

Abstract
We tested the ability of white participants to encode and retain over a brief period of time information about the identity of white and black people, using faces as stimuli in a standard change detection task and tracking neural activity using electroencephalography. Neural responses recorded over the posterior parietal cortex reflecting visual working memory activity increased in amplitude as a function of the number of faces that had to be maintained in memory. Critically, these memory-related neural responses varied as a function of participants’ implicit racial prejudice toward black people. High-prejudiced participants encoded black people faces with a lower degree of precision compared to low-prejudiced participants, suggesting that the class of mental operations affected by implicit racial prejudice includes basic cognitive mechanisms underpinning the encoding and maintenance of faces’ visual representations in visual working memory.
Keywords: implicit racial prejudice, visual working memory, electroencephalography, event-related potentials
INTRODUCTION
Interacting with people of a different race is naturally bound to several cognitive, affective and behavioral consequences (e.g. Macrae and Bodenhausen, 2000), some of which related to the degree of prejudice toward other-race members (e.g. Richeson et al., 2003). Although recent evidence suggests overt expressions of racial prejudice are decreasing (e.g. Cunningham et al., 2004), social psychological research has long disclosed that people simply tend to hide their racial prejudices in a way that largely depends on social context. Evidence for the persistence of racial prejudice comes, however, from covert measures that by-pass people’s awareness (e.g. the Implicit Association Test, IAT; Greenwald et al., 1998). Understanding the nature of racial prejudice and the class of mental operations and behaviors covertly affected by it is therefore of paramount importance and a central topic in social cognition. Recent work in neuroscience has provided critical new insights into the cognitive and neural systems that are influenced by racial prejudice.
In a seminal fMRI study, for instance, Richeson et al. (2003) explored the relationship between implicit racial prejudice and high-order aspects of human cognition, and demonstrated that high-order cognition may be affected by racial prejudice. In this study, white participants were instructed to carry out an experimental task hypothesized to hinge on so-called executive (or control) functions, following a brief interaction with black or white actors. High-prejudiced white participants (classified based on IAT scores) displayed a substantial performance reduction in the experimental task after interacting with black actors, in conjunction with an increase in blood oxygen level-dependent (BOLD) response recorded in regions of the prefrontal cortex following the exposition to black faces. These effects were reduced to absent in low-prejudiced participants, and this suggests that interracial contacts may impact cognitive control functions in a way that correlates with IAT-based individual implicit prejudice.
Although the study briefly described in the foregoing paragraph suggests that the influence of racial prejudice impacts high-level aspects of human cognition, like the active control of mental subroutines subtended in the execution of a complex cognitive task, other fMRI work demonstrated that racial prejudice exerts an influence also on low level, automatic emotional responses elicited by the exposure to other-race members. Phelps et al. (2000) reported that hemodynamic activation at the level of the amygdala is modulated by implicit prejudice toward members of a different race. In this study, white participants were exposed visually to black and white faces. On average, comparable BOLD responses were observed at the level of the amygdala when participants were exposed to black and white faces. However, individual amplitude differences obtained by subtracting BOLD responses elicited by white faces from BOLD responses elicited by black faces correlated with implicit racial prejudice estimated using IAT, suggesting that an important node of the neural circuitry subtended in emotional processing and learning is affected by implicit prejudice toward members of a different race.
The present study examines the pervasiveness of racial prejudice in visual processing demonstrating that the class of processes affected by the implicit racial prejudice includes basic cognitive and neural systems such as those involved in the encoding and maintenance of facial representations in visual working memory (VWM). Linchpin of the present study is the observation that a complete understanding of racial biases cannot be achieved without an in-depth examination of how racial prejudice influences basic cognitive processes such as those involved in face representation. It is only by understanding racial prejudice at this level that one can provide answers to fundamental questions such as whether the control mechanisms suggested by Richeson et al. (2003) are capable of affecting how we see and built-up representations of other-race people, or whether they alter people’s mood and possibly temper their emotional responses and overt behavior independently on how people process mentally other-race faces. This scope was pursued by examining whether racial prejudice influences the maintenance of visual information of other-race faces in VWM, and considering that providing evidence for the impact of racial prejudice on such basic cognitive skill may have several consequences on crucial aspects of social life such as those highlighted by the scenario hypothesized in the forthcoming paragraph.
Encoding visual information for storage in VWM is the privileged route to higher order mental processes, such as long-term memory storage, learning and response selection (Luck, 2008). Systematic deficits in the efficacy of maintenance of other-race faces representations in VWM can therefore bias the information available to higher order aspects of human cognition. If, for some reason, retaining in VWM information about the identity of members of other races is modulated by racial prejudice then this has important consequences for how we understand situations in which an expert is requested to judge the reliability of a witness to a crime committed by people of a different race. Here is a question that would naturally cross everybody’s mind: would the witness be thoroughly reliable in court when reporting about the identities of the people populating the crime scene, or could implicit racial prejudice in this imaginary situation be raised as an element during an eventual trial?
