since the landing is then sufficiently well synchronized but not too delayed in time. The importance of a well-adjusted degree of interindividual interdependence in collective decisions has also been suggested by List et al.'s [4] model (§2.3).
Figure 8.

The proportion of birds in a flock that have landed as a function of time and of interdependence between neighbouring birds (after Bhattacharya & Vicsek [83]). If the interdependence factor between birds J is zero, all birds land independently of each other at their optimal times (solid line). If the interdependence factor is medium (dashed line) or large (dotted line), the landing slope is steeper and the flock's landing is much more synchronized. However, the larger the interdependence factor J, the later the average landing time (see main text for further details).
4. Discussion
Apart from decision-making in eusocial insects, animal collective decision-making has only started to attract wider attention in the past decade. Since then, a range of very different modelling methods has been developed to unravel complex collective decision-making processes and strategies in animals. These diverse modelling methods have nevertheless several fundamental aspects in common. With only one exception [28], all models are, in principle, agent-based models. They start with a hypothesis about the decision-making strategies of individual animals in a group, and then proceed by making assumptions about the integration process of individual strategies into an aggregated collective decision outcome.
The hypotheses about individual strategies include quorum response strategies [11,43,44], state-dependent strategies [33,82,83], assertiveness/submission strategies [76,78,80] and compromise strategies [4,10,15,27,82,83]. Strategies can be continuous [10,11,43,44,82] or discrete [78,80]. Some of the hypotheses on individual strategies are based on descriptions of observed behaviours [4,11,43,44], while the remainder are founded on theoretical considerations. In order to be able to judge how appropriate those hypotheses are (and, thus, the respective modelling approaches), more empirical data are required.
The integration process is usually modelled through simulations, owing to its complexity. Two main types of integration processes can be distinguished: those that consider global interactions of individuals within relatively small groups [11,27,33,43,44,78,80] and those that assume local interactions of individuals (self-organization) within relatively large groups [4,10,15,76,82,83]. Again, since very little is known empirically about integration processes, it is difficult to judge between different methods. A potentially very useful method, which has not yet been employed on animal collective decision-making, is the voter model [84].
Models investigating animal collective decisions in situations with information uncertainty often ask mechanistic questions about the effective dissemination of information within groups [10,15], but also functional questions relating to collective decision accuracy [4,11,43,44]. Models examining animal collective decisions in situations that involve conflicts of interest between group members usually ask functional and evolutionary questions, particularly about which strategies animals are likely to use in order to maximize their own benefits [27,28,33,76,78,80,82]. The evolutionary aspects of those models usually require some form of game-theory modelling. Some of those game-theory models use analytical solution methods [78]. However, because of the high complexity of n-player games, several models have to fall back on numerical and simulation solution methods [33,76,80]. The game-theory parts of all the models are tailor-made to the biological assumptions of the system. Therefore, without more empirical information, it is not possible to judge between methods, and probably different methods are appropriate for different biological situations.
Despite the very different modelling approaches and the different types of collective decisions that are addressed, results have started to show a reassuringly coherent picture. It is particularly reassuring that models dealing with relatively small groups of two or three animals make similar predictions to models dealing with large groups of up to 1000 animals.
In situations with information uncertainty, the models suggest that relatively simple cognitive mechanism can lead to effective information dissemination, which are in reach of most social animal species. An important principle that also emerges is the relationship between speed and accuracy of decision-making. Often, behaviours that increase decision accuracy do so at the expense of decision speed [4]. However, this is not invariably so, and there appear to be mechanisms, at least in principle, whereby animals can attain high decision accuracy without great loss of decision speed [4,11]. Further, the information uncertainty models highlight the importance of the right balance of interdependence and independence between individuals [4,83]. If interdependence is too low, information pooling is impaired, animals often do not reach consensus, they do not benefit from other animals' knowledge, and group cohesion and synchronization of group behaviour can be weak. On the other hand, if independence is too low, groups can quickly congregate on suboptimal decision choices. Two models also highlight the problem of temporally asymmetric information, and indicate that too high interdependence of individuals can be particularly disadvantageous in such asymmetric situations [27,83].
