

Abstract
The prototypical representatives of the Euryarchaeota—the methanogens—are oxygen sensitive and are thought to occur only in highly reduced, anoxic environments. However, we found methanogens of the genera Methanosarcina and Methanocella to be present in many types of upland soils (including dryland soils) sampled globally. These methanogens could be readily activated by incubating the soils as slurry under anoxic conditions, as seen by rapid methane production within a few weeks, without any additional carbon source. Analysis of the archaeal 16S ribosomal RNA gene community profile in the incubated samples through terminal restriction fragment length polymorphism and quantification through quantitative PCR indicated dominance of Methanosarcina, whose gene copy numbers also correlated with methane production rates. Analysis of the δ13C of the methane further supported this, as the dominant methanogenic pathway was in most cases aceticlastic, which Methanocella cannot perform. Sequences of the key methanogenic enzyme methyl coenzyme M reductase retrieved from the soil samples before incubation confirmed that Methanosarcina and Methanocella are the dominant methanogens, though some sequences of Methanobrevibacter and Methanobacterium were also detected. The global occurrence of only two active methanogenic archaea supports the hypothesis that these are autochthonous members of the upland soil biome and are well adapted to their environment.
Keywords: upland soils, biological soil crust, methanogens, 16S rRNA, mcrA, δ13C
Introduction
The methanogenic Archaea (methanogens) produce nearly all biogenic methane on Earth. All known methanogens fall into the phylum Euryarchaeota, where they form six distinct orders: Methanomicrobiales, Methanocellales, Methanosarcinales, Methanobacteriales, Methanococcales and Methanopyrales. The pathways for generating CH4 in methanogens vary and include methanol and CO2 reduction, acetate cleavage, as well as CH4 production from a variety of methylated compounds. With the exception of marine sediments and hypersaline mats (Oremland, 1988), methanogenesis in natural systems is dominated by CO2 reduction (hydrogenotrophic methanogenesis) and acetate cleavage (aceticlastic methanogenesis; Conrad, 2005).
Although the biochemical machinery in methanogens varies with the pathway used, few functional genes, which encode for key enzymes in the production of methane, are common to all known methanogens (Hedderich and Whitman, 2006). Of those, the methyl coenzyme M reductase (mcrA) that is responsible for the last step in all methanogenic pathways—the reduction of the methyl group bound to coenzyme M—is typically used as functional gene marker for analysis of methanogenic communities along with the 16S ribosomal RNA (rRNA) gene (Luton et al., 2002).
Methanogens are strict anaerobes, and methanogenesis was shown to be fully suppressed upon exposure to oxygen in both pure culture and in soil (Fetzer et al., 1993; Fetzer and Conrad, 1993; Yuan et al., 2009). Nevertheless, the ability of methanogens in pure culture and soil to survive oxygen and desiccation stress conditions for several hours to days has also been shown (Kiener and Leisinger, 1983; Fetzer et al., 1993; Ueki et al., 1997; Liu et al., 2008; Ma and Lu, 2011). Although numbers of viable cells decreased with time, in many types of methanogens a core population was able to survive the stress and proliferate again.
In addition, the typical energetic yields for hydrogenotrophic and aceticlastic methanogenesis (ΔG0′ = −131 and −36 kJ, respectively) are lower than those obtained by iron, nitrate and sulphate reduction (ΔG0′ = −228.3, −163.2 and −152.2 kJ, respectively), and organisms performing these respirations pathways successfully compete with methanogens over hydrogen (Thauer et al., 1977, 1989). As a result, methanogenesis typically occurs only in highly reduced anoxic environments, where oxygen is absent and alternative electron acceptors, such as nitrate and sulphate, are depleted (Whitman et al., 2006).
In contrast to the notions presented above, Peters and Conrad (1995) demonstrated that several types of aerated soils could become methanogenic when incubated under anoxic conditions as slurry. Among these soils was a South African arid soil, which was stored in dry state at room temperature for nearly 10 years before incubation. Few other researchers have reported the occurrence of methanogenesis in aerated soils and/or the presence of methanogens. For example, West and Schmidt (2002) were able to induce methanogenesis in alpine soils when incubated under an H2/CO2-enriched atmosphere (but not under pure N2 atmosphere), and Teh et al. (2005) found methanogenesis occurring in tropical forest soils containing up to 19% O2. In three other cases, researchers measured methanogenic activity in pasture soil and were able to retrieve sequences affiliated to methanogens (Nicol et al., 2003; Gattinger et al., 2007; Radl et al., 2007). However, these authors have attributed most of this activity to the effect that livestock had on the soil by enriching it with nutrients from urine and manure and by introducing rumen microflora to the soil. Last, Poplawski et al. (2007) retrieved sequences of Methanosarcina and Methanocella in a Swedish barley field but did not attempt to measure potential methanogenic activity.