In order to understand how the retention of visual information is affected by racial prejudice, white participants were asked to perform a change detection task (e.g. Vogel and Machizawa, 2004). Change detection requires to encode and retain a memory array of visual objects for a brief interval (i.e. ∼1 s). After the interval has elapsed, memory is tested by displaying an identical test array of objects, or a test array in which one of the objects has been changed in some pre-specified physical property. The task requirement is to report whether a change between memory and test arrays had occurred or not. In the present study, white participants were asked to detect a change in identity between memory and test arrays composed of white (own-race) or black (other-race) faces. Event-related potentials (ERPs) time-locked to the onset of the memory array of faces recorded at posterior parietal sites were inspected for the presence of an ERP component that many neuroscientists have hypothesized to reflect the amount and quality of visual information retained in VWM, namely the sustained posterior contralateral negativity (SPCN; e.g. Dell'Acqua et al., 2006; Jolicœur et al., 2006a, b; also labeled contralateral delay activity, CDA, Vogel and Machizawa, 2004; Vogel et al., 2005; McCollough et al., 2007). Sessa et al. (2011) have recently shown that SPCN amplitude varies proportionally to how precisely faces are represented in VWM. In these authors’ study, faces encoded as low-precision VWM representations elicited reduced SPCN amplitude relative to faces encoded as high-precision VWM representations. In the present context, differences in SPCN as a function of racial prejudice would therefore provide clear-cut evidence that racial prejudice influences basic cognitive and neural systems involved in visual processing and memory for faces.
To assess whether racial prejudice influenced SPCN, individual estimates of memory precision for faces were estimated by subtracting the amplitude of the SPCN elicited by white faces from the amplitude of the SPCN elicited by black faces. If not null, this subtracted SPCN amplitude would indicate a difference in precision of VWM representations of own- and other-race faces during VWM maintenance. Crucially, individual subtracted SPCN amplitudes were subsequently analyzed to check for a possible correlation with implicit racial prejudice, quantified as individual IAT scores collected at the end of the memory task. Evidence of such a correlation would indicate that anti-black prejudice could affect the quality of VWM representations of black faces, in a way commensurate to the degree of prejudice held by an individual observer.
METHODS
Participants
Data were collected from 16 white students (aged 18–24 years) who reported normal or corrected-to-normal vision and no history of neurological disorders. All participants gave their informed consent according to the ethical principles approved by the University of Padova.
Stimuli and procedure
Memory task
The stimuli were grayscale digital photographs of 12 white (own-race) and 12 black (other-race) male faces selected from Minear and Park’s (2004) database, cropped with an oval-shaped mask to remove most hair, and finally scaled so as to fit in a 3.3° × 4.5° (width × height) rectangle from a viewing distance of ∼70 cm. The memory task was a spatially cued variant of a standard change detection task typically used in the field of VWM studies (e.g. Vogel and Machizawa, 2004). Examples of two different trials are reported in Figure 1a and b and described in detail in the respective captions. Memory arrays consisted of one or two faces displayed in each visual hemifield, preceded by arrow cues pointing to the hemifield including the to-be-memorized faces. The face/s included in the opposite hemifield had to be ignored. The center of each face was located at the corners of a notional 7.0° × 5.7° rectangular region centered on fixation. Following the memory array, participants were required to examine the test array for a possible change in the identity of one face, which occurred unpredictably on 50% of trials. When a change occurred, one face was replaced with a different face of the same race. The experiment consisted of 192 trials per condition (white vs black faces, and one vs two faces; each face was shown on average 32 times).
Fig. 1.

Examples of change detection task trials for the right hemifield when either (a) one black face or (b) two black faces had to be memorized. ISI: interstimulus interval. The relative distance between faces in this figure is approximately to scale with the spatial arrangement of the stimuli when displayed on screen. At the beginning of each trial, two central arrows (both pointing to the left or to the right) instructed participants to memorize the faces in only one visual hemifield of the memory array, and ignore the faces in the opposite hemifield of the memory array. Following the memory array, a blank interval elapsed before the onset of the test array. On each trial, memory and test arrays contained faces of the same race (all faces were either white or black), and trials with white and black faces were presented intermixed at random in each block. A change in identity of one face between memory and test arrays could occur unpredictably on 50% of the trials. Participants responded without speed pressure by pressing one of two appropriately labeled buttons of the computer keyboard to indicate whether a change in identity between memory and test arrays had occurred or not.
