

Abstract
Plasmodium falciparum malaria (Pf-malaria) and Epstein Barr Virus (EBV) infections coexist in children at risk for endemic Burkitt's lymphoma (eBL); yet studies have only glimpsed the cumulative effect of Pf-malaria on EBV-specific immunity. Using pooled EBV lytic and latent CD8+ T-cell epitope-peptides, IFN-γ ELISPOT responses were surveyed three times among children (10 months to 15 years) in Kenya from 2002–2004. Prevalence ratios (PR) and 95% confidence intervals (CI) were estimated in association with Pf-malaria exposure, defined at the district-level (Kisumu: holoendemic; Nandi: hypoendemic) and the individual-level. We observed a 46% decrease in positive EBV lytic antigen IFN-γ responses among 5–9 year olds residing in Kisumu compared to Nandi (PR: 0.54; 95% CI: 0.30–0.99). Individual-level analysis in Kisumu revealed further impairment of EBV lytic antigen responses among 5–9 year olds consistently infected with Pf-malaria compared to those never infected. There were no observed district- or individual-level differences between Pf-malaria exposure and EBV latent antigen IFN-γ response. The gradual decrease of EBV lytic antigen but not latent antigen IFN-γ responses after primary infection suggests a specific loss in immunological control over the lytic cycle in children residing in malaria holoendemic areas, further refining our understanding of eBL etiology.
Introduction
Plasmodium falciparum (Pf) malaria and Epstein Barr Virus (EBV) have been identified as co-factors in the pathogenesis of endemic Burkitt's lymphoma (eBL) [1] which is estimated to account for 70% of cancers among children in equatorial Africa [2], [3]. In areas with intense perennial malaria transmission (holoendemic), the highest incidence of eBL is in children aged 4–8 years [4], [5], [6], [7], [8], [9], in contrast to areas with low malaria transmission (hypoendemic) where eBL is rarely reported [7], [10], [11].
It has been hypothesized that Pf-malaria infections promote eBL in two mutually-compatible ways. In developing countries, most children experience primary EBV infection by 3 years of age, followed by life-long infection in memory B-lymphocytes [12], [13]. P. falciparum induces polyclonal B-cell expansion and lytic EBV reactivation [14], thus increasing the number of latently-infected B-cells. In otherwise healthy individuals, interferon-gamma (IFN-γ) secreting cytotoxic CD8+ T-cells mediate immunosurveillance of EBV [5], [12], [15], [16], [17], [18]. Repeated Pf-malaria infections could hence lead to exhaustion or hypo-responsiveness of EBV latent or lytic antigen CD8+ T-cells, thus increasing the chance for this EBV-associated malignancy to arise.
Limited evidence supports an impaired EBV-specific T-cell response in association with Pf-malaria. Using an in vitro regression assay as a measure of cytotoxicity, children with acute Pf-malaria demonstrated a transient loss of control over B-cell outgrowth [19], [20], [21]. Furthermore, case-control studies comparing acutely Pf-malaria infected individuals with healthy adults came to the same conclusion [22], [23]. However, the cumulative effect of repeated often asymptomatic Pf-malaria infections on EBV persistence has not been thoroughly studied [1], [6], [24], [25]. Two ecological studies provide the minimum understanding we have on the relationship. A study among adults found a loss of EBV-specific T-cell control among those exposed to holoendemic compared to hypoendemic malaria [26]. A second study only found significantly lower EBV latent and lytic antigen IFN-γ responses in children 5–9 years old residing in the holoendemic area compared to other age groups and children from a hypoendemic area [16].
The objective of this study was to examine the influence of cumulative Pf-malaria on EBV latent and lytic antigen CD8+ T-cell IFN-γ ELISPOT responses in children over a two-year period.
Results
Participant summary
Of the 236 children enrolled, 230 (97.5%) were seropositive for EBV [27]. Our weighted analysis included 149 children who participated in all surveys and had interpretable EBV-specific T-cell responses (Table 1). The age and sex distribution between the districts were not significantly different (P = .11 and P = .30, respectively). Children in Kisumu experienced more Pf-malaria infections than children in Nandi (P<.001); only 3% of Kisumu children were never infected compared to 78% in Nandi. This was despite a classically defined malaria outbreak in Nandi during the survey periods (Figure 1).
Table 1. Summary of participants in the Kisumu/Nandi cohort, Kenya 2002–2004a.
| Site | Total | ||||
| Kisumu (holoendemic) | Nandi (hypoendemic) | ||||
| n | % | n | % | ||
| Sex | |||||
| Male | 39 | 59.1 | 38 | 45.8 | 77 |
| Female | 27 | 40.9 | 45 | 54.2 | 72 |
| Age (in years) | |||||
| 0–4 | 16 | 24.2 | 30 | 36.1 | 46 |
| 5–9 | 33 | 50.0 | 35 | 42.2 | 68 |
| ≥10 | 17 | 25.8 | 18 | 21.7 | 35 |
| Malaria infections | |||||
| All surveys | 38 | 57.6 | 0 | 0 | 38 |
| Two surveys | 20 | 30.3 | 4 | 4.8 | 24 |
| One survey | 6 | 9.1 | 14 | 16.9 | 20 |
| Never | 2 | 3.0 | 65 | 78.3 | 67 |
| Total | 66 | 83 | 149 | ||
n, number; %, percentage.
Data in the table are weighted according to the 149 children who participated in all surveys and had interpretable Epstein-Barr virus (EBV) specific CD8+ T-cell IFN-γ response.
