

Abstract
We investigated the antioxidant potential, cytotoxic effect, and TNF-α inhibition activity with NF-κB activation response in a chloroform fraction of Centratherum anthelminticum seeds (CACF). The antioxidant property of CACF was evaluated with DPPH, ORAC, and FRAP assays, which demonstrated significant antioxidant activity. The cytotoxicity of CACF was tested using the MTT assay; CACF effective inhibitory concentrations (IC50) for A549, PC-3, MCF-7, and WRL-68 cells were 31.42 ± 5.4, 22.61 ± 1.7, 8.1 ± 0.9, and 54.93 ± 8.3 μg/mL, respectively. CACF effectively and dose-dependently inhibited TNF-α release, in vitro and in vivo. CACF inhibited TNF-α secretion in stimulated RAW264.7 macrophage supernatants with an IC50 of 0.012 μg/mL, without affecting their viability; the highest dose tested reduced serum TNF-α by 61%. Acute toxicity testing in rats revealed that CACF was non-toxic at all doses tested. Matching the cytotoxic activity towards a mechanistic approach, CACF dose-dependently exhibited in vitro inhibitory effects against the activation of NF-κB translocation in MCF-7 cells. Preliminary phytochemical screening with GC/MS analysis detected 22 compounds in CACF, of which morpholinoethyl isothiocyanate was the most abundant (29.04%). The study reveals the potential of CACF in the treatment of breast cancer and in oxidative stress conditions with associated inflammatory responses.
1. Introduction
Since time immemorial, traditional herbs have been used as remedies for several diseases [1]. Even though new synthetic drugs are available, traditional medicine is still being utilized today as part of primary healthcare in several parts of the world. Traditional medicine is often sought because it is economical, readily available, and trusted by its advocate [2]. Studies on herbal medicines have shown the positive correlation of traditional claims and scientific data [3]. Some of these traditional treatments are part of clinical practice, and few are found to be not beneficial [4]. Support for traditional medicine and the numerous natural products with biological activity have led to multidisciplinary investigations utilizing preliminary screening procedures as well as advanced mechanistic studies to develop drugs which may be used clinically.
Centratherum anthelminticum (L.) KUNTZE (family: Asteraceae) is an erect, pubescent annual herb found widely in the Indian subcontinent which is locally known as “Somraj,” and its seeds are known as “Kalijiri” in Hindi [5, 6]; scientific synonyms for this plant include Vernonia anthelmintica and Conyza anthelmintica, among others. This plant is used extensively in Ayurveda for the treatment of cough and diarrhoea, as well as an anthelmintic, stomachic, diuretic, and antiphlegmatic agent [5, 7]. Experimental studies proved the pharmacological effects of this plant's seeds extract, including antihelminthic [8], larvicidal [9], antipyretic [10], antifilarial [11], antihyperglycemic [12], antimicrobial [13], and diuretic [14] activities. In addition to primary metabolites, the seeds of this plant contain glycosides, phenolic compounds, tannins, flavonoids, saponins, and sterols [15]. Examples of secondary metabolites found in C. anthelminticum include the following: flavonoids such as 2′,3,4,4-tetrahydroxychalcone (Butein); 5,6,7,4′-tetrahydroxy flavone and 7,3′,4′-trihydroxydihydroflavone [16]; sterols such as sterol-4-alpha-methylvernosterol, vernosterol, and avernosterol [17]; steroids such as (24a/R)-stigmasta-7-en-3-one, 24(a/R)-stigmasta-7, 9(11)-dien-3-one, 24(a/S)-stigmasta-5 and 22-dien-3β-ol, stigmasta-7, and 22-dien-3β-ol [18].
While several studies investigated the pharmacological effects of this plant, the antiinflammatory and anti-cancer activities are yet to be investigated. Inflammation is a natural defense mechanism by the host which plays a role in various diseases. Immune cells are particularly important in inflammation because they orchestrate the release of several mediators such as cytokines, prostaglandins, and nitric oxide, which play a part in the defense process. However, uncontrolled production of these mediators is associated with tissue damage due to oxidative stress [19]. Oxidative stress resulting from reactive oxygen and nitrogen species (ROS and RNS) was shown to cause DNA mutations and cell death and affect cell proliferation. Cells which survive the DNA damage caused by oxidative stress are likely to have aberrant repair mechanisms, the proliferation of these genetically instable cells might eventually progress toward carcinogenesis. Therefore, this study looked into the antioxidant approaches, the in vitro and in vivo TNF inhibition activity counting cytotoxic effect in the chloroform fraction with further evaluation of linkage between NF-κB activation on breast cancer (MCF-7) cell lines.
2. Materials and Methods
2.1. Cell Lines and Reagents
All cell lines were purchased from ATCC (Rockville, MD, USA). RPMI medium, penicillin and streptomycin solution, and phosphate buffer Saline (PBS) were purchased from Invitrogen (Rockville, MD, USA). MTT, DMSO, and heat-inactivated fetal bovine serum and 0.25% trypsin solution were purchased from Sigma-Aldrich Chemicals (Saint Louis, MO, USA). Cell culture treated 96-well plates, and cell culture flasks were purchased from Orange Scientific (Braine-l'Alleud, Belgium). PBS 75 nM, pH Dulbecco's Modified Eagle Medium (DMEM), phosphate buffered saline, Hanks' balanced salt solution (HBSS), and 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) were from Invitrogen (Carlsbad, USA). Fetal bovine serum (FBS), LPS from E. coli serotype 0111:B4, and dimethylsulfoxide (DMSO) were obtained from Sigma (St. Louis, USA). Murine TNF-α ELISA kit was from eBioscience (San Diego, USA). NF-κB activation kit from Thermo Scientific Cellomics, fluorescence sodium salt, DPPH and FRAP reagents, Pentoxifylline and Paclitaxel (Sigma-Aldrich), AAPH, Quercetin, and Trolox were from Sigma-Aldrich Chemicals (S. Louis, MO, USA). Plates were read using Chameleon V Multilabel microplate reader (Hidex, Turku; Finland) in 96-well format black plate.
2.2. Plant Materials
The seeds of the plant Centratherum anthelminticum were procured from the medicinal plant cultivation zone of Amritum Bio-Botanica Herbs Research Laboratory Pvt. Ltd, Betul, Madhya Pradesh, India. The seeds of the plant were authenticated by the Quality Control department of the company itself. Voucher specimen (CA-9) is deposited in the Pharmacology Department of the University of Malaya, Malaysia.
2.3. Extraction Procedure
The coarsely powdered seeds (100 g) were extracted with water : ethanol (80 : 20) using a Soxhlet extractor for 24 h (Figure 1). The solvent was completely evaporated using rotary evaporator. The brown viscous crude extract weighing (49% w/w) obtained was further fractionated successively with hexane, chloroform and methanol, then the solvent from each fraction was completely recovered with the help of rotary evaporator under reduced pressure. After drying, the final yields with hexane (CAHF), chloroform (CACF), and methanol fraction (CAMF) were (19.41% w/w), (4.11 w/w), and (11.3% w/w), respectively. Then the dried fractions were kept below −20°C before being used. The in pilot bioactivity testing, chloroform fraction (CACF) had shown the positive effect and hence was chosen for further analysis.