The discovery of potential methanogenesis in arid soils is of particular interest, as it is least expected in a soil that is dry and oxic throughout most of the year and where input of organic carbon is particularly low and grazing ruminants are scarce. Particularly for these reasons, if active methanogens are to be found in arid soils, they can demonstrate their autochthonous origin as soil microorganisms (rather than as allochthones being introduced by ruminants) and ability to survive in anoxic microniches of upland soils around the world.
Arid soils differ from soils in humid areas by their unique profile. Unlike most soils, soils in arid parts do not display a profile of layers representing different stages of bedrock erosion and a gradient of nutrient concentrations. Rather, the bulk soil is undifferentiated, eroded bedrock poor in nutrients, which is in turn covered by a densely populated mat of microorganisms, termed biological soil crust (BSC). The BSC is an association of microorganisms, which is formed practically in any soil whose water budget restricts the development of higher plants. The primary members and first colonizers of BSCs are filamentous cyanobacteria, which aggregate the soil using exopolysaccharides. Fungi, microalgae and lichens are often also associated with mature BSCs and sometimes bryophytes as well (Belnap et al., 2003). An extensive literature exists on the abovementioned members of BSCs but studies (particularly molecular) on the prokaryotic inhabitants of this mat (other than cyanobacteria) are limited (Garcia-Pichel, 2002).
Our goals in this study were to compare the methanogenic potential of different aerated soils collected globally and, more importantly, to uncover the identity of the active methanogens in these soils and the potential methanogenic pathways. We focused primarily on dryland environments and collected samples not only from Israel, Australia and Utah, but also from a forest and a meadow in Germany for comparison. We screened the soil samples for methanogenic potential, and attempted to decipher the methanogenic pathways through stable isotopes analysis and by the use of an inhibitor for aceticlastic methanogenesis. We then analyzed the identity of the archaeal populations in these soils and quantified their sizes through specific quantitative PCR (qPCR) assays.
Materials and methods
Soil samples and soil characterization
Soil samples were collected from several sites around the world as listed in Table 1. The sampling done in the Negev Desert, Israel, comprises our most comprehensive sampling. The sites were previously described by Angel and Conrad (2009). Samples were taken from the BSC where it existed, from 0 to 10 cm and from 10 to 20 cm depth. Apart from the Negev Desert samples, samples from Utah and Australia were also from dryland environments, whereas the samples obtained in Germany were from temperate forest and grassland. The gravimetric water content of the samples was determined before all further analyses.
Table 1. Characteristics of the study sites and samples and methanogenic potentials.
| Location | Site | Soil type (USDA)a | Sampling layer | pH | Corg, Ntot (%) | Methane production rate (nmol gdw−1 d−1) | Lagb (days) | R2 | Inhibited methane production rate (nmol gdw−1 d−1)c | Lag (days) | R2 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Avdat, Negev Plateau, Israel | Natural field—intershrub 30°47′N 34°45′E | Silt loam | BSC | 8.3 | 0.46, 0.30 | 3081.6 | 20 | 0.99 | 629.02 | 24 | 0.96 |
| 0–10 cm | 0.33, 0.01 | 1.03 | 24 | 0.90 | |||||||
| 10–20 cm | 0.38, 0.01 | 0.46 | 23 | 0.94 | |||||||
| -‘-Shrub 30°47′N 34°45′E | Silt loam | BSC | 8.4 | 0.66, 0.05 | 3710.73 | 16 | 0.96 | 1793.76 | 21 | 0.96 | |
| 0–10 cm | 0.34, 0.01 | 1.54 | 22 | 0.93 | |||||||
| 10–20 cm | 0.26, BLD | 1.23 | 23 | 0.82 | |||||||
| Liman (constructed mini-catchment) 30°48′N 34°45′E | Silt loam | BSC | 7.9 | 2.00, 0.15 | 5657.59 | 11 | 0.99 | 3471.14 | 15 | 1.00 | |
| 0–10 cm | 1.27, 0.09 | 540.95 | 16 | 1.00 | |||||||