IAT
The stimuli were digital photographs of 10 white males and 10 black males different than faces used in the memory task. Attributes were 10 positive words (e.g. peace, love and paradise) and 10 negative words (e.g. disgust, vomit and earthquake). A standard race-IAT was administered to each participant following the memory task. Upon IAT administration, participants were asked to categorize visually displayed faces as belonging to white or black individuals and words as pleasant or unpleasant via button press. A detailed description of the IAT procedure is illustrated in Table 1. Racial prejudice is said to exert an effect on behavior when reaction time (RT) to faces and word concepts is modulated by stimulus–response mapping. Shorter RT when black faces and negative attributes are mapped to one button and white faces and positive attributes are mapped to the other button (prejudice-consistent block) relative to when the mapping is reversed (prejudice-inconsistent block) is taken as the hallmark of an anti-black implicit prejudice.
Table 1.
Schematic description of the IAT procedure
| Block | Type of judgment | Instruction | Number of trials |
|---|---|---|---|
| 1 | Race discrimination | Press ‘D’ to categorize pictures as Whites and ‘K’ to categorize pictures as Blacks | 20 |
| 2 | Attribute discrimination | Press ‘D’ to categorize positive words and ‘K’ to categorize negative words | 20 |
| 3 | Critical Prejudice-consistent combination | Press ‘D’ to categorize pictures of Whites and positive words and ‘D’ to categorize pictures of Blacks and negative words | 40 |
| 4 | Race discrimination-reversed | Press ‘D’ to categorize pictures as Blacks and ‘K’ to categorize pictures as Whites | 20 |
| 5 | Critical Prejudice-inconsistent combination | Press ‘D’ to categorize pictures of Blacks and positive words and ‘D’ to categorize pictures of Whites and negative words | 40 |
IAT consisted of five classification tasks administered in a sequential order on the computer screen. Blocks 1, 2 and 4 were learning blocks while blocks 3 and 5 were critical in order to compute the IAT score. The order of the critical blocks was counterbalanced across participants. In each block, participants were required to classify stimuli by pressing one of two different keys on the computer keyboard (‘D’ and ‘K’). Instructions and key assignments were displayed on the computer screen before each block. Stimuli were presented at the center of the screen and remained visible until response.
ERPs recording and analysis
EEG activity was recorded from 19 standard electrodes distributed over the scalp according to the international 10/20 system referenced to the left earlobe. The EEG was re-referenced offline to the average of the left and right earlobes. Trials contaminated by eye blinks, large horizontal eye movements or incorrect responses in the change detection task were discarded from analysis. We computed contralateral waveforms by averaging the activity recorded at right hemisphere electrodes when participants were cued to encode the face stimuli on the left side of the memory array with the activity recorded from the left hemisphere electrodes when they were cued to encode the face stimuli on the right side of the memory array. SPCN was quantified at posterior electrodes sites (P7/P8) as the difference in mean amplitude between the ipsilateral and contralateral waveforms in a time window of 300–900 ms relative to the onset of the memory array. We computed, for each participant and condition, an SPCN index reflecting race-based differential activity in VWM maintenance based on the following equation:
SPCN is a negative-going ERP component. Positive SPCN[Race] indices would thus indicate that white faces were maintained in VWM as higher resolution representations relative to black faces and negative SPCN[Race] indices would indicate the obverse.
RESULTS
IAT
Mean RTs in prejudice-consistent blocks and in prejudice-inconsistent blocks were calculated using the standard algorithm proposed by Greenwald et al. (1998). RTs were then submitted to ANOVA considering block type (prejudice-consistent vs prejudice-inconsistent) as within-subject factor and block order (prejudice-consistent first vs prejudice-inconsistent first) as between-subject factor. Shorter RTs were observed in prejudice-consistent trials (M = 650 ms, s.d. = 112) than in prejudice-inconsistent trials [M = 779 ms, s.d. = 218; F (1, 14) = 6.29; P < 0.05; 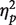 = 0.310]. No other main effect or interaction was significant (all P’s > 0.21). Individual IAT scores were calculated by subtracting the mean RT in the consistent block from the mean RT in the inconsistent block. Greater IAT scores were taken to reflect a higher degree of racial prejudice toward black people.
= 0.310]. No other main effect or interaction was significant (all P’s > 0.21). Individual IAT scores were calculated by subtracting the mean RT in the consistent block from the mean RT in the inconsistent block. Greater IAT scores were taken to reflect a higher degree of racial prejudice toward black people.