Figure 1.
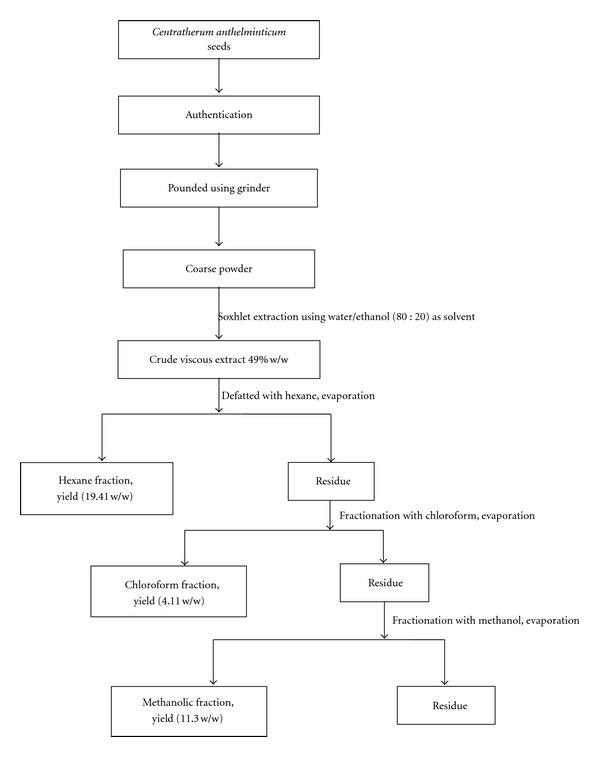
Schematic diagram of extraction/fractionation of C. anthelminticum seeds.
2.4. Animals
Altogether 72 Sprague Dawley rats of 150–200 g were obtained from the University of Malaya Medical Centre Animal House and maintained under standard conditions of lighting (12 h of light and darkness) and nutrition (food and water ad libitum) throughout the experimental period. Studies were performed in accordance with the Medical Research Council Guidelines on Ethics in Animal Experimentation. This study was approved by the Animal Experimentation ethics committee at the University of Malaya Medical Center-(UMMC), Animal Ethics no: FAR/10/11/2008/AA(R).
2.5. Total Phenolic Content (TPC)
TPC of CACF was determined using Folin-Ciocalteu method [20]. CACF was prepared in a concentration of 10 mg/mL in methanol. Five microliters of this solution were transferred to 96-well mircoplate (TPP, USA). To this, 80 μL of Folin-Ciocalteu reagent (1 : 10) was added and mixed thoroughly. After 5 min, 160 μL of sodium bicarbonate solution (NaHCO3 7.5%) was added, and the mixture was allowed to stand for 30 min with intermittent shaking. Absorbance was measured at 765 nm using microplate reader (Molecular Devices, Sunnyvale, USA). The TPC was expressed as gallic acid equivalent (GAE) in mg/g extract and obtained from the standard curve of gallic acid. The gallic acid standard curve was established by plotting concentration (mg/mL) versus absorbance (nm) (y = 0.001x + 0.045; R 2 = 0.9975), where y is absorbance and x is concentration in GAE (n = 3).
2.6. Antioxidant Activity of C. anthelminticum
2.6.1. 1,1-Diphenyl-2-picrylhydrazyl (DPPH) Radical Scavenging Activity of CACF
The scavenging activity of CACF on DPPH was determined using the method described in [21]. This method is based on the reduction of purple DPPH to a yellow-colored diphenyl picrylhydrazine. Changes in color were measured at 518 nm. CACF was tested at final concentrations ranging from 600 to 10 μg/mL in ethanol. One milliliter of 0.3 mM DPPH ethanol solution was added to 2.5 mL of sample solution of different concentrations to make the test solutions, while 1 mL of ethanol was added to 2.5 mL of samples to make the blank solutions. The negative control (blank) consisted of 1 mL DPPH solution plus 2.5 mL of ethanol. These solutions were allowed to react at room temperature for 30 minutes in the dark. The absorbance values were measured at 518 nm and converted into percentage antioxidant activity using the following equation:
| (1) |
where A B is the absorption of blank sample; A A is the absorption of tested samples.
The IC50 as well as the kinetics of DPPH scavenging activity was determined. Ascorbic acid and butylated hydroxytoluene (BHT) was used as a positive control in this assay.
2.6.2. ORAC Antioxidant Activity Assay
The oxygen radical absorbance capacity (ORAC) assay was done based on the procedure described earlier by Choi et al. [21] with slight modifications. Briefly, 175 μL of the sample/blank were dissolved with PBS at concentrations of 160 μg/mL, pH 7.4. Serial dilutions of the standard Trolox were prepared from 75 mM. The assay was performed in 96-well black microplates 25 μL of samples (CACF), standard (Trolox), blank (solvent/PBS), or the positive control (quercetin) was added to the wells. Subsequently, 150 μL of fluorescent sodium salt solution was added, and the plate was then incubated for 45 minutes at 37°C. Twenty five microliters of 2,20-azobis (2-amidinopropane) dihydrochloride (AAPH) solution was added for a total volume of 200 μL/well. Fluorescence was recorded until it reached zero (excitation at 485 nm, emission at 535 nm) using a fluorescence spectrophotometer (Perkin—Elmer LS 55), equipped with an automatic thermostatic autocell holder at 37°C. Data were collected every 2 mins for a duration of 2 hrs and were analyzed by calculating the differences of areas under the fluorescein decay curve (AUC) between the blank and the sample. Values were expressed as Trolox equivalents.
2.6.3. Ferric Reducing/Antioxidant Power (FRAP) Assay
The FRAP assay was slightly modified from the method of Benzie and Strain [22]. The stock solutions included 300 mM acetate buffer (pH 3.6), 10 mM TPTZ (2,4,6-tripyridyl-s-triazine) solution in 40 mM HCl, and 20 mM FeCl3 ·6H2O. The fresh working solution was prepared by mixing 25 mL acetate buffer, 2.5 mL TPTZ, and 2.5 mL FeCl3 ·6H2O. The temperature of the solution was raised to 37°C before use. CACF (10 μL) was allowed to react with 190 μL of the FRAP solution for 30 min in the dark. Colorimetric readings of the product ferrous tripyridyltriazine complex were taken at 593 nm. The standard curve was linear between 200 and 1000 μM FeSO4. Results are expressed as μM Fe (II)/g dry mass and compared with those of Ascorbic acid and BHT.
