

Abstract
Saponins are detergent-like substances showing antibacterial as well as anticancer potential. In this study, the effects of saponins from Quillaja saponaria were analyzed against prokaryotic and eukaryotic cells. Multidrug-resistant clinical E. coli strains were isolated from human urine. As eukaryotic cells, the CHO-K1 cell lines were applied. Antibacterial effect of ampicillin, streptomycin, and ciprofloxacin in the presence of saponins was measured by cultivation methods. Properties of saponins against CHO-K1 cells were measured by the MTT test, hemolysis assay and flow cytometry. Saponin from Quillaja saponaria has a cytotoxic effect at concentrations higher than 25 μg/mL and in the range of 12–50 μg/mL significantly increases the level of early apoptotic cells. Saponin at dose of 12 μg/mL enhances the six E. coli strains growth. We postulate that saponins increase the influx of nutrients from the medium into E. coli cells. Saponins do not have synergetic effects on antibacterial action of tested antibiotics. In contrary, in the presence of saponins and antibiotics, more CFU/mL E. coli cells were observed. This effect was similar to saponins action alone towards E. coli cells. In conclusion, saponins was cytotoxic against CHO-K1 cells, whereas against E. coli cells this effect was not observed.
1. Introduction
Saponins contain a steroidal or a triterpenoid aglycone to which one or more sugar chains are attached [1]. These chemical structures determine their biological properties as natural nonionic detergents which have cytotoxic, hemolytic, molluscicidal, anti-inflammatory, antifungal, antiyeast, antibacterial, and antiviral activities [2–4]. Saponins are nonionic detergents belonging to the group of glycosides. They are used in the pharmaceutical industry because some forms are the starting point for the semisynthesis of steroidal drugs. Many have pharmacological properties and are used in phytotherapy and in the cosmetic industry. Saponins can be classified into two groups based on the nature of their aglycone skeleton: steroidal (consisting of a C27 spirostane skeleton comprising a six-ring structure) and triterpenoid (consisting of a C30 skeleton comprising a pentacyclic structure) [1]. The biological properties of saponins depend on the structure of the aglycone and/or the number of sugar units involved [5]. The therapeutic potential of saponins against eukaryotic cells is associated with their cell membrane-permeabilizing properties by complexing with cholesterol [6]. Our previous studies with a series of synthetic saponins showed their potential as anticancer drugs [7]. Saponins, having surface-active properties, might insert into the lipid bilayer, bind to cholesterol, form domains enriched with cholesterol-saponin complexes, and finally lyse cells [8]. On the other hand, the absence of cholesterol in the membrane structure does not inhibit pore formation by saponins [9, 10]. In our previous study, we suggested that saponin might disturb the permeability of the bacterial outer membrane. About 90% of the surface of naturally cholesterol-free Gram-negative bacteria cell-wall outer membranes are covered by lipopolysaccharide (LPS). We concluded that saponin might interact with the lipid A part of Proteus LPSs and thereby increase the permeability of bacterial cell wall. Lipid A-saponin complexes might promote antibiotic (colistin, ampicillin) uptake to inherently resistant bacteria cells [11].
The potentially antibacterial effect of saponins from clinical point of view is associated with a comprehensive investigation of its antibacterial effect and minimal cytotoxic properties in eukaryotic cells. We previously showed that colistin or ampicillin in the presence of 15 μg/mL saponins reduced the numbers of cells of the laboratory strains P. mirabilis S1959 and R45 [11]. An important medical problem is the elimination of pathogenic Gram-positive and Gram-negative bacterial strains. Several types of actions are published, such as bacteriophage treatment [12], inhibition of bacterial adhesion [13], and increasing the permeability of bacterial cell walls [14].
The aims of the present study are
testing the saponin abilities to induce hemolytic and cytotoxic effects against eukaryotic cells,
studies of saponin influences on growth of clinical E. coli strains,
testing of saponin abilities to interfere with antibiotic action against E. coli cells.