Memory task
SPCN
SPCN amplitude values were submitted to ANOVA considering race (white vs black) and number of to-be-memorized faces (one vs two) as within-subjects factors. Figure 2 illustrates SPCN (contra minus ipsi waveforms) recorded at electrode sites P7/P8 time-locked to the onset of the memory array of faces in each cell of the present experimental design. Figure 3 illustrates posterior views of isovoltage contour maps of the scalp distribution of SPCN activity in each cell of the present experimental design. Consistent with prior reports on the scalp localization of this ERP component (e.g. McCollough et al., 2007; Robitaille et al., 2010), the maps revealed a source of SPCN activity over posterior parietal and occipital areas. Prior fMRI and fNIRS investigation using the change detection task make a strong case for the involvement of neural circuitries in the intraparietal sulcus and intraoccipital sulcus as neural generators of the SPCN component (e.g. Todd and Marois, 2004; Xu and Chun, 2006; Cutini et al., 2011).
Fig. 2.

SPCN (contralateral minus ipsilateral) recorded at the P7/P8, time-locked to the onset of the memory array plotted as a function of race (white vs black) and number of faces (one vs two). The gray rectangle indicates the SPCN temporal window (300–900 ms).
Fig. 3.

Posterior views of isovoltage contour-maps for each experimental condition (one vs two faces and white vs black faces) generated from contralateral-minus-ipsilateral activity recorded during a 300–900 ms temporal window following the onset of the memory array.
SPCN amplitude did not differ, on average, between white and black faces, F(1, 15) = 1.06, P = 0.32, 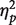 = 0.066, or between one or two faces, F < 1. The interaction between race and number of faces was, however, significant, F(1, 15) = 6.74, P < 0.05,
= 0.066, or between one or two faces, F < 1. The interaction between race and number of faces was, however, significant, F(1, 15) = 6.74, P < 0.05, 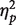 = 0.310. Planned comparisons indicated that SPCN amplitude increased significantly as the number of to-be-memorized white faces was increased, t(15) = 3.22, P < 0.01 (one white face: −1.32 μV; two white faces: −1.59 μV), whereas SPCN amplitude did not differ between one and two black faces, t(15) = −0.70, P = 0.49 (one black face: −1.40 μV; two black faces: −1.28 μV). This may be a first symptom of a processing difference between white and black faces, suggesting that only when faces were encoded with sufficient resolution (as we hypothesize for white faces), VWM resources were allocated as though separate faces representations had to be retained prior to the onset of the test array. Importantly, two white faces elicited a larger SPCN amplitude than two black faces, t(15) = 3.64, P < 0.01, likely reflecting that differences in precision of VWM representations of white vs black faces emerged more clearly either as VWM approached capacity (i.e. when two faces had to be memorized; see Sessa et al., 2011), or as the redundancy of race information increased (when the number of to-be-memorized faces was increased from one to two; e.g. Pettigrew and Tropp, 2010).
= 0.310. Planned comparisons indicated that SPCN amplitude increased significantly as the number of to-be-memorized white faces was increased, t(15) = 3.22, P < 0.01 (one white face: −1.32 μV; two white faces: −1.59 μV), whereas SPCN amplitude did not differ between one and two black faces, t(15) = −0.70, P = 0.49 (one black face: −1.40 μV; two black faces: −1.28 μV). This may be a first symptom of a processing difference between white and black faces, suggesting that only when faces were encoded with sufficient resolution (as we hypothesize for white faces), VWM resources were allocated as though separate faces representations had to be retained prior to the onset of the test array. Importantly, two white faces elicited a larger SPCN amplitude than two black faces, t(15) = 3.64, P < 0.01, likely reflecting that differences in precision of VWM representations of white vs black faces emerged more clearly either as VWM approached capacity (i.e. when two faces had to be memorized; see Sessa et al., 2011), or as the redundancy of race information increased (when the number of to-be-memorized faces was increased from one to two; e.g. Pettigrew and Tropp, 2010).
As can be seen in the Figure 4a and b, individual SPCN[Race] indices correlated with IAT scores, with high-prejudice white participants exhibiting magnified SPCN[Race] indices relative to low-prejudice white participants, independently on the number of faces displayed in the memory array (one face: r = 0.64, P < 0.01; two faces: r = 0.52, P < 0.05). Based on the known properties of SPCN, this finding suggests that the higher the prejudice held by participants, the greater the difference in precision of mental representations of white faces relative to mental representations of black faces at a stage of VWM maintenance prior to memory testing.
Fig. 4.

Correlations between individual IAT scores and SPCN[Race] indices when one face (a) or two faces (b) had to be memorized.