2.7. Cell Viability and Cytotoxicity
2.7.1. Cell Culture
All cell lines were obtained from American Type Cell Collection (ATCC) and maintained in a 37°C incubator with 5% CO2 saturation. Human breast carcinoma (MCF-7) and normal hepatic (WRL-68) cell lines were maintained in Dulbecco's modified Eagle's medium (DMEM), while nonsmall cell lung cancer cells (A549) and prostate adenocarcinoma cells (PC-3) were maintained in RPMI medium. Both media were supplemented with 10% fetus calf serum (FCS), 100 units/mL penicillin, and 0.1 mg/mL streptomycin. Cells were cultured using standard aseptic techniques and were seeded at the indicated densities below.
2.7.2. Cellular Viability
The inhibitory effects of CACF on the indicated cell lines' growth were tested using the MTT assay [23]. Cells were seeded at a density of 1 × 105cells/mL in a 96-well plate and incubated for 24 hours at 37°C and 5% CO2. Cells were then treated with CACF and incubated for another 24 hours, after which the MTT solution at 2 mg/mL was added for 1 hour. The insoluble formazan product was dissolved in DMSO, and absorbance was measured at 570 nm using Plate Chameleon V microplate reader (Hidex, Turku, Finland). Results were expressed as a percentage calculated from the ratio of absorbance of treated cells to untreated cells. The concentration that caused a 50% loss of cell growth (IC50) was used to measure the CACF growth inhibition potency.
2.8. TNF-α Inhibition Activity of CACF
2.8.1. In Vitro Cell Viability and TNF-α Production Assays
Murine macrophage cells RAW 264.7 were seeded in 96-well plates at 5 × 105cells/mL. Cells were either left untreated in DMEM or pretreated with CACF at the indicated concentrations for 30 minutes. Cell stimulation, viability, and TNF production measurements were conducted exactly as in [24]. Percentage viability was calculated as follows: cell viability (%) = [(OD570 (sample)/OD570 (control)) ×100].
Whereas percentage TNF inhibition was calculated as follows:
| (2) |
where control indicates cells treated in LPS alone.
2.8.2. Measurement of In Vivo Serum TNF
Healthy male rats were selected and divided into 6 groups (n = 8 for each group). The rats were pretreated with the indicated solutions for 30 minutes before lipopolysaccharide (LPS) stimulation: group 1 was pretreated intraperitoneally (i. p.) with 1 mL of PBS alone, group 2 with dexamethasone (6 mg/kg BW) in 25% DMSO, group 3 with CACF (25 mg/kg BW) in 25% DMSO, group 4 with CACF (50 mg/kg BW) in 25% DMSO, and group 5 with CACF (100 mg/kg BW) in 25% DMSO. Groups 1–5 were treatment groups stimulated with LPS, while group 6 made up the untreated negative control in which rats were given a solution of 25% DMSO in PBS and no LPS. Subsequent to pretreatment, LPS (1 mg/kg) was then administered in 1 mL of pyrogen-free normal saline i. p for five treatment groups, and PBS was administered i. p. for the negative control group. Blood was withdrawn from the animals under ether anesthesia after 90 minutes of LPS or PBS administration. Serum was collected and stored at −80°C until analysis. Serum levels of TNF-α were determined using rat TNF-α ELISA kit according to the manufacturer's protocol (e Bioscience, San Diego, USA).
2.9. Acute Toxicity Study
Healthy adult rats of either sex were divided into 4 groups (n = 6) and were orally fed with increasing doses of CACF: 10, 20, 100, and 1000 mg/kg body weight (BW). The rats were observed continuously for 2 hours for behavioral, neurological, and autonomic profiles and after 24 and 72 hours for any lethality. All procedures were according to the guidelines stated by OECD.
2.10. NF-κB Translocation Assay
NF-kB translocation in MCF-7 cells was examined using NF-kB activation HCS kit which contains Hoechst 33342 and Alexa Fluor 488 conjugated anti-NF-kB dyes. MCF-7 cells were seeded into 96-well plates (Perkin-Elmer Inc., Wellesley, MA, USA) at 6000 cells/well. After overnight, cells were treated with different concentrations of CACF for 1 hr, followed by treatment with 10 μg/mL TNF-α for another 30 minutes. Fixation, permeabilization, and immunofluorescence staining of cells were performed according to the manufacturer's instructions. ArrayScan reader was used to quantify the difference between the intensity of nuclear and cytoplasmic NF-κB-associated fluorescence, reported as translocation parameter.
2.11. Identification and Chemical Analysis Using GC-MS
Gas chromatography mass spectrometry (GC-MS) analysis of the selected fraction was carried out on a Shimadzu GC-17 A network GC system coupled to a mass-selective detector: MS-QP50-50. Separation was conducted on an HP-5 MS column (30 m × 0.32 mm × 3.0 μm), with helium as the carrier gas at a flow rate of 1.0 mL/min. The injection volume was 1 μL with a split ratio of 10 : 1. The column temperature was initially held at 100°C for 3 min and then increased to 290°C at a rate of 10°C/min. The column temperature was then maintained at 290°C for 3 min. The temperatures of the injector and detector were 250°C and 280°C, respectively. Mass acquisition was performed in the range of 40–550 atomic mass units (a. m. u) using electron impact ionization at 70 eV. The major components in this sample were predicted by a spectral database matching against the library of National Institute of Standards and Technology (NIST21 and NIST Wiley).
2.12. Statistical Analysis
Experimental values were expressed as the means ± standard deviation (SD) of the number of experiments indicated in the legends. Statistical significance was assessed using one-way analysis of variance (ANOVA) followed by a multiple comparison test (Tukey's post-hoc test), were P < 0.001, P < 0.01, and P < 0.05 were considered significant. Pearson correlation coefficient was used to assess the correlation between phenolic content and antioxidant activities.
3. Results
3.1. Antioxidant Activity
Table 1 provides a summary of the antioxidant and total phenolic content of CACF. The total phenolic content of CACF was determined to be 37.16 ± 0.85 μg GAE/mg extract.
Table 1.
Antioxidant activity profile of the chloroform fraction of Centratherum anthelminticum (CACF).
| Samples | DPPH IC50, μg/mL | FRAP (μmol/L) | ORAC Equivalent conc. Trolox (20 μg/mL (μM)** IC50 | TPC μg GAE/mg | |
|---|---|---|---|---|---|
| CACF | 22.56a ± 1.4 | 1048.3a ± 21.2 | 992.34a ± 45.12 | 86.62 | 37.16 ± 0.85 |
| Ascorbic acid | 15a ± 0.3 | 6240b ± 56.2 | — | — | — |
| BHT | 17a ± 0.4 | 907.7a ± 54.8 | — | — | — |
| Quercetin | — | — | 1018.00b ± 34.82 | 74.52 | — |
*The net AUC was calculated by subtracting the blank AUC from the AUC of each sample, the standards, and the positive control. Final ORAC values were expressed as the equivalent concentration of Trolox (TE) that gives the same level of antioxidant activity as the samples at 20 μg/mL. **Means with different alphabets are statistically significant.
