

Abstract
In recent years recombinant adeno-associated viral vectors (AAV) have become increasingly valuable for in vivo studies in animals, and are also currently being tested in human clinical trials. Wild-type AAV is a non-pathogenic member of the parvoviridae family and inherently replication-deficient. The broad transduction profile, low immune response as well as the strong and persistent transgene expression achieved with these vectors has made them a popular and versatile tool for in vitro and in vivo gene delivery. rAAVs can be easily and cheaply produced in the laboratory and, based on their favourable safety profile, are generally given a low safety classification. Here, we describe a method for the production and titering of chimeric rAAVs containing the capsid proteins of both AAV1 and AAV2. The use of these so-called chimeric vectors combines the benefits of both parental serotypes such as high titres stocks (AAV1) and purification by affinity chromatography (AAV2). These AAV serotypes are the best studied of all AAV serotypes, and individually have a broad infectivity pattern. The chimeric vectors described here should have the infectious properties of AAV1 and AAV2 and can thus be expected to infect a large range of tissues, including neurons, skeletal muscle, pancreas, kidney among others. The method described here uses heparin column purification, a method believed to give a higher viral titer and cleaner viral preparation than other purification methods, such as centrifugation through a caesium chloride gradient. Additionally, we describe how these vectors can be quickly and easily titered to give accurate reading of the number of infectious particles produced.
Keywords: Immunology, Issue 57, adeno-associated virus, AAV, virus titer, stereotaxic injection, viral gene transfer
Protocol
See Figure 1 for an illustration summarizing the following protocol.
Safety Note: All material that has been in contact with assembled viral particles needs to be disinfected with Virkon solution or other suitable disinfectant.
1. Preparation of plasmid DNA stocks (~ 2 days)
- The following plasmids are required 1:
- pRV1 - Containing the AAV2 Rep and Cap sequences
- pH21 - Containing the AAV1 Rep and Cap sequences
- pFdelta6 - Adenovirus-helper plasmid
- AAV plasmid containing the recombinant expression cassette flanked by AAV2 packaging signals (inverted terminal repeats, ITRs)
While pFdelta6 should be grown in Stbl2 competent cells to prevent partial deletion, pRV1 and pH21 can be grown in DH5alpha competent cells. AAV plasmids can be grown in Stbl2 cells if partial deletions occur in DH5alpha cells.
- Plasmid DNA should be of high quality and free of RNA contaminants. Plasmids can be screened for integrity using the following digests:
- pRV1 - digest with XbaI to give bands of 7.5 kb and 3.8 kb
- pH21 - digest with EcoRI to give bands of 4.5 kb, 2.8 kb and 0.2 kb
- pFdelta6 - digest with HindIII to give bands of 5.5 kb, 3 kb, 3 kb, 2.3 kb and 1.5 kb
2. Preparation of Human Embryonic Kidney 293 (Hek293) cells for transfection (2 - 3 days)
Plate two 80% confluent 150 cm2 flasks of Hek293 cells into five 15 cm diameter Nunc tissue culture dishes. Cells should be 70 - 80% confluent before transfection (approximately 48 hours after plating). Culture cells in standard Dulbecco's modified Eagle medium (DMEM) with low glucose containing 10% foetal calf serum and 100 U/ml penicillin/ 100 μg/ml streptomycin.
3 hours before transfection remove DMEM and replace with Iscove's modified Dulbecco's medium (IMDM) containing 5% foetal calf serum.
3. Transfection of viral plasmids (~1 hour for transfection, 3 days for incubation)
- Prepare the following for a single batch (5 x 15 cm tissue culture plates) of virus:
- 62.5 μg AAV plasmid
- 125 μg pFdelta6
- 31.25 μg pRV1
- 31.25 μg pH21
- 1650 μl 2.5 M CaCl2
- 12 ml dH2O
In a class 2 tissue culture hood sterile filter the transfection mixture into a 50 ml tube.
Whilst vortexing the solution, quickly add 13 ml of 2 x HEPES buffered saline (pH 7.05). Replace lid of 50 ml tube and continue to vortex for 15 seconds. Leave to stand for 1 minute 45 seconds, a fine white precipitate should form.