In situations with conflicting preferences of individuals (i.e. conflicts of interest), models generally predict a relatively high degree of decision-sharing between individuals [28], transient leadership [27,33], and most importantly leadership according to needs and physiological status [33,82]. That is, those animals which have the lowest energy reserves, and the greatest needs with respect to the decision outcome, are likely to influence collective decision outcomes most or even emerge as single leaders. One important aspect that all conflict-of-interest models highlight is the trade-off that animals face between maintaining group cohesion and influencing the decision outcome in the direction of their own personal preferences. There does not appear to be any obvious strategy whereby animals can maximize group cohesion and their own influence at the same time. Thus, the more they use a strategy that influences decision outcomes in their own favour, the more they risk group fragmentation, and vice versa.
Some of the conflict models also stress that it is important whether consensus costs are distributed continuously or discretely across decision options, and that continuous distributions can lead to exactly opposite evolutionary predictions from discrete distributions [78,80]. For example, in collective decisions about when a group leaves a foraging patch and which patch it goes to next, individual animals might use completely different strategies for the two different aspects of the decision. As a consequence, the timing part of the decision (when to leave) could, for example, be a shared decision, while the destination part of the same decision (where to go to) might at the same time be made dictatorially by the dominant [81].
Despite the great progress that has been made in the past 10 years in understanding animal collective decision-making, there remains one big gap in our knowledge. The above models give us a good idea about how animals can, and are likely to, behave in situations when information is uncertain, and in situations when there are conflicts of interests. However, most decisions in the real world involve both factors simultaneously. Moreover, it is highly likely that these two factors interact in a complex, non-trivial manner [27,53,77]. Thus, how can, and should, animals behave during collective decisions that involve uncertain information and conflicts of interest simultaneously? For example, how should quorum responses be modified in case of conflict? What are the options to maximize gains in self-organizing systems, if there is conflict but it is also unclear which individuals hold information? How does information uncertainty modify evolutionary games in conflict decisions? Here is a broad scope for future modelling.
Another area in which more research is urgently needed is empirical work on animal collective decision-making. Although the last few years have seen a surge in empirical work, this is still lagging behind the advances in modelling, and many model assumptions and predictions are as yet untested. One interesting question is why so many studies report unshared and dictatorial decision-making, while the models predominantly predict shared decision-making. Obviously, the models are missing some aspects of the problem about which only more empirical data can inform us. In particular, secondary considerations about maintaining long-term good social relationships might prohibit some animals from contributing fully to decisions [24], and social network structure is likely to play a role [85]. Much more empirical information and experimental testing of hypotheses is needed (see [86] for a review on primates).
Acknowledgements
L.C. was supported by a Royal Society URF grant. I thank Edward Codling for the organization of a very interesting conference, and three reviewers for comments on the manuscript.
References