Figure 1. Malaria incidence in the highland area of Kipsamoite, 2001–2004.

The magnitude of EBV-specific IFN-γ responses did not differ significantly by malaria endemicity
The proportion of positive IFN-γ responses to PHA (positive control) demonstrates that children from both districts were equally able to elicit an IFN-γ response indicating no global signs of immune dysfunction (Table 2). There were no significant differences in median values of EBV lytic or latent CD8+ T-cell IFN-γ responses between children of similar age groups across districts. Therefore, Pf-malaria exposure does not appear to influence the magnitude of EBV-specific IFN-γ responses.
Table 2. EBV-specific CD8+ T-cell IFN-γ Response by Residence and Age Groupa.
| EBV lytic antigens | EBV latent antigens | PHAb | ||||||
| n | % | Medianc | n | % | Medianc | n | % | |
| (range) | (range) | |||||||
| Baseline (July–August 2002) | ||||||||
| Kisumu | ||||||||
| 0–4 years | 7/16 | 43.8 | 96 (14–166) | 8/16 | 50.0 | 43 (20–98) | 13/16 | 81.3 |
| 5–9 years | 8/33 | 24.2 | 67 (18–170) | 6/33 | 18.2 | 47 (16–448) | 31/33 | 93.9 |
| ≥10 years | 5/17 | 29.4 | 150 (20–350) | 5/17 | 29.4 | 46 (16–404) | 15/17 | 88.2 |
| Nandi | ||||||||
| 0–4 years | 12/30 | 34.3 | 98 (28–836) | 8/30 | 26.7 | 70 (18–146) | 28/30 | 93.3 |
| 5–9 years | 15/35 | 42.9 | 50 (22–792) | 11/35 | 31.4 | 84 (42–668) | 33/35 | 94.3 |
| ≥10 years | 8/18 | 22.9 | 53 (36–304) | 8/18 | 44.4 | 88 (26–1322) | 18/18 | 100 |
| First follow-up (February–March 2003) | ||||||||
| Kisumu | ||||||||
| 0–4 years | 4/16 | 25.0 | 46 (40–128) | 2/16 | 12.5 | 55 (32–78) | 16/16 | 100 |
| 5–9 years | 1/33 | 3.0 | 20 (20) | 6/33 | 18.2 | 15 (14–132) | 31/33 | 93.9 |
| ≥10 years | 5/17 | 29.4 | 30 (18–162) | 2/17 | 11.8 | 23 (18–28) | 16/17 | 94.1 |
| Nandi | ||||||||
| 0–4 years | 6/30 | 20.0 | 98 (24–744) | 4/30 | 13.3 | 77 (32–128) | 26/30 | 86.7 |
| 5–9 years | 8/35 | 22.9 | 82 (16–1742) | 8/35 | 22.9 | 58 (20–248) | 34/35 | 97.1 |
| ≥10 years | 5/18 | 27.8 | 54 (32–382) | 3/18 | 16.7 | 54 (14–354) | 18/18 | 100 |
| Second follow-up (July–August 2004) | ||||||||
| Kisumu | ||||||||
| 0–4 years | 5/16 | 31.3 | 76 (30–84) | 2/16 | 12.5 | 106 (64–148) | 15/16 | 93.8 |
| 5–9 years | 3/33 | 9.1 | 60 (56–150) | 3/33 | 9.1 | 42 (24–74) | 33/33 | 100 |
| ≥10 years | 3/17 | 17.7 | 250 (40–288) | 1/17 | 5.9 | 16 (16) | 17/17 | 100 |
| Nandi | ||||||||
| 0–4 years | 8/30 | 26.7 | 50 (14–384) | 2/30 | 6.7 | 69 (58–80) | 25/30 | 83.3 |
| 5–9 years | 7/35 | 20.0 | 76 (14–278) | 6/35 | 17.1 | 59 (14–214) | 31/35 | 88.6 |
| ≥10 years | 5/18 | 27.8 | 26 (14–130) | 5/18 | 27.8 | 56 (22–122) | 18/18 | 100 |
n, number; %, percentage; EBV, Epstein-Barr Virus; PHA, Phytohemagglutinin.
Data in the table are weighted according to the 149 children who participated in all surveys and had interpretable Epstein-Barr Virus (EBV) specific CD8+ T-cell IFN-γ response.
Phytohemagglutinin (PHA) was used as a positive control.
Median EBV-specific CD8+ T-cell IFN-γ responses were calculated among children with positive responses and is expressed as spot forming units (SFU) per 1×106 peripheral blood mononuclear cells (PBMC).
Pf-malaria exposure (district-level) and EBV-specific T-cell IFN-γ responses
EBV lytic antigen CD8+ T-cell IFN-γ responses
We observed a few intriguing patterns in the prevalence of positive EBV lytic antigen CD8+ T-cell IFN-γ response when children were stratified into age groups by their baseline age (age group cohorts) (Figure 2A,C). In Kisumu, the prevalence of positive responses in the 0–4 year and 5–9 year cohorts decreased from baseline to first follow-up, but remained unchanged in the ≥10 year cohort. By the second follow-up, responses increased among the 0–4 and 5–9 year cohorts while responses decreased in ≥10 year cohort. However, children in the 5–9 year cohort had the lowest prevalence at each survey period. In Nandi, responses declined in all age group cohorts from baseline to first follow-up and remained almost unchanged in the 5–9 year and ≥10 year cohorts by the second follow-up. In the 0–4 year cohort, however, responses increased. The patterns and prevalence of responses among the age group cohorts were similar at all survey periods, varying <10%.