Behavior
We computed response accuracy and estimated the number of faces successfully retained in VWM by each participant and in each condition of the present experimental design using the formula developed by Cowan (2001):
where K is an estimate of the number of faces retained in VWM, S is the number of to-be-memorized faces displayed in the memory array (one or two faces), H is the hit rate in the change detection task and FA is the false alarm rate in the change detection task. Mean accuracy and K-values did not differ significantly for white and black faces (F’s < 1; one white face: mean accuracy 85%, K = 0.69; two white faces: mean accuracy 72%, K = 0.87; one black face: mean accuracy 86%, K = 0.71; two black faces: mean accuracy 71%, K = 0.85). Participants were less accurate when presented with two faces than one face, F(1, 15) = 144.28, P < 0.001, 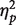 = 0.906 (one face: mean accuracy 85% vs two faces: mean accuracy 71%), and a greater amount of information was retained in VWM when two faces were displayed than when one face was displayed in the memory array, F(1, 15) = 11.48, P < 0.005,
= 0.906 (one face: mean accuracy 85% vs two faces: mean accuracy 71%), and a greater amount of information was retained in VWM when two faces were displayed than when one face was displayed in the memory array, F(1, 15) = 11.48, P < 0.005, 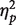 = 0.43 (one face: K = 0.70 vs two faces: K = 0.86). Differences in K between own- and other-race faces were correlated with the IAT scores when one face had to be encoded (r = 0.56, P < 0.05), indicating that higher K-values for white rather than black faces were associated with higher IAT scores.
= 0.43 (one face: K = 0.70 vs two faces: K = 0.86). Differences in K between own- and other-race faces were correlated with the IAT scores when one face had to be encoded (r = 0.56, P < 0.05), indicating that higher K-values for white rather than black faces were associated with higher IAT scores.
DISCUSSION
We used the ERP approach to assess whether implicit racial prejudice influenced the efficiency of VWM maintenance of information about the identity of own- and other-race individuals. Using a spatially cued variant of a standard change detection task, neural activity associated with VWM maintenance of own-race (white) and other-race (black) faces was quantified by monitoring SPCN responses (Jolicœur et al., 2006a, b; Luria et al., 2010; Sessa et al., 2011) elicited by a varying number of to-be-memorized faces of white and black individuals. SPCN amplitude did not differ, on average, for one own-race and one other-race face. SPCN amplitude was larger for two white faces than two black faces, suggesting that differences in precision of VWM representations of white and black faces became more evident as the VWM system approached capacity, or as the redundancy of race information increased (Pettigrew and Tropp, 2010). Crucially, individual estimates of the difference in SPCN elicited by own- vs other-race faces (i.e. SPCN[Race] indices) correlated with estimates of implicit racial prejudice (i.e. IAT scores), suggesting that high-prejudice participants encoded and retained black faces representations in VWM with a lower degree of precision than white faces. These findings support the view that implicit racial prejudice toward members of a different racial group does not simply alter people’s mood and temper overt behavior but it also affects a crucial low-order processing routine, namely, the maintenance of visual information about faces in VWM.
The behavioral results, though generally in line with the ERP results, failed to capture an analogous correlation between IAT scores and K indices of VWM load when two faces, rather than just one, were displayed in the memory array. A shared view about these discrepancies is that brain responses may be more sensitive to subtle processing differences than behavioral measures, as shown in studies where variations at the neural level did not find an analogous correspondence at the behavioral level of observation (e.g. Luck et al., 1996; Phelps et al., 2000; Heil et al., 2004). Owing to their higher sensitivity, neural measures of brain activity may convey information on possible modulations affecting the processing underlying a particular cognitive task even when such modulations do not translate into overt behavioral manifestations (see Wilkinson and Halligan, 2004). Note also that SPCN and K (or accuracy indices) are separate reflections of distinct processes enabling change detection. Differently from SPCN, the production of an overt response engages a set of processing subroutines that take place after the test array is displayed, such as an attention movement toward the changed object, a serial comparison of the changed object with the objects stored in WVM, and a final decision for or against a given response (Awh et al., 2007; Hyun et al., 2009; Dell'Acqua et al., 2010). Each of these processing subroutines may distort behavioral indices of performance in unpredictable ways.