Gently add 5 ml of the transfection solution to each 15 cm tissue culture dish. Swirl plates to mix and return to incubator.
16 hours after transfection remove IMDM medium and replace with DMEM.
4. Lysing of cells and harvesting of rAAVs (2 hours)
72 hours after transfection, remove media from cell culture plates and discard. All waste should be treated with Virkon solution or other suitable disinfectant.
Gently wash the cells in warm 1x phosphate buffered saline (PBS; pH 7.4).
Add 25 ml warm PBS to each plate and gently remove cells with a cell scraper. Collect suspension in 50 ml tubes.
Pellet cells at 800 x g for 10 minutes.
Discard supernatant and resuspend pellet in 150 mM NaCl, 20 mM Tris pH 8.0, use 10 ml per tissue culture plate. Split into two 50 ml tubes.
Prepare a fresh solution of 10% sodium deoxycholate in dH2O. Add 1.25 ml of this to each tube for a final concentration of 0.5%. Add benzonase nuclease to a final concentration of 50 units per ml. Mix tube thoroughly.
Incubate at 37°C for 1 hour.
Remove cellular debris by centrifuging at 3000 x g for 15 mins. Transfer to fresh 50 ml tube, ensure all cell debris has been removed to prevent blocking of heparin columns (see step 5). At this stage, the samples can be stored at -20°C before continuing. We have stored samples for several weeks at -20°C without a reduction in infectivity.
5. Heparin column purification of rAAVs (2 - 3 hours)
Setup HiTrap heparin columns using a peristaltic pump so that solutions flow through the column at 1 ml per minute for steps 5.2 to 5.4. It is important to ensure no air bubbles are introduced into the heparin column.
Equilibrate the column with 10 ml 150 mM NaCl, 20 mM Tris, pH 8.0.
Apply 50 ml virus solution to column and allow to flow through.
Wash column with 20 ml 100 mM NaCl, 20 mM Tris, pH 8.0.
Using a 5 ml syringe continue to wash the column with 1 ml 200 mM NaCl, 20 mM Tris, pH 8.0, followed by 1 ml 300 mM NaCl, 20 mM Tris, pH 8.0. Discard the flow-through.
- Using 5 ml syringes and gentle pressure elute the virus from the column by applying:
- 1.5 ml 400 mM NaCl, 20 mM Tris, pH 8.0
- 3.0 ml 450 mM NaCl, 20 mM Tris, pH 8.0
- 1.5 ml 500 mM NaCl, 20 mM Tris, pH 8.0
- Collect these in a 15 ml centrifuge tube.
6. Concentration and sterile filtration of rAAVs (1 hour)
Concentrate vector using Amicon ultra-4 centrifugal filter units with a 100,000 molecular weight cutoff. Load 4 ml of column eluate into the concentrator and centrifuge at 2000 x g for 2 minutes (at room temperature). Discard flowthrough and reload concentrator with remaining virus solution and repeat centrifugation. The concentrated volume should be approximately 250 μl. If concentrated volume is significantly more than this, discard the flow through and continue to centrifuge in one minute steps until volume is approximately 250 μl.
Add 250 μl of PBS to virus for a final volume of 500 μl and remove from concentrator.
Filter vector through a 13 mm diameter 0.2 μm syringe filter. Vector should be aliquoted and stored at -80°C until required.
7. Titering of viral stocks (3 days)
This requires a promoter that is active in Hek293 cells driving a reporter gene or an immunocytochemically detectable gene product. When producing a vector that is not compatible with Hek293 cells other cell lines or primary cell cultures can be used.
Prepare 18 poly-L-lysine coated glass coverslips or 18 wells of Nunc chamber slides by seeding Hek293 cells so that they are 40 - 50% confluent. For titration of Cre-dependent rAAVs use Hek293 cells that stably express Cre recombinase 2.
Infect each well with serial dilutions of the AAV vector (Fig 2A). Use 3 wells per dilution. We routinely add virus directly to each well.
After 72 hours fix Hek293 cells by adding an equal volume of 4% paraformaldehyde (PFA) to medium (giving a final concentration of 2% PFA) for ten minutes at room temperature.