Figure 2. Change in prevalence of EBV-specific CD8+ T-cell IFNγ responses with age.

Changes in prevalence of positive EBV lytic (A and C) and latent (B and D) antigen CD8+ T-cell IFNγ response from 2002–2004. Age group at each survey period is based on age at baseline. In Kisumu: 16 (0–4 years), 33 (5–9 years) and 17 (≥10 years). In Nandi: 30 (0–4 years), 35 (5–9 years) and 18 (≥10 years). Solid black line: 0–4 year olds; hash-mark green line: 5–9 year olds; and dotted blue line: ≥10 year old children.
Using the district-level definition of Pf-malaria and the weighted model, we estimated the prevalence of positive responses in Kisumu was 0.70 (95% CI: 0.45–1.08) times the prevalence in Nandi although this 30% difference was not significant. In Kisumu, there were no significant differences in positive responses in children 0–4 years (PR: 1.39, 95% CI: 0.60–3.20) and 5–9 years (PR: 0.74, 95% CI: 0.37–1.48) when compared to children ≥10 years (Figure 3A). Likewise in Nandi, the prevalence of positive responses in children 0–4 years (PR: 1.10, 95% CI: 0.60–2.02) and 5–9 years (PR: 1.04, 95% CI: 0.61–1.76) did not differ significantly from children ≥10 years. When similar age groups were compared between districts, we detected a significant difference in children 5–9 years where the prevalence of positive responses in Kisumu was 0.54 (95% CI: 0.30–0.99) that of children in Nandi (Figure 3A). No other differences by age group were found.
Figure 3. Prevalence of EBV-specific CD8+ T-cell IFNγ response by age group and residence.

Prevalence of positive EBV lytic (A) and latent (B) antigen CD8+ T-cell IFNγ response by age group and site of residence, Kenya 2002–2004. Age group was classified as a time-varying factor. For both graphs, the number of observations for children in each age group in Kisumu was: 33 (0–4 years), 87 (5–9 years) and 78 (≥10 years). The number of observations for children in each age group in Nandi was: 54 (0–4 years), 125 (5–9 years) and 70 (≥10 years). P values for differences between areas of residence by age group are indicated.
EBV latent antigen CD8+ T-cell IFN-γ responses
Examining the patterns in the prevalence of positive EBV latent antigen CD8+ T-cell IFN-γ response by age group cohorts, there was variation within and between districts (Figure 2B,D). In Kisumu, the prevalence at baseline was highest among the 0–4 year cohort but then decreased to nearly the same prevalence as the other age group cohorts. In Nandi, there was a decreasing trend from baseline to second follow-up for the 0–4 year and 5–9 year cohorts. However, the ≥10 years cohort had the highest prevalence of response at baseline that decreased by the first follow-up but rebounded by the second follow-up.
From our weighted model, we observed the prevalence of positive responses in Kisumu was 0.80 (95% CI: 0.51–1.25) times the prevalence in Nandi, although not significant. In Kisumu, the prevalence of positive responses in children 0–4 years (PR: 1.93, 95% CI: 0.91–4.13) and 5–9 years (PR: 1.22, 95% CI: 0.61–2.45) was not significantly different from children ≥10 years, although there was a decrease in prevalence with increasing age group (Figure 3B). Similarly in Nandi, responses among children 0–4 years (PR: 0.72, 95% CI: 0.35–1.48) and 5–9 years (PR: 0.84, 95% CI: 0.47–1.49) did not differ significantly from children ≥10 years old, although there was a slight increase in response with increasing age. Despite these interesting trends, there were no significant differences in the prevalence of positive responses when similar age groups were compared between districts.
Pf-malaria infection (individual-level) and EBV-specific T-cell IFN-γ responses
EBV lytic antigen CD8+ T-cell IFN-γ responses
Using the individual-level definition of Pf-malaria in our weighted model, we found the association between recurrent Pf-malaria infections and EBV lytic antigen CD8+ T-cell IFN-γ response varied by age group and survey period. We therefore used two weighted models. In the first model, we stratified results by age group, while adjusting for sex and survey period. Similarly in the second model, we stratified by survey period while adjusting for sex and age group.
We noted three observations from our analysis. First, the PR of recurrent Pf-malaria infections and positive IFN-γ responses among Kisumu children were consistently lower than Nandi children for all age groups and survey periods (Table 3). In general, there is a two-fold difference in the PR between Kisumu and Nandi although not significant (P = .32). Secondly, the association between recurrent Pf-malaria infections and IFN-γ responses varied by age group. In both Kisumu and Nandi, the prevalence of positive responses among children 0–4 years with recurrent Pf-malaria infections was higher than that of similarly aged children never infected. In Nandi, the difference was statistically significant. Finally, the PR of recurrent Pf-malaria infections and IFN-γ responses to EBV lytic antigens varied by survey period in both districts. At baseline, for both districts, the PR of positive responses among children with recurrent Pf-malaria infections was greater compared to children never infected; this result was statistically significant in Nandi, but not Kisumu. However, the PR decreased at subsequent study periods; the prevalence of positive responses among children with recurrent Pf-malaria infection diminished over time compared to children never infected. This could reflect functional diminishment of responsive EBV lytic antigen T-cells under continuous pressure from Pf-malaria.
