

Abstract
The current rapid event-related fMRI study used optional parasitic gap constructions, such as [Which paper] did the tired student submit [gap--] after reviewing [p-gap--/it]?, to test 3 potential roles for Broca’s area in sentence processing. These 3 functional options are: I. any intra-sentential Dependency relation activates Broca’s area. II. this region specifically processes syntactic Displacement or movement. III. Broca’s area handles any dependency relation, as long as it is predictable at an early stage of processing. Broca’s area was only activated by the contrast that tested predictability within BA 45, as determined by its overlap with cytoarchitectonic probability maps. These results imply that an alternative or modified functional account of Broca’s area, from those presently available, is required. Constraints on either a displacement account to movements that are not parasitic or a Working Memory one to predicted dependencies that cross verbal arguments or noun phrases would achieve the necessary consistency. Further, the results from the minimal contrasts investigating displacement and dependency have implications to potential language regions outside of Broca’s area. The minimal contrast investigating displacement activated the left anterior Middle Temporal Gyrus, which has more recently been claimed to play a role in syntactic operations.
Keywords: fMRI, Syntax, Parasitic Gap, Working Memory, Predictability
1 Introduction
This paper addresses several central issues regarding the human sentence processing mechanisms, and their neural underpinnings. It explores the nature of mechanisms that compute filler-gap dependencies in connection with Broca’s area and potentially other language regions. It does so by an fMRI investigation of parasitic-gap constructions. We report an attempt to distinguish between 3 possible roles of this brain region in sentence processing: I. a Working Memory (WM) approach, which contends that any intra-sentential Dependency relation activates Broca’s area. II. a syntax specific approach, according to which this region is involved in the processing of syntactic Displacement or movement. III. a Predictability approach, which holds that Broca’s area handles any dependency relation, as long as it is syntactically (not statistically) predictable at an early stage of processing. The study therefore touches on critical questions in psycho- and neuro-linguistics.
1.1 Background
Neurolinguistic accounts of sentence comprehension have long been a source of debate. The current picture identifies Broca’s area as critical for the comprehension of sentences in which an argument is displaced, or moved (1), as Broca’s aphasic patients with focal lesions in and around Broca’s area (i.e., Brodmann’s Areas 44, 45, insula, and surrounding white matter) perform at chance levels on comprehension tests of these, but not on sentences that do not contain displaced constituents (2) (e.g., Ansell & Flowers, 1982; Caplan & Futter, 1986; Caramazza & Zurif, 1976; Drai & Grodzinsky, 2006a; Grodzinsky, 1989; Hickok & Avrutin, 1995; Shapiro & Levine, 1990; Sherman & Schweickert, 1989):
1. a. [fillerWhich teacher] did the student push [gap--]? Chance b. [filler The teacher] was pushed [gap--] by the student Chance c. [filler The teacher] that the student pushed [gap--] was strong Chance 2. a. Which student pushed the teacher? Above chance1 b. The student pushed the teacher Above chance c. The teacher who pushed the student was strong Above chance
Note that we focus on A’-dependencies2. In psycholinguistic terms, these are filler-gap dependencies in which the filler is identified as such early on. We steer clear from more complicated cases, as they appear to have little direct bearing on the question that concerns is.
Related real-time studies of filler-gap relations in aphasia, mostly carried out through Cross Modal Lexical Priming (e.g., Shapiro, Gordon, & Killackey, 1993; Swinney & Zurif, 1995; Zurif, Swinney, Prather, Solomon, & Bushell, 1993) have likewise documented highly selective deficits that pertain to displacement.
While the distinction between (1) and (2) is expressible in syntactic terms, it can also be viewed as a deficit to processing mechanisms of various sorts. Indeed, accounts of the receptive deficit in Broca’s aphasia range from those postulating a specific movement-related deficit, such as the Trace-Deletion Hypothesis (TDH) and related views (e.g., Friedmann, 2008; Friedmann & Shapiro, 2003; Grillo, 2009; Grodzinsky, 1984, 2000; Mauner, Fromkin, & Cornell, 1993), to accounts based on the ease of processing canonically sequenced sentences, and the difficulty of processing deviations from such sequences (e.g., Caplan & Futter, 1986; Hagiwara, 1993), to WM accounts (e.g., Caplan & Waters, 1999), and timing-based accounts, according to which the selective sentence-reception deficit in Broca’s aphasia stems from overall slowed processing (e.g., Avrutin, 2006; Zurif, 2003). Importantly, it has been shown that despite considerable individual variability, structural generalizations can be maintained when appropriate statistical methods are used (cf. Drai & Grodzinsky, 2006a, b, passim). In addition, finer distinctions may lead to revisions of these generalization (see Discussion page 33).
While accounts of the processing deficit have been relatively precise, anatomical precision has been lacking. Focal brain lesions (mostly subsequent to stroke in this case), being accidents of nature, do not always align with anatomical borders. The advent of neuroimaging techniques has shed new light on this matter, as it provides new angles on the relevant processes, as well as greater anatomical precision. In many of the fMRI studies of sentence processing (especially the more recent ones, e.g., Santi & Grodzinsky 2007a,b, 2010, Makuuchi et al., 2009, as well as the present one), Broca’s area and its parts are localized through the cytoarchitectonic probability maps that have come out of the Jülich brain mapping project (cf. Amunts et a., 1999, passim), which allows localization in Brodmann areas (and some of their newly discovered subparts, cf. Amunts et al., 2009) while taking individual variation into account. This method is widely recognized as the best anatomical localization tool in existence (cf. Eickhoff et al., 2007).
fMRI studies currently show that healthy participants indeed demonstrate greater activation within Broca’s area for sentences with displacement than sentences without (Ben-Shachar, Hendler, Kahn, Ben-Bashat, & Grodzinsky, 2003; Ben-Shachar, Palti, & Grodzinsky, 2004; Caplan et al., 2002; Friederici, Fiebach, Schlesewsky, Bornkessel, & Von Cramon, 2006; Grodzinsky, 2000; Just, Carpenter, Keller, Eddy, & Thulborn, 1996; Santi & Grodzinsky, 2007a,b, 2010; Stromswold, Caplan, Alpert, & Rauch, 1996). For example, experiments in German have demonstrated that activation in Broca’s area (identified through visual inspection of the MR images in this case) is incrementally intensified as the number of displacements in a sentence is increased (Friederici, Fiebach, Schlesewsky, Bornkessel, & Von Cramon, 2006):
3. a. Heute hat der Opa dem Jungen den Lutscher geschenkt
Today has the grandpa the boy the candy given
b. Heute hat [filler1 dem Jungen] der Opa [gap1--] den Lutscher geschenkt
c. Heute hat [filler1 dem Jungen] [filler2 den Lutscher] der Opa [gap1--] [gap2--] geschenkt
Sentences without displacement (3a) induced a weaker signal in Broca’s area than (3b) which contains 1 displacement, and which in turn was lower in intensity than (3c) which contains 2 displacements, indicating that degree of displacement is related to signal intensity in Broca’s area. Thus the results that pertain to displacement are relatively stable across experiments, laboratories, and languages, and still, the particular process(es) that Broca’s area supports remain a matter of debate. Similar to the aphasia literature, views of the role of this area in the neuroimaging literature range from specific syntactic displacement operations (in line with the TDH), to WM, and Syntactic Complexity (Caplan & Waters, 1999; Friederici et al., 2006; Grodzinsky, 1984, 2000; Just, Carpenter, Keller, Eddy, & Thulborn, 1996; Stromswold, Caplan, Alpert, & Rauch, 1996).
Broca’s area seems to play a crucial role in the processing of intra-sentential dependency relations: when this brain regions is lesioned, chance comprehension of these relations is documented in aphasia (1); in health, such relations activate it, relative to control conditions (3b–c). Below, we focus on different properties of intrasentential dependencies. In particular, the contrasts in (1)–(3) may not be easily distinguishable along 3 dimensions, which we detail from general to more specific:
Dependency: The presence or absence of 2 or more non-locally linked positions (positions in a sentence that contain information (e.g., phonological, thematic) about the same argument), whose parsing requires WM
Displacement: The presence or absence of a filler-gap dependency
Predictability: The ability of the parser to identify a dependency relation at an early stage of processing based on syntactic factors. In the movement relations in (1), the critical identifier is a constituent in an A’ position.
Dependency regards any intra-sentential nonlocal (ie, non adjacent) linking (e.g., between a gap and its filler, or an anaphor and its antecedent), Displacement relates to leftward syntactic movement, and by Predictability we mean a Dependency in which the antecedent can be identified as such long before the dependent element is encountered. All Displacements are Dependencies (Table 1i, ii); some, but not all Displacements are Predictable (Table 1i); finally, some (Table 1iii), but not all (Table 1iv) Dependencies that are not Displacements are Predictable. In this study, we attempt to distinguish between these, using sentence types that have never been tested in fMRI. These terms are not novel, as one – displacement - has featured extensively in linguistic and neurolinguistic theories and the other two – predictable, dependency – have featured extensively in psycholinguistic theories of syntactic complexity. The use of these terms in psycholinguistic theories of syntactic complexity is described below.
Table 1.
Types of Dependencies
| +Predictable | −Predictable | |
|---|---|---|
| +Displacement | i. Which paper did the student submit [---] | ii. Which paper did the student submit after reviewing [---] |
| −Displacement | iii. The papers that you wrote are/*is on the table | iv. The boy who likes Mary pinched himself |
Notice that we focus on A’ dependencies. The reason we do so is that we want to contrast displacement, predictability and dependency. These three factors can only be minimally contrasted in A’ dependencies. Whether or not the conclusions extend to other types of dependencies remains to be resolved by future work.