3.1.1. DPPH Scavenging Activity of CACF
CACF exhibited a significant dose-dependent inhibition of DPPH activity (P < 0.05), with an IC50 value of 22.56 ± 1.4 μg/mL (Table 1). Maximal DPPH scavenging activity occurred at 41 ± 1.2 μg/mL of CACF with an inhibition of 89%.
3.1.2. Ferric Reducing Antioxidant Power of CACF
CACF showed a significant dose-dependent FRAP value (P < 0.05) with a 1048.3 μmol/L for the fraction, while the positive control used in this study displayed a value of 6240 and 907.7 μmol/L for ascorbic acid and BHT, respectively (Table 1).
3.1.3. ORAC Activity of CACF
The area under the curve (AUC) was calculated for oxygen radical absorbance capacity of CACF, trolox, and the positive control quercetin. ORAC results are demonstrated in Table 1. CACF had an ORAC value of 992.34 ± 45.12 μM trolox equivalent at 20 μg/mL. On the other hand, quercetin had an ORAC value of 1018.00 ± 34.82 μM of Trolox equivalent at 5 μg/mL.
3.2. Cytotoxic Activity of CACF
To evaluate the cytotoxic activity, CACF was tested with a series of different doses on nonsmall cell lung cancer (A549), prostate cancer (PC-3), breast cancer (MCF-7), and normal hepatic cells (WRL-68), respectively. After 24 hours, cell viability was determined by the MTT assay. CACF induced cell cytotoxicity in a concentration-dependent manner. These dose titration curves allowed determination of IC50 for the CACF towards different cell lines. CACF demonstrated dose-dependent cytotoxic effects with IC50 values of 31.42 ± 5.4, 22.61 ± 1.7, 8.1 ± 0.9, and 54.93 ± 8.3 μg/mL; in A549, PC-3, MCF-7 and WRL-68, respectively (Figure 2). In line with these screening, reference drug (Paclitaxel) was used as a positive control whose IC50 on tested A549, PC-3, MCF-7 and WRL-68 were 5.675 ± 1.03, 0.37 ± 0.03, 1.583 ± 0.24, and 0.666 ± 0.05, respectively. Hence, these results point out that cell lines vary in their sensitivity.
Figure 2.
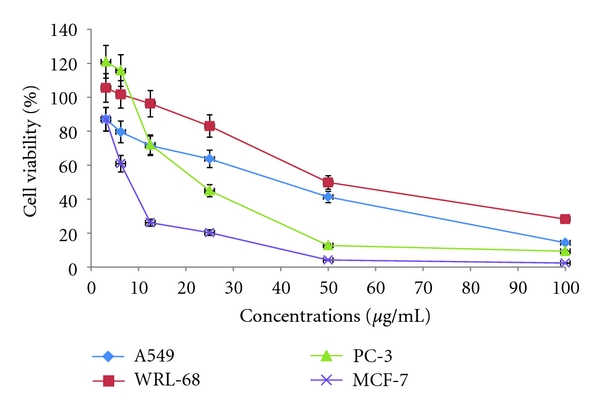
CACF was tested with a series of different doses on A549, MCF-7, PC-3, and WRL-68 cells, respectively. After 24 hours, cell viability was determined by the MTT assay. Test agents induced cell cytotoxicity in a concentration-dependent manner. These dose titration curves allowed determining IC50 for the test agents towards different cell lines. The IC50 value of CACF on the viability of A549, PC-3, MCF-7, and WRL-68 has been determined to be 31.42 ± 5.4, 22.61 ± 1.7, 8.1 ± 0.9, and 54.93 ± 8.3 μg/mL, respectively.
3.3. In Vitro Inhibitory Effects of CACF on TNF Production and RAW264.7 Cell Viability
Stimulation of RAW264.7 with LPS for 4 hours caused a significant increase in TNF-α production (Figure 3). CACF effectively and dose dependently inhibited TNF-α release with an IC50 of 0.012 μg/mL as depicted in Figure 4. CACF exhibited maximal TNF inhibition of 90% at 0.31 μg/mL. This significant inhibitory effect was observed at noncytotoxic doses ranging from 0.031 to 0.002 μg/mL (Figure 5).
Figure 3.
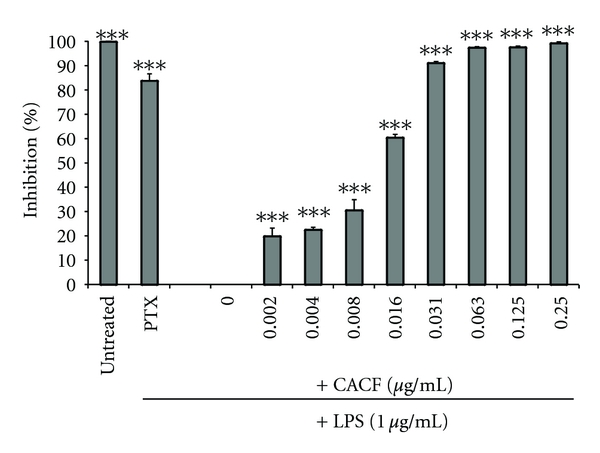
The effects of CACF on TNF production in RAW264.7 cells. Cells were pretreated with the indicated concentrations of CACF, or the TNF inhibitor pentoxifylline (PTX). The cells were stimulated with LPS (1 μg/mL) for four hours or were left untreated (DMSO). The protein concentration was measured using ELISA. Data is representative of three independent experiments and was analyzed using one-way ANOVA with Tukey's post hoc test. The inhibitory effect of CA chloroform fraction was significantly different from stimulated cells (LPS) (***P < 0.001).
Figure 4.
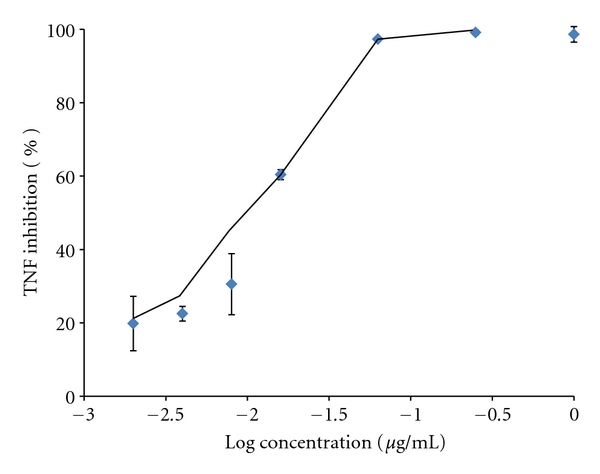
Dose-response effect of the CACF on TNF inhibition: the chloroform fraction had an IC50 of 0.012 μg/mL (LOG IC50: −1.931).
Figure 5.