- 1.Seeley T. D., Buhrman S. C. 1999. Group decision making in swarms of honey bees. Behav. Ecol. Sociobiol. 45, 19–31 10.1007/s002650050536 (doi:10.1007/s002650050536) [DOI] [Google Scholar]
- 2.Conradt L., Roper T. J. 2005. Consensus decision making in animals. Trends Ecol. Evol. 20, 449–456 10.1016/j.tree.2005.05.008 (doi:10.1016/j.tree.2005.05.008) [DOI] [PubMed] [Google Scholar]
- 3.Trillmich J., Fichtel C., Kappeler P. M. 2004. Coordination of group movements in wild Verreaux's sifakas Propithecus verreauxi. Behaviour 141, 1103–1120 10.1163/1568539042664579 (doi:10.1163/1568539042664579) [DOI] [Google Scholar]
- 4.List C., Elsholtz C., Seeley T. D. 2009. Independence and interdependence in collective decision making: an agent-based model of nest-site choice by honeybee swarms. Phil. Trans. R. Soc. B 364, 755–762 10.1098/rstb.2008.0277 (doi:10.1098/rstb.2008.0277) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Franks N. R., Dechaume-Moncharmont F. X., Hanmore E., Reynolds J. K. 2009. Speed versus accuracy in decision-making ants: expediting politics and policy implementation. Phil. Trans. R. Soc. B 364, 845–852 10.1098/rstb.2008.0224 (doi:10.1098/rstb.2008.0224) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kerth G., Ebert C., Schmidtke C. 2006. Group decision making in fission-fusion societies: evidence from two-field experiments in Bechstein's bats. Proc. R. Soc. B 273, 2785–2790 10.1098/rspb.2006.3647 (doi:10.1098/rspb.2006.3647) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ame J. M., Halloy J., Rivault C., Detrain C., Deneubourg J. L. 2006. Collegial decision making based on social amplification leads to optimal group formation. Proc. Natl Acad. Sci. USA 103, 5835–5840 10.1073/pnas.0507877103 (doi:10.1073/pnas.0507877103) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Sueur C., Petit O. 2008. Shared or unshared consensus decision in macaques? Behav. Process. 78, 84–92 10.1016/j.beproc.2008.01.004 (doi:10.1016/j.beproc.2008.01.004) [DOI] [PubMed] [Google Scholar]
- 9.Sumpter D. J. T. 2010. Collective animal behavior. Princeton, NJ: Princeton University Press [Google Scholar]
- 10.Couzin I. D., Krause J., Franks N. R., Levin S. A. 2005. Effective leadership and decision-making in animal groups on the move. Nature 433, 513–516 10.1038/nature03236 (doi:10.1038/nature03236) [DOI] [PubMed] [Google Scholar]
- 11.Sumpter D. J. T., Pratt S. C. 2009. Quorum responses and consensus decision making. Phil. Trans. R. Soc. B 364, 743–753 10.1098/rstb.2008.0204 (doi:10.1098/rstb.2008.0204) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Wallraff H. G. 1978. Social interrelations involved in migratory orientation of birds—possible contribution of field studies. Oikos 30, 401–404 10.2307/3543490 (doi:10.2307/3543490) [DOI] [Google Scholar]
- 13.Simons A. M. 2004. Many wrongs: the advantage of group navigation. Trends Ecol. Evol. 19, 453–455 10.1016/j.tree.2004.07.001 (doi:10.1016/j.tree.2004.07.001) [DOI] [PubMed] [Google Scholar]
- 14.Dyer J. R. G., Johansson A., Helbing D., Couzin I. D., Krause J. 2009. Leadership, consensus decision making and collective behaviour in humans. Phil. Trans. R. Soc. B 364, 781–789 10.1098/rstb.2008.0233 (doi:10.1098/rstb.2008.0233) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Codling E. A., Pitchford J. W., Simpson S. D. 2007. Group navigation and the ‘many-wrongs principle’ in models of animal movement. Ecology 88, 1864–1870 10.1890/06-0854.1 (doi:10.1890/06-0854.1) [DOI] [PubMed] [Google Scholar]
- 16.Surowiecki J. 2004. The wisdom of crowds. New York, NY: Doubleday [Google Scholar]