1.2 Predictability in Psycholinguistic Theories
Unlike in the field of psycholinguistics (Gibson, 1998; Hale, 2001; Levy, 2008), “predictability” has not played a major role in neurolinguistic investigations. Of particular interest, here, are those psycholinguistic theories that have related the difficulty of object over subject extraction (in relative clauses) to predictability. These theories are of two types: “Surprisal” and “Memory Resource”. “Surprisal” type theories assume object-extracted relatives are more difficult because they are less common (predictable). The parser, thus, expects the extraction to be from subject position and upon encountering an overt subject needs to make a revision. “Memory Resource” theories, such as the Syntactic Prediction Locality Theory (SPLT; (Gibson, 1998)) attribute the greater difficulty to holding the extracted constituent in a store until its predicted extraction site is located (ie, storage cost). More generally, the prediction of “surprisal” type theories is the less predictable the dependency (displacement) the greater the difficulty, whereas for the SPLT the longer the prediction is maintained (where length is measured in discourse referents) the greater is the difficulty. Although we will not be able to directly test these two theories, the version of predictability we describe above is in line with the SPLT, as previous fMRI studies have shown an effect of distance over the predicted dependency in Broca’s area (Makuuchi, Bahlmann, Anwander, & Friederici, 2009; Santi & Grodzinsky, 2007b).
1.3 Dependency and Psycholinguistic Theories
Additionally Memory Resource theories such as the SPLT predict difficulty effects for dependencies irrespective of predictability. According to the SPLT an additional source of complexity for object extraction over subject extraction in relative clauses emerges from the greater integration cost postverbally where the head of the relative needs to be integrated at its gap. Integration is higher here than at the subject gap, as there are multiple discourse referents along the path of the dependency. Given work by Gordon et al. (2001) the number of intervening discourse referents may not be sufficient to account for the difficulty effects. Rather the similarity, in terms of NP type (referential, pronoun, or proper name), between the extracted NP (the object) and the NP intervening (the subject) the dependency needs to be taken into consideration. Thus, according to these Memory Resource theories, a dependency (without predictability) is sufficient to generate difficulty effects as long as there are similar nouns/discourse referents positioned within the dependency.
1.4 Displacement and Psycholinguistic Theories
Psycholinguistic theories of complexity diverge from the neurolinguistic ones in that none of the latter allocates a special role to displacement. The psycholinguistic complexity dimensions do not distinguish displacement dependencies from other predictable dependencies, such as subject-verb agreement dependencies.
1.5 Psycholinguistic Theories and Brain Location
While none of these theories predict brain location, it is of interest to understand which, if any, of these psycholinguistic complexity dimensions Broca’s area operates on.
In previous work, we tried to tease apart Displacement and Dependency neurologically, as others have argued that as a filler (i.e., a leftward-displaced argument) is separated from its gap (i.e., a thematic position) over a distance, the establishment of the dependency requires WM, which activates Broca’s area in fMRI (Cooke et al., 2001; Fiebach, Schlesewsky, Lohmann, von Cramon, & Friederici, 2005; Santi & Grodzinsky, 2007b; Smith & Jonides, 1999). The more distant the dependent elements, the more WM resources their linking should recruit.
Broca’s aphasic patients, who are deficient in analyzing Displacement in receptive language, nonetheless retain considerable ability in handling a Dependency that crosses a discourse referent, such as reflexive binding (Grodzinsky, Wexler, Chien, Marakovitz, & Solomon, 1993; Santi & Grodzinsky, 2007a).
Against this background, we designed a parametric experiment that measured Blood Oxygen Level Dependent (BOLD) response in healthy people in fMRI, while contrasting a filler-gap relation with an antecedent-reflexive relation. We parameterized the distance between a displaced element and its gap (4), and compared the resulting growth in signal intensity in various brain regions to the effects of parameterized distance between a reflexive and its antecedent (5). In both cases distance was parameterized by number of intervening NPs (consistent with both (Gibson, 1998; Gordon et al., 2001). When we measured the BOLD response, Brodmann Area 45 responded as follows: there was a linear relation between the degree of activation in it, and the number of NPs that intervened between a gap and its filler; conversely, there was no such result for reflexive-antecedent relations. No other brain area showed this pattern (Santi & Grodzinsky, 2007b):
4. a. The mailman and the mother of Jim love the woman [filler who] Kate burnt [gap--] b. The mother of Jim loves the woman [filler who] the mailman and Kate burnt [gap--] c. Kate loves the woman [filler who] the mailman and the mother of Jim burnt [gap--] 5. a. The sister of Kim assumes that Anne loves the mailmani who burnt himselfi b. The sister of Kim assumes that the mailmani who loves Anne burnt himselfi c. Anne assumes that the mailmani who loves the sister of Kim burnt himselfi
While Broca’s area appears to have demonstrated selectivity for Displacement, Not that this response patterns characterized a single cluster throughout the brain. Note also that localization was done not by visual inspection, but quantitatively: this cluster was co-registered with Amunts et al.’s (1999) probability map of Broca’s area, and its determination as belonging in Brodmann’s Area 45 is as precise as current technology enables.
Note also that the sentence types that modulated a linear effect in it are also related to Predictability: in all of them, leftward deviation from canonicity is observed, as the displaced constituent not only resides outside the clause to which it belongs (in an A’-position), but also precedes it. Arguably, a leftward displaced constituent, identified as such early on in the string, might affect the analytic process: the parser, upon reaching the constituent following the relative pronoun who, recognizes that the constituent the woman in (4) cannot be the subject of the clause that immediately follows. The woman must therefore be a displaced filler. This makes the existence of a gap predictable. Displacement and Predictability go hand in hand in these cases.
Moreover, this coupling is not true for reflexives (6a): they satisfy Dependency, but as they are always A-bound, they satisfy neither Displacement nor Predictability. Indeed, a free lexical NP can replace the reflexive (6b), making this dependency unpredictable at the antecedent:
6. a. The waitress pinched herself b. The waitress pinched Mary
Finally, we note that in the past, there have been 2 experiments with Hebrew speaking Broca’s aphasics that compared the comprehension of Hebrew relative clauses with predictable gaps to comprehension of relative clauses with resumptive pronouns. These are relatively rare, A’-bound pronouns, and they are relevant here because as pronouns, they represent a Dependency; being A’-bound they are Predictable; and most analyses do not view them as maintaining a Displacement relation with their antecedent (Chomsky, 1977; Doron, 1982; Sharvit, 1999; Shlonsky, 1992):
7. a. ra’iti ‘et ha-’ishi she-ha-‘isha daxafa ‘otoi
saw-I ACC the-mani that-the woman pushed himi
I saw the mani whom the woman pushed himi
b. ra’iti ‘et ha-’ishi she- she-ha-‘isha daxafa [GAP--]i
saw-I ACC the-mani that-the woman pushed [GAP--]i
I saw the mani whom the woman pushed [GAP--]i
While (7a–b) contrast in Displacement (as only the latter is displaced), they are on a par with respect to Predictability, as both pronoun and gap are A’-bound, and so the existence of a Dependency can be identified early on. The patients failed with both gaps and their resumptive pronoun counterparts, when all else was kept constant (Friedmann, 2008; Grodzinsky, 1984). They were above chance, though, on relevant controls. This provides preliminary evidence that displacement alone cannot account for the results.
Given the complexity of the picture, we sought a contrast between Displacement, Dependency, and Predictability to further understand the role of Broca’s area in sentence comprehension. For that, we used a novel design. The predictions made by the two neurolinguistic theories being tested in the current study are presented in Table 2.
Table 2.
Functional accounts of Broca’s area and their predictions regarding activation in Broca’s area for the three types of contrasts.
| FUNCTIONAL ACCOUNT OF BROCA’S AREA | |||
|---|---|---|---|
| TDH | WM | ||
| Contrast | Displacement | Yes | No |
| Dependency | No | Yes | |
| Predictability | No | Yes | |
1.6 The present experiment
To tease apart the 3 factors in relation to Broca’s area we had three main contrasts. First we will provide a brief summary of the contrasts and analyses and then we will provide some detail about the structural facts of the sentences and contrasts used. We tested Displacement by comparing the fMRI signal for (+Dependent, +Displaced, −Predictable) parasitic gaps in PP adjuncts (Table 3a), with A’-bound pronouns (Table 3b) that freely vary in the same syntactic environment (+Dependent, −Displaced, −Predictable). Next, we tested Dependency by keeping Predictability and Displacement constant by contrasting the parasitic gap and optional A’-bound pronoun (Table 3a, b) to a PP adjunct without any dependency (Table 3c, d); and finally, we tested Predictability through contrasting a wh-object question (Table 3c) with a wh-subject question (Table 3d).
Table 3.
Example sentences from the four conditions1.
| Condition | Example Sentence |
|---|---|
| a. Parasitic Gap PAR | |
| b. Pronoun PRO | |
| c. Wh-Obj. Question WHO | |
| d. Wh-Subj. Question WHS | Which tired student submitted a paper after jogging? |
We fortified our results by complementing the minimal contrasts with a robust, conservative analytic technique called Conjunction Analysis, to which our experimental setup lends itself. Next, we discuss the sentence types we used, and their properties before discussing the contrasts and the conjunctions they entered into.
We start with the central sentence type – one having an unpredictable displacement, namely (+Dependent, +Displaced, −Predictable). Sentences with a parasitic gap in a PP adjunct satisfy this requirement (8):
8. [filler Which paper] did the tired student submit [gap--] after reviewing [p-gap--]?