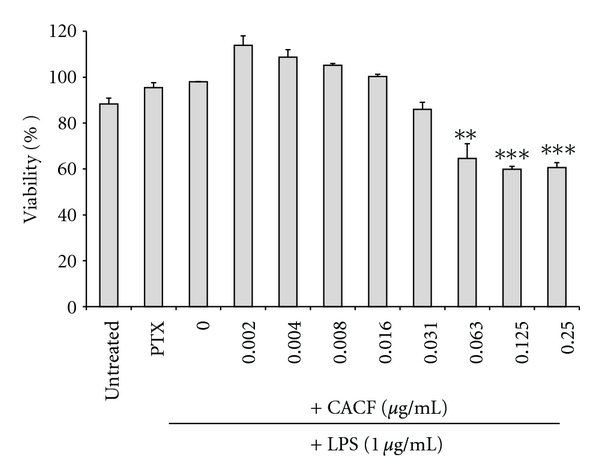
The effects of CACF on RAW264.7 cells' viability. Cells were pretreated with the indicated concentrations of CACF for 4 hours or were left untreated (DMEM). Data is the average of three independent experiments (± SD) and was analyzed using one-way ANOVA with Tukey's posttest (*P < 0.05, **P < 0.01, and ***P < 0.001).
3.4. In Vivo Activity
3.4.1. Acute Toxic Effects of CACF
The acute toxicity study revealed the nontoxic nature of CACF. There was a 100% survival rate of the animals treated with CACF at doses ranging from 10 to 1000 mg/kg. Moreover, no toxic effects were observed throughout the study.
3.4.2. The Inhibitory Effect of CACF on Serum TNF
Stimulation of animals with LPS leads to an increase in serum TNF levels up to 2 ng/mL in rats pretreated with PBS alone. On the other hand, animals pretreated with dexamethasone and CACF have shown a significant reduction in TNF levels (Figure 6). Moreover, CACF showed a dose-dependent inhibitory effect of serum TNF in LPS-stimulated rats, and this effect in rats pretreated with 100 mg/kg was almost 61%, to that of dexamethasone 67%, respectively.
Figure 6.
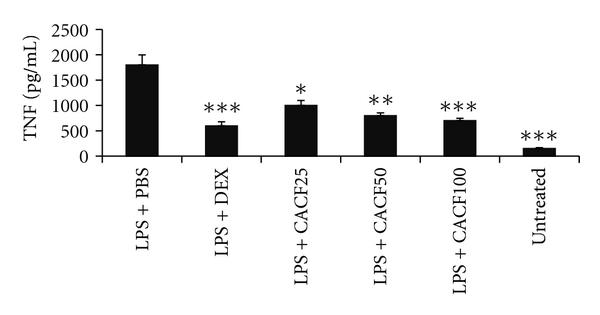
Effect of CACF on serum TNF levels in rats. Different treatment groups were pretreated i.p with 25, 50, and 100 mg/kg of (LPS + CACF), 6 mg/kg of dexamethasone (LPS + DEX) or with phosphate buffer saline (LPS + PBS), and DMSO (Untreated) for 30 mins. All the treatment groups were then either injected with 1 mg/kg of lipopolysaccharides (LPS) or with PBS for 90 mins. Blood was withdrawn, and serum TNF was quantified using ELISA. One-way ANOVA with Tukey's post-analysis was used to calculate the statistical significance among the groups when compared to LPS + PBS. ***P < 0.001, **P < 0.01, and *P < 0.05.
3.5. CACF Inhibits NF-kB Activity
In this study, we tested CACF for its in vitro inhibitory effects against NF-κB translocation activated by TNF-α and illustrated by HCS assay. CACF displayed significant inhibitory effects on the activation of NF-κB (Figure 7). In parallel, the morphological changes of NF-κB translocation indicated by immunofluorescence staining (Figure 8) showed an inhibitory effect of CACF on TNF-α-induced NF-κB translocation in a dose-dependent manner. When cells remain untreated, most of the fluorescence staining for NF-κB were in the cytoplasm and rare NF-κB staining found in nuclei area. While cells were stimulated with the TNF-α alone, NF-κB staining significantly increased in nuclei area, suggesting that NF-κB translocated from cytoplasm into the nucleus. However, MCF-7 cells were treated with 8, 4, and 2 μg/mL of CACF, and NF-κB translocation induced by TNF-α was inhibited.
Figure 7.
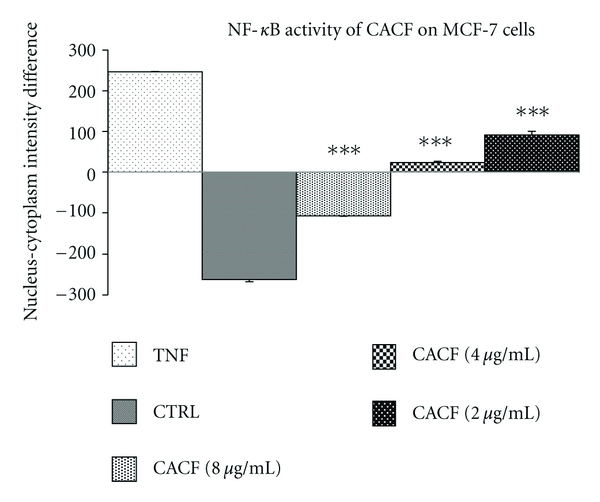
Dose-response histogram of CACF treated MCF-7 cells for 1 hours and then stimulated for 30 minutes with 10 ng/mL TNF-α for quantitative image analysis of intracellular targets.
Figure 8.
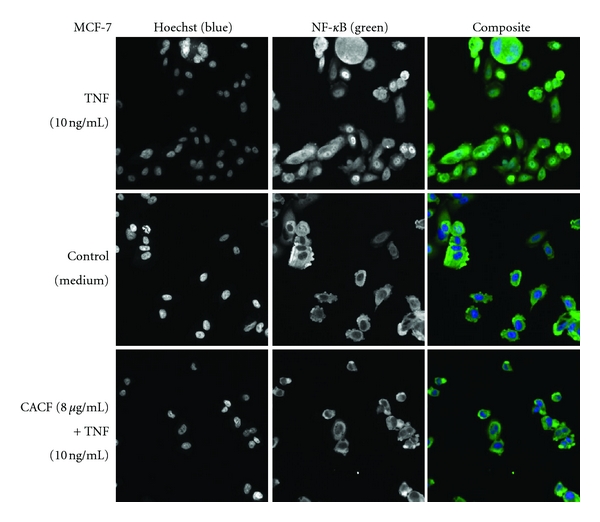
Stained MCF-7 cells were treated with CACF (8 μg/mL) for 1 hour and then stimulated for 30 minutes with 10 ng/mL TNF-α (NF-κB activation).
3.6. Chemical Composition
The possible chemical composition of CACF analyzed by GC/MS is presented in (Table 2). Based on the similarity index out of major peaks, total 22 compounds were detected in this fraction. The most abundant component comprise of 2-Morpholinoethyl isothiocyanate (29.04%), 1,E-11,Z-13-Octadecatriene (16.48%), and Octadecanoic acid, butyl ester (16.15%). Eight compounds (<5%) were not reported in Table 2 due to their minority.
Table 2.