Wash cell three times in PBS, rinse in dH2O and coverslip.
Count the number of transduced cells from the three wells that have the highest dilution factor, but still contain infected cells, find the average of these and multiply by the dilution factor to give the number of infectious units per microliter (Fig 2B).
8. Anticipated Results
Hek293 cells transduced with a virus encoding enhanced green fluorescent protein (eGFP) are shown in Figure 2B. We generally achieve consistent titers of around 6 x 106 infectious particles per micro litre. We routinely use these vectors for stereotaxic injection into the adult rodent brain. This provides a powerful technique for region-specific gene expression 2. To combine this region specificity with cell-type selective gene expression we inject Cre recombinase-dependent rAAVs into target regions of Cre-transgenic mice 2. Figure 2C-E shows examples of stereotaxic injections of Cre-dependent rAAVs encoding eGFP into different brain regions of adult parvalbumin-Cre transgenic mice 3.
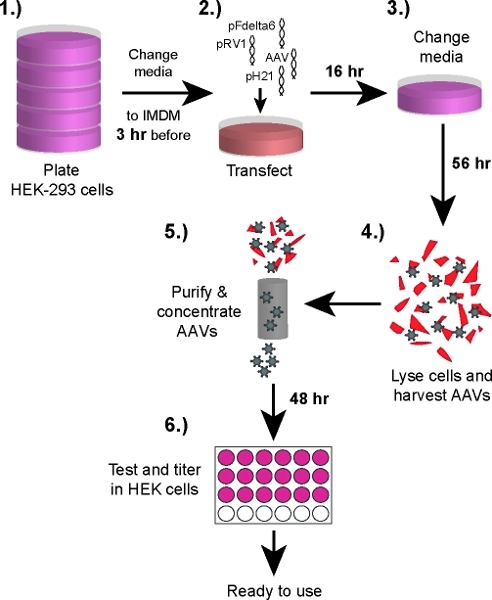 Figure 1. Schematic illustration of the protocol for rAAV production. 1.) Hek293 cells are plated and grown to 70 - 80% confluence. 2.) AAV and helper plasmids are prepared and transfected into Hek293 cells. 3.) Medium is changed back to DMEM 16 hours after transfection and Hek293 cells are cultured for another 56 hours. 4.) Hek293 cells are harvested and lysed. rAAV vectors are separated from cell debris by centrifugation. 5.) Viral particles are purified through heparin columns before concentration and sterilisation. 6.) Hek293 cells are grown on 24 well plates and infect with serial dilutions of the AAV vector to determine the number of infectious units.
Figure 1. Schematic illustration of the protocol for rAAV production. 1.) Hek293 cells are plated and grown to 70 - 80% confluence. 2.) AAV and helper plasmids are prepared and transfected into Hek293 cells. 3.) Medium is changed back to DMEM 16 hours after transfection and Hek293 cells are cultured for another 56 hours. 4.) Hek293 cells are harvested and lysed. rAAV vectors are separated from cell debris by centrifugation. 5.) Viral particles are purified through heparin columns before concentration and sterilisation. 6.) Hek293 cells are grown on 24 well plates and infect with serial dilutions of the AAV vector to determine the number of infectious units.
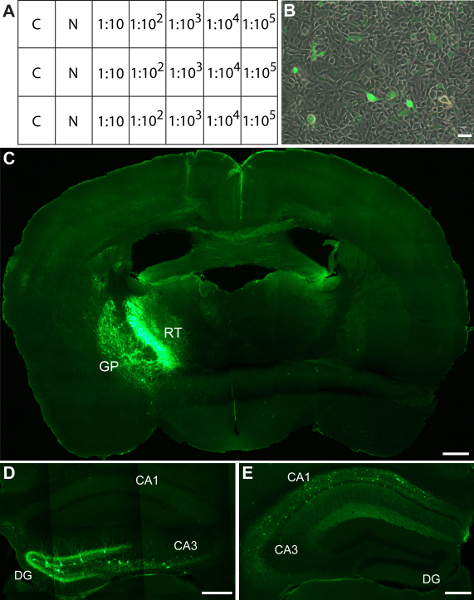 Figure 2. Titering and in vivo application of rAAV1/2. (A) Setup of Hek293 cells for measuring viral titer. Hek293 cells are transduced with serial dilutions of rAAVs. C, uninfected control; N, 1 μl of undiluted viral stock. (B) Hek293 cells infected with rAAVs expressing eGFP. (C-E) Viral eGFP expression is restricted to Cre-expressing neurons in different target regions of parvalbumin-Cre transgenic mice after stereotaxic injection of Cre-activated rAAVs. Examples images show GFP immunoreactivity in (C) the reticular thalamus and globus pallidus, (D) the dentate gyrus and (E) the CA1 region of the hippocampus. DG, dentate gyrus; RT, reticular thalamic nucleus; GP, globus pallidus. Scale bars: B, 500 μm; C, 100 μm; D, 100 μm; E, 500 μm.