The first gap in (8) is a regular, object-gap. The second gap, however, is special, and is known as a “parasitic gap” because its grammatical existence depends on the first (Engdahl, 1983; Nissenbaum, 2000; Ross, 1967; Taraldsen, 1981). If the first “matrix” gap is filled, the sentence becomes ungrammatical (9); yet the matrix gap is not dependent on the parasitic one (10). In the class of structures we discuss here, the parasitic gap always follows the “real” gap. The requirement that each filler must be associated with a gap is thus satisfied at the “real” gap, and the parasitic one, that appears in an untensed adverbial adjunct PP, is optional, as this adjunct may be gapless and contain an overt object (10a), an intransitive verb (10b), or it can be dropped altogether (10c):
9. *[filler Which paper] did the tired student submit the exam [PP after reviewing [p-gap--] ]?
10. a. [filler Which paper] did the tired student submit [gap--] [PP after reviewing the literature]?
b. [filler Which paper] did the tired student submit [gap--] [PP after jogging]?
c. [filler Which paper] did the tired student submit [gap--]?3
Importantly, the parasitic gap in (8) is on a par with the regular gap found with syntactic movement in relative clauses (Nissenbaum, 2000)4. This structure thus satisfies the Displacement property, but not Predictability because the syntax does not require it (ie, it provides multiple options). Psycholinguistic studies confirm that the parser does not go looking for a gap in post-verbal adjuncts that host optional parasitic gaps, as it does earlier for the real gap (Wagers & Phillips, 2009). This has been shown through demonstrating durational differences in reading times following a real gap between plausible and implausible fillers, but no such difference at the point of an optional parasitic gap within the adjunct. The converse finding that the parser does go looking for a gap in optional subject parasitic gap environments (Phillips, 2006) is also fully compatible with our construal of Predictability. At a subject parasitic gap the need to find a gap for the filler is not yet satisfied by a real gap (Wagers & Phillips, 2009). Thus, the parser is looking to satisfy the predictability of a gap. This finding of course further enriches our understanding of the parser in island environments and incremental structure building, but critically does not contradict our definition of Predictability with respect to optional object parasitic gaps.
This structural configuration, moreover, lends itself to another desirable contrast: the parasitic gap in (8) is not only A’-bound and unpredictable, but also optional in another respect: it freely varies with a (relatively rare) A’-bound, unstressed pronoun (11), to produce a sentence with (almost) the same meaning5 (Engdahl, 1983):
11. a. [filler Which paper] did the tired student submit [gap--] after reviewing it?
In (11), the pronoun inside the PP adjunct stands in an Unpredictable Dependency relation with its A’-antecedent, and it is not Displaced. In this respect, this pronoun is reminiscent of the Hebrew resumptive pronoun with the exception of predictability (since its A’ antecedent is satisfied at the real gap), as it is an (+Dependent, −Displaced, −Predictable), A’-bound pronoun. Thus the contrast between (11) and (8) provides a (unpredictable) Displacement contrast where dependency and predictable displacements are controlled.
The sentences in (8), (10)–(11) lend themselves to a test of Dependency. This is done through a comparison between the (+Dependent with respect to the adjunct) parasitic gap (8) and pronoun (11) and the similar structure (10b) that is (−Dependent) with respect to the adjunct.
Finally, we included a control contrast, so that the neural substrates of the two contrasts just discussed can be compared on the same data set to the object-movement vs. no movement effect in Broca’s area. This contrast has previously modulated Broca’s area in both health and disease (Ben-Shachar et al., 2003; Ben-Shachar, Palti, & Grodzinsky, 2004; Bornkessel, Zysset, Friederici, Von Cramon, & Schlesewsky, 2005; Caplan, Alpert, & Waters, 1999; Caplan, Alpert, Waters, & Olivieri, 2000; Friedmann, 2008; Friedmann & Gvion, 2003; Grewe et al., 2005; Grodzinsky, 1989, 2000; Just et al., 1996; Stromswold et al., 1996). Its presence would help to create a baseline against which our new exploration can be studied. To obtain a minimal contrast for it, we used subject and object wh-questions, with an adjunct, but with no dependency in the latter. We expected the contrast in (12)–(13) to activate Broca’s area, in line with past results:
12. [filler Which paper] did the tired student submit [gap--] after jogging? 13. Which tired student submitted a paper after jogging
Further, this is the only contrast that compares a sentence with +Predictability to -Predictability. Thus, it allows for the ability to test the effect of Predictability. Although the contrast also tests Displacement and Dependency, their effects are tested in the previous two contrasts.
In sum, we tried to further understand the role of Broca’s area in filler gap dependency comprehension, through an fMRI study of all 3 dimensions - Displacement, Dependency, and Predictability. Examples of the 4 sentence types are presented in Table 3.
1.7 The analytic idea – Contrasts and Conjunctions
We studied parasitic gaps in fMRI for the first time. Yet, what makes this study special is not just the technology and the constructions we studied, but also, the basic approach. That is, in addition to standard comparisons between conditions, we used an analytic technique called Conjunction analysis in order to quantify brain responses to psycholinguistic generalizations, which complement the minimal contrasts by providing a more conservative analysis that tests their robustness.
Our analysis proceeded in 2 stages: first, we set up minimal pairs of conditions that formed statistical contrasts. The result was an array of voxel clusters, marking contiguous chunks of brain voxels for which the difference in signal intensity between members of a minimal pair was statistically significant. Second, we took these clusters – the results of the contrasts between our minimal pairs – and arranged them according to each of our 3 dimensions – Displacement, Dependency and Predictability. As Table 4 shows, each dimension clusters different contrasts. The Displacement conjunction identifies regions sensitive to an unpredictable displacement. Given the unpredictable displacement (PAR) depends on a predictable displacement, the contrast factors out the effect of a predictable displacement by subtracting (PRO, WhO, or WhS) across 3 contrasts. The Dependency conjunction identifies effects of a dependency in the adjunct (PAR, PRO) by subtracting out the effect of each of these the dependency in the main clause (WhO, WhS) across two contrasts. The Predictability contrast takes cases of a predictable displacement (WhO, PAR, PRO) and subtracts from each of them the case of no displacement (WhS). A corresponding conjunction would quantify this psycholinguistic generalization by marking those brain voxels for which a significant difference was found for all the contrasts being conjoined. A conjunction, then, essentially identifies the cross-contrast intersection. We carried out conjunction analyses for all 3 dimensions. Next, we provide the details of our contrasts and analyses.
Table 4.
Conjunction of contrasts used in statistical analyses. Par, Parasitic; Pro, Pronoun; WhO, Wh-Object; WhS, Wh-Subject.
| Type of Conjunction | Contrasts |
|---|---|
| i. Displacement | [Par – Pro] & [Par – WhO] & [Par – WhS] |
| ii. Dependency | [Par – WhO] & [Par – WhS] & [Pro – WhO] & [Pro – WhS] |
| iii. Predictability | [Par – WhS] & [WhO – WhS] & [Pro – WhS] |
1.8 Main Contrasts
The minimal Displacement contrast [PAR–PRO] between (8 and 11), is between an A’-bound gap with a (relatively rare) A’-bound pronoun, where all else (meaning, number of dependencies, distance/storage requirements and other variables) is equal. A [PRO–WHO] comparison, between a sentence with an unpredictable pronominal dependency in its PP adjunct, and another that has no dependency in its PP adjunct (10b) along with the parasitic gaps, provides a test of Dependency. Finally, there is a control comparison [WHO–WHS], aimed to replicate past work, and to provide a test of Predictability.
Additional contrasts were computed to provide further support to the conclusions drawn from the results of the main contrasts. These additional contrasts, (PRO-WHS) and (PAR-WHS), along with those already discussed, entered into three conjunction analyses (Nichols, Brett, Andersson, Wager, & Poline, 2005).
1.9 Conjunction Analyses
The contrasts (PAR-WHS) and (PAR-WHO) are similar to (PAR-PRO) in that they contrast a condition with two displacements to a condition with one or no displacement, thus investigating a displacement effect. The logic of this conjunction analysis assumes that Broca’s area linearly responds to number of displacements. Evidence that Broca’s area is sensitive to number of displacements rather than only activating for a single displacement has previously been provided (Friederici et al., 2006). Though from that study it is unclear, whether it is due to dependency, displacement, or predictability.
Additional contrasts were computed to investigate the effect of an unpredicted dependency. The contrasts, (PAR-WHS) and (PRO-WHS) are similar to (PAR-WHO) and (PRO-WHO), in that they differ with respect to whether or not there is a dependency extending into the adjunct. Thus, a conjunction analysis was computed with the following contrasts (PAR-WHO), (PAR-WHS), (PRO-WHO), and (PRO-WHS) to test for a dependency effect.
The additional contrasts, (PAR-WHS) and (PRO-WHS), are similar to the contrast (WHO-WHS) in that they contrast a condition with an object displacement to a condition with no displacement, thus, providing a standard control that does not distinguish between dependency, displacement or prediction, but is unique in its test of prediction. Thus, these three contrasts were entered into a third conjunction analysis to test for a predictability effect.
2 Materials and Methods
2.1 Participants
Fourteen students in Linguistics at McGill University volunteered. One of the participants was excluded from the analysis due to low behavioral scores. All participants were native English speakers, right-handed and had normal hearing (9 females; X̅age=23.68yrs). Participants gave informed consent in accordance with the ethics committee of the Montreal Neurological Institute (MNI) and in accordance with the Helsinki Declaration of 1975, as revised in 1983.
2.2 Stimuli
There were four conditions (Parasitic (PAR), Pronoun (PRO), Wh-Object (WHO), and Wh-Subject (WHS); see examples in Table 3 and Appendix A for complete list). A general description of each of these syntactic conditions was already provided in the Introduction. Consistent with the examples provided in the Introduction, the object argument was most frequently inanimate, given the difficulty of constructing parasitic gap constructions with animate objects. This presents a conservative bias on our part, given that performance differences between object and subject extraction is greater when the two arguments are animate than when the subject is animate and the object is inanimate (Traxler, Williams, Blozis, & Morris, 2005).