Table 3. Prevalence Ratios for EBV Lytic Antigen CD8+ T-cell IFN-γ Response.
| Kisumu | Nandi | |||
| Constant Pf-malaria infection versus no infection | Constant Pf-malaria infection versus no infection | |||
| PR | 95% CI | PR | 95% CI | |
| Unadjusted | 0.64 | 0.23–1.77 | 1.43 | 0.73–2.81 |
| Age groupsa | ||||
| 0–4 years | 1.31 | 0.28–6.18 | 3.00 | 1.72–5.23 |
| 5–9 years | 0.53 | 0.15–1.88 | 1.16 | 0.39–3.45 |
| ≥10 years | 0.78 | 0.16–3.53 | 0.98 | 0.33–2.95 |
| Survey periodsb | ||||
| Baseline | 1.24 | 0.49–3.11 | 1.76 | 1.07–2.91 |
| Six months | 0.29 | 0.05–1.62 | 0.73 | 0.17–3.22 |
| Two years | 0.21 | 0.05–0.92 | 0.22 | 0.02–3.23 |
Pf-malaria, Plasmodium falciparum malaria; EBV, Epstein-Barr virus; PR, prevalence ratio; CI, confidence interval; Ref, referent group.
Adjusted for sex and survey period. Unstratified estimates for constant Pf-malaria infections compared to never infected in Kisumu (P = 0.72) and Nandi (P = 0.97) were not significant. Specific details on the number and prevalence of positive responses for each age group are included in Table 2.
Adjusted for sex and age group. Unstratified estimates for constant Pf-malaria infections compared to never infected was not significant in Kisumu (P = 0.65) but significant in Nandi (P = 0.03). The number of children in Kisumu for each survey period was 66 and the number of children in Nandi was 83.
EBV latent antigen CD8+ T-cell IFN-γ responses
Using our weighted model, we did not observe any variation by age group (Table 4) or survey period (data not shown). In Kisumu, for all age groups, the adjusted prevalence of positive EBV latent antigen CD8+ T-cell IFN-γ response was higher among children with recurrent Pf-malaria infections compared to those never infected (Table 4). There was a two-fold difference in the PR for children 0–4 years and >10 years with recurrent Pf-malaria infections than children 5–9 years. In Nandi, children 0–4 years with recurrent Pf-malaria infections had fewer positive responses than children never infected, and a PR that was three-fold lower than older children. However, children in older age groups with recurrent Pf-malaria infections had higher positive responses than similarly aged children never infected. Despite estimates for Kisumu and Nandi being imprecise and not statistically significant, the observations suggest that children 5–9 years in Kisumu are unable to mount the type of T-cell response as younger and older children. Meanwhile, in Nandi, the increasing PR with age may reflect how a maturing immune system, not continuously exposed to Pf-malaria, is able to induce a T-cell response to latent antigens even when co-infected with Pf-malaria.
Table 4. Prevalence Ratio for EBV Latent Antigen CD8+ T-cell IFN-γ Response.
| Kisumu | Nandi | |||
| Constant Pf-malaria infection versus no infection | Constant Pf-malaria infection versus no infection | |||
| PR | 95% CI | PR | 95% CI | |
| Unadjusted | 1.60 | 0.37–6.92 | 1.54 | 0.71–3.35 |
| Age groupsa | ||||
| 0–4 years | 2.10 | 0.22–19.65 | 0.51 | 0.08–3.37 |
| 5–9 years | 1.14 | 0.26–4.99 | 1.47 | 0.58–3.63 |
| ≥10 years | 2.68 | 0.38–18.73 | 1.82 | 0.83–3.99 |
Pf-malaria, Plasmodium falciparum malaria; EBV, Epstein - Barr virus; PR, prevalence ratio; CI, confidence interval.
Adjusted for sex and survey period. Unstratified estimates for constant Pf-malaria infections compared to never infected in Kisumu (P = 0.32) and Nandi (P = 0.13) were not significant. Specific details on the number and prevalence of positive responses for each age group are included in Table 2.
Discussion
Our study demonstrates that the prevalence of positive EBV lytic- but not latent-antigen CD8+ T-cell IFN-γ responses decreases in a malaria holoendemic area and not a hypoendemic area. This suggests that children repeatedly infected with Pf-malaria eventually lose functional IFN-γ producing CD8+ T-cells in response to EBV lytic antigens. In an effort to control viral replication induced by recurrent Pf-malaria infections [14], we hypothesize that EBV lytic antigen CD8+ T-cells have become exhausted and unable to produce IFN-γ or alternatively these cells were culled through apoptosis. As a result of the loss of responsive EBV lytic antigen CD8+ T-cells, more B-lymphocytes could become latently infected by EBV, and thus gradually increasing the risk of eBL. These findings are consistent with previous studies of this cohort, which detected significantly higher median EBV viral load and EBV-specific IgG antibodies to EBV lytic and latent antigens in the holoendemic compared to hypoendemic area [27],[28].
Furthermore, the association between Pf-malaria infections and positive EBV lytic antigen CD8+ T-cell IFN-γ responses varied by age group. The EBV lytic antigen deficiency was most pronounced among children 5–9 years old in the malaria holoendemic area and was further potentiated in those recurrently infected with Pf-malaria. In our individual-level analysis, these children had the lowest PR of positive responses while this same age group in the hypoendemic area appeared to be affected little. Additionally, the patterns observed in the age group cohorts clearly showed that the 5–9 year cohort in Kisumu had the lowest prevalence of positive responses among all age group cohorts, in both districts, at each survey period. The sustained inability to produce an effective EBV lytic antigen CD8+ T-cell IFN-γ response among 5–9 year olds may be an etiologically relevant event in eBL development since eBL is most often diagnosed in this age group. Finally, the inconsistency of patterns between age group cohorts within a district suggests there is an age-dependent interaction between Pf-malaria and EBV-specific T-cell response. Studies of immune mechanisms that induce exhaustion or deletion are needed to understand maintenance of EBV-specific T-cell immunity, especially in children. In support of this premise is the observation that the ELISPOT responses in both groups of children were lower than those described for healthy adults [29] and our Kenyan adult controls (data not shown). To our knowledge there have been no studies of EBV-specific T-cell immunity in healthy children from non-malaria endemic countries. However our studies of malaria-specific immunity also demonstrate an age-associated instability in cytokine recall responses more pronounced in children compared to adults [30], [31].