This consideration leads us to propose an explanatory framework of the SPCN results, and of the correlation between SPCN[Race] indices and IAT scores, that is intertwined with the important question of the functional locus of racial prejudice in our model of VWM and face representation. Showing that racial prejudice exerts a modulatory effect on the amount and/or quality of information stored in VWM does not necessarily imply that the locus of the effect is confined to a stage of VWM maintenance. An obvious alternative, which we cannot exclude at present, is that racial prejudice perturbates a subset of stages involved in face encoding and/or consolidation prior to storage in VWM. Evidence hinting at the fact that this hypothesis may indeed be correct comes from ERP studies showing an effect of race on face processing occurring prior to the 250/300 ms at which SPCN usually onsets (e.g. Ito and Urland, 2003, 2005), suggesting that some sort of structural or attentional limitation in the processing of face features—for purposes related to make faces distinguishable one from another—acts at early stages along the face processing chain. Perhaps this may be due to the very simple fact a given individual has more opportunity to interact with own-race members than with other-race members, developing visual processing subroutines ‘tuned’ to specific (configuration of) features which would be just more familiar and processed more efficiently. One influential model of face perception hypothesizes in fact that facial representations occupy locations in a multidimensional psychological space by virtue of continuous variations along many learned dimensions representing peculiar physiognomic aspects useful for face discrimination (Valentine, 1991). Valentine’s model accounts for race effects in face perception and recognition assuming that other-race faces are less distinctive than own-race faces due to the lack of visual experience at processing phenotypical features of other-race faces. Spatial differences among individual other-race faces’ representations within such multidimensional psychological space would be reduced compared to those characterizing own-race faces’ representations, leading to perception and recognition deficits for other-race faces. Papesh and Goldinger (2010) have recently tested this tenet of Valentine’s (1991) model using a speeded same-different judgment task to generate empirically psychological spaces of own- and other-race faces. To note, own- and other-race faces in this study were physically identical and were varied only in skin tone, so as to equate stimuli for the level of expertise with the phenotypical features hypothesized to determine their relative distribution within a psychological space. Remarkably, the results of this study showed that other-race faces were more densely clustered than own-race faces within a given psychological space, suggesting that other-race faces were encoded and stored less efficiently than own-race faces, a result that could hardly be reconciled with Valentine’s (1991) explanation for differences in the spatial organization of own- and other-race faces based to the lack of experience at processing features from other-race faces (in the Papesh and Goldinger’s study indeed own- and other-race faces were structurally identical; see also Levin, 2000). Taken together, the findings above are strongly suggestive of a pre-VWM mechanism in the cognitive architecture underpinning face processing and recognition that reacts promptly (Ito and Urland, 2003) to the exposure of an individual’s face on the basis of a class of input features (e.g. color, as of the face skin tone) that convey information about the individual’s race. We cannot argue against or in favor of this mechanism as subjected to structural limitations affecting a stage of encoding of face features that may differ in the degree of familiarity, nevertheless Papesh and Goldinger’ (2010) findings strongly suggest that such filtering mechanism may be triggered by input features as the skin tone and would be flexible in gating the quantity of face features (or resolution in their relative spatial arrangement; Woodman et al., 2003) ultimately encoded for purposes related to building a face’s perceptual representation that can be input to VWM, and used to recognize an individual through his/her face. The proposed filtering mechanism may be permeable to the influence of automatic emotional reactions to race information processed by subcortical structures (Phelps et al., 2000), which would, however, exert a re-entrant influence on extra-striate portions of the visual cortex, including the intraparietal and intraoccipital sulci. Our results help characterize this filtering mechanisms further, by suggesting that race would not determine the band-width of information gated by the filtering mechanism in an all-or-none fashion, but flexibly, in a way proportional to the degree of racial prejudice.
There is no question that the accuracy of any decision process is ultimately constrained by the quality of information it receives. The present observation that racial prejudice influences the precision of basic visual representations of other-race faces in VWM suggests that there will be systematic deficiencies in the quality of information used by all subsequent cognitive processes. On this hypothesis, the impact of implicit racial prejudice on VWM efficiency may in turn have a ‘domino effect’ on many cognitive subsystems, including long-term memory and learning, providing the basis for a broad-spectrum of race-dependent behaviors, including mechanisms devoted to the control of such behaviors (e.g. Richeson et al., 2003). A corollary hypothesis in this architecture is that VWM faces representations characterized by a low degree of precision should be translated into poor long-term memory representations. A well-established finding in the long-term memory literature is in fact that people are notoriously more prone to make errors at recognizing faces of other racial groups than faces of their own race (i.e. other-race effect; see Meissner and Brigham, 2001, for a review). Although the influential working memory model of Baddeley and Hitch (1974) proposed that visual information may directly reach long-term memory by-passing VWM, recent evidence challenged this assumption suggesting that VWM and long-term memory represent a highly interactive functional continuum (e.g. Olson et al., 2006). This view makes it plausible that implicit racial prejudice, while affecting the precision of visual information of other-race faces retained in VWM, determines as a consequence a quality reduction of inputs to long-term memory storage processes, which in turn would bring about effects during the recovery of long-term face representations like the other-race effect.
Conflict of Interest
None declared.