There were 30 sentences per condition repeated in each of the 2 runs, for a total of 120 unique sentences, and 240 trials. Over the entire experiment, sentences contained between 13 and 17 syllables, but minimal pairs between conditions differed by at most 2 syllables. On average the sentences were 3600 ms long. Each trial lasted 6000 ms, with approximately the first 2000 ms being occupied by silence and the following 4000ms occupied by the sentence. The slight variability in the duration of the sentences caused for slight variability in the duration of silence at the beginning of the trial, which was 2000ms at a minimum.
2.3 Task
In order to be assured that the participants were attending and understanding the sentences, they were given a comprehension task. Randomly, on one-third of the trials, participants were asked a comprehension question. The decision to have comprehension questions on one-third rather than all of the trials was based on maximizing the number of experimental trials while minimizing the duration the subject would be in the scanner. Given that the stimulus sequence was random, the subject could not predict when there would be a comprehension question forcing them to constantly attend.
On trials with a task, participants would hear the word “question” 500ms after the offset of the experimental stimulus to prompt them to the upcoming comprehension question. Further, the comprehension questions were recorded by a male speaker, whereas the experimental sentences were recorded by a female speaker. This distinction in the sex of the speaker was used to distinguish experimental sentences from comprehension questions. The experimental instructions provided to the participants included a description of these features of the comprehension trial to prepare them. The subject was required to press one of two buttons on a response box to indicate whether their response was “yes” or “no”.
The comprehension question asked either about the matrix clause or the information contained within the adjunct but not both. The comprehension questions requiring a false response altered any of the following: the adjective on the subject, the subject noun, the verb (in the matrix or the adjunct), or the object noun. There were equal numbers of true and false comprehension questions. An example stimulus sentence and comprehension question is presented in (14) and (15), respectively.
14. Which paper did the tired student submit _ after reviewing _?
15. Did a lazy student submit a paper? [Correct Response: NO]
2.4 Procedure
One to two days prior to MRI scanning the subject came into the lab and they were provided with task instructions and given a short practice session. This practice session provided an opportunity to address any questions or concerns participants may have about the task, or otherwise, prior to them being inside the imaging machine. On the day of MRI scanning the subject was positioned on the bed of the MRI machine. An air-vacuum pillow was used to minimize head movement. Stimuli were presented through high quality pneumatic-based Headphones (Silent Scan 3100 System, Avotec Inc.) that attenuated surrounding noise by ~30dB. A high-resolution anatomical scan was first acquired, followed by a short practice run of 12 sentences. The practice run provided an opportunity to confirm that the volume was adequate and that all equipment was properly connected, in addition to giving the subject a chance to warm up to the task. The subject then performed the task across two runs of stimuli. Stimuli were presented in a rapid event related design. The inter stimulus interval (ie, silence) was minimally 1 TR, but was further jittered by multiples of the TR (ie, 0–5 additional volumes of silence could follow). The number of additional null volumes used to jitter stimulus onset was approximately equivalent to the number of volumes per experimental condition (60). Both the order of conditions and amount of jitter per trial was optimized by the Linux program Optseq (http://surfer.nmr.mgh.harvard.edu/optseq/). Presentation order of the two runs was counterbalanced across participants.
2.5 fMRI Parameters
Image acquisition was performed with a 1.5T Siemens Vision imager at the MNI in Montréal, Canada. A localizer was performed followed by whole-brain T1-weighted imaging for anatomical localization (256×256 matrix; 160–176 continuous 1.00mm sagittal slices). Each functional volume was acquired with a 64 × 64 matrix size and a total volume acquisition time of 2000 ms. Each imaging run produced 582 acquisitions of the brain volume (TE = 50ms; TR = 2000 ms; FA = 40°; FOV = 320×320 mm; 20, 5mm thick slices).
2.6 Behavioural Data Analysis
The behavioral percent correct comprehension data were averaged by condition across runs for each subject and was then submitted to a one-way within-subject ANOVA with the factor Syntax (PAR, PRO, WHO, WHS) in SPSS.
2.7 fMRI Data Analyses
Statistical analyses were performed with Brainvoyager QX v1.7 software (www.brainvoyager.com Goebel, Esposito, & Formisano, 2006). The data were slice-scan time corrected (sinc interpolation), motion corrected, and processed to filter out linear trends. The first two volumes of silence (in a sequence of 7) in each run were excluded from the analysis. The functional data was coregistered to the anatomical data through a two step process. An initial alignment was performed based on position information contained in the header of the functional and anatomical files, followed by a fine-tuning alignment based on intensity correlation between the functional (inverted intensities values were used) and anatomical data in order to compensate for any head motion between the acquisition of the anatomical and the functional. In many cases where the fine-tuning alignment did not produce a perfect alignment, minor manual adjustments were made. The individual functional and anatomical data was transformed into talairach space and the talairached functional data were smoothed with a Gaussian spatial filter (FWHM=8mm).
2.8 Deconvolution Analysis
Statistical analyses were based on the General Linear Model (GLM). Given that a rapid event-related design was used, the hemodynamic response function (HRF) of one condition overlapped with that of others. The signal was deconvolved over 10 time points from stimulus onset (ie, 10 predictors per condition) for each of the four experimental conditions and the comprehension condition. It should be noted that increasing the number of predictors to 15 or 20 did not affect the pattern of results, indicating their robustness. Stimulus onset was taken as the beginning of the 4000ms following the first 2000ms of silence in a trial, with the onset of the 4000ms containing a variable amount of silence given sentence length varied. The comprehension condition included the two volumes following the experimental sentence, which included the 500ms silent interval as well as the comprehension question. The experimental sentence was not included in the comprehension condition and the comprehension condition was not divided up according to the experimental condition to which it was associated. The comprehension condition did not enter into any contrast as it was used to enforce participants to attend to the experimental stimuli and was used to exclude participants from the analysis.
The deconvolution analysis produced beta-weights for the series of volumes (ie, 10 for a TR=2sec) post stimulus onset over which the signal is deconvolved, unlike the regular convolved GLM which produces one beta-weight for how well the data fit a standard HRF. The deconvolution analysis provides an advantage over the regular convolution analysis, as it allows for a flexible HRF. This is an advantage, as different brain regions may demonstrate different hemodynamic response parameters. For example, more recent studies (with the use of slow event related designs) have demonstrated that the HRF in temporal cortex has a shorter latency to peak from stimulus onset than that of the frontal cortex (Belin, Zatorre, Hoge, Evans, & Pike, 1999; Dehaene-Lambertz et al., 2006; Friederici et al., 2006). Given the flexibility of the HRF in the deconvolution analysis, it does not present a bias, as the standard HRF may if it is more prototypical of some cortical areas than others.
2.9 Contrasts
The main minimal pairs that formed statistical contrasts were (PAR-PRO), (WHO-WHS), (PAR-WHO) and (PRO-WHO). For all contrasts, the beta-weights for scans 2–6 (corresponding to 4–12sec post sentence onset), which corresponds to the approximate timing of the peak in the HRF, were entered into the contrast. Group maps (Random Effects) were corrected for serial correlations. Additional contrasts were computed to provide further support to the conclusions drawn from the results of the main contrasts. These additional contrasts, (PRO-WHS) and (PAR-WHS), along with those already discussed, entered into three conjunction analyses (Nichols et al., 2005).
2.10 Conjunction Analyses
All conjunction analyses were first thresholded at a voxel-wise level of p < .05 and then corrected for multiple comparisons (p < .05). In Brainvoyager there are two different methods for running a conjunction analysis, we used the method that demonstrates an intersection of all 3 maps at p<.05. Thus, for every cluster each of the contrasts activate every voxel of that cluster at p<.05. Given that, a voxel-wise p-value of .05 is quite stringent when considering that this value needs to be reached in all three individual contrast maps at any particular voxel to appear in the resulting conjunction map. This results in a .000125 (.05*.05*.05) probability that a voxel is active simply due to chance in the conjunction map. The correction for multiple comparisons was based on a cluster size thresholding analysis (Forman et al., 1995). The cluster size correction is determined by two factors: (1) the map’s spatial smoothness and (2) Monte Carlo simulation for estimating cluster-level false-positive rates. The simulation provides the minimum cluster size associated with a cluster-level false-positive rate of 5%. Given that each map has a different number of clusters and sizes the cluster size for correction will differ across the maps. Further, if there are too many false positives, a value will not be reported.
The displacement effect and prediction effect conjunction contrasts had a cluster criterion size of 840mm3 and 803mm3. The dependency effect conjunction contrast did not produce any clusters for a multiple comparison correction.
2.11 Cytoarchitectonic Probability Calculations
For activation clusters that appeared within Broca’s area, the probability that these clusters were within BA 44 and/or 45 was calculated using the probability maps of Amunts et al. (1999; http://www.bic.mni.mcgill.ca/cytoarchitectonics/). The probability calculations required a 5-step process: (1) Extract talairach coordinates for all voxels within a cluster (2) Convert the coordinates to MNI coordinates using the matlab script tal2mni (http://imaging.mrc-cbu.cam.ac.uk/imaging/MniTalairach) as the Amunts map (1999) was in the MNI coordinate system and ours were in Talairach and Tournoux (1988), (3) Convert the MNI world coordinates to voxel coordinates of the MINC files (with respect to the probability maps of BA44 and BA45 that are within MNI space), (4) Extract probability values from the voxels within the probability maps, and (5) average all values within a cluster to obtain the final number. Since the values represent the number of subjects with overlapping cytoarchitectonic areas at any particular voxel, the average value was divided by the number of brains used in generating the cytoarchtectonic maps (ie, 10) and multiplied by 100%6.