- 17.Ward A. J. W., Herbert-Read J. E., Sumpter D. J. T., Krause J. 2011. Fast and accurate decisions through collective vigilance in fish shoals. Proc. Natl Acad. Sci. USA 108, 2312–2315 10.1073/pnas.1007102108 (doi:10.1073/pnas.1007102108) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Ward A. J. W., Sumpter D. J. T., Couzin L. D., Hart P. J. B., Krause J. 2008. Quorum decision-making facilitates information transfer in fish shoals. Proc. Natl Acad. Sci. USA 105, 6948–6953 10.1073/pnas.0710344105 (doi:10.1073/pnas.0710344105) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Biro D., Sumpter D. J. T., Meade J., Guilford T. 2006. From compromise to leadership in pigeon homing. Curr. Biol. 16, 2123–2128 10.1016/j.cub.2006.08.087 (doi:10.1016/j.cub.2006.08.087) [DOI] [PubMed] [Google Scholar]
- 20.Reebs S. G. 2000. Can a minority of informed leaders determine the foraging movements of a fish shoal? Anim. Behav. 59, 403–409 10.1006/anbe.1999.1314 (doi:10.1006/anbe.1999.1314) [DOI] [PubMed] [Google Scholar]
- 21.List C. 2004. Democracy in animal groups: a political science perspective. Trends Ecol. Evol. 19, 168–169 10.1016/j.tree.2004.02.004 (doi:10.1016/j.tree.2004.02.004) [DOI] [PubMed] [Google Scholar]
- 22.Katsikopoulos K. V., King A. J. 2011. Swarm intelligence in animal groups: when can a collective out-perform an expert? PLoS ONE 5, e15505 10.1371/journal.pone.0015505 (doi:10.1371/journal.pone.0015505) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Katsikopoulos K. V., Martignon L. 2006. Naive heuristics for paired comparisons: some results on their relative accuracy. J. Math. Psychol. 50, 488–494 10.1016/j.jmp.2006.06.001 (doi:10.1016/j.jmp.2006.06.001) [DOI] [Google Scholar]
- 24.King A. J., Douglas C. M. S., Huchard E., Isaac N. J. B., Cowlishaw G. 2008. Dominance and affiliation mediate despotism in a social primate. Curr. Biol. 18, 1833–1838 10.1016/j.cub.2008.10.048 (doi:10.1016/j.cub.2008.10.048) [DOI] [PubMed] [Google Scholar]
- 25.McComb K., Shannon G., Durant S. M., Sayialel K., Slotow R., Poole J., Moss C. 2011. Leadership in elephants: the adaptive value of age. Proc. R. Soc. B 278, 3270–3276 10.1098/rspb.2011.0168 (doi:10.1098/rspb.2011.0168) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lusseau D., Conradt L. 2009. The emergence of unshared consensus decisions in bottlenose dolphins. Behav. Ecol. Sociobiol. 63, 1067–1077 10.1007/s00265-009-0740-7 (doi:10.1007/s00265-009-0740-7) [DOI] [Google Scholar]
- 27.Dostálková I., Špinka M. 2007. Synchronization of behaviour in pairs: the role of communication and consequences in timing. Anim. Behav. 74, 1735–1742 10.1016/j.anbehav.2007.04.014 (doi:10.1016/j.anbehav.2007.04.014) [DOI] [Google Scholar]
- 28.Conradt L., Roper T. J. 2003. Group decision-making in animals. Nature 421, 155–158 10.1038/nature01294 (doi:10.1038/nature01294) [DOI] [PubMed] [Google Scholar]
- 29.Clutton-Brock T. H., Guinness F. E., Albon S. D. 1982. Red deer: behaviour and ecology of two sexes. Chicago, IL: University of Chicago Press [Google Scholar]
- 30.Ruckstuhl K. E., Neuhaus P. 2002. Sexual segregation in ungulates: a comparative test of three hypotheses. Biol. Rev. 77, 77–96 10.1017/S1464793101005814 (doi:10.1017/S1464793101005814) [DOI] [PubMed] [Google Scholar]
- 31.Krause J., Bumann D., Todt D. 1992. Relationship between the position preference and nutritional state of individuals in schools of Juvenile Roach Rutilus-Rutilus. Behav. Ecol. Sociobiol. 30, 177–180 10.1007/BF00166700 (doi:10.1007/BF00166700) [DOI] [Google Scholar]
- 32.Conradt L., Roper T. J. 2000. Activity synchrony and social cohesion: a fission-fusion model. Proc. R. Soc. Lond. B 267, 2213–2218 10.1098/rspb.2000.1271 (doi:10.1098/rspb.2000.1271) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Rands S. A., Cowlishaw G., Pettifor R. A., Rowcliffe J. M., Johnstone R. A. 2003. Spontaneous emergence of leaders and followers in foraging pairs. Nature 423, 432–434 10.1038/nature01630 (doi:10.1038/nature01630) [DOI] [PubMed] [Google Scholar]
- 34.Lingle S., Pellis S. M., Wilson W. F. 2005. Interspecific variation in antipredator behaviour leads to differential vulnerability of mule deer and white-tailed deer fawns early in life. J. Anim. Ecol. 74, 1140–1149 10.1111/j.1365-2656.2005.01014.x (doi:10.1111/j.1365-2656.2005.01014.x) [DOI] [Google Scholar]