Figure 2. Titering and in vivo application of rAAV1/2. (A) Setup of Hek293 cells for measuring viral titer. Hek293 cells are transduced with serial dilutions of rAAVs. C, uninfected control; N, 1 μl of undiluted viral stock. (B) Hek293 cells infected with rAAVs expressing eGFP. (C-E) Viral eGFP expression is restricted to Cre-expressing neurons in different target regions of parvalbumin-Cre transgenic mice after stereotaxic injection of Cre-activated rAAVs. Examples images show GFP immunoreactivity in (C) the reticular thalamus and globus pallidus, (D) the dentate gyrus and (E) the CA1 region of the hippocampus. DG, dentate gyrus; RT, reticular thalamic nucleus; GP, globus pallidus. Scale bars: B, 500 μm; C, 100 μm; D, 100 μm; E, 500 μm.
Discussion
rAAV vectors are becoming increasingly valuable for in vivo studies in animals. Here, we have described a simple and inexpensive protocol for producing rAAVs, which may facilitate the widespread application of this useful vector by avoiding the costly outsourcing of virus production to companies. The protocol (based on reference 1) describes the production of chimeric rAAV1/2 vectors containing capsid proteins from both parental serotypes at equal ratios 4. The purification of the rAAV vectors by binding to heparin columns relies on the expression of AAV2 capsid proteins, but can be combined with serotypes other than AAV1 outlined here. Additionally, AAV6 has been reported to bind to heparin, although with a reduced affinity, and could potentially be purified with heparin columns using this procedure 5,6. It should be noted however, that other serotypes will require different concentrations of NaCl for heparin column elution 5,7.
Packaging rAAV genomes was shown to be optimal between 4.1 and 4.9 kb with a sharp reduction in packaging efficiency up to 5.2 kb 8. This packaging limit can be considered the largest disadvantage of the rAAV system, since cell type-specific regulation of transgene expression is usually achieved by large cis acting elements which can not be accommodated within the small AAV particles. To overcome low titer batches, such as those resulting from attempting to package genes that are close to the AAV packing limit we are combining separate viral batches concentrated to 250 μl into one 500 μl batch, rather than adding PBS as outlined in step 6.3. Reliable cell-type specific transduction can be achieved by combining Cre-driver mice and conditional rAAV cassettes (see below) to avoid stretching the packaging limit.
Of note, while this protocol attempts to provide easy-to-follow instructions on the production of high titer rAAV, it requires the presence of AAV2 capsid protein moieties for purification by heparin affinity chromatography. Serotypes other than AAV2 must be purified using alternate procedures 9. One advantage of heparin column purification is it is believed to produce AAV vectors that have greater infectivity rate than those produced using a caesium chloride gradient 10. However, there are also some drawbacks to this method. For example, other proteins that bind heparin may also be present in the purified viral stock. While the tropism of heparin-purified vectors can be influenced by the vector titer10, our approach to specifically target either excitatory20 or inhibitory neurons (Fig. 2) employs Cre-induced activation of AAV-mediated transgene expression and is titre-independent. An alternative to heparin column purification is the use of an iodixanol density gradient, which produces a higher viral titer and purer preparation than the use of a caesium chloride gradient 10. This method can also be combined with heparin column purification to produce a vector that is greater than 99% pure 11. Therefore, careful consideration needs to be taken by individual groups as to which viral purification method is best for their particular downstream application.
The most common problem associated with this protocol is low viral titer. In our experience this can usually be traced to low transfection efficiency, or the elution of virus from the heparin columns. All transfection materials should be at room temperature before transfection, and test transfections should be performed to find the optimal conditions in each laboratory. Other methods of transfection, such as lipofectamine, are highly efficient, but can be prohibitively expensive. Additionally, the concentration and pH of the heparin column elution solutions is critical for complete elution of virions from heparin columns without damage. Another important point is that the packaging cells must adhere well to the surface of the tissue culture dishes. If the cells detach at some stage during the procedure it is advised to terminate the experiment and defrost a new vial of cells.
Spatio-temporal control of transgene expression in rodents is easily achieved by accurate anatomical targeting and the developmental stage of the animal. Efficient AAV-mediated gene transfer has been shown in utero 12,13. In the adult brain, the area infected with rAAV particles after stereotaxic injection can be adjusted from very small targeted regions, to much larger areas not only by changes in the viral titer but also by altering the injection parameters. These include the volume and speed of injections and the inclusion of the polyol mannitol with the virus suspension 14. Mannitol can also be used to enhance infection of a variety of tissues after systemic rAAV injection 15.
The packaging capacity of rAAV (approximately 4.7 kb) is probably the most limiting factor in terms of the genes that can be expressed from these viral vectors. However, recent studies have shown that this can be partially overcome by splitting larger genes or expression cassettes into separate rAAV vectors and introducing a bridging sequence, initiating gene expression when virions infect the same cell 16. Expression levels of genes carried by rAAV vectors can also be greatly enhanced by using self-complimentary rAAV vectors 17. Stereotaxic injection of rAAV vectors provides a rapid, inexpensive and powerful method for inducing gene expression in a region-specific manner. In combination with 2nd generation RNA interference strategies 18 rAAVs can also be used for region-specific gene-knock down. rAAVs can be combined with Cre-transgenic mice or cell-type specific promoters to achieve circuit- and cell-type-specific gene expression, permitting genetic manipulations with high spatial resolution 2,19-21, while fitting rAAVs with e.g. the tet system adds temporal control over virus-mediated gene expression 22,23.These examples illustrate the enormous combinatorial potential of these vectors and predict a rapid increase in rAAV-based studies over the years to come.
Disclosures
No conflicts of interest declared.
References
- Klugmann M. AAV-mediated hippocampal expression of short and long Homer 1 proteins differentially affect cognition and seizure activity in adult rats. Molecular and Cellular Neurosciences. 2005;28:347–360. doi: 10.1016/j.mcn.2004.10.002. [DOI] [PubMed] [Google Scholar]
- Murray AJ. Parvalbumin-positive CA1 interneurons are required for spatial working but not for reference memory. Nature Neuroscience. 2011;14:297–299. doi: 10.1038/nn.2751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hippenmeyer S. A developmental switch in the response of DRG neurons to ETS transcription factor signaling. PLoS Biology. 2005;3:e159–e159. doi: 10.1371/journal.pbio.0030159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hauck B. Generation and characterization of chimeric recombinant AAV vectors. Molecular Therapy. 2003;7:419–425. doi: 10.1016/s1525-0016(03)00012-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halbert CL, Allen JM, Miller A. Adeno-associated virus type 6 (AAV6) vectors mediate efficient transduction of airway epithelial cells in mouse lungs compared to that of AAV2 vectors. Journal of Virology. 2001;75:6615–6615. doi: 10.1128/JVI.75.14.6615-6624.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blankinship MJ. Efficient transduction of skeletal muscle using vectors based on adeno-associated virus serotype 6. Molecular Therapy. 2004;10:671–678. doi: 10.1016/j.ymthe.2004.07.016. [DOI] [PubMed] [Google Scholar]
- Wu Z. Single amino acid changes can influence titer, heparin binding, and tissue tropism in different adeno-associated virus serotypes. Journal of Virology. 2006;80:11393–11397. doi: 10.1128/JVI.01288-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dong JY, Fan PD, Frizzell RA. Quantitative analysis of the packaging capacity of recombinant adeno-associated virus. Human gene therapy. 1996;7:2101–2112. doi: 10.1089/hum.1996.7.17-2101. [DOI] [PubMed] [Google Scholar]
- Zolotukhin S. Production and purification of serotype 1, 2, and 5 recombinant adeno-associated viral vectors. Methods. 2002;1:158–167. doi: 10.1016/s1046-2023(02)00220-7. [DOI] [PubMed] [Google Scholar]
- Burova E, Loffe E. Chromatographic purification of recombinant adenoviral and adeno-associated viral vectors: methods and implications. Gene Therapy. 2005;12:5–17. doi: 10.1038/sj.gt.3302611. [DOI] [PubMed] [Google Scholar]
- Zolotukhin S. Recombinant adeno-associated virus purification using novel methods improves infectious titer and yield. Gene Therapy. 1999;6:973–985. doi: 10.1038/sj.gt.3300938. [DOI] [PubMed] [Google Scholar]
- Bilbao R. Patterns of gene expression from in utero delivery of adenoviral-associated vector serotype 1. Human Gene Therapy. 2005;16:678–684. doi: 10.1089/hum.2005.16.678. [DOI] [PubMed] [Google Scholar]
- Pilpel N. reproducible transduction of select forebrain regions by targeted recombinant virus injection into the neonatal mouse brain. Journal of Neuroscience Methods. 2009;182:55–63. doi: 10.1016/j.jneumeth.2009.05.020. [DOI] [PubMed] [Google Scholar]
- Mastakov MY. Combined injection of rAAV with mannitol enhances gene expression in the rat brain. Molecular Therapy. 2001;3:225–232. doi: 10.1006/mthe.2001.0246. [DOI] [PubMed] [Google Scholar]
- Fu H. Self-complementary adeno-associated virus serotype 2 vector: global distribution and broad dispersion of AAV-mediated transgene expression in mouse brain. Molecular Therapy. 2003;8:911–917. doi: 10.1016/j.ymthe.2003.08.021. [DOI] [PubMed] [Google Scholar]
- Ghosh A, Yue Y, Duan D. Efficient Transgene Reconstitution with Hybrid Dual AAV Vectors Carrying the Minimized Bridging Sequences. Human Gene Therapy. 2011;22:77–83. doi: 10.1089/hum.2010.122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCarty DM. Self-complementary AAV vectors; advances and applications. Molecular Therapy. 2008;16:1648–1656. doi: 10.1038/mt.2008.171. [DOI] [PubMed] [Google Scholar]
- Georgiadis A. AAV-mediated knockdown of peripherin-2 in vivo using miRNA-based hairpins. Gene Therapy. 2010;17:486–493. doi: 10.1038/gt.2009.162. [DOI] [PubMed] [Google Scholar]
- Atasoy D. A FLEX switch targets Channelrhodopsin-2 to multiple cell types for imaging and long-range circuit mapping. The Journal of Neuroscience. 2008;28:7025–7030. doi: 10.1523/JNEUROSCI.1954-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guggenhuber S. AAV Vector-Mediated Overexpression of CB1 Cannabinoid Receptor in Pyramidal Neurons of the Hippocampus Protects against Seizure-Induced Excitoxicity. PLoS One. 2010;5:e15707–e15707. doi: 10.1371/journal.pone.0015707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lawlor PA. Efficient gene delivery and selective transduction of glial cells in the mammalian brain by AAV serotypes isolated from nonhuman primates. Molecular therapy : the journal of the American Society of Gene Therapy. 2009;17:1692–1702. doi: 10.1038/mt.2009.170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chtarto A. Tetracycline-inducible transgene expression mediated by a single AAV vector. Gene Therapy. 2003;10:84–94. doi: 10.1038/sj.gt.3301838. [DOI] [PubMed] [Google Scholar]
- Han Y. Lack of humoral immune response to the tetracycline (Tet) activator in rats injected intracranially with Tet-off rAAV vectors. Gene Therapy. 2010;17:616–625. doi: 10.1038/gt.2010.6. [DOI] [PMC free article] [PubMed] [Google Scholar]