2. Material and Methods
2.1. Chemicals
Saponin from Quillaja saponaria bark was obtained from Sigma Chemical Co., St. Louis, MO, USA. The main aglycone (sapogenin, 20–35%) moiety is quillaic acid, a triterpene of predominantly 30 carbon atoms of the ∆12-oleanane type. The aglycone is bound to various sugars, including glucose, glucuronic acid, galactose, xylose, apiose, rhamnose, fucose, and arabinose.
2.2. Cell Culture
CHO-K1 cells were cultured at 37°C in a humidified 5% CO2 atmosphere in plastic dishes in McCoy's 5A medium supplemented with 10% heat-inactivated fetal calf serum, 2 mM L-glutamine, and antibiotics (100 units/mL penicillin and 100 μg/mL streptomycin).
2.3. MTT Test
Saponin at final concentrations of 12 to 100 μg/mL was added to 500 μl of the CHO-K1 cell suspension (1.5 × 106/mL of complete McCoy's 5A medium) onto plates. CHO-K1 cells were treated with saponin for 72 hours at 37°C in 5% CO2. After treatment, the viability of cells was evaluated by the MTT assay. MTT reagent was added to each plate, and after 6 h of incubation in Hera Cell, 1 mL of SDS (10% in 0.01 N HCl) was added to dissolve the water-insoluble formazan salt. One hour later, the OD650 nm-OD570 nm difference was measured. Unexposed cells were regarded as 100% viable [15].
2.4. Apoptosis
CHO-K1 cells were treated with saponin at different concentrations (6, 12, 24, and 50 μg/mL) for 72 hours. After treatment, the frequencies of early apoptotic, late apoptotic, and necrotic cells were evaluated with the Annexin V-FITC apoptosis detection Kit I (BD Pharmingen, USA) described elsewhere [16]. Briefly, the cells were washed two times with cold PBS and then resuspended in a 1 × binding buffer at a concentration of 1 × 106 cells/mL. An aliquot of 125 μL of the cell suspension was incubated with 5 μL of Annexin V-FITC and 5 μL of propidium iodide (PI) at room temperature for 15 min in the dark. The cells were resuspended in 400 μL of 1 × binding buffer. The fluorescence was determined using a Becton Dickinson FACSscan flow cytometer. A computer system (CellQuest Pro, Becton Dickinson) was used for data acquisition and analysis. Data for 20000 events were stored. A cell gate containing CHO-K1 was established on the basis of forward and side light scatter. Four different populations of cells were detected with the Annexin V-FITC kit: normal cells that are Annexin negative and PI negative and express no fluorescence, early apoptotic cells that are Annexin positive and PI negative and express green fluorescence, late apoptotic/necrotic cells that are Annexin positive and PI positive and express green and orange fluorescence, and necrotic cells that are Annexin negative and PI positive and express orange fluorescence.
2.5. Hemolysis Assay
Blood was obtained from healthy young male donors. The red blood cells (RBCs) were washed three times and resuspended in sterile PBS to give about 15 × 106 cells per mL and further processed [17]. The erythrocytes were incubated with saponin in a range of 0.5–50 μg/mL for 1 h at 37°C. After centrifugation of the nonhemolysed erythrocytes (1400 rpm, 15 min), the absorbance of the released hemoglobin in the wavelength range of 400–600 nm was measured. The percentage of hemolysis was determined by comparing the absorbance of hemoglobin at 435, 540, and 575 nm released from the RBCs in the presence of saponin. The positive control (100% hemolysis) was determined by the amount of hemoglobin released from 15 × 106 RBCs after 10 min of incubation with water.
2.6. Bacteria Growth in the Presence of Saponin and Antibiotics
Six strains of E. coli isolated from patients at the Department of Microbiology, Holy Cross Cancer Center in Kielce, Poland, were included in this study. The stock solution of saponin (Sigma Chemical Co., St. Louis, MO, USA) was diluted in LB medium or minimal M9 medium described elsewhere [18]. The reaction mixtures contained saponin at concentrations of 0.1–12 μg/mL and a bacterial suspension of E. coli strains at 102 cells/mL in each probe in final volumes of 300 μl of proper medium. Incubation was carried out for 18 h at 37°C. Bacterial viability was expressed in terms of colony-forming units (CFU/mL). Additionally, the antimicrobial susceptibility of the clinical E. coli strains to ampicillin, streptomycin, and ciprofloxacin at 25–400 μg/mL was prospectively tested in the presence of saponin at 12 μg/mL in LB medium by the cultivation methods.