Compounds tentatively identified in the chloroform fraction of Centratherum anthelminticum (CACF).
| Peak number | RTa | Percentage of the peakb | Molecular weight | Molecular formula | Similarity index | Compoundc |
|---|---|---|---|---|---|---|
| 1 | 5.279 | 5.97 | 116 | C6H12O2 | 89 | 2-Pentanone-4-hydroxy-4-methyl |
| 2 | 8.086 | 29.04 | 172 | C7H12N2OS | 92 | 2-Morpholinoethyl isothiocyanate |
| 3 | 11.278 | 5.97 | 180 | C6H12O6 | 85 | d-Allose |
| 4 | 11.547 | 3.62 | 346 | C22H34O3 | 65 | Drostanolone AC |
| 5 | 11.650 | 1.62 | 162 | C10H10O2 | 62 | 3(2H)-Benzofuranone, 2,6-dimethyl |
| 6 | 15.533 | 6.36 | 326 | C21H42O2 | 67 | Nonadecanoic acid, ethyl ester |
| 7 | 17.676 | 16.48 | 248 | C18H32 | 78 | 1,E-11,Z-13-Octadecatriene |
| 8 | 17.933 | 1.85 | 312 | C20H40O2 | 68 | Hexadecanoic acid, 1,1-dimethylethyl ester |
| 9 | 17.950 | 0.45 | 145 | C6H11NO3 | 46 | Adipic acid monoamide |
| 10 | 17.967 | 2.45 | 282 | C12H26O5S | 55 | d-Mannitol, 1-thiohexyl |
| 11 | 18.117 | 0.83 | 173 | C7H11NO2 | 45 | Ethyl 4-isothiocyanatobutyrate |
| 12 | 21.450 | 16.15 | 340 | C22H44O2 | 85 | Octadecanoic acid, butyl ester |
| 13 | 23.350 | 1.68 | 172 | C9H16O3 | 50 | Arabino-hetitol, 2,3 : 5,6-dianhydro-1,7-dideoxy-2,6-di-e-methyl |
| 14 | 23.942 | 2.62 | 298 | C17H14O5 | 42 | 4H-1-Benzopyran-4-one,5-hydroxy-7-methoxy-2-(3-methoxyphenyl) |
|
| ||||||
| Total | 95.09 | |||||
aRT: retention time (min).
bRelative area percentage (peak area relative to the total peak area percentage).
cCompounds listed in order of their relative area percentage.
4. Discussion
The current study revealed the pleiotropic bioactivities of the chloroform fraction of Centratherum anthelminticum (CACF) seeds. The antioxidant assays performed using the DPPH, ORAC, and FRAP methodologies revealed the free radical-scavenging possibilities of this fraction. Antioxidants are substances that may protect cells from the damage caused by unstable molecules known as free radicals. Several factors can lead to the accumulation of free radicals in the cell; examples are certain chemicals, ultraviolet radiations, inflammatory cytokines, and bacterial lipopolysacharides, and other pesticides further generate oxidative stress conditions and extend their contribution in the progression of various ailments including neurodegenerative diseases, atherosclerosis, diabetes, inflammation, and carcinogenesis [19, 25–27]. However, present work demonstrated that CACF fraction is grouped with certain antioxidant compounds. Our results warrant the probability of the CACF as a natural source of antioxidants which could be promising in hunting free radicals and treating diseases related to free radical reactions. Hence, the ability to scavenge free radicals may attenuate the several signalling pathways triggering tissue damage and inflammation, which in turn will have a protective effect on the cells [28, 29].
Having uncovered the antioxidant nature of CACF, we then turned to look at its effects in a living cell system. Cytotoxic screening models provide important preliminary data to select plant extracts or natural compounds with potential anticancer properties [30]. In this study, the cytotoxic effect of CACF was investigated by the addition of the MTT tetrazolium salt [23] to various cancer cell lines previously treated with CACF. CACF showed selective cytotoxic effect on MCF-7 compared with other tested cell lines with low IC50 value, nevertheless it was not cytotoxic to normal cell line, WRL-68. These results specify that cell lines differ in their sensitivity to the same CACF, which may be determined by multiple-cell type-specific signalling cascades and transcription factor activities [31].
In addition to that, CACF exhibited inhibition of TNF production both in vitro and in vivo without affecting cell viability and animal survival, respectively. This also warrants the ethnomedical uses of this plant in the treatment of inflammatory conditions. The ability of CACF to inhibit TNF production may attribute to several factors. Previous studies on C. anthelminticum have shown the presence of several bioactive components [18], for example, butein which is one of the secondary metabolites isolated from the seeds of this plant shown to possess both antioxidant and anti-inflammatory activities [14]. The ability of CACF to interfere with inflammatory signalling may in turn explain its cytotoxic effects on cancer cells, since these pathways are also involved in the survival, proliferation, invasion, angiogenesis and metastasis of tumours [32].
Moreover, this fraction along with pleotropic bioactivities demonstrated the in vitro inhibitory effects on cell proliferation in MCF-7 cells, which moves our line of study towards carrying out additional experiments to get a better understanding in elucidating mechanisms of breast carcinogenesis with possible future strategies. It is well known that antioxidants are known for their ability to scavenge free radicals from the various stimuli, resulting in activation of transcription factors involved in the transcription of survival and inflammatory genes such as nuclear factor kappa B (NF-κB) [24]. NF-κB is a transcription factor involved in copious inflammatory and cancer-related ailments and has developed as a foremost target in drug discovery [33]. The consequences of NF-κB transcription factors in constitutive activation includes increased survival signalling, cell proliferation, angiogenesis, and invasion, which are key features of the malignant phenotype [34]. Our study demonstrated that CACF in a concentration-dependent manner inhibited constitutive activation of NF-kB translocation stimulated by TNF-α in MCF-7 cells. TNF-α-induced transcript levels for the adhesion molecules, and this might interfer at an early stage of signaling event induced by TNF-α. This suggests that CACF may inhibit the expression of the cell adhesion molecules by interfering with the transcription of their respective genes and may inhibit either the initiation of transcription or the stability of the mRNAs encoding in these molecules. It is well understood that NF-κB is an important transcription factor involved in the gene regulation and contributing in immune and inflammatory responses, including genes encoding [35, 36]. Moreover, tumors with constitutive NF-kB activity have inherent resistance to many anticancer therapies. Thus, NF-κB is believed to play an important role in the regulation of inflammatory response associated with cancer therapy [37, 38].
Analysis of the nonpolar extractable of plant material using GC/MS has been applied before [39]. Our results demonstrate that CACF contains various bioactive components such as 2-morpholinoethyl isothiocyanate which represents 29.04%. Isothiocyanates have been shown to inhibit carcinogenesis and also useful as chemopreventive agents against cancers. They work on a variety of levels. These compounds are shown to induce apoptosis in certain cancer cell lines and in some cases and are even able to induce apoptosis in cells that are resistant to some currently used chemotherapeutic drugs [40–44].