| 10–20 cm | 0.96, 0.05 | 224.6 | 16 | 1.00 | |||||||
| Agricultural field 30°52′N 34°48′E | Loam | 0–10 cm | 8.5 | 0.28, BLD | 0.62 | 3 | 0.71 | ||||
| 10–20 cm | 0.25, 0.03 | BLD | |||||||||
| Arava Valley South, Israel | Wadi (dry riverbed) 29°58′N 35° 05′ E | Sand | BSC | 8.4 | 0.98, BLD | 4.39 | 22 | 0.99 | |||
| 0–10 cm | 0.06, BLD | BLD | |||||||||
| 10–20 cm | 0.08, BLD | BLD | |||||||||
| Agricultural field 30° 04′ N 35° 08′ E | Sand | 0–10 cm | 8.5 | 0.38, 0.14 | BLD | ||||||
| 10–20 cm | 0.18, BLD | BLD | |||||||||
| North Utah, USA | Natural field 41°44′N 112°20′E | Silt loam | 0–10 cm | 6.5 | 4.71, 0.37 | 222.98 | 52 | 0.95 | 23.3 | 28 | 1.00 |
| 41°42′N 112°20′E | Sandy loam | 0–10 cm | 7.7 | 2.03, 0.11 | 138.35 | 13 | 0.93 | 49.53 | 26 | 0.99 | |
| 41°40′N 113°10′E | Silt loam | 0–10 cm | 8.2 | 2.61, 0.14 | 3.49 | 19 | 0.99 | ||||
| 41°51′N 113°05′E | Silt loam | 0–10 cm | 8.1 | 2.63, 0.16 | 8.98 | 62 | 0.94 | ||||
| Nifold Plain, Australia | Savannah −14°45′N 144°04′E | Silt loam | 0–10 cm | 4.8 | 0.72, 0.10 | 11.61 | 59 | 0.91 | |||
| −14°45′N 144°04′E | Loamy sand | 0–10 cm | 5 | 0.35, 0.43 | 374.5 | 38 | 0.93 | 62.88 | 37 | 0.96 | |
| −14°39′N 143°58′E | Loamy sand | 0–10 cm | 3.9 | 1.21, 0.10 | 0.63 | 38 | 0.97 | ||||
| Giessen, Germany | Meadow 50°32′N 8°41′E | Silt loam | 0–7.5 cm | 5.9 | 4.24, 0.40 | 1860.07 | 33 | 0.92 | 305.48 | 27 | 0.97 |
| 7.5–15 cm | 6.0 | 3.48, 0.35 | 1428.4 | 42 | 0.85 | 171.91 | 37 | 0.98 | |||
| Marburg, Germany | Forest 50°48′N 8°48′E | Sandy loam | 0–10 cm | 3.5 | 20.83, 0.92 | 1.2 | 55 | 0.99 | |||
| 0–10 cm | 4.0 | 20.07, 0.90 | 1.58 | 62 | 0.98 |
Abbreviations: BLD, below limits of detection; BSC, biological soil crust.
Initiation of linear methane production (extrapolated from linear regressions).
Inhibition of acetoclastic methanogenesis using 2% CH3F (see Materials and methods).
Incubation conditions and gas measurements
For determining the methanogenic potential of the soils, the samples were incubated under wet anoxic conditions, as supposed to occur after flooding: 5 g of soil sample was placed in a 26-ml glass tube, supplemented with 5 ml of sterile distilled deionized water and the tube was capped with a butyl rubber stopper (cleaned by boiling in water before use). The headspace was repeatedly evacuated and flushed with N2 for 10 min and left with a 0.5 bar overpressure. Each sample was set up in triplicate and incubated at 25 °C in the dark for 48 days. Samples that demonstrated methanogenic activity were later incubated again using fresh soil, but supplemented with 2% methyl fluoride (CH3F) to inhibit aceticlastic methanogenesis (Janssen and Frenzel, 1997). Gas samples were taken from the tube headspaces every 6 days using a gas-tight pressure-lok syringe (Vici, Baton Rouge, LA, USA) and analyzed immediately. Methane and CO2 concentrations were analyzed using a gas chromatograph equipped with a methanizer (Ni-catalyst at 350 °C, Chrompack, Varian Deutschland GmbH, Darmstadt, Germany) and a flame ionization detector (Shimadzu Deutschland, Duisburg, Germany).
Analysis of stable carbon isotopes
The carbon isotope ratios 13C:12C in the CH4 and CO2 were determined using a gas chromatograph combustion isotope ratio mass spectrometer against the Vienna Pee Dee Belemnite (V-PDB) standard, as described previously (Conrad et al., 2009). Isotopic calculations and estimation of the approximate fraction of hydrogenotrophic methanogenesis of the total methanogenesis were done after Conrad (2005). Briefly, the fraction of the newly formed CH4 between two time points is given by:
where δ1, δ2 and δn are the isotopic signatures of the CH4 at times 1 and 2 and of the newly formed, respectively, whereas fn is the fraction of the newly formed CH4 between times 1 and 2.
The apparent fractionation factor for the conversion of CO2 to CH4 is given by:
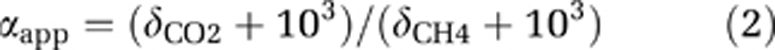 |
where δCO2 and δCH4 are the isotopic signatures of the carbon in CO2 and CH4, respectively.
For convenience, ɛ is often used instead of α. The two can be