2.7. Data Analysis
The data were analyzed using the Statistica software package (StatSoft, Tulsa, OK, USA). All the values in this study are expressed as the mean ± SD of three experiments. If no significant differences between variations were found as assessed by the Snedecor-Fisher test, the differences were compared by the ANOVA test.
3. Results
3.1. The Effect of Saponin on Eukaryotic Cells
3.1.1. Drug Resistance
Figure 1 shows the percent viability of the CHO-K1 cells after 4-day treatment with saponin at concentrations ranging from 12 to 100 μg/mL measured by the MTT assay. It was found that saponin at concentrations 25 and 50 μg/mL caused 70% and 50% cell killing, respectively.
Figure 1.
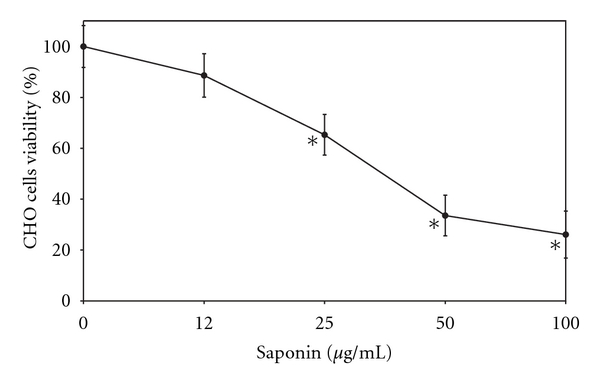
Percentage of the viability of CHO-K1 cells following treatment with saponin measured by the MTT assay; mean of three experiments ± SD. *P < 0.005.
3.1.2. Apoptosis
Table 1 shows the level of apoptosis measured by flow cytometer with the Annexin V-FITC kit and propidium iodide staining according to [16]. Saponin dose-depended lyses of CHO-K1 cells were observed, as in MTT test (Figure 1). The number of early and late apoptotic cells was detected after incubation with saponin in a concentration range of 12–50 μg/mL. The flow cytometric analyses of the CHO-K1 cells population showed that saponins at concentrations of 12–50 μg/mL significantly increased the percentage of early apoptotic cells up to 30.88%. It was in a saponin dose-dependent manner compared with the untreated control. The increase in the number (6.21%) of late apoptosis was notice at the dose of 12 μg/mL of saponin. The necrosis of cells varied and did not excite 2.52% at saponin dose 50 μg/mL.
Table 1.
Percentage of early and late apoptotic and necrotic CHO-K1 cells following treatment with saponin measured by flow cytometry; mean of three experiments ± SD. IP: propidium iodide.
| Saponin (μg/mL) | Normal cells (Annexin−/IP−) | Apoptosis | Necrosis (Annexin−/IP+) | |
|---|---|---|---|---|
| Early (Annexin+/IP−) | Late (Annexin+/IP+) | |||
| 0 | 96.08 ± 0.07 | 0.07 ± 0.06 | 2.53 ± 0.06 | 0.69 ± 0.06 |
| 12 | 85.75 ± 0.62 | 5.99 ± 0.33 | 6.21 ± 0.19 | 2.06 ± 0.13 |
| 25 | 79.73 ± 2.52 | 18.67 ± 2.49 | 1.89 ± 0.04 | 0.70 ± 0.04 |
| 50 | 64.59 ± 0.32 | 30.88 ± 0.10 | 2.09 ± 0.16 | 2.52 ± 0.17 |
3.1.3. Hemolysis Assay
In addition to CHO-K1 cell line, the human red blood cells (RBCs) were used as hemolysis marker. Figure 2 shows the amount of hemoglobin released from RBCs that was determined by the absorbance at 435, 540, and 575 nm in the presence of saponin at concentrations ranging from 3 to 100 μg/mL. We observed a statistically significant, dose-depended increase in hemolysis at concentrations from 25 up to 100 μg/mL. The proportion of pick at 435, 540, and 575 nm was similar. This may suggest that porfirine rings and iron molecules are not released from hemoglobin, and observed hemolysis is the effect of RBCs wall destruction. The saponin in range 3–12 μg/mL does not induce hemolysis. This indicates that critical amount of saponin molecules 25 μg/mL is required to destabilize cell wall of erythrocytes.