3 Results
3.1 Behavioral Results
The participants comprehended the sentences, as they were 92–96% accurate, depending on the condition. There was no significant main effect of sentence type (F(3,36)=2.63, p=.07) (see Figure 1).
Figure 1.
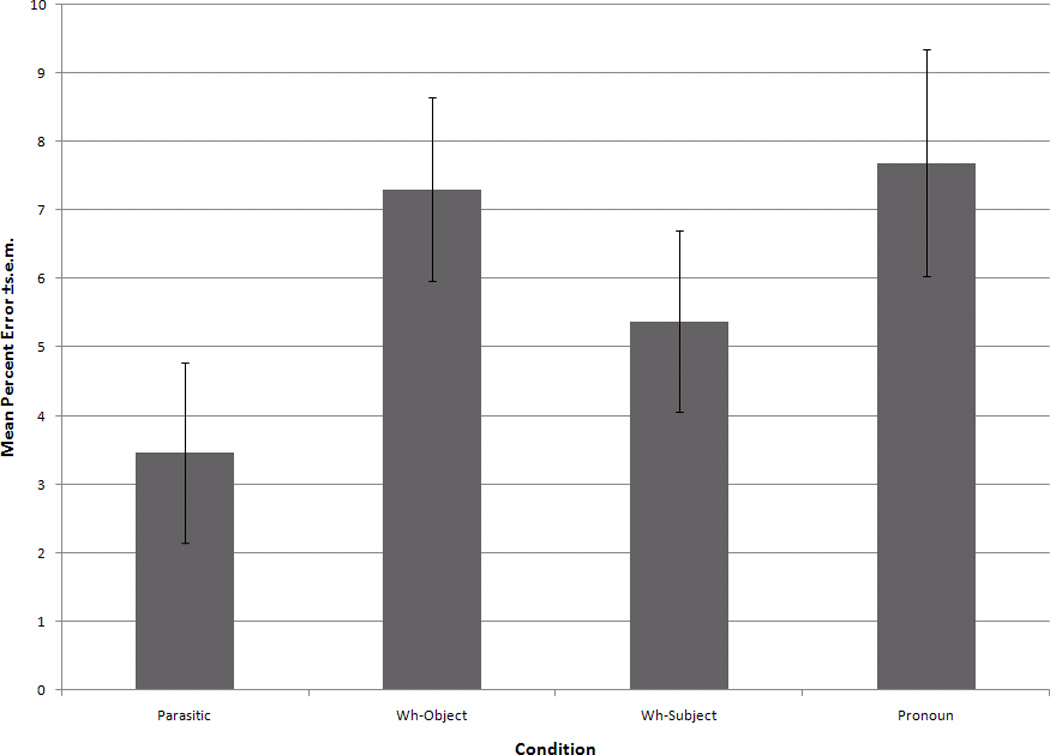
Mean percent errors +s.e.m. across all four conditions.
3.2 fMRI Results
We first focus on the results from the conjunction analyses since they are based on rigorous linguistic generalizations and then look at minimal pairs to further understand the individual contributions.
The conjunction analysis used to isolate the predictability effect resulted in a cluster of activation within the left Inferior Frontal Gyrus (IFG; see Table5). The activation included 1220 mm3 (see Table 5; see Figure 2). The probability that the voxels are within BA 45 is relatively high at 23%. For a discussion of Amunts et al.’s (1999) probability maps and why a percentage of 23% is high see Santi and Grodzinsky (2007b).
Table 5.
fMRI results from the conjunction of Random Effects analysis, corrected for serial correlations. BA, Brodmann area.
| Predictability Conjunction [(WHO-WHS), (PAR-WHS), (PRO-WHS)] | ||||
|---|---|---|---|---|
| Landmark | Hemisphere | Talairach coordinates (x, y, z) |
BA | mm3 |
| Inferior Frontal Gyrus | Left | −46, 38, 6 | 45 | 1220 |
Figure 2.

Activation of the Left Inferior Frontal Gyrus by the conjunction of contrasts that investigated a predictability effect. Maps are thresholded at p<.05 (voxel-wise) and above a cluster of criterion size such that p<.05 (corrected for multiple comparisons). The activation is overlaid on an average of all 13 participants’ anatomical scans. The deconvolution plots present deconvolution coefficients over 10 scans from stimulus onset.
The minimal pairs that contributed to the Predictability conjunction analysis were individually investigated at a higher voxel-wise threshold (p<.01) and corrected for multiple comparisons (p<.05). The [who-whs] contrast resulted in the greatest volume of activation with two clusters measuring 2063 and 2585 mm3 in volume. The contrast that resulted in the second largest cluster was [pro-whs] with a 2845 mm3 volume. The [par-whs] contrast had the smallest volume of activation of 485 mm3 (see Figure 3).
Figure 3.

Activation of the Left Inferior Frontal Gyrus by the individual contrasts that contributed to the predictability effect conjunction map. Maps are thresholded at p<.01 (voxel-wise) and above a cluster of criterion size such that p<.05 (corrected for multiple comparisons). The activation is overlaid on an average of all 13 participants’ anatomical scans.
The conjunction analysis investigating a displacement effect did not result in any clusters of activation. Similarly, the conjunction analysis used to investigate the effect of dependency did not result in any clusters of activation.
A summary of the results from the minimal contrasts is provided in Table 6. Consistent with the conjunction analysis, the only minimal contrast that activated Broca’s area was the one investigating a predictability effect. The minimal contrasts for predictability and dependency demonstrated effects at a voxel-wise p<.01 and corrected for cluster size (p<.05), whereas the displacement effect only appeared at a voxel-wise p<.05 when corrected for cluster size (p<.05). However, lowering the voxel-wise threshold to p<.05 for the dependency contrast did not change the pattern of results. Interestingly the novel parasitic gap condition (displacement effect) activated the left anterior Middle Temporal Gyrus (see Figure 4).
Table 6.
Minimal Contrasts
| Landmark | Hemisphere | Talairach coordinates (x, y, z) |
BA | mm3 |
|---|---|---|---|---|
| who-whs | ||||
| Middle Temporal Gyrus | Right | 38, −1, −15 | 21 | 446 |
| Middle Frontal Gyrus | Right | 24, 48, 18 | 10 | 1125 |
| Basal Ganglia | Right | 25, 0, 6 | -- | 344 |
| Parahippocampal Gyrus | Right | 21, −18, −13 | 36 | 399 |
| Cingulate Gyrus | Right | 11, 26, −6 | 32 | 1184 |
| Basal Ganglia | Left | −23, 0, 8 | -- | 264 |
| Inferior Frontal Gyrus | Left | −32, 34, −9 | 45 | 2064 |
| Inferior Frontal Gyrus | Left | −41, 44, 15 | 45 | 2585 |
| Ventral Precentral Sulcus | Left | −49, 3, 37 | 6/8 | 424 |
| Superior Temporal Sulcus | Left | −51, −2, −12 | 22 | 225 |
| Superior Temporal Gyrus | Left | −55, 8, 0 | 22 | 555 |
| Par-Pro | ||||
| Middle Temporal Gyrus | Left | −51, −12, −11 | 21/22 | 1666 |
| Pro-who | ||||
| Middle Frontal Gyrus | Right | 29, 33, 3 | 10 | 354 |
| Ventral Precentral Sulcus | Left | −40, 0, 39 | 6/8 | 1315 |
| Supramarginal Gyrus | Left | −43, −42, 34 | 40 | 203 |
Figure 4.

Activation of the anterior Left Middle Temporal Gyrus by the minimal contrast for the Displacement effect (par-pro). Maps are thresholded at p<.05 (voxel-wise) and above a cluster of criterion size such that p<.05 (corrected for multiple comparisons). The activation is overlaid on an average of all 13 participants’ anatomical scans.
4 Discussion
4.1 Summary of the design and results
This study compared sentences with multiple dependency relations. The novel use of parasitic gaps and their pronoun alternatives enabled us to simultaneously look for the brain signature of three effects potentially associated with processing filler-gap dependencies: (1) displacement, (2) dependency and (3) predictability. The displacement effect was assessed by a conjunction of contrasts that increased the number of gaps across two sentences from one to two, while maintaining all else constant (including the number of dependencies and dependency distance). Dependency was investigated through a conjunction of contrasts between sentences with two dependencies to sentences with one. Lastly, predictability was investigated by manipulating the type of wh-question (object or subject). This rich design enabled the use of conjunction analyses that pertained to broad generalizations (all carried out on the same data set), in addition to subsequent tests of individual contrasts. By investigating how these factors relate to one another, we were able to break down the filler-gap relation into finer neurological component parts.
The central finding, a predictability cluster of activation in Brodmann’s Area 45, is in keeping with earlier results from both aphasia and neuroimaging with healthy subjects (Ben-Shachar et al., 2003; Ben-Shachar et al., 2004; Bornkessel et al., 2005; Caplan et al., 1999; Caplan et al., 2000; Fiebach et al., 2005; Friedmann & Shapiro, 2003; Grewe et al., 2005; Grodzinsky, 1989; Santi & Grodzinsky, 2007a, 2007b). The absence of activation in Broca’s area by the two additional novel minimal pairs and their conjunctions, dependency and displacement, runs counter to any current functional theory of this region in sentence comprehension. However, it is consistent with Gibson’s (1998) storage component of syntactic complexity. We now discuss the neurolinguistic theories, and consider possible modifications.