This study is an important early step to understanding the cumulative effect of Pf-malaria infections on EBV-specific T-cell immunity over time. Availability of data over two-years permitted identification of potentially important biological and environmental mechanisms that only became apparent over time. For example, the association between Pf-malaria infection and positive EBV lytic antigen CD8+ T-cell IFN-γ responses varied by age group and survey period. The variation noted with age group is expected because there is an age-dependent increase in T-cell immunity as children develop protection against Pf-malaria after repeated infections [32]. Children in malaria holoendemic areas acquire immunity to Pf-malaria and EBV during the first years of life, and ongoing studies will compare the development of Pf-malaria to EBV-specific T-cell memory. An impairment of EBV-specific T-cell control with progressive EBV reactivation has been described in HIV-infected individuals [33], lending support to a sequential series of events in the etiology of eBL.
Using data, collected during a two-year period, also allowed us to use an individual-level definition for Pf-malaria infections. Unlike other studies, our definition accounted for the cumulative effect of Pf-malaria infection which has been hypothesized to be critical in the pathogenesis of eBL, rather than the transient effect typically observed with acute Pf-malaria infection [1]. However, our definition was vulnerable to misclassification because Pf-malaria infection was assessed only twice during the two-year follow-up. Therefore, we may not have captured participants' malaria histories accurately. This misclassification was likely to be differential because children in the holoendemic area were exposed to Pf-malaria parasites at a higher frequency, averaging two malaria infections per year, than children in the hypoendemic area [34]. Therefore, we may have underestimated or overestimated the PR for Pf-malaria infections and EBV-specific T-cell responses in the holoendemic area.
A strength of our study was the use of two definitions for Pf-malaria: 1) district-level according to malaria transmission intensity, and 2) individual-level based on measured Pf-malaria infection. Although our findings of EBV lytic antigen CD8+ T-cell IFN-γ responses were consistent with both definitions, our findings of EBV latent antigen CD8+ T-cell IFN-γ responses were inconsistent. This may have been due to the limited power or an underestimation of the influence of Pf-malaria infections in hypoendemic areas. However, it also highlights the potential pitfall in attributing district-level results to the individual, also known as the ecological fallacy. The inconsistency may have been due to other factors that differed between the districts and unrelated to malaria transmission intensities. Therefore, we conclude that the use of malaria transmission intensity as a surrogate for malaria infection has been informative yet future studies should endeavor to prospectively collect Pf-malaria and EBV co-infections information from individuals to more accurately describe this complex relationship.
There were several potential confounders that were not captured in our study, specifically HIV status, nutritional status, schistosomiasis infection, and socioeconomic status. However, we do not believe the absence of these confounders materially affected our findings. When data were collected in western Kenya from 2002–2004, HIV testing in infants was conducted only when medically warranted. All children enrolled in this study were examined by a clinician and had no obvious signs of illness or malnourishment, and no deaths were reported as of 2009. Therefore, even though it was possible that an underlying HIV infection might influence the rate of malaria parasitaemia [35] the likelihood that a significant number of undiagnosed children remained in this study was low. Schistosomiasis infection was unmeasured yet an examination of the Pf-malaria and EBV response relationship indicated adjusting for schistosomiasis infection would have biased our analysis. Finally, is socioeconomic status had been measures, participants and their families would likely have been classified as low socioeconomic status because the main occupation was fishing (Kisumu) and farming (Nandi) in both rural study areas with homes constructed of locally available materials.
Our findings on EBV lytic antigen CD8+ T-cell IFN-γ responses were consistent with the studies that have used residence area (malaria transmission intensity) to explore the cumulative effect of Pf-malaria infections on EBV-specific T-cell response. We observed fewer positive EBV lytic antigen CD8+ T-cell IFN-γ responses among 5–9 year old than older children [16]. We also identified a reduction in EBV-specific T-cell response among children living in a holoendemic compared a hypoendemic area [26]. The consistency of our findings with previous studies is important given our limited sample size and precision. Meanwhile, our analysis of EBV lytic antigen CD8+ T-cell IFN-γ response at the individual-level supports findings from previous studies that used residence area as a surrogate for Pf-malaria infection.
However, we did not detect the same statistically significant district-level difference in positive EBV latent antigen CD8+ T-cell IFN-γ responses among 5–9 year olds as a previous study [16]. This discrepancy may be due to the limited power of our study. Furthermore, the difference between our individual-level analysis and the previous study may also be due to the use of a surrogate definition of Pf-malaria.
This study design marks a step toward examining the individual-level association of Pf-malaria infections and EBV-specific T-cell IFN-γ responses and identifies a potential difference between children recurrently infected with Pf-malaria compared to children never infected. To adequately quantify this effect, a longitudinal study should be considered which could accurately measure Pf-malaria infection and changes in Pf-malaria and EBV-specific T-cell immunity over time. The temporal aspects of future studies will be vital to elucidating the precise mechanism by which repeated Pf-malaria infections affect EBV persistence and immunity.