Figure 2.
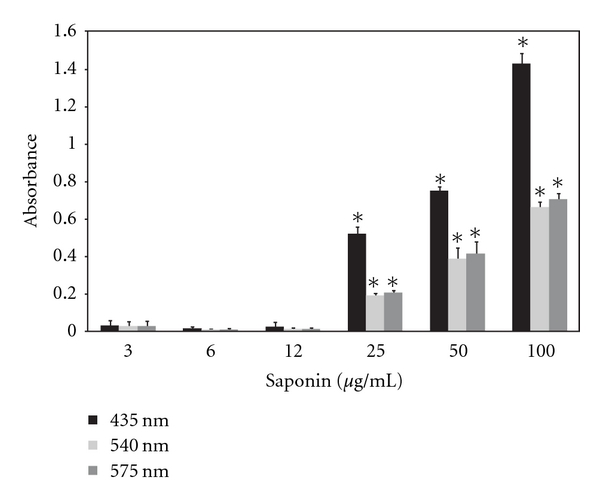
Hemolysis of human erythrocytes incubated with saponin. Hemolysis was measured at 435, 540, and 570 nm; mean of three independent experiments ± SD. *P < 0.005.
3.2. The Effect of Saponin on Bacterial Cells
3.2.1. The Effect of Saponin on Bacterial Growth
Figure 3(a) shows the effect of saponin at concentrations ranging from 1 to 12 μg/mL on the growth of the six clinical E. coli strains incubated for 18 h at 37°C in LB medium. The presence of saponin does not inhibite E.coli cells growth. It is worth to notice that we observed statistically significant increases in CFU of the multidrug resistant- (MDR-) E. coli strains in the presence of saponin at a concentration of 6 and 12 μg/mL in LB medium, in contrast to growth of bacterial strains in M9 medium which was influenced by saponin presence (Figure 3(b)).
Figure 3.
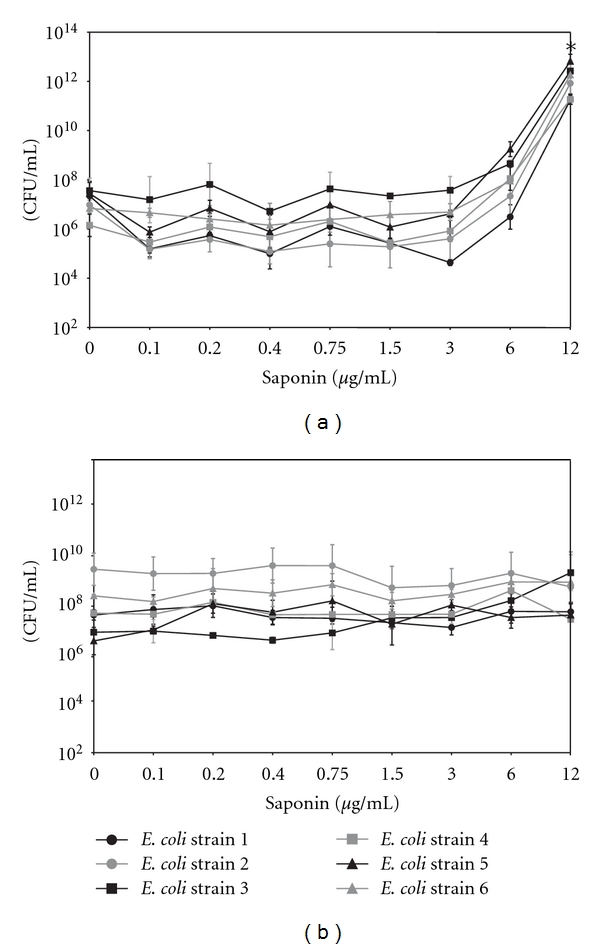
The effect of saponin on the growth of the E. coli strains incubated for 18 h at 37°C in LB (a) and M9 medium (b); mean of three independent experiments ± SD. *P < 0.005.