4.2 Implications of Absent Dependency and Displacement Effect
The absence of a dependency effect in Broca’s area speaks against accounts that attribute the role of linking non-adjacent positions in a sequence to this region. The dependency conjunction included contrasts with dependencies that cross an overt subject in the matrix clause, or a covert one in an adjunct PP. The WM and syntactic complexity accounts predict a dependency effect, and its absence suggest that some modifications to their formulations may prove fruitful.
Next, consider related accounts, that consider dependency to be an insufficient condition to account for the object-subject movement contrast in Broca’s area, but rather focus on properties of crossing. In English, movement from object (but not subject) position crosses an argument (ie., intervener). The claim is that the existence of an intervener of similar type to the head of the dependency (ie., NP or argument) is the source of the contrast in Broca’s area. These accounts are based on results from contrasts between a dependency that crosses one intervener, and a dependency in which no interveners are crossed (Friedmann, 2008; Gordon et al., 2001; Grillo, 2008, 2009; Grodzinsky & Amunts, 2006). Yet, these accounts do not appeal to incremental intervention (i.e., when the number of interveners changes from 1 to 2 to 3). As the current test compares a movement dependency across two interveners to a dependency across one, it is difficult to formulate the prediction that these accounts would make for the dependency contrast. Later, we will propose an amendment to the WM and intervener-based accounts that might cover the current findings.
Finally, consider the movement-related account of the function of Broca’s area (Trace-Deletion Hypothesis, (Grodzinsky, 1995, 2000). The absence of a displacement effect likewise calls for its refinement, at a minimum. Initially designed to cover all traces, the TDH later restricted the role of Broca’s area to the processing of traces of phrasal movement, as verb movement proved intact in Broca’s aphasia (Grodzinsky, 1995, 2000). Our present results seem to suggest further restriction of the scope of this account, to the effect that only predictable traces are processed in Broca’s area. Is such a restriction justified? To ponder this question, we discuss predictable and unpredictable traces.
Predictable traces are, essentially, those linked to overt antecedents on their left, outside the basic clausal boundaries – antecedents in A’-positions (mostly marked in a manner that identifies this position, e.g., by a wh-word, or focus stress). This setup allows for early realization that an NP encountered in this position is displaced (a filler), which should provide a hearer an early warning that she should be storing that NP in memory and go hunting for a gap. Our results only suggest that filler-gap relations of this sort are processed in Broca’s area.
But what about other filler-gap relations, in particular those generated through A-movement? Unlike A’-moved antecedents, A-moved NPs (typically subjects of passive and Raising constructions) cannot be immediately identified as displaced antecedents in left-to-right scanning, which makes their traces unpredictable. If Broca’s area only supports the analysis of predictable dependencies, then an active/passive contrast should not result in fMRI activation of this region, nor should Broca’s aphasic patients be impaired in comprehension of passive.
Little fMRI evidence that bears on this issue is available: an fMRI study of passives (Yokoyama et al., 2007) found activation within the left frontal operculum (in close vicinity to Broca’s area). Notably, this study was carried out in Japanese, a language in which the status of passive is under debate (Hoshi, 1994). No reliable evidence from other languages –whether from passive or from other sources of “unpredictable” movement – is available.7
We might seek additional clues in the comprehension performance of Broca’s aphasic patients – an early data source for the TDH. The picture is complex: while comprehension of passive is no doubt compromised (Goodglass, Barton, & Kaplan, 1968); whether or not it presents a pattern of impairment and sparing that is exactly the same as the pattern for predictable movement is still under discussion (for a comprehensive review and a quantitative analysis see (Drai & Grodzinsky, 2006). At this point, though, the following descriptive statements can be made with considerable certainty: in both predictable and unpredictable movement constructions, patients perform around chance, and significantly lower than their baseline (X̅passive=0.63; X̅active=0.83; X̅object-relative =0.57; X̅subject-relative=0.84, see (Drai & Grodzinsky, 2006), for distribution curves). However, the variances in performance differ (passive=0.21; σactive=0.13; σobject-relative=0.12; σsubject-relative=0.09): passive is almost twice that of all other conditions, and X̅passive > X̅object-relative. Broca’s area (or the surrounding cortex), then, is undoubtedly involved in the analysis of passive, but the manner of its involvement may be different from A’-movement. The nature of this difference remains to be characterized. Hopefully, future fMRI studies will help resolve this important issue.
4.3 Two Possibilities
We find ourselves in a dilemma: we can either (a) account for our results by restricting the role of Broca’s area in movement to the processing of relations pertaining to predictable traces, or (b) maintain that it has a broader role in movement. Either possibility fails to capture all the data: A restricted role would exclude the results from passive that pertain to unpredictable A-movement; while a broader role would fail to exclude the unpredictable traces in the current experiment, namely those found in PP adjuncts. Further, both possibilities fail to explain the gradation in performance from the best performance on no movement, to worse performance on A movement to the worst performance on A’ movement that is hinted at by current aphasia results. It seems the most prudent approach is to pursue additional research that directly compares relative signal intensity between A and A’ movement on the same data set to more conclusively understand their relation before expecting current theories to account for this suspected gradation.
Further, if the region is more broadly engaged in movement, then we should wonder why in the current study we did not see activation from the contrast that tested displacement. It could well be that the absence of a displacement effect for an unpredictable A’-movement, in the current study, may be a mere artifact of contrasting a sentence with two displacements to a sentence with one displacement: a second displacement may have less of an effect than an initial one, because sensitivity to local demands (as reflected by activation levels) diminishes as processing of a sentence progresses, thereby hampering the statistical detectability of a contrast that pertains to a second gap in a sentence. This seems less likely, given that Broca’s area has been shown to be sensitive to multiple +Predictable displacements (Friederici et al., 2006). While there are multiple possible interpretations of the Friederici et al (2006) study, it nonetheless demonstrates that neural activity does not saturate after one instance of a syntactic operation.
4.4 Integrating the Accounts
An alternative to the accounts we considered is a hybrid of those from the Introduction: Broca’s area hosts a syntactically constrained WM. This WM would be constrained to syntactic dependencies that involve displacements with a distance metric measured according to the number of intervening verbal arguments. Given results from the current study and earlier studies on reflexive binding dependencies (Grodzinsky et al., 1993; Santi & Grodzinsky, 2007b), it appears that the presence of a dependency is not a sufficient condition for activating Broca’s area, but rather a predictable displacement is required. This conclusion is very similar to Gibson’s (1998) storage dimension with the exception that we propose to calculate distance in a distinct way. Although there are many ways one can construe distance (e.g., number of syllables, words, phrases, clauses etc), we and others (Friedmann, 2008; Gordon et al., 2001; Grillo, 2009; Santi & Grodzinsky, 2007b) have similarly argued that a critical property of the object vs subject contrast in Broca’s area is the NP intervener, as they are of the same category as fillers (Frazier, Clifton, & Randall, 1983). We have further argued that complexity increases with increasing number of these intervening NPs (Santi & Grodzinsky, 2007b). In the current study the object wh-question has an NP/argument intervening between the filler and the gap, whereas the subject wh-question clearly does not, which is consistent with this account. The cases in which NPs and verbal arguments that are made up of complex NPS are distinguishable require further study in establishing the appropriate distance metric (NPs or verbal argument)8. Thus, it seems likely that the contrast between object and subject wh-questions is related to a specialized form of syntactic WM for predicted dependencies with NP/argument interveners. Though here we have primarily focused on predicted displacements, it would be interesting to test further implications of such an account, such as predictable relations between a triadic predicate and its arguments.
There is some indication that this account may generalize to other predictable dependencies than displacement. Studies that modulate the distance of subject-verb agreement have found Broca’s area sensitive to this distance (in NPs) as well (Makuuchi et al., 2009; Santi & Grodzinsky, 2010). However, the situation is complicated by the fact that there is some indirect evidence that A’ dependencies rely to some degree on distinct anatomy within Broca’s area than A dependencies (Santi & Grodzinsky, 2010). A direct test of these two dependencies is still required.
4.5 Other brain regions
In addition to the main result within Broca’s area across the conjunction analyses, the minimal contrasts demonstrated clusters of activation. The minimal contrast investigating displacement activated the left anterior temporal lobe, which is consistent with previous studies that has found this area activated by syntax (Humphries, Love, Swinney, & Hickok, 2005; Noppeney & Price, 2004; Rogalsky & Hickok, 2009; Stowe et al., 1999). The predictability minimal contrast also activated a very similar region, although the activation spread more superiorly. One of the minimal contrasts [pro-who] investigating the dependency effect activated the left ventral precentral gyrus, which has been activated by a variety of additional neuroimaging experiments of sentence comprehension that put a load on WM (Ben-Shachar et al., 2004; Fiebach et al., 2005; Friederici, Meyer, & von Cramon, 2000). The minimal contrast of predictability [who-whs] activated a variety of additional regions from Broca’s area, including the left STG, left vPCS, right PHG, and right temporal regions. Many of these regions have been previously activated by displacement (Ben-Shachar et al., 2003; Ben-Shachar et al., 2004; Fiebach et al., 2005; Just et al., 1996) and the others have been activated by dependency distance (right PHG; (Santi & Grodzinsky, 2007b).
The results from the minimal contrasts demonstrate that the absence of activation in Broca’s area in the more conservative conjunction analyses, other than for Predictability, was not due to a weak effect that simply got washed out. Each of the minimal contrasts generated activation clusters, but none were located in Broca’s area. The distinction between the novel contrasts (Dependency, Displacement) and the Predictability contrast in Broca’s area provides a novel angle in uncovering the functional role of this region. Important insight into the particular aspects of sentence processing that this region is sensitive to is critical to advancing our understanding of the linguistic selectivity of this region and ultimately the computations undertaken. Critically, the results from this study, in addition to others, have shown that the critical sentence comprehension process in Broca’s area, if tied to WM storage, is not general. This process demonstrates specificity to predicted displacements with a distance metric respecting the number of intervening verbal arguments or NPs.