REFERENCES
- Awh E, Barton B, Vogel EK. Visual working memory represents a fixed number of items regardless of complexity. Psychological Science. 2007;18:622–8. doi: 10.1111/j.1467-9280.2007.01949.x. [DOI] [PubMed] [Google Scholar]
- Baddeley AD, Hitch G. Working memory. In: Spence KW, Spence JT, editors. The Psychology of Learning and Motivation. Vol. 8. New York: Academic Press; 1974. pp. 67–89. [Google Scholar]
- Cowan N. The magical number 4 in short-term memory: A reconsideration of mental storage capacity. Behavioral and Brain Science. 2001;24:87–185. doi: 10.1017/s0140525x01003922. [DOI] [PubMed] [Google Scholar]
- Cunningham WA, Nezlek JB, Banaji MR. Implicit and explicit ethnocentrism: Revisiting the ideologies of prejudice. Personality and Social Psychology Bulletin. 2004;30:1332–46. doi: 10.1177/0146167204264654. [DOI] [PubMed] [Google Scholar]
- Cutini S, Scarpa F, Scatturin P, et al. A hemodynamic correlate of lateralized visual short-term memories. Neuropsychologia. 2011;49:1611–21. doi: 10.1016/j.neuropsychologia.2010.12.009. [DOI] [PubMed] [Google Scholar]
- Dell'Acqua R, Sessa P, Jolicœur P, Robitaille N. Spatial attention freezes during the attentional blink. Psychophysiology. 2006;43:394–400. doi: 10.1111/j.1469-8986.2006.00411.x. [DOI] [PubMed] [Google Scholar]
- Dell'Acqua R, Sessa P, Toffanin P, Luria R, Jolicœur P. Orienting attention to objects in visual short-term memory. Neuropsychologia. 2010;48:419–28. doi: 10.1016/j.neuropsychologia.2009.09.033. [DOI] [PubMed] [Google Scholar]
- Greenwald AG, McGhee DE, Schwartz JLK. Measuring individual differences in implicit cognition: The Implicit Association Test. Journal of Personality and Social Psychology. 1998;74:1464–80. doi: 10.1037//0022-3514.74.6.1464. [DOI] [PubMed] [Google Scholar]
- Heil M, Rolke B, Pecchinenda A. Automatic semantic activation is no myth: Semantic context effects on the N400 in the letter-search task in the absence of response time effects. Psychological Science. 2004;15:852–7. doi: 10.1111/j.0956-7976.2004.00766.x. [DOI] [PubMed] [Google Scholar]
- Hyun J-s, Woodman GF, Vogel EK, Hollingworth A, Luck SJ. The comparison of visual working memory representations with perceptual inputs. Journal of Experimental Psychology: Human Perception and Performance. 2009;35:1140–60. doi: 10.1037/a0015019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ito TA, Urland GR. Race and gender on the brain: Electrocortical measures of attention to race and gender of multiply categorizable individuals. Journal of Personality and Social Psychology. 2003;85:616–26. doi: 10.1037/0022-3514.85.4.616. [DOI] [PubMed] [Google Scholar]
- Ito TA, Urland GR. The influence of processing objectives on the perception of faces: an ERP study of race and gender perception. Cognitive, Affective, and Behavioral Neuroscience. 2005;5:21–36. doi: 10.3758/cabn.5.1.21. [DOI] [PubMed] [Google Scholar]
- Jolicœur P, Sessa P, Dell’Acqua R, Robitaille N. Attentional control and capture in the attentional blink paradigm: Evidence from human electrophysiology. European Journal of Cognitive Psychology. 2006a;18:560–78. [Google Scholar]
- Jolicœur P, Sessa P, Dell'Acqua R, Robitaille N. On the control of visual spatial attention: evidence from human electrophysiology. Psychological Research. 2006b;70:414–24. doi: 10.1007/s00426-005-0008-4. [DOI] [PubMed] [Google Scholar]
- Levin DT. Race as a visual feature: using visual search and perceptual discrimination tasks to understand face categories and the cross-race recognition deficit. Journal of Experimental Psychology: General. 2000;129:559–74. doi: 10.1037//0096-3445.129.4.559. [DOI] [PubMed] [Google Scholar]
- Luck SJ. Visual short-term memory. In: Luck SJ, Hollingworth A, editors. Visual memory. New York: Oxford University Press; 2008. pp. 43–86. [Google Scholar]
- Luck SJ, Vogel EK, Shapiro KL. Word meanings are accessed but cannot be reported during the attentional blink. Nature. 1996;383:616–8. doi: 10.1038/383616a0. [DOI] [PubMed] [Google Scholar]