- 35.Prins H. H. T. 1996. Ecology and behaviour of the African Buffalo. London, UK: Chapman & Hall [Google Scholar]
- 36.Conradt L. 1998. Could asynchrony in activity between the sexes cause intersexual social segregation in ruminants? Proc. R. Soc. Lond. B 265, 1359–1363 10.1098/rspb.1998.0442 (doi:10.1098/rspb.1998.0442) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Ruckstuhl K. E., Neuhaus P. 2000. Sexual segregation in ungulates: a new approach. Behaviour 137, 361–377 10.1163/156853900502123 (doi:10.1163/156853900502123) [DOI] [Google Scholar]
- 38.Conradt L., List C. 2009. Group decisions in humans and animals: a survey. Phil. Trans. R. Soc. B 364, 719–742 10.1098/rstb.2008.0276 (doi:10.1098/rstb.2008.0276) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Hix S., Noury A., Roland G. 2009. Voting patterns and alliance formation in the European Parliament. Phil. Trans. R. Soc. B 364, 821–831 10.1098/rstb.2008.0263 (doi:10.1098/rstb.2008.0263) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Seeley T. D., Buhrman S. C. 2001. Nest-site selection in honey bees: how well do swarms implement the ‘best-of-N’ decision rule? Behav. Ecol. Sociobiol. 49, 416–427 10.1007/s002650000299 (doi:10.1007/s002650000299) [DOI] [Google Scholar]
- 41.Krause J., Ruxton G. D., Krause S. 2010. Swarm intelligence in animals and humans. Trends Ecol. Evol. 25, 28–34 10.1016/j.tree.2009.06.016 (doi:10.1016/j.tree.2009.06.016) [DOI] [PubMed] [Google Scholar]
- 42.Deneubourg J. L., Aron S., Goss S., Pasteels J. M. 1990. The selforganising exploratory pattern of the Argentine ant. J. Insect Behav. 3, 159–168 10.1007/BF01417909 (doi:10.1007/BF01417909) [DOI] [Google Scholar]
- 43.Deneubourg J. L., Goss S. 1989. Collective patterns and decision-making. Ethol. Ecol. Evol. 1, 295–311 10.1080/08927014.1989.9525500 (doi:10.1080/08927014.1989.9525500) [DOI] [Google Scholar]
- 44.Beckers R., Deneubourg J. L., Goss S. 1993. Modulation of trail laying in the ant Lasius niger and its role in the collective selection of a food source. J. Insect Behav. 6, 751–759 10.1007/BF01201674 (doi:10.1007/BF01201674) [DOI] [Google Scholar]
- 45.Pratt S. C., Sumpter D. J. T. 2006. A tunable algorithm for collective decision-making. Proc. Natl Acad. Sci. USA 103, 15 906–15 910 10.1073/pnas.0604801103 (doi:10.1073/pnas.0604801103) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Couzin I. 2007. Collective minds. Nature 445, 715. 10.1038/445715a (doi:10.1038/445715a) [DOI] [PubMed] [Google Scholar]
- 47.Couzin I. D., Krause J. 2003. Self-organization and collective behavior in vertebrates. Adv. Study Behav. 32, 1–75 10.1016/S0065-3454(03)01001-5 (doi:10.1016/S0065-3454(03)01001-5) [DOI] [Google Scholar]
- 48.Parrish J. K., Viscido S. V., Grunbaum D. 2002. Self-organized fish schools: an examination of emergent properties. Biol. Bull. 202, 296–305 10.2307/1543482 (doi:10.2307/1543482) [DOI] [PubMed] [Google Scholar]
- 49.Vicsek T., Czirok A., Benjacob E., Cohen I., Shochet O. 1995. Novel type of phase-transition in a system of self-driven particles. Phys. Rev. Lett. 75, 1226–1229 10.1103/PhysRevLett.75.1226 (doi:10.1103/PhysRevLett.75.1226) [DOI] [PubMed] [Google Scholar]
- 50.Ballerini M., et al. 2008. Empirical investigation of starling flocks: a benchmark study in collective animal behaviour. Anim. Behav. 76, 201–215 10.1016/j.anbehav.2008.02.004 (doi:10.1016/j.anbehav.2008.02.004) [DOI] [Google Scholar]
- 51.Hemelrijk C. K., Hildenbrandt H. 2008. Self-organized shape and frontal density of fish schools. Ethology 114, 245–254 10.1111/j.1439-0310.2007.01459.x (doi:10.1111/j.1439-0310.2007.01459.x) [DOI] [Google Scholar]
- 52.Dyer J. R. G., Ioannou C. C., Morrell L. J., Croft D. P., Couzin I. D., Waters D. A., Krause J. 2008. Consensus decision making in human crowds. Anim. Behav. 75, 461–470 10.1016/j.anbehav.2007.05.010 (doi:10.1016/j.anbehav.2007.05.010) [DOI] [Google Scholar]