Supplementary Material
Acknowledgments
This work was supported by NIH (grant #000494), SSHRC (standard grant #410-2009-0431), and Canada Research Chairs Canada Research Chairs, Natural Sciences and Engineering Research Council. The authors would like to thank Jon Nissenbaum, Isabelle Deschamps, and Katie Clark for their thoughtful comments and assistance during stimuli construction.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
For simplicity of presentation, we suppress the representation of vacuous movement from subject position. This suppression will have no consequences in the present context.
Roughly speaking, A’-positions (or “non-argument” positions) are distinct from A(rgument)-positions in that NPs in them cannot be assigned a thematic role directly. NPs in A’-positions receive thematic roles indirectly, via a link with a position, to which a thematic role is directly assigned. Normally, if an NP is in an A’-position (i.e., it is a “filler” in psychological talk), it is linked to an empty position (a “gap”), which is the direct recipient of the relevant thematic role. This role is then transmitted to the filler NP.
Note that adjunct parasitic gaps are not licensed by A-movement. Cf. the contrast between which paper did you file [gap--] [before reading [gap--]]? To *The man was pushed [gap--] [before killing [gap--].
For reasons related to making the stimuli pragmatically equivalent across conditions, this contrast also contains a transitive vs. intransitive verb distinction. Based on aphasia (Shapiro, Gordon, & Killackey, 1993) and neuroimaging (Ben-Shachar, Hendler, Kahn, Ben-Bashat, & Grodzinsky, 2003) results, however, there is no reason to expect an effect of a transitive vs. intransitive verb in Broca’s area, which is our main interest.
(i) [filler Which woman] did every tired student invite [gap--] after meeting [p-gap--]? (ii) [filler Which woman] did every tired student invite [gap--] after meeting her?In analogy to Doron’s and Sharvit’s sentences, (i) is ambiguous between a single- and multiple-individual readings – there could be one or more women associates with all the tired students; however (ii) only allows for a single-individual reading – only one woman is associated with all the tired students.
Alternative methods for calculating probability values from these maps exist (Eickhoff et al., 2007). The basis for the current method was to integrate both information about the size of the cluster and the distribution of probability values in one calculation.
Other studies also activated Broca’s region, but we are not sure about how indicative their results are: two studies featured uneven contrasts, that compared passive stimuli to Raising constructions, to other passive stimuli in which lexical changes were made, and to active sentences (Dapretto & Bookheimer, 1999; Grodzinsky, Shapiro, & Swinney, 2000; Wartenburger, Heekeren, Burchert, De Bleser, & Villringer, 2003)(Dapretto & Bookheimer, 1999; Wartenburger, Heekeren, Burchert, De Bleser, & Villringer, 2003); a third is briefly reported without sufficient details to make further comment (Grodzinsky et al., 2000)(Caplan, 2000).
The distinction between complex verbal arguments and NPs is captured by the following sentence “The brother of John kicked Mary”. The complex NP “The brother of John” is the subject (ie, one argument of the verb), and is composed of two NPs (the brother and John).
The lines representing the two dependencies in Table 2 are for exposition purposes and do not represent a commitment to any particular analysis. Specifically, we are not committed to an explicit relationship between the wh-phrase and the parasitic gap.
References
- Amunts K, Schleicher A, Börgel U, Mohlberg H, Uylings HBM, Zilles K. Broca's region revisited: Cytoarchitecture and intersubject variability. Journal of Comparative Neurology. 1999;412(2):319–341. doi: 10.1002/(sici)1096-9861(19990920)412:2<319::aid-cne10>3.0.co;2-7. [DOI] [PubMed] [Google Scholar]
- Avrutin S. Weak syntax. In: Grodzinsky Y, Amunts K, editors. Broca's Region. New York: Oxford University Press; 2006. pp. 49–62. [Google Scholar]
- Belin P, Zatorre R, Hoge R, Evans AC, Pike B. Event-related fMRI of the auditory cortex. NeuroImage. 1999;10(4):417–429. doi: 10.1006/nimg.1999.0480. [DOI] [PubMed] [Google Scholar]
- Ben-Shachar M, Hendler T, Kahn I, Ben-Bashat D, Grodzinsky Y. The neural reality of syntactic transformations: Evidence from functional magnetic resonance imaging. Psychological Science. 2003;14(5):433–440. doi: 10.1111/1467-9280.01459. [DOI] [PubMed] [Google Scholar]
- Ben-Shachar M, Palti D, Grodzinsky Y. Neural correlates of syntactic movement: Converging evidence from two fMRI experiments. NeuroImage. 2004;21(4):1320–1336. doi: 10.1016/j.neuroimage.2003.11.027. [DOI] [PubMed] [Google Scholar]
- Bornkessel I, Zysset S, Friederici A, Von Cramon DY, Schlesewsky M. Who did what to whom? The neural basis of argument hierarchies during language comprehension. Neuroimage. 2005;26(1):221–233. doi: 10.1016/j.neuroimage.2005.01.032. [DOI] [PubMed] [Google Scholar]
- Caplan D, Alpert N, Waters G. PET studies of syntactic processing with auditory sentence presentation. NeuroImage. 1999;9(3):343–351. doi: 10.1006/nimg.1998.0412. [DOI] [PubMed] [Google Scholar]
- Caplan D, Alpert N, Waters G, Olivieri A. Activation of Broca's area by syntactic processing under conditions of concurrent articulation. Human Brain Mapping. 2000;9(2):65–71. doi: 10.1002/(SICI)1097-0193(200002)9:2<65::AID-HBM1>3.0.CO;2-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caplan D, Futter C. Assignment of thematic roles to nouns in sentence comprehension by an agrammatic patient. Brain and Language. 1986;27(1):117–134. doi: 10.1016/0093-934x(86)90008-8. [DOI] [PubMed] [Google Scholar]
- Caplan D, Waters GS. Verbal working memory and sentence comprehension. Behavioral and Brain Sciences. 1999;22(1):77–126. doi: 10.1017/s0140525x99001788. [DOI] [PubMed] [Google Scholar]
- Chomsky N. On wh-movement. In: Culicover P, Wasow T, Akmajian A, editors. Formal Syntax. New York: Academic Press; 1977. [Google Scholar]
- Cooke A, Zurif EB, DeVita C, Alsop D, Koenig P, Detre J, et al. Neural basis for sentence comprehension: Grammatical and short-term memory components. Human Brain Mapping. 2001;15(2):80–94. doi: 10.1002/hbm.10006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dapretto M, Bookheimer SY. Form and content: Dissociating syntax and semantics in sentence comprehension. Neuron. 1999;24(2):427–432. doi: 10.1016/s0896-6273(00)80855-7. [DOI] [PubMed] [Google Scholar]
- Dehaene-Lambertz G, Dehaene S, Anton J-L, Campagne A, Ciuciu P, Dehaene G, et al. Functional segregation of cortical language areas by sentence repetition. Human Brain Mapping. 2006;27(5):360–371. doi: 10.1002/hbm.20250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doron E. On the syntax and semantics of resumptive pronouns. Texas Linguistics Forum. 1982;19:1–48. [Google Scholar]
- Drai D, Grodzinsky Y. A new empirical angle on the variability debate: Quantitative neurosyntactic analyses of a large data set from Broca’s Aphasia. Brain and Language. 2006;96(2):117–128. doi: 10.1016/j.bandl.2004.10.016. [DOI] [PubMed] [Google Scholar]
- Eickhoff SB, Paus T, Caspers S, Grosbras M, Evans AC, Zilles K, et al. Assignment of functional activations to probabilistic cytoarchitectonic areas revisited. NeuroImage. 2007;36:511–521. doi: 10.1016/j.neuroimage.2007.03.060. [DOI] [PubMed] [Google Scholar]
- Engdahl E. Linguistics and Philosophy. Dordrecht, Holland and Boston, USA: D. Reidel; 1983. Parasitic gaps; pp. 35–45. [Google Scholar]
- Fiebach CJ, Schlesewsky M, Lohmann G, von Cramon DY, Friederici AD. Revisiting the role of Broca's area in sentence processing: Syntactic integration versus syntactic working memory. Human Brain Mapping. 2005;24(2):79–91. doi: 10.1002/hbm.20070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forman SD, Cohen JD, Fitzgerald M, Eddy WF, Mintun MA, Noll DC. Improved assessment of significant activation in functional magnetic resonance imaging (fMRI): use of a cluster-size threshold. Magnetic Resonance in Medicine. 1995;33(5):636–647. doi: 10.1002/mrm.1910330508. [DOI] [PubMed] [Google Scholar]
- Frazier L, Clifton C, Randall J. Filling gaps: Decision principles and structure in sentence comprehension. Cognition. 1983;13(2):187–222. [Google Scholar]
- Friederici AD, Fiebach CJ, Schlesewsky M, Bornkessel ID, Von Cramon DY. Processing linguistic complexity and grammaticality in the left frontal cortex. Cerebral Cortex. 2006;16(12):1709–1717. doi: 10.1093/cercor/bhj106. [DOI] [PubMed] [Google Scholar]
- Friederici AD, Meyer M, von Cramon DY. Auditory language comprehension: an event-related fMRI study on the processing of syntactic and lexical information. Brain and Language. 2000;74(2):289–300. doi: 10.1006/brln.2000.2313. [DOI] [PubMed] [Google Scholar]