3.2.2. The Effect of Saponin and Antibiotics on Bacterial Growth
In next experiment, dose of 12 μg/mL of saponin in LB medium was used for testing antibiotic action against E. coli cells. Figure 4 shows the effect of the presence of saponin and ampicillin, streptomycin, or ciprofloxacin at concentrations ranging from 25 to 400 μg/mL on the growth of the clinical E. coli strains incubated for 18 h at 37°C in LB medium. All six strains were sensitive to ciprofloxacin and resistant to ampicillin and streptomycin. The addition of saponin caused in strains 3 and 6 that ciprofloxacin was less effective (Figure 4 compares open circle and open triangle). Having a similar effect, increases of amount of cells were observed with ampicillin or streptomycin and saponin mixtures in all strains tested (black or gray circle and triangle, Figure 4). The presented results indicate that saponin enhances clinical E. coli cells growth in the presence of antibiotics.
Figure 4.
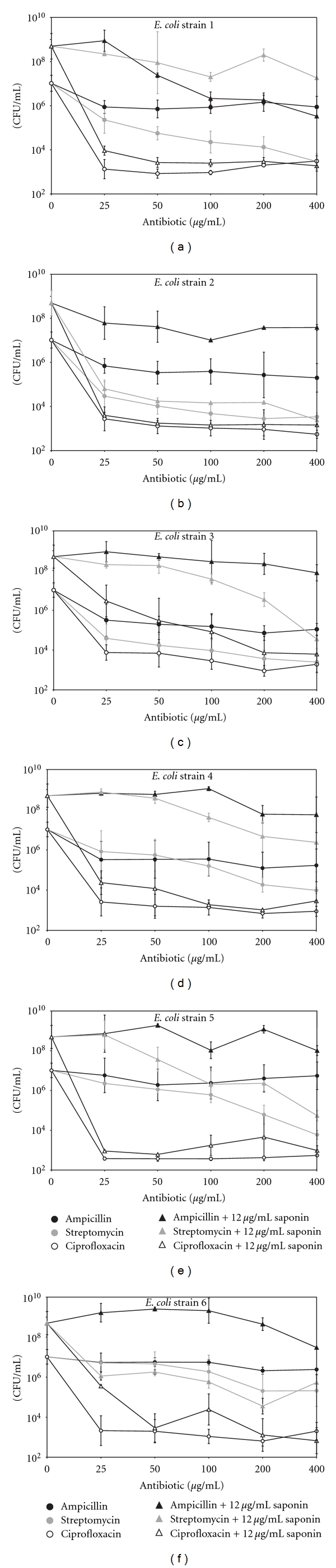
The effect of saponin at 12 μg/mL and ampicillin, streptomycin, or ciprofloxacin at concentrations ranging from 25 to 400 μg/mL on the growth of the E. coli strains incubated for 18 h at 37°C in LB medium; mean of three independent experiments ± SD.