In summary, the chloroform fraction of Centratherum anthelminticum seed (CACF) displayed a range of interrelated in vitro activities, ranging between antioxidant, cytotoxic and in vitro and in vivo TNF-α inhibition while remaining safe. Furthermore, CACF is maintained concerning mechanistic approach linked with cytotoxic activity on targeted cell line which positively inhibited NF-kB translocation in MCF-7 cells. Therefore, these results merit further pharmacological investigations with detailed phytochemical analysis on C. anthelminticum.
Acknowledgments
This study, which was carried out as part of the corresponding author's PhD research, was supported by an Institute of Research Management and Consultancy (IPPP) research grant (No.:PS144/2008C) and by a University of Malaya Research Grant (HIR: E00002-20001). We are also grateful to Nitika Rai, chief executive of Amritum Bio-Botanica Herbs Research Laboratory Pvt. Ltd., for contributing the plant material. These sources were not involved in the study design; in the collection, analysis, and interpretation of data; in the writing of the report; and in the decision to submit the article for publication.
References
- 1.Ang-Lee MK, Moss J, Yuan CS. Herbal medicines and perioperative care. Journal of the American Medical Association. 2001;286(2):208–216. doi: 10.1001/jama.286.2.208. [DOI] [PubMed] [Google Scholar]
- 2.Lambert J, Srivastava J, Vietmeyer N. Medicinal Plants: Rescuing a Global Heritage. Washington, DC, USA: World Bank Publications; 1997. [Google Scholar]
- 3.Ernst E. The risk-benefit profile of commonly used herbal therapies: Ginkgo, St. John’s Wort, Ginseng, Echinacea, Saw Palmetto, and Kava. Annals of Internal Medicine. 2002;136(1):42–53. doi: 10.7326/0003-4819-136-1-200201010-00010. [DOI] [PubMed] [Google Scholar]
- 4.Sparreboom A, Cox MC, Acharya MR, Figg WD. Herbal remedies in the United States: potential adverse interactions with anticancer agents. Journal of Clinical Oncology. 2004;22(12):2489–2503. doi: 10.1200/JCO.2004.08.182. [DOI] [PubMed] [Google Scholar]
- 5.Mehta BK, Mehta D, Itoriya A. Structure elucidation by NMR spectroscopy of a new acetylated saponin from Centratherum anthelminticum . Carbohydrate Research. 2004;339(18):2871–2874. doi: 10.1016/j.carres.2004.10.001. [DOI] [PubMed] [Google Scholar]
- 6.Rastogi RP, Mehrotra BN. Compendium of Indian Medicinal Plants. Vol. 1. New Delhi, India: Research Institute, Lucknow and Publication and Information Directorate; 1995. [Google Scholar]
- 7.Nadkarni KM. The Indian Materia Medica. Bombay, India: 1927. [Google Scholar]
- 8.Iqbal Z, Lateef M, Jabbar A, Akhtar MS, Khan MN. Anthelmintic activity of Vernonia anthelmintica seeds against trichostrongylid nematodes of sheep. Pharmaceutical Biology. 2006;44(8):563–567. [Google Scholar]
- 9.Srivastava A, Bartarya R, Tonk S, Srivastava SS, Kumari KM. Larvicidal activity of an indigenous plant, Centratherum anthelminticum . Journal of Environmental Biology. 2008;29(5):669–672. [PubMed] [Google Scholar]
- 10.Purnima A, Koti BC, Tikare VP, Viswanathaswamy AHM, Thippeswamy AHM, Dabadi P. Evaluation of analgesic and antipyretic activities of Centratherum anthelminticum (L) kuntze seed. Indian Journal of Pharmaceutical Sciences. 2009;71(4):461–464. doi: 10.4103/0250-474X.57302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Singhal KC, Sharma S, Mehta BK. Antifilarial activity of Centratherum anthelminticum seed extracts on Setaria cervi. Indian Journal of Experimental Biology. 1992;30(6):546–548. [PubMed] [Google Scholar]
- 12.Ani V, Naidu KA. Antihyperglycemic activity of polyphenolic components of black/bitter cumin Centratherum anthelminticum (L.) Kuntze seeds. European Food Research and Technology. 2008;226(4):897–903. [Google Scholar]
- 13.Sharma S, Mehta BK. In vitro antimicrobial efficacy of Centratherum anthelminticum seeds extracts. Journal of Hygiene Epidemiology Microbiology and Immunology. 1991;35(2):157–161. [PubMed] [Google Scholar]
- 14.Koti BC, Purnima A. Diuretic activity of extracts of Centratherum anthelminticum . International Journal of Green Pharmacy. 2008;2(4):228–231. [Google Scholar]
- 15.Bhatia D, Gupta MK, Gupta A, Singh M, Kaithwas G. Pharmacognosticol studies on seeds of Centratherum anthelminticum Kuntze. Natural Product Radiance. 2008;7(4):326–329. [Google Scholar]
- 16.Tian G, Zhang U, Zhang T, Yang F, Ito Y. Separation of flavonoids from the seeds of Vernonia anthelmintica Willd by high-speed counter-current chromatography. Journal of Chromatography A. 2004;1049(1-2):219–222. [PubMed] [Google Scholar]
- 17.Akihisa T, Hayashi Y, Patterson GW, Shimizu N, Tamura T. 4α-methylvernosterol and other sterols from Vernonia anthelmintica seeds. Phytochemistry. 1992;31(5):1759–1763. [Google Scholar]
- 18.Mehta BK, Mehta D, Verma M. Novel steroids from the seeds of Centratherum anthelminticum . Natural Product Research. 2005;19(5):435–442. doi: 10.1080/14786410512331330729. [DOI] [PubMed] [Google Scholar]
- 19.Gomez-Cambronero LG, Sabater L, Pereda J, et al. Role of cytokines and oxidative stress in the pathophysiology of acute pancreatitis: therapeutical implications. Current Drug Targets-Inflammation & Allergy. 2002;1(4):393–403. doi: 10.2174/1568010023344544. [DOI] [PubMed] [Google Scholar]
- 20.Turkmen N, Sari F, Velioglu YS. Effects of extraction solvents on concentration and antioxidant activity of black and black mate tea polyphenols determined by ferrous tartrate and Folin-Ciocalteu methods. Food Chemistry. 2006;99(4):835–841. [Google Scholar]
- 21.Choi CW, Kim SC, Hwang SS, et al. Antioxidant activity and free radical scavenging capacity between Korean medicinal plants and flavonoids by assay-guided comparison. Plant Science. 2002;163(6):1161–1168. [Google Scholar]
- 22.Benzie IFF, Strain JJ. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: the FRAP assay. Analytical Biochemistry. 1996;239(1):70–76. doi: 10.1006/abio.1996.0292. [DOI] [PubMed] [Google Scholar]