- Friedmann N. Traceless relatives: Agrammatic comprehension of relative clauses with resumptive pronouns. Journal of Neurolinguistics. 2008;21(2):138–149. [Google Scholar]
- Friedmann N, Gvion A. Sentence comprehension and working memory limitation in aphasia: A dissociation between semantic-syntactic and phonological reactivation. Brain and Language. 2003;86(1):23–39. doi: 10.1016/s0093-934x(02)00530-8. [DOI] [PubMed] [Google Scholar]
- Friedmann N, Shapiro LP. Agrammatic comprehension of simple active sentences with moved constituents: Hewbrew OSV and OVS structures. Journal of Speech language and Hearing Research. 2003;46(2):288–297. doi: 10.1044/1092-4388(2003/023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibson E. Linguistic complexity: Locality of syntactic dependencies. Cognition. 1998;68(1):1–76. doi: 10.1016/s0010-0277(98)00034-1. [DOI] [PubMed] [Google Scholar]
- Goebel R, Esposito F, Formisano E. Analysis of functional image analysis contest (FIAC) data with Brainvoyager QX: From single-subject to cortically aligned group general linear model analysis and self-organizing group independent component analysis. Human Brain Mapping. 2006;27(5):392–401. doi: 10.1002/hbm.20249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goodglass H, Barton MI, Kaplan EF. Sensory modality and object-naming in aphasia. Journal of Speech and Hearing Research. 1968;11(3):488–496. doi: 10.1044/jshr.1103.488. [DOI] [PubMed] [Google Scholar]
- Gordon PC, Hendrick R, Johnson M. Memory interference during language processing. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2001;27(6):1–13. doi: 10.1037//0278-7393.27.6.1411. [DOI] [PubMed] [Google Scholar]
- Grewe T, Bornkessel I, Zysset S, Wiese R, Von Cramon DY, Schlesewsky M. The emergence of the unmarked: A new perspective on the language-specific function of Broca's Area. Human Brain Mapping. 2005;26(3):178–190. doi: 10.1002/hbm.20154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grillo N. Generalized Minimality: Syntactic Underspecification in Broca's Aphasia. Utrecht: Utrecht University; 2008. [Google Scholar]
- Grillo N. Generalized minimality: Feature impoverishment and comprehension deficits in agrammatism. Lingua. 2009;119:1426–1443. [Google Scholar]
- Grodzinsky Y. Language deficits and linguistic theory. Boston: Brandeis University; 1984. [Google Scholar]
- Grodzinsky Y. Agrammatic Comprehension of relative clauses. Brain and Language. 1989;37(3):480–499. doi: 10.1016/0093-934x(89)90031-x. [DOI] [PubMed] [Google Scholar]
- Grodzinsky Y. A Restrictive theory of agrammatic comprehension. Brain and Language. 1995;50(1):27–51. doi: 10.1006/brln.1995.1039. [DOI] [PubMed] [Google Scholar]
- Grodzinsky Y. The neurology of syntax: language use without Broca's area. Behavioral and Brain Sciences. 2000;23(1):1–21. doi: 10.1017/s0140525x00002399. [DOI] [PubMed] [Google Scholar]
- Grodzinsky Y, Amunts K. Broca's Region. New York: Oxford University Press; 2006. [Google Scholar]
- Grodzinsky Y, Shapiro L, Swinney D, editors. Language and the Brain: Representation and Processing. Academic Press; 2000. [Google Scholar]
- Grodzinsky Y, Wexler K, Chien YC, Marakovitz S, Solomon J. The breakdown of binding relations. Brain and Language. 1993;45(3):396–422. doi: 10.1006/brln.1993.1052. [DOI] [PubMed] [Google Scholar]
- Hagiwara H. The breakdown of Japanese passives and theta-role assignment principle in Broca's aphasics. Brain and Language. 1993;45(3):318–339. doi: 10.1006/brln.1993.1049. [DOI] [PubMed] [Google Scholar]
- Hale J. A probabilistic earley parser as a psycholinguistic model. Paper presented at the Proceedings of the 2nd North American Chapter of the Association for Computational Linguistics on Language Technologies.2001. [Google Scholar]
- Hoshi H. Theta-role assignment, passivization, and excorporation. Journal of East Asian Linguistics. 1994;3:147–178. [Google Scholar]
- Humphries C, Love T, Swinney D, Hickok G. Response of anterior temporal cortex to syntactic and prosodic manipulations during sentence processing. Human Brain Mapping. 2005;26(2):128–138. doi: 10.1002/hbm.20148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Just MA, Carpenter PA, Keller TA, Eddy WF, Thulborn KR. Brain activation modulated by sentence comprehension. Science. 1996;274(5284):114–116. doi: 10.1126/science.274.5284.114. [DOI] [PubMed] [Google Scholar]
- Levy R. Expectation-based syntactic comprehension. Cognition. 2008;106:1126–1177. doi: 10.1016/j.cognition.2007.05.006. [DOI] [PubMed] [Google Scholar]
- Makuuchi M, Bahlmann A, Anwander A, Friederici AD. Segregating the core computational faculty of human language from working memory. Proceedings of the National Academy of Sciences. 2009;106:8362–8367. doi: 10.1073/pnas.0810928106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mauner G, Fromkin VA, Cornell TL. Comprehension and acceptability judgments in agrammatism: Disruptions in the syntax of referential dependency. Brain and Language. 1993;45(3):304–370. doi: 10.1006/brln.1993.1050. [DOI] [PubMed] [Google Scholar]
- Nichols T, Brett M, Andersson J, Wager T, Poline J-B. Valid conjunction inference with the minimum statistic. Neuroimage. 2005;25(3):653–660. doi: 10.1016/j.neuroimage.2004.12.005. [DOI] [PubMed] [Google Scholar]
- Nissenbaum J. Investigations of covert phrase movement. Cambridge, MA: MITWPL; 2000. [Google Scholar]
- Noppeney U, Price CJ. An fMRI study of syntactic adaptation. Journal of Cognitive Neuroscience. 2004;16(4):702–713. doi: 10.1162/089892904323057399. [DOI] [PubMed] [Google Scholar]
- Phillips C. The real-time status of island phenomena. Language. 2006;82(4):795–8223. [Google Scholar]
- Rogalsky C, Hickok G. Selective attention to semantic and syntactic features modulates sentence processing networks in anterior temporal cortex. Cerebral Cortex. 2009;19:786–796. doi: 10.1093/cercor/bhn126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ross JR. Constraints on variables in syntax. MIT; 1967. [Google Scholar]
- Santi A, Grodzinsky Y. Taxing Working Memory with Syntax: Bi-hemispheric Modulations. Human Brain Mapping. 2007a;28(11):1089–1097. doi: 10.1002/hbm.20329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santi A, Grodzinsky Y. Working memory and syntax interact in Broca's area. Neuroimage. 2007b;37(1):8–17. doi: 10.1016/j.neuroimage.2007.04.047. [DOI] [PubMed] [Google Scholar]
- Santi A, Grodzinsky Y. fMRI adaptation dissociates syntactic complexity dimensions. NeuroImage. 2010;51:1285–1293. doi: 10.1016/j.neuroimage.2010.03.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shapiro LP, Gordon B, Killackey J. Verb-argument structure processing in complex sentences in Broca's and Wernicke's aphasia. Brain and Language. 1993;45(3):423–447. doi: 10.1006/brln.1993.1053. [DOI] [PubMed] [Google Scholar]
- Sharvit Y. resumptive pronouns in relative clauses. Natural Language & Linguistic Theory. 1999;17(3):587–612. [Google Scholar]
- Shlonsky U. Resumptive pronouns as a last resort. Linguistic Inquiry. 1992;23(3):443–468. [Google Scholar]
- Smith EE, Jonides J. Storage and executive processes in the frontal lobes. Science. 1999;283(5408):1657–1661. doi: 10.1126/science.283.5408.1657. [DOI] [PubMed] [Google Scholar]
- Stowe LA, Paans AMJ, Wijers AA, Zwarts F, Mulder G, Vaalburg W. Sentence comprehension and word repetition: A positron emission tomography investigation. Psychophysiology. 1999;36:786–801. [PubMed] [Google Scholar]
- Stromswold K, Caplan D, Alpert N, Rauch S. Localization of syntactic comprehension by positron emission tomography. Brain and Language. 1996;52(3):452–473. doi: 10.1006/brln.1996.0024. [DOI] [PubMed] [Google Scholar]
- Talairach J, Tournoux P. Co-planar stereotaxic atlas of the human brain. New York: Thieme; 1988. [Google Scholar]
- Taraldsen T. The theoretical implications of a class of marked extraction. Pisa: Scuola Normale Superiore; 1981. [Google Scholar]
- Traxler MJ, Williams RS, Blozis SA, Morris RK. Working memory, animacy, and verb class in the processing of relative clauses. Journal of Memory and Language. 2005;53(2):204–224. [Google Scholar]
- Wagers MW, Phillips C. Multiple dependencies and the role of the grammar in real-time comprehension. Journal of Linguistics. 2009;45:395–433. [Google Scholar]
- Wartenburger I, Heekeren HR, Burchert F, De Bleser R, Villringer A. Grammaticality judgments on sentences with and without movement of phrasal constituents - an event-related fMRI study. Journal of Neurolinguistics. 2003;16:301–314. [Google Scholar]
- Yokoyama S, Watanabe J, Iwata K, Ikuta N, Haji T, Usui N, et al. Is Broca's area involved in the processing of passive sentences? An event-related fMRI study. Neuropsychologia. 2007;45:989–996. doi: 10.1016/j.neuropsychologia.2006.09.003. [DOI] [PubMed] [Google Scholar]
- Zurif E. The neuroanatomical organization of some features of sentence comprehension: Studies of real-time syntactic and semantic composition. Psychologica. 2003;32:13–24. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.