- 53.Austen-Smith D., Feddersen T. J. 2009. Information aggregation and communication in committees. Phil. Trans. R. Soc. B 364, 763–769 10.1098/rstb.2008.0256 (doi:10.1098/rstb.2008.0256) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Passino K. M., Seeley T. D. 2006. Modeling and analysis of nest-site selection by honeybee swarms: the speed and accuracy trade-off. Behav. Ecol. Sociobiol. 59, 427–442 10.1007/s00265-005-0067-y (doi:10.1007/s00265-005-0067-y) [DOI] [Google Scholar]
- 55.Ruckstuhl K. E. 1999. To synchronise or not to synchronise: a dilemma for young bighorn males? Behaviour 136, 805–818 10.1163/156853999501577 (doi:10.1163/156853999501577) [DOI] [Google Scholar]
- 56.King A. J., Cowlishaw G. 2009. All together now: behavioural synchrony in baboons. Anim. Behav. 78, 1381–1387 10.1016/j.anbehav.2009.09.009 (doi:10.1016/j.anbehav.2009.09.009) [DOI] [Google Scholar]
- 57.McClure M., Ralph M., Despland E. 2011. Group leadership depends on energetic state in a nomadic collective foraging caterpillar. Behav. Ecol Sociobiol. 65, 1573–1579 10.1007/s00265-011-1167-5 (doi:10.1007/s00265-011-1167-5) [DOI] [Google Scholar]
- 58.Sueur C., et al. 2011. Collective decision-making and fission-fusion dynamics: a conceptual framework. Oikos 120, 1608–1617 10.1111/j.1600-0706.2011.19685.x (doi:10.1111/j.1600-0706.2011.19685.x) [DOI] [Google Scholar]
- 59.Black J. M. 1988. Preflight signaling in swans—a mechanism for group cohesion and flock formation. Ethology 79, 143–157 10.1111/j.1439-0310.1988.tb00707.x (doi:10.1111/j.1439-0310.1988.tb00707.x) [DOI] [Google Scholar]
- 60.Stolba A. 1979. Entscheidungsfinden in Verbaenden von Papio hamadryas. Zurich, Switzerland: Universitaet Zuerich [Google Scholar]
- 61.Boinski S., Campbell A. F. 1995. Use of trill vocalizations to coordinate troop movement among white-faced Capuchins—a 2nd field-test. Behaviour 132, 875–901 10.1163/156853995X00054 (doi:10.1163/156853995X00054) [DOI] [Google Scholar]
- 62.Byrne R. W. 2000. How monkeys find their way: leadership, coordination and cognitive maps of African baboons. In On the move (eds Boinski S., Garber P. A.). Chicago, IL: University of Chicago Press [Google Scholar]
- 63.Bousquet C. A. H., Manser M. B. 2011. Resolution of experimentally induced symmetrical conflicts of interest in meerkats. Anim. Behav. 81, 1101–1107 10.1016/j.anbehav.2011.02.030 (doi:10.1016/j.anbehav.2011.02.030) [DOI] [Google Scholar]
- 64.Stewart K. J., Harcourt A. H. 1994. Gorillas vocalizations during rest periods—signals of impending departure. Behaviour 130, 29–40 10.1163/156853994X00127 (doi:10.1163/156853994X00127) [DOI] [Google Scholar]
- 65.Sellers W. I., Hill R. A., Logan B. S. 2007. An agent-based model of group decision making in baboons. Phil. Trans. R. Soc. B 362, 1699–1710 10.1098/rstb.2007.2064 (doi:10.1098/rstb.2007.2064) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Milton K. 2000. Quo vadis? Tactics of food search and group movement in primates and other animals. In On the move (eds Boinski S., Garber P. A.). Chicago, IL: University of Chicago Press [Google Scholar]
- 67.Meunier H., Leca J. B., Deneubourg J. L., Petit O. 2006. Group movement decisions in capuchin monkeys: the utility of an experimental study and a mathematical model to explore the relationship between individual and collective behaviours. Behaviour 143, 1511–1527 10.1163/156853906779366982 (doi:10.1163/156853906779366982) [DOI] [Google Scholar]
- 68.Ramseyer A., Boissy A., Dumont B., Thierry B. 2009. Decision making in group departures of sheep is a continuous process. Anim. Behav. 78, 71–78 10.1016/j.anbehav.2009.03.017 (doi:10.1016/j.anbehav.2009.03.017) [DOI] [Google Scholar]