Materials and Methods
The Kisumu/Nandi cohort has been previously described [27]. In brief, the cohort consists of 236 children, randomly selected and between 10 months and 15 years at enrollment, from two districts in western Kenya with disparate Pf-malaria transmission intensities: Kisumu is characterized as holoendemic and Nandi as hypoendemic. Due to the age-related incidence of eBL, an equal distribution of children by age and sex were enrolled from each area: children 0–4 years have an elevated risk of eBL whereas 5–9 year olds are at highest risk and ≥10 years old have the lowest risk. Data were collected from 2002–2004 using a standardized survey. Three face-to-face interviews were conducted at baseline (July–August 2002), six month follow-up (February–March 2003), and two-year follow-up (July–August 2004). Blood was also collected for malaria and EBV testing.
Pf-malaria infection was confirmed on thick and thin blood smears by microscopy. Testing of EBV-specific T-cell response by IFN-γ ELISPOT has been previously described [16]. Lytic (BRLF1, BZLF1, and BMLF1) and latent (Epstein-Barr nuclear antigen [EBNA] 3A, EBNA 3B, and EBNA3C) antigens were selected and pooled for testing. One positive control (mitogen phytohemmagglutinin [PHA]) was used to stimulate wells and a negative control (phosphate buffer saline [PBS]) was used to measure background IFN-γ response in unstimulated wells. Assays were condensed into a three-week period using the same reagents and personnel to minimize inter-assay variability. Cytotoxic T-lymphocyte (CTL) ImmunoSpot scanning and imaging software (version 4; Cellular Technology Ltd, Shaker Heights, OH) was used to count the number of spot-forming units (SFU) per well; results were expressed as SFU per million peripheral blood mononuclear cells (PBMC). Using a two-sided Fisher's exact test (P<.05), EBV lytic and latent epitope-peptide CD8+ T-cell IFN-γ responses were categorized as positive or negative. A positive response was recorded if the proportion of SFUs in the stimulated well was significantly different from the proportion of SFU in the unstimulated well. The magnitude of response was calculated by subtracting the SFU in PBS wells (negative control) from the SFU in the stimulated wells. The median value for the negative control wells was 4 SFU per million PBMCs (range 0 to 772 SFU/million PBMC). Median values were calculated among positive responders only.
Analyses were restricted to EBV seropositive children at baseline [27]. We used two definitions of cumulative Pf-malaria. First, Pf-malaria exposure was defined according to the malaria transmission intensity of the district (district-level definition): Kisumu (holoendemic) or Nandi (hypoendemic). Next, Pf-malaria infection was defined as the cumulative average of P. falciparum infection (parasitemia) in a participant over the three survey periods (individual-level definition). The value ranged from 0 (never infected) to 1 (always infected); results and discussion focus on children who were always infected (referred to as recurrent) and never infected. With the individual-level definition, we also included the covariates age group, district, sex, and when the survey was conducted (referred to as survey period) in the analysis. We first examined covariates as potential effect measure modifiers using an a priori cutoff of P = .20. In the absence of evidence of effect measure modification, we included covariates in the model as potential confounders.
For descriptive analyses, we used the Chi-square statistic to measure associations between categorical exposures and outcomes. We used the two-sided Wilcoxon rank sum (Mann-Whitney U)/Kruskal Wallis test for continuous outcomes. For multivariable analyses, we used weighted log-binomial regression with robust variances to estimate the prevalence ratios (PR) and corresponding 95% confidence intervals (CI). We used generalized estimating equations (GEE) with robust variance estimators to account for correlation due to repeated measurements. A weighted model with inverse probability weights (IPWs) was used to address missing observations due to children not participating in all surveys. When using IPW, observations are assumed to be missing at random; missing data are dependent on observed data but independent of unobserved data [36]. To calculate IPWs, the probability of participation was modeled using available predictor variables. The inverse of the predicted probabilities were calculated and assigned to each individual with complete data. Individuals with complete data were weighted to represent individuals with similar characteristics who have missing data.
To calculate the predicted probabilities for our study, we used logistic regression with first order interaction terms using the following equation:
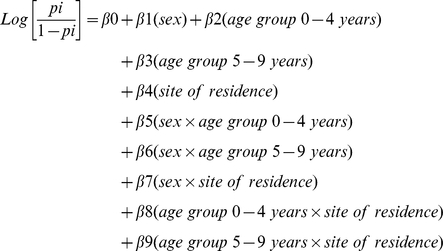 |
Where pi was the probability of child i (i = 1,2,3…n) participating in all surveys. Taking the inverse of the predicted probabilities, the mean IPW was 1.44 and ranged from 1.14–1.92. Children who participated in all three surveys were assigned the mean IPW value whereas children with missing observations were assigned an IPW of 0. We also conducted complete case analyses and found no differences in the PR or 95% CI; therefore we report results from the weighted analyses. Data were analyzed in SAS 9.1.3 (Cary, NC).
Written informed consent was obtained from a parent or guardian of the participant. This study was approved by the Institutional Review Boards at the University Hospitals of Cleveland, Case Western Reserve University where Dr. Moormann was affiliated at the time this study was done and also obtained from the Ethical Review Committee for the Kenya Medical Research Institute. It was deemed exempt by the Institutional Review Board at the University of North Carolina at Chapel Hill.