4. Discussion
Medically desired situation is the concentration of saponins with antibacterial properties that has minimal cytotoxic activity against eukaryotic cells. Saponins possess detergent-like properties and might increase the permeability of bacterial cell membranes without destroying them [19]. In theory, this activity might facilitate antibiotic influx through the bacterial cell wall membrane. In the presented studies, we compared saponins' activities against prokaryotic and eukaryotic cells. We used commercial saponin from Quillaja saponaria, which is one of the major sources of industrial triterpenoid saponins. Saponins extracts from Quillaja saponaria have been in practical use as foaming agents in beverages or emulsifiers in foods [20]. In our investigations, we analyzed the activity of saponin from Quillaja saponaria against CHO-K1 cell line. The saponin extracts increase the number of early apoptotic cells in a dose-dependent manner. Saponins do not induce significantly necrosis and late apoptosis. Effect of saponin on membrane receptors for apoptosis was shown also by others [5]. In our studies, we also have shown that saponin at concentrations higher than 12 μg/mL has significantly cytotoxic effect on CHO-K1 cells. Similar effect was observed with human erythrocytes at dose 25 μg/mL of saponins. Interestingly, lower doses do not lyse of RBC at all. That may indicate a defined amount of saponins intercalating to cell wall membranes of erythrocytes needed to lyse it. To analyze the antibacterial properties of saponin, we use saponin at 12 μg/mL taking into consideration its minimal cytotoxic effect against eukaryotic cells. Saponin does not inhibites E. coli growth in minimal M9 and Luria broth. In contrary, we observed statistically significant increases in CFU/mL of six tested MDR E. coli strains in the presence of saponin at a concentration of 12 μg/mL. This effect was not observed on bacterial growth in M9 minimal medium. Our study of analysis of susceptibility of clinical E. coli strains against ampicillin, streptomycin, and ciprofloxacin in presence of saponin at a concentration of 12 μg/mL confirmed that saponins enhance bacteria growth, even in antibiotic presence.
In conclusion, we presented varied potencies of saponins against eukaryotic and prokaryotic cells. That is probably due to differences on cell membranes of two cell types. The biological properties of chemical substances potentially useful in clinical use should be characterized by their defined activity in correlation with the optimal concentration.
Acknowledgments
The work was supported by a Grant no. N N304 044639. The authors would like to thank Bonita Durnaś from the Department of Microbiology, Holy Cross Cancer Center in Kielce, Poland, for the collected E. coli strains and Marta Kowalik, Marianna Jagódka, and Katarzyna Pastuszka for technical support.
References
- 1.Sparg SG, Light ME, Van Staden J. Biological activities and distribution of plant saponins. Journal of Ethnopharmacology. 2004;94(2-3):219–243. doi: 10.1016/j.jep.2004.05.016. [DOI] [PubMed] [Google Scholar]
- 2.Leung YM, Ou YJ, Kwan CY, Loh TT. Specific interaction between tetrandrine and Quillaja saponins in promoting permeabilization of plasma membrane in human leukemic HL-60 cells. Biochimica et Biophysica Acta. 1997;1325(2):318–328. doi: 10.1016/s0005-2736(97)00002-3. [DOI] [PubMed] [Google Scholar]
- 3.Sen S, Makkar HPS, Muetzel S, Becker K. Effect of Quillaja saponaria saponins and Yucca schidigera plant extract on growth of Escherichia coli. Letters in Applied Microbiology. 1998;27(1):35–38. doi: 10.1046/j.1472-765x.1998.00379.x. [DOI] [PubMed] [Google Scholar]
- 4.Bachran C, Sutherland M, Heisler I, Hebestreit P, Melzig MF, Fuchs H. The saponin-mediated enhanced uptake of targeted saporin-based drugs is strongly dependent on the saponin structure. Experimental Biology and Medicine. 2006;231(4):412–420. doi: 10.1177/153537020623100407. [DOI] [PubMed] [Google Scholar]