- 69.Ramseyer A., Petit O., Thierry B. 2009. Decision-making in group departures of female domestic geese. Behaviour 146, 351–371 10.1163/156853909X410955 (doi:10.1163/156853909X410955) [DOI] [Google Scholar]
- 70.Sueur C., Deneubourg J. L., Petit O. 2011. From the first intention movement to the last joiner: macaques combine mimetic rules to optimize their collective decisions. Proc. R. Soc. B 278, 1697–1704 10.1098/rspb.2010.2084 (doi:10.1098/rspb.2010.2084) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Nagy M., Akos Z., Biro D., Vicsek T. 2010. Hierarchical group dynamics in pigeon flocks. Nature 464, 890–899 10.1038/nature08891 (doi:10.1038/nature08891) [DOI] [PubMed] [Google Scholar]
- 72.Lewis J. S., Wartzok D., Heithaus M. R. 2011. Highly dynamic fission-fusion species can exhibit leadership when traveling. Behav. Ecol. Sociobiol. 65, 1061–1069 10.1007/s00265-010-1113-y (doi:10.1007/s00265-010-1113-y) [DOI] [Google Scholar]
- 73.Krause J., Ruxton G. D. 2002. Living in groups. Oxford, UK: Oxford University Press [Google Scholar]
- 74.Krause J. 1993. The relationship between foraging and shoal position in a mixed shoal of roach Rutilus-Rutilus and chub Leuciscus-Cephalus—a field-study. Oecologia 93, 356–359 10.1007/BF00317878 (doi:10.1007/BF00317878) [DOI] [PubMed] [Google Scholar]
- 75.Fischhoff I. R., Sundaresan S. R., Cordingley J., Larkin H. M., Sellier M. J., Rubenstein D. I. 2007. Social relationships and reproductive state influence leadership roles in movements of plains zebra, Equus burchellii. Anim. Behav. 73, 825–831 10.1016/j.anbehav.2006.10.012 (doi:10.1016/j.anbehav.2006.10.012) [DOI] [Google Scholar]
- 76.Johnstone R. A., Manica A. 2011. Evolution of personality differences in leadership. Proc. Natl Acad. Sci. USA 108, 8373–8378 10.1073/pnas.1102191108 (doi:10.1073/pnas.1102191108) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Schulte E. 2010. Information aggregation and preference heterogeneity in committees. Theory Decis. 69, 97–118 10.1007/s11238-008-9118-y (doi:10.1007/s11238-008-9118-y) [DOI] [Google Scholar]
- 78.Conradt L., Roper T. J. 2007. Democracy in animals: the evolution of shared group decisions. Proc. R. Soc. B 274, 2317–2326 10.1098/rspb.2007.0186 (doi:10.1098/rspb.2007.0186) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Gompper M. E. 1996. Sociality and asociality in white-nosed coatis Nasua narica: foraging costs and benefits. Behav. Ecol. 7, 254–263 10.1093/beheco/7.3.254 (doi:10.1093/beheco/7.3.254) [DOI] [Google Scholar]
- 80.Conradt L., Roper T. J. 2009. Conflicts of interest and the evolution of decision sharing. Phil. Trans. R. Soc. B 364, 807–819 10.1098/rstb.2008.0257 (doi:10.1098/rstb.2008.0257) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Conradt L., Roper T. J. 2010. Deciding group movements: where and when to go. Behav. Process. 84, 675–677 10.1016/j.beproc.2010.03.005 (doi:10.1016/j.beproc.2010.03.005) [DOI] [PubMed] [Google Scholar]
- 82.Conradt L., Krause J., Couzin I. D., Roper T. J. 2009. ‘Leading according to need’ in self-organizing groups. Am. Nat. 173, 304–312 10.1086/596532 (doi:10.1086/596532) [DOI] [PubMed] [Google Scholar]
- 83.Bhattacharya K., Vicsek T. 2010. Collective decision making in cohesive flocks. New J. Phys. 12, 093019. 10.1088/1367-2630/12/9/093019 (doi:10.1088/1367-2630/12/9/093019) [DOI] [Google Scholar]
- 84.Holley R., Liggett T. M. 1975. Ergodic theorems for weakly interacting infinite systems and the voter model. Ann. Probab. 3, 643–663 10.1214/aop/1176996306 (doi:10.1214/aop/1176996306) [DOI] [Google Scholar]
- 85.Bode N. W. F., Wood A. J., Franks D. W. 2011. The impact of social networks on animal collective motion. Anim. Behav. 82, 29–38 10.1016/j.anbehav.2011.04.011 (doi:10.1016/j.anbehav.2011.04.011) [DOI] [Google Scholar]
- 86.King A. J., Sueur C. 2011. Where next? Group coordination and collective decision making by primates. Int. J.Primatol. 32, 1245–1267 10.1007/S10764-011-9526-7 (doi:10.1007/S10764-011-9526-7) [DOI] [Google Scholar]