Acknowledgments
We are grateful to the Kenya Medical Research Institute staff for organizing blood sample collection and processing. We thank Nancy Raab-Trab for reviewing early versions of the manuscript. Special thanks to John Oyombe, John Ogone and Fred Opinya for recruiting and retaining study participants. We are also grateful to the parents/guardians of the study participants and the children that participated in the study. These data are published with the approval of the Director of the Kenya Medical Research Institute.
Footnotes
Competing Interests: The authors have declared that no competing interests exist.
Funding: This work was supported by the Infectious Disease Epidemiology Training Program from the National Institutes of Health [5-T32-AI070114-03 to CJS]. The original study was supported by the National Institutes of Health [K08 AI51565, CA134051, AI01572, and AI056270]. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Klein G. Epstein-Barr virus, malaria and Burkitt's lymphoma. Scand J Infect Dis. 1982;Suppl 36:15–23. [PubMed] [Google Scholar]
- 2.Donati D, Espmark E, Kironde F, Mbidde EK, Kamya M, et al. Clearance of circulating Epstein-Barr virus DNA in children with acute malaria after antimalaria treatment. J Infect Dis. 2006;193:971–977. doi: 10.1086/500839. [DOI] [PubMed] [Google Scholar]
- 3.Roberts DE, Quincey E. Epstein-Barr virus serology and malaria exposure in a small group of Liberian children with splenomegaly. J Trop Pediatr. 1985;31:209–212. doi: 10.1093/tropej/31.4.209. [DOI] [PubMed] [Google Scholar]
- 4.Brady G, Macarthur GJ, Farrell PJ. Epstein-Barr virus and Burkitt lymphoma. Postgrad Med J. 2008;84:372–377. doi: 10.1136/jcp.2007.047977. [DOI] [PubMed] [Google Scholar]
- 5.Facer CA, Playfair JH. Malaria, Epstein-Barr virus, and the genesis of lymphomas. Adv Cancer Res. 1989;53:33–72. doi: 10.1016/s0065-230x(08)60278-x. [DOI] [PubMed] [Google Scholar]
- 6.Kafuko GW, Burkitt DP. Burkitt's lymphoma and malaria. Int J Cancer. 1970;6:1–9. doi: 10.1002/ijc.2910060102. [DOI] [PubMed] [Google Scholar]
- 7.Burkitt D, Wright D. Geographical and tribal distribution of the African lymphoma in Uganda. Br Med J. 1966;1:569–573. doi: 10.1136/bmj.1.5487.569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.De-The G. The epidemiology of Burkitt's lymphoma: evidence for a causal association with Epstein-Barr virus. Epidemiol Rev. 1979;1:32–54. doi: 10.1093/oxfordjournals.epirev.a036213. [DOI] [PubMed] [Google Scholar]
- 9.Rasti N, Falk KI, Donati D, Gyan BA, Goka BQ, et al. Circulating epstein-barr virus in children living in malaria-endemic areas. Scand J Immunol. 2005;61:461–465. doi: 10.1111/j.1365-3083.2005.01589.x. [DOI] [PubMed] [Google Scholar]
- 10.Burkitt DP. Etiology of Burkitt's lymphoma–an alternative hypothesis to a vectored virus. J Natl Cancer Inst. 1969;42:19–28. [PubMed] [Google Scholar]
- 11.Kafuko GW, Baingana N, Knight EM, Tibemanya J. Association of Burkitt's tumour and holoendemic malaria in West Nile District, Uganda: malaria as a possible aetiologic factor. East Afr Med J. 1969;46:414–436. [PubMed] [Google Scholar]
- 12.Rochford R, Cannon MJ, Moormann AM. Endemic Burkitt's lymphoma: a polymicrobial disease? Nat Rev Microbiol. 2005;3:182–187. doi: 10.1038/nrmicro1089. [DOI] [PubMed] [Google Scholar]
- 13.Rickinson AB, Kieff E. Epstein-Barr Virus. In: Knipe DM, Howley PM, editors. Fields Virology. Fifth ed. Philadelphia, PA: Lippincott Williams & Wilkins; 2007. [Google Scholar]
- 14.Chene A, Donati D, Guerreiro-Cacais AO, Levitsky V, Chen Q, et al. A molecular link between malaria and Epstein-Barr virus reactivation. PLoS Pathog. 2007;3:e80. doi: 10.1371/journal.ppat.0030080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Moss DJ, Rickinson AB, Pope JH. Long-term T-cell-mediated immunity to Epstein-Barr virus in man. I. Complete regression of virus-induced transformation in cultures of seropositive donor leukocytes. Int J Cancer. 1978;22:662–668. doi: 10.1002/ijc.2910220604. [DOI] [PubMed] [Google Scholar]
- 16.Moormann AM, Chelimo K, Sumba PO, Tisch DJ, Rochford R, et al. Exposure to holoendemic malaria results in suppression of Epstein-Barr virus-specific T cell immunosurveillance in Kenyan children. J Infect Dis. 2007;195:799–808. doi: 10.1086/511984. [DOI] [PubMed] [Google Scholar]
- 17.Young LS, Rickinson AB. Epstein-Barr virus: 40 years on. Nat Rev Cancer. 2004;4:757–768. doi: 10.1038/nrc1452. [DOI] [PubMed] [Google Scholar]
- 18.Munz C. Immune Response and Evasion in the Host-EBV Interaction. In: Robertson ES, editor. Epstein-Barr Virus. Norfolk, England: Cister Academic Press; 2005. pp. 197–231. [Google Scholar]