- 23.Mosmann T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. Journal of Immunological Methods. 1983;65(1-2):55–63. doi: 10.1016/0022-1759(83)90303-4. [DOI] [PubMed] [Google Scholar]
- 24.Achoui M, Appleton D, Abdulla MA, Awang K, Mohd MA, Mustafa MR. In vitro and in vivo anti-inflammatory activity of 17-O-acetylacuminolide through the inhibition of cytokines, Nf-κB translocation and IKKβ activity. PLoS One. 2010;5(12) doi: 10.1371/journal.pone.0015105. Article ID e15105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Raingeaud J, Gupta S, Rogers JS, et al. Pro-inflammatory cytokines and environmental stress cause p38 mitogen-activated protein kinase activation by dual phosphorylation on tyrosine and threonine. Journal of Biological Chemistry. 1995;270(13):7420–7426. doi: 10.1074/jbc.270.13.7420. [DOI] [PubMed] [Google Scholar]
- 26.Valko M, Rhodes CJ, Moncol J, Izakovic M, Mazur M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chemico-Biological Interactions. 2006;160(1):1–40. doi: 10.1016/j.cbi.2005.12.009. [DOI] [PubMed] [Google Scholar]
- 27.Lee IM, Cook NR, Manson JE, Buring JE, Hennekens CH. β-carotene supplementation and incidence of cancer and cardiovascular disease: the Women’s Health study. Journal of the National Cancer Institute. 1999;91(24):2102–2106. doi: 10.1093/jnci/91.24.2102. [DOI] [PubMed] [Google Scholar]
- 28.Chandra J, Samali A, Orrenius S. Triggering and modulation of apoptosis by oxidative stress. Free Radical Biology and Medicine. 2000;29(3-4):323–333. doi: 10.1016/s0891-5849(00)00302-6. [DOI] [PubMed] [Google Scholar]
- 29.Reilly PM, Schiller HJ, Bulkley GB. Pharmacologic approach to tissue injury mediated by free radicals and other reactive oxygen metabolites. The American Journal of Surgery. 1991;161(4):488–503. doi: 10.1016/0002-9610(91)91120-8. [DOI] [PubMed] [Google Scholar]
- 30.Itharat A, Houghton PJ, Eno-Amooquaye E, Burke PJ, Sampson JH, Raman A. In vitro cytotoxic activity of Thai medicinal plants used traditionally to treat cancer. Journal of Ethnopharmacology. 2004;90(1):33–38. doi: 10.1016/j.jep.2003.09.014. [DOI] [PubMed] [Google Scholar]
- 31.Taha MME, Abdul AB, Abdullah R, Ibrahim TAT, Abdelwahab SI, Mohan S. Potential chemoprevention of diethylnitrosamine-initiated and 2-acetylaminofluorene-promoted hepatocarcinogenesis by zerumbone from the rhizomes of the subtropical ginger (Zingiber zerumbet) Chemico-Biological Interactions. 2010;186(3):295–305. doi: 10.1016/j.cbi.2010.04.029. [DOI] [PubMed] [Google Scholar]
- 32.Aggarwal S, Ichikawa H, Takada Y, Sandur SK, Shishodia S, Aggarwal BB. Curcumin (diferuloylmethane) down-regulates expression of cell proliferation and antiapoptotic and metastatic gene products through suppression of IκBα kinase and Akt activation. Molecular Pharmacology. 2006;69(1):195–206. doi: 10.1124/mol.105.017400. [DOI] [PubMed] [Google Scholar]
- 33.Haefner B. NF-κB: arresting a major culprit in cancer. Drug Discovery Today. 2002;7(12):653–663. doi: 10.1016/s1359-6446(02)02309-7. [DOI] [PubMed] [Google Scholar]
- 34.Baldwin AS., Jr. The NF-κB and IκB proteins: new discoveries and insights. Annual Review of Immunology. 1996;14:649–681. doi: 10.1146/annurev.immunol.14.1.649. [DOI] [PubMed] [Google Scholar]
- 35.May MJ, Ghosh S. Signal transduction through NF-κB. Immunology Today. 1998;19(2):80–88. doi: 10.1016/s0167-5699(97)01197-3. [DOI] [PubMed] [Google Scholar]
- 36.Mo SJ, Son EW, Lee SR, Lee SM, Shin DH, Pyo S. CML-1 inhibits TNF-α-induced NF-κB activation and adhesion molecule expression in endothelial cells through inhibition of IkBα kinase. Journal of Ethnopharmacology. 2007;109(1):78–86. doi: 10.1016/j.jep.2006.07.006. [DOI] [PubMed] [Google Scholar]
- 37.Collins T, Read MA, Neish AS, Whitley MZ, Thanos D, Maniatis T. Transcriptional regulation of endothelial cell adhesion molecules: NF-κB and cytokine-inducible enhancers. The FASEB Journal. 1995;9(10):899–909. [PubMed] [Google Scholar]
- 38.Das KC, White CW. Activation of NF-κB by antineoplastic agents. Role of protein kinase C. Journal of Biological Chemistry. 1997;272(23):14914–14920. doi: 10.1074/jbc.272.23.14914. [DOI] [PubMed] [Google Scholar]
- 39.Jonsson P, Gullberg J, Nordström A, et al. A strategy for identifying differences in large series of metabolomic samples analyzed by GC/MS. Analytical Chemistry. 2004;76(6):1738–1745. doi: 10.1021/ac0352427. [DOI] [PubMed] [Google Scholar]
- 40.Damiano JS, Cress AE, Hazlehurst LA, Shtil AA, Dalton WS. Cell adhesion mediated drug resistance (CAM-DR): role of integrins and resistance to apoptosis in human myeloma cell lines. Blood. 1999;93(5):1658–1667. [PMC free article] [PubMed] [Google Scholar]
- 41.Johnstone RW, Ruefli AA, Lowe SW. Apoptosis: a link between cancer genetics and chemotherapy. Cell. 2002;108(2):153–164. doi: 10.1016/s0092-8674(02)00625-6. [DOI] [PubMed] [Google Scholar]
- 42.Xiao D, Powolny AA, Moura MB, et al. Phenethyl isothiocyanate inhibits oxidative phosphorylation to trigger reactive oxygen species-mediated death of human prostate cancer cells. Journal of Biological Chemistry. 2010;285(34):26558–26569. doi: 10.1074/jbc.M109.063255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Wang H, Nair MG, Strasburg GM, et al. Antioxidant and antiinflammatory activities of anthocyanins and their aglycon, cyanidin, from tart cherries. Journal of Natural Products. 1999;62(2):294–296. doi: 10.1021/np980501m. [DOI] [PubMed] [Google Scholar]
- 44.Lai KC, Huang ANC, Hsu SC, et al. Benzyl isothiocyanate (BITC) inhibits migration and invasion of human colon cancer HT29 cells by inhibiting matrix metalloproteinase-2/-9 and urokinase plasminogen (uPA) through PKC and MAPK signaling pathway. Journal of Agricultural and Food Chemistry. 2010;58(5):2935–2942. doi: 10.1021/jf9036694. [DOI] [PubMed] [Google Scholar]