- Luria R, Sessa P, Gotler A, Jolicœur P, Dell'Acqua R. Visual short-term memory capacity for simple and complex objects. Journal of Cognitive Neuroscience. 2010;22:496–512. doi: 10.1162/jocn.2009.21214. [DOI] [PubMed] [Google Scholar]
- Macrae CN, Bodenhausen GV. Social cognition: Thinking categorically about others. Annual Review of Psychology. 2000;51:93–120. doi: 10.1146/annurev.psych.51.1.93. [DOI] [PubMed] [Google Scholar]
- McCollough AW, Machizawa MG, Vogel EK. Electrophysiological measures of maintaining representations in visual working memory. Cortex. 2007;43:77–94. doi: 10.1016/s0010-9452(08)70447-7. [DOI] [PubMed] [Google Scholar]
- Meissner CA, Brigham JC. Thirty years of investigating the own-race bias in memory for faces: A meta-analytic review. Psychology, Public Policy, and Law. 2001;7:3–35. [Google Scholar]
- Minear M, Park DC. A lifespan database of adult facial stimuli. Behavior Research Methods, Instruments, & Computers. 2004;36:630–3. doi: 10.3758/bf03206543. [DOI] [PubMed] [Google Scholar]
- Olson IR, Moore KS, Stark M, Chatterjee A. Visual working memory is impaired when the medial temporal lobe is damaged. Journal of Cognitive Neuroscience. 2006;18:1087–97. doi: 10.1162/jocn.2006.18.7.1087. [DOI] [PubMed] [Google Scholar]
- Papesh MH, Goldinger SD. A multidimensional scaling analysis of own- and cross-race face spaces. Cognition. 2010;116:283–8. doi: 10.1016/j.cognition.2010.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pettigrew TF, Tropp L. Philadelphia: Psychology Press; 2010. When groups meet: The dynamics of intergroup contact. [Google Scholar]
- Phelps EA, O’Connor KJ, Cunningham WA, et al. Performance on indirect measures of race evaluation predicts amygdala activation. Journal of Cognitive Neuroscience. 2000;12:729–38. doi: 10.1162/089892900562552. [DOI] [PubMed] [Google Scholar]
- Richeson JA, Baird AA, Gordon HL, et al. An fMRI investigation of the impact of interracial contact on executive function. Nature Neuroscience. 2003;6:1323–8. doi: 10.1038/nn1156. [DOI] [PubMed] [Google Scholar]
- Robitaille N, Marois R, Todd J, Grimault S, Cheyne D, Jolicœur P. Distinguishing between lateralized and nonlateralized brain activity associated with visual short-term memory: fMRI, MEG, and EEG evidence from the same observers. NeuroImage. 2010;53:1334–45. doi: 10.1016/j.neuroimage.2010.07.027. [DOI] [PubMed] [Google Scholar]
- Sessa P, Luria R, Gotler A, Jolicœur P, Dell'Acqua R. Inter-hemispheric ERP asymmetries over inferior parietal cortex reveal differential visual working memory maintenance for fearful versus neutral facial identities. Psychophysiology. 2011;48:187–97. doi: 10.1111/j.1469-8986.2010.01046.x. [DOI] [PubMed] [Google Scholar]
- Todd JJ, Marois R. Capacity limit of visual short-term memory in human posterior parietal cortex. Nature. 2004;428:751–4. doi: 10.1038/nature02466. [DOI] [PubMed] [Google Scholar]
- Valentine T. A unified account of the effects of distinctiveness, inversion and race in face recognition. Quarterly Journal of Experimental Psychology. 1991;43A:161–204. doi: 10.1080/14640749108400966. [DOI] [PubMed] [Google Scholar]
- Vogel EK, Machizawa MG. Neural activity predicts individual differences in visual working memory capacity. Nature. 2004;428:748–51. doi: 10.1038/nature02447. [DOI] [PubMed] [Google Scholar]
- Vogel EK, McCollough AW, Machizawa MG. Neural measures reveal individual differences in controlling access to working memory. Nature. 2005;438:500–3. doi: 10.1038/nature04171. [DOI] [PubMed] [Google Scholar]
- Wilkinson DT, Halligan PW. The relevance of behavioural measures for functional imaging studies of cognition. Nature Reviews Neuroscience. 2004;5:67–73. doi: 10.1038/nrn1302. [DOI] [PubMed] [Google Scholar]
- Woodman GF, Vecera SP, Luck SJ. Perceptual organization influences visual working memory. Psychonomic Bulletin & Review. 2003;10:80–7. doi: 10.3758/bf03196470. [DOI] [PubMed] [Google Scholar]
- Xu Y, Chun MM. Dissociable neural mechanisms supporting visual short-term memory for objects. Nature. 2006;440:91–5. doi: 10.1038/nature04262. [DOI] [PubMed] [Google Scholar]