- 19.Lam KM, Syed N, Whittle H, Crawford DH. Circulating Epstein-Barr virus-carrying B cells in acute malaria. Lancet. 1991;337:876–878. doi: 10.1016/0140-6736(91)90203-2. [DOI] [PubMed] [Google Scholar]
- 20.Whittle HC, Brown J, Marsh K, Blackman M, Jobe O, et al. The effects of Plasmodium falciparum malaria on immune control of B lymphocytes in Gambian children. Clin Exp Immunol. 1990;80:213–218. doi: 10.1111/j.1365-2249.1990.tb05236.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Whittle HC, Brown J, Marsh K, Greenwood BM, Seidelin P, et al. T-cell control of Epstein-Barr virus-infected B cells is lost during P. falciparum malaria. Nature. 1984;312:449–450. doi: 10.1038/312449a0. [DOI] [PubMed] [Google Scholar]
- 22.Gunapala DE, Facer CA, Davidson R, Weir WR. In vitro analysis of Epstein-Barr virus: host balance in patients with acute Plasmodium falciparum malaria. I. Defective T-cell control. Parasitol Res. 1990;76:531–535. doi: 10.1007/BF00931060. [DOI] [PubMed] [Google Scholar]
- 23.Njie R, Bell AI, Jia H, Croom-Carter D, Chaganti S, et al. The effects of acute malaria on Epstein-Barr virus (EBV) load and EBV-specific T cell immunity in Gambian children. J Infect Dis. 2009;199:31–38. doi: 10.1086/594373. [DOI] [PubMed] [Google Scholar]
- 24.Morrow RH, Gutensohn N, Smith PG. Epstein-Barr virus-malaria interaction models for Burkitt's lymphoma: implications for preventive trials. Cancer Res. 1976;36:667–669. [PubMed] [Google Scholar]
- 25.Morrow RH, Kisuule A, Pike MC, Smith PG. Burkitt's lymphoma in the Mengo Districts of Uganda: epidemiologic features and their relationship to malaria. J Natl Cancer Inst. 1976;56:479–483. doi: 10.1093/jnci/56.3.479. [DOI] [PubMed] [Google Scholar]
- 26.Moss DJ, Burrows SR, Castelino DJ, Kane RG, Pope JH, et al. A comparison of Epstein-Barr virus-specific T-cell immunity in malaria-endemic and -nonendemic regions of Papua New Guinea. Int J Cancer. 1983;31:727–732. doi: 10.1002/ijc.2910310609. [DOI] [PubMed] [Google Scholar]
- 27.Moormann AM, Chelimo K, Sumba OP, Lutzke ML, Ploutz-Snyder R, et al. Exposure to holoendemic malaria results in elevated Epstein-Barr virus loads in children. J Infect Dis. 2005;191:1233–1238. doi: 10.1086/428910. [DOI] [PubMed] [Google Scholar]
- 28.Piriou E, Kimmel R, Chelimo K, Middeldorp JM, Odada PS, et al. Serological evidence for long-term Epstein-Barr virus reactivation in children living in a holoendemic malaria region of Kenya. J Med Virol. 2009;81:1088–1093. doi: 10.1002/jmv.21485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Hislop AD, Annels NE, Gudgeon NH, Leese AM, Rickinson AB. Epitope-specific evolution of human CD8(+) T cell responses from primary to persistent phases of Epstein-Barr virus infection. J Exp Med. 2002;195:893–905. doi: 10.1084/jem.20011692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Moormann AM, John CC, Sumba PO, Tisch D, Embury P, et al. Stability of interferon-gamma and interleukin-10 responses to Plasmodium falciparum liver stage antigen-1 and thrombospondin-related adhesive protein in residents of a malaria holoendemic area. Am J Trop Med Hyg. 2006;74:585–590. [PubMed] [Google Scholar]
- 31.Moormann AM, Sumba PO, Tisch DJ, Embury P, King CH, et al. Stability of interferon-gamma and interleukin-10 responses to Plasmodium falciparum liver stage antigen 1 and thrombospondin-related adhesive protein immunodominant epitopes in a highland population from Western Kenya. Am J Trop Med Hyg. 2009;81:489–495. [PMC free article] [PubMed] [Google Scholar]
- 32.Moormann AM. How might infant and paediatric immune responses influence malaria vaccine efficacy? Parasite Immunol. 2009;31:547–559. doi: 10.1111/j.1365-3024.2009.01137.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Legoff J, Amiel C, Calisonni O, Fromentin D, Rajoely B, et al. Early impairment of CD8+ T cells immune response against Epstein-Barr virus (EBV) antigens associated with high level of circulating mononuclear EBV DNA load in HIV infection. J Clin Immunol. 2004;24:125–134. doi: 10.1023/B:JOCI.0000019777.75784.6f. [DOI] [PubMed] [Google Scholar]
- 34.Bloland PB, Boriga DA, Ruebush TK, McCormick JB, Roberts JM, et al. Longitudinal cohort study of the epidemiology of malaria infections in an area of intense malaria transmission II. Descriptive epidemiology of malaria infection and disease among children. Am J Trop Med Hyg. 1999;60:641–648. doi: 10.4269/ajtmh.1999.60.641. [DOI] [PubMed] [Google Scholar]
- 35.Abu-Raddad LJ, Patnaik P, Kublin JG. Dual infection with HIV and malaria fuels the spread of both diseases in sub-Saharan Africa. Science. 2006;314:1603–1606. doi: 10.1126/science.1132338. [DOI] [PubMed] [Google Scholar]
- 36.Singer JD, Willett JB. Applied Longitudinal Data Analysis: Modeling Change and Event Occurrence. New York: Oxford University Press; 2003. [Google Scholar]