- 5.Chwalek M, Lalun N, Bobichon H, Plé K, Voutquenne-Nazabadioko L. Structure-activity relationships of some hederagenin diglycosides: haemolysis, cytotoxicity and apoptosis induction. Biochimica et Biophysica Acta. 2006;1760(9):1418–1427. doi: 10.1016/j.bbagen.2006.05.004. [DOI] [PubMed] [Google Scholar]
- 6.Gögelein H, Hüby A. Interaction of saponin and digitonin with black lipid membranes and lipid monolayers. Biochimica et Biophysica Acta. 1984;773(1):32–38. doi: 10.1016/0005-2736(84)90547-9. [DOI] [PubMed] [Google Scholar]
- 7.Myszka H, Bednarczyk D, Najder M, Kaca WS. Synthesis and induction of apoptosis in B cell chronic leukemia by diosgenyl 2-amino-2-deoxy-β-D-glucopyranoside hydrochloride and its derivatives. Carbohydrate Research. 2003;338(2):133–141. doi: 10.1016/s0008-6215(02)00407-x. [DOI] [PubMed] [Google Scholar]
- 8.Baumann E, Stoya G, Völkner A, Richter W, Lemke C, Linss W. Hemolysis of human erythrocytes with saponin affects the membrane structure. Acta Histochemica. 2000;102(1):21–35. doi: 10.1078/0065-1281-00534. [DOI] [PubMed] [Google Scholar]
- 9.Segal R, Shatkovsky P, Milo Goldzweig I. On the mechanism of saponin hemolysis. I. Hydrolysis of the glycosidic bond. Biochemical Pharmacology. 1974;23(5):973–981. doi: 10.1016/0006-2952(74)90027-6. [DOI] [PubMed] [Google Scholar]
- 10.Winter WP. Mechanism of saponin induced red cell hemolysis: evidence for the involvement of aquaporin CHIP28. Blood. 1994;84(supplement 1–10, abstract 445) [Google Scholar]
- 11.Arabski M, Wasik S, Dworecki K, Kaca W. Laser interferometric and cultivation methods for measurement of colistin/ampicilin and saponin interactions with smooth and rough of Proteus mirabilis lipopolysaccharides and cells. Journal of Microbiological Methods. 2009;77(2):178–183. doi: 10.1016/j.mimet.2009.01.020. [DOI] [PubMed] [Google Scholar]
- 12.O’Flaherty S, Ross RP, Coffey A. Bacteriophage and their lysins for elimination of infectious bacteria: Review article. FEMS Microbiology Reviews. 2009;33(4):801–819. doi: 10.1111/j.1574-6976.2009.00176.x. [DOI] [PubMed] [Google Scholar]
- 13.Montdargent B, Letourneur D. Toward new biomaterials. Infection Control and Hospital Epidemiology. 2000;21(6):404–410. doi: 10.1086/501782. [DOI] [PubMed] [Google Scholar]
- 14.Mamelli L, Petit S, Chevalier J, et al. New antibiotic molecules: bypassing the membrane barrier of gram negative bacteria increases the activity of peptide deformylase inhibitors. PLoS ONE. 2009;4(7, article e6443) doi: 10.1371/journal.pone.0006443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Majsterek I, Arabski M, Czechowska A, et al. Imatinib (STI571) inhibits DNA repair in human leukemia oncogenic tyrosine kinase-expressing cells. Zeitschrift fur Naturforschung C. 2006;61(11-12):896–902. doi: 10.1515/znc-2006-11-1219. [DOI] [PubMed] [Google Scholar]
- 16.Darzynkiewicz Z. Current Protocols in Cell Biology. John Wiley & Sons; 2003. [Google Scholar]
- 17.Arabski M, Gwoździnski K, Sudak B, Kaca W. Effects of Proteus mirabilis lipopolysaccharides with different O-polysaccharide structures on the plasma membrane of human erythrocytes. Zeitschrift fur Naturforschung C. 2008;63(5-6):460–468. doi: 10.1515/znc-2008-5-624. [DOI] [PubMed] [Google Scholar]
- 18.Sambrook J, Russell DW. Molecular Cloning: A Laboratory Manual. 3rd edition. New York, NY, USA: Cold Spring Harbor Laboratory Press; 1989. [Google Scholar]
- 19.Jacob MC, Favre M, Bensa JC. Membrane cell permeabilisation with saponin and multiparametric analysis by flow cytometry. Cytometry. 1991;12(6):550–558. doi: 10.1002/cyto.990120612. [DOI] [PubMed] [Google Scholar]
- 20.Pelah D, Abramovich Z, Markus A, Wiesman Z. The use of commercial saponin from Quillaja saponaria bark as a natural larvicidal agent against Aedes aegypti and Culex pipiens. Journal of Ethnopharmacology. 2002;81(3):407–409. doi: 10.1016/s0378-8741(02)00138-1. [DOI] [PubMed] [Google Scholar]