

Abstract
Antibody-driven phagocytosis is induced via the engagement of Fc receptors on professional phagocytes, and can contribute to both clearance as well as pathology of disease. While the properties of the variable domains of antibodies have long been considered critical to in vivo function, the ability of antibodies to recruit innate immune cells via their Fc domains has become increasingly appreciated as a major factor in their efficacy, both in the setting of recombinant monoclonal antibody therapy, as well as in the course of natural infection or vaccination1-3.
Importantly, despite its nomenclature as a constant domain, the antibody Fc domain does not have constant function, and is strongly modulated by IgG subclass (IgG1-4) and glycosylation at Asparagine 2974-6. Thus, this method to study functional differences of antigen-specific antibodies in clinical samples will facilitate correlation of the phagocytic potential of antibodies to disease state, susceptibility to infection, progression, or clinical outcome.
Furthermore, this effector function is particularly important in light of the documented ability of antibodies to enhance infection by providing pathogens access into host cells via Fc receptor-driven phagocytosis7. Additionally, there is some evidence that phagocytic uptake of immune complexes can impact the Th1/Th2 polarization of the immune response8.
Here, we describe an assay designed to detect differences in antibody-induced phagocytosis, which may be caused by differential IgG subclass, glycan structure at Asn297, as well as the ability to form immune complexes of antigen-specific antibodies in a high-throughput fashion. To this end, 1 μm fluorescent beads are coated with antigen, then incubated with clinical antibody samples, generating fluorescent antigen specific immune complexes. These antibody-opsonized beads are then incubated with a monocytic cell line expressing multiple FcγRs, including both inhibitory and activating. Assay output can include phagocytic activity, cytokine secretion, and patterns of FcγRs usage, and are determined in a standardized manner, making this a highly useful system for parsing differences in this antibody-dependent effector function in both infection and vaccine-mediated protection9.
Protocol
1. Culture Phagocytic Cells
Culture THP-1 cells10 in RPMI 1640 supplemented with 10% fetal bovine serum as a stationary suspension in T flasks. Cell density should be maintained below 0.5x106/mL in order to maintain consistent levels of FcγR expression and assay performance.
2. Prepare Biotinylated Antigen
Calculate the amount of sulfo-NHS LC biotin reagent kit necessary to biotinylate the target antigen of interest according to the manufacturer’s instructions.
Dissolve the biotinylation reagent in water and immediately add the calculated amount to antigen in a buffer that does not contain primary amines. Allow the reaction to proceed for 1 hour at room temperature, mixing occasionally.
Remove excess, unconjugated biotin by buffer exchange in an Amicon centrifugal concentration unit of an appropriate molecular weight cutoff to retain the target antigen. Add sample to the top chamber of the concentration unit and add PBS to bring the total volume up to 15 mL fill line. Centrifuge at 4000 x g to bring volume down to approximately 1.5 mls. Repeating this process twice will remove 99% of the free biotin to ensure maximal coating of antigen to beads.
3. Prepare Antigen Saturated Beads
Wash 100 μl of 1 mm fluorescent neutravidin beads twice in 1 ml 0.1% PBS-BSA after spinning down in a microcentrifuge at high speed. Resuspend washed beads in 100 μl PBS-BSA, and aliquot into 10 tubes.
To determine antigen coating conditions which saturate the beads, combine 10 ul of the washed bead suspension with varying concentrations of biotinylated antigen in two-fold steps. Incubate overnight at 4°C in a microcentrifuge tube on a rotator.
NOTE: Saturation of beads must be determined experimentally. This can be accomplished by identifying bead-coating conditions that yield maximal phagocytosis when beads are subsequently opsonized with a control monoclonal antibody.
Remove unbound antigen by washing with 1 mL PBS-BSA and centrifuge at high speed until beads are pelleted. Remove PBS-BSA and repeat.
Resuspend antigen-coated beads in a final volume of 1 ml in PBS-BSA. Beads can be stored for up to a week at 4°C prior to use.
4. Prepare Antibody Samples
Clinical antibody samples can be purified from plasma using a Melon gel IgG purification kit according to manufacturers instructions. Purified IgG can be stored at 4°C until ready for use.
NOTE: Proper precautions regarding handling human samples must always been taken.
Determine the concentration of purified antibodies by absorbance at A280, and dilute samples to 1 mg/ml in PBS.
Prepare positive and negative monoclonal control antibodies by dilution to 1 mg/ml in PBS.
5. Plating the Experiment
Resuspend the washed, antigen saturated bead solution prepared above by vortexing, and transfer 10 μl into each well of a round-bottom 96 well plate. Care must be taken to continually agitate the bead suspension to ensure equal numbers of fluorescent beads are added to each well.
Add varying concentrations of the antibodies of interest to each well, creating a dose-response curve for each antibody. Optimal concentrations will differ between samples depending on the titer of antibodies present, but a range of 0.01- 100 μg/ml final concentration will provide good coverage and allow identification of the concentration range of interest. Ensure antibodies are added in volumes no larger than 20 μl.
Incubate beads and antibody samples for 2 hours at 37°C to allow the antibodies to opsonize the beads.
Prepare a suspension of THP-1 cells at 2.5 x 105 cells/ml, and add 200 μl of this suspension to each well, for a total of 5 x 104 THP-1 cells in each well.
Incubate overnight at 37°C, 5% CO2, in a stationary incubator, allowing cells and beads to pellet via gravity.
NOTE: Signal to noise may be improved by determining the optimal bead:antibody:THP-1 cell ratios for a given antigen and antibody source.
6. Flow Cytometric Analysis
Remove 100 μl of supernatant from each well being careful not to disturb the cell pellet. Supernatant can be saved if cytokine secretion determinations or other analyses are desired.
For fixation, add 100 μl of 4% paraformaldehyde to each well and pipet to resuspend and mix cells.
High throughput flow cytometric analysis can be performed using a BD LSR II equipped with an HTS plate reader.
Program software to mix each well three times (100 μl mix volume) and analyze 30 μl, or at least 2000 cell events, of each sample.
Data collected can be analyzed in FlowJo or equivalent software. Useful metrics include the percent of bead+, or fluorescent cells, which provides a measure of the number of phagocytic cells present, as well as the fluorescence intensity of the phagocytic cells, which provides a measure of the number of phagocytosed beads. Multiplying these values generates an integrated mean fluorescent intensity, or iMFI.
The average number of beads phagocytosed by each phagocytic cell can be calculated by dividing the iMFI by the mean fluorescence of 1 bead.
7. Representative Results
There should be clear differentiation of antibody samples from affected and unaffected subjects. Figure 1A presents the FACS histograms of an antibody sample from an HIV negative (black trace) and an HIV positive (gray trace) subject, and demonstrates the increased phagocytosis driven by the presence of antigen-specific antibodies.
Optimal sensitivity of the assay is dependent on saturation of the beads with biotinylated antigen. Figure 1B presents the phagocytosis observed when beads coated with differing amounts of antigen were opsonized with 3 control monoclonal antibodies (including a non-binding antibody, triangles), and establishes 2μg antigen/μl beads as a saturating concentration for this antigen.
A dose-response curve is presented in Figure 2, and demonstrates the differential capacity of subject antibody samples to induce phagocytosis over an antibody concentration range of 0.05-5 μg/ml. This differential phagocytosis may be driven by either differences in titer or Fc domain properties such as IgG subclass and glycosylation state.
When THP-1 cells are imaged by fluorescent microscopy, there is clear evidence of bead phagocytosis. Figure 3 presents two 63x still images of THP-1 cells after incubation with antibody opsonized (green) and non-opsonized (red) beads, demonstrating the lack of phagocytic uptake in the absence of antibody. When time lapse microscopy is performed, the antibody-specific phagocytic uptake of fluorescent beads is even more striking (Movie 1, 20x magnification).
Previous work has confirmed internalization of the beads associated with the cells, and experiments with primary monocytes have agreed well with phagocytic scores (iMFI values) in this high throughput assay (data not shown).
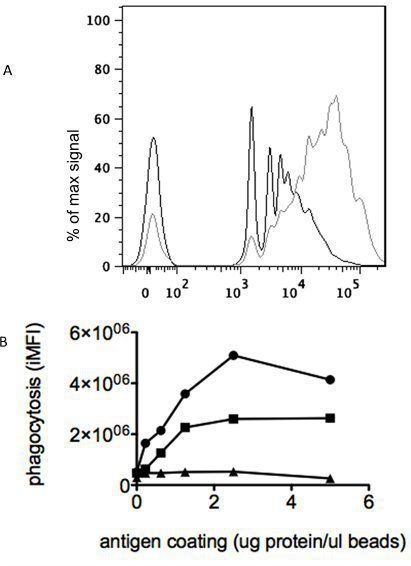 Figure 1.
Assay Quality Control. 1A, Flow cytometry histograms of phagocytosis for an antibody sample from an HIV negative subject (black trace) and an HIV positive subject (gray trace). 1B: Experimental determination of the optimal bead coating conditions for a sample antigen. Antigen-specific monoclonal antibodies (circle, square) demonstrate maximal phagocytosis of beads coated with >2 μg antigen/μl of beads, while a control antibody (triangle) demonstrates no phagocytic activity.
Figure 1.
Assay Quality Control. 1A, Flow cytometry histograms of phagocytosis for an antibody sample from an HIV negative subject (black trace) and an HIV positive subject (gray trace). 1B: Experimental determination of the optimal bead coating conditions for a sample antigen. Antigen-specific monoclonal antibodies (circle, square) demonstrate maximal phagocytosis of beads coated with >2 μg antigen/μl of beads, while a control antibody (triangle) demonstrates no phagocytic activity.
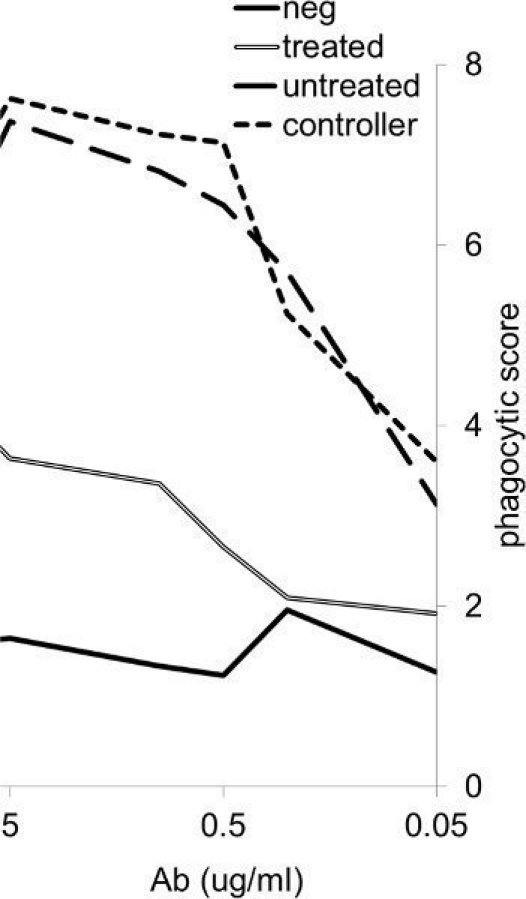 Figure 2.
Phagocytosis Dose-Response Curve. Clinical antibody samples from HIV positive (treated, untreated, and exhibiting control of viral replication in the absence of anti-retroviral therapy) and HIV negative subjects drive phagocytosis of gp120 (HIV envelope) coated beads differentially.
Figure 2.
Phagocytosis Dose-Response Curve. Clinical antibody samples from HIV positive (treated, untreated, and exhibiting control of viral replication in the absence of anti-retroviral therapy) and HIV negative subjects drive phagocytosis of gp120 (HIV envelope) coated beads differentially.
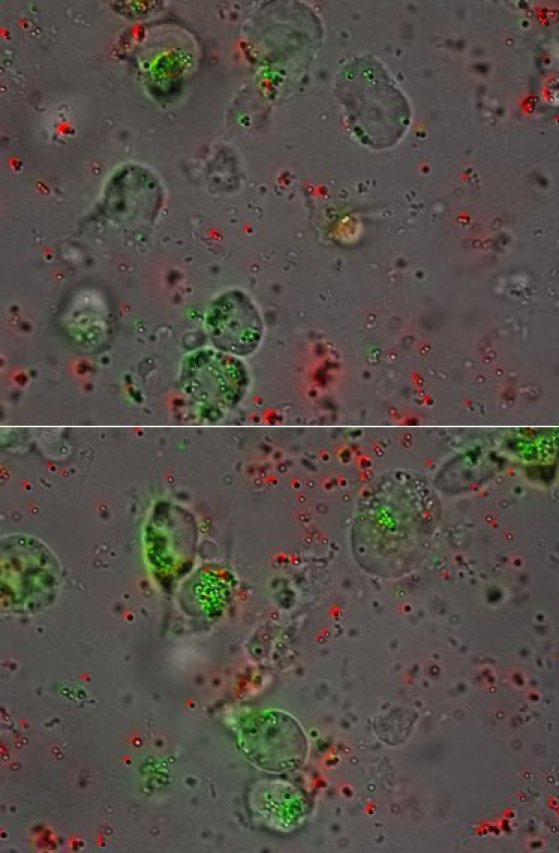 Figure 3.
Efficient Internalization. Microscopy confirms the internalization of antibody-opsonized beads (green), while non-opsonized beads remain in solution (63x magnification).
Figure 3.
Efficient Internalization. Microscopy confirms the internalization of antibody-opsonized beads (green), while non-opsonized beads remain in solution (63x magnification).
Movie 1. Time-lapse microscopy of antibody-driven phagocytosis was performed over the course of 14 hours and allows visualization of the phagocytic activity of the THP-1 cells utilized in the high-throughput assay. Green fluorescent beads are antibody opsonized, red fluorescent beads provide a negative control. Click here to watch the movie.
Discussion
The assay described here allows high-throughput analysis of the phagocytic activity of clinical antibody samples by utilizing a monocytic cell line long utilized to study phagocytic processes10 and plate based automated flow cytometry. This assay is advantageous over others in its ability to precisely characterize antigen-specific antibody subsets, allowing not only for study of differences in phagocytosis driven by disease-specific antibodies from different subjects, but also study of multiple antigen specificities within the same subject. Because antibody recruitment of innate effector cells, including monocytes and other antigen presenting cells strongly impacts disease outcome11-13, and is biologically variable depending antibody geometry, IgG subclass, and glycosylation at Asparagine297 of the Fc domain14-16, this method provides for evaluation of a critical antibody mechanism of action. Furthermore, because antibody-mediated phagocytosis is exploited by some pathogens during the course of infection7,17,18, this assay holds promise in evaluation of the process of antibody-dependent enhancement, and highlights the dual nature of phagocytosis as a potentially protective, as well as potentially detrimental antibody activity.
The assay described can be utilized to analyze antibodies from clinical samples of patient populations, but also of vaccinees, and may be adapted to utilize serum rather than purified IgG. Additionally, cytokine secretion in response to phagocytosis can be analysed from the culture supernatant, and the influence of specific FcR can be determined by using FcR-blocking antibodies, allowing not only differences in phagocytic potency to be determined, but downstream signaling events, with insight into mechanism, and may help to resolve and refine understanding of this immune mechanism.
Disclosures
No conflicts of interest declared.
Acknowledgments
The authors wish to acknowledge funding from NIH 3R01AI080289-02S1, and a Harvard University Center for AIDS Research Scholar Fellowship under NIH/NIAID 2P30AI060354-07.
References
- Cartron G. Therapeutic activity of humanized anti-CD20 monoclonal antibody and polymorphism in IgG Fc receptor FcgammaRIIIa gene. Blood. 2002;99:754–758. doi: 10.1182/blood.v99.3.754. [DOI] [PubMed] [Google Scholar]
- Ahmad R. Evidence for a correlation between antibody-dependent cellular cytotoxicity-mediating anti-HIV-1 antibodies and prognostic predictors of HIV infection. J. Clin. Immunol. 2001;21:227–233. doi: 10.1023/a:1011087132180. [DOI] [PubMed] [Google Scholar]
- Hashimoto G, Wright PF, Karzon DT. Antibody-dependent cell-mediated cytotoxicity against influenza virus-infected cells. J. Infect Dis. 1983;148:785–794. doi: 10.1093/infdis/148.5.785. [DOI] [PubMed] [Google Scholar]
- Anthony RM, Nimmerjahn F. Th . Curr. Opin. Organ Transplant. 2010 doi: 10.1097/MOT.0b013e328342538f. [DOI] [PubMed] [Google Scholar]
- Takai T. Fc receptors and their role in immune regulation and autoimmunity. J. Clin. Immunol. 2005;25:1–18. doi: 10.1007/s10875-005-0353-8. [DOI] [PubMed] [Google Scholar]
- Jefferis R. Antibody therapeutics: isotype and glycoform selection. Expert. Opin. Biol. Ther. 2007;7:1401–1413. doi: 10.1517/14712598.7.9.1401. [DOI] [PubMed] [Google Scholar]
- Halstead SB, O'Rourke EJ. Dengue viruses and mononuclear phagocytes. I. Infection enhancement by non-neutralizing antibody. J. Exp. Med. 1977;146:201–217. doi: 10.1084/jem.146.1.201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subramaniam KS. The absence of serum IgM enhances the susceptibility of mice to pulmonary challenge with Cryptococcus neoformans. J. Immunol. 2010;184:5755–5767. doi: 10.4049/jimmunol.0901638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ackerman ME. A robust, high-throughput assay to determine the phagocytic activity of clinical antibody samples. J. Immunol. Methods. 2011;366:8–19. doi: 10.1016/j.jim.2010.12.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsuchiya S. Establishment and characterization of a human acute monocytic leukemia cell line (THP-1. Int. J. Cancer. 1980;26:171–176. doi: 10.1002/ijc.2910260208. [DOI] [PubMed] [Google Scholar]
- Ward ES, Ghetie V. The effector functions of immunoglobulins: implications for therapy. Ther. Immunol. 1995;2:77–94. [PubMed] [Google Scholar]
- Winiarska M, Glodkowska-Mrowka E, Bil J, Golab J. Molecular mechanisms of the antitumor effects of anti-CD20 antibodies. Front. Biosci. 2011;16:277–306. doi: 10.2741/3688. [DOI] [PubMed] [Google Scholar]
- Takai T. Fc receptors: their diverse functions in immunity and immune disorders. Springer Semin. Immunopathol. 2006;28:303–304. doi: 10.1007/s00281-006-0055-y. [DOI] [PubMed] [Google Scholar]
- Yamaguchi Y. Glycoform-dependent conformational alteration of the Fc region of human immunoglobulin G1 as revealed by NMR spectroscopy. Biochim. Biophys. Acta. 2006;1760:693–700. doi: 10.1016/j.bbagen.2005.10.002. [DOI] [PubMed] [Google Scholar]
- Anthony RM, Ravetch JV. A novel role for the IgG Fc glycan: the anti-inflammatory activity of sialylated IgG. Fcs. J. Clin. Immunol. 2010;30(Suppl 1):S9–S14. doi: 10.1007/s10875-010-9405-6. [DOI] [PubMed] [Google Scholar]
- Nimmerjahn F, Anthony RM, Ravetch JV. Agalactosylated IgG antibodies depend on cellular Fc receptors for in vivo activity. Proc. Natl. Acad. Sci. U. S. A. 2007;104:8433–8437. doi: 10.1073/pnas.0702936104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marchette NJ. Effect of immune status on dengue 2 virus replication in cultured leukocytes from infants and children. Infect. Immun. 1979;24:47–50. doi: 10.1128/iai.24.1.47-50.1979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fust G. Neutralizing and enhancing antibodies measured in complement-restored serum samples from HIV-1-infected individuals correlate with immunosuppression and disease. AIDS. 1994;8:603–609. doi: 10.1097/00002030-199405000-00005. [DOI] [PubMed] [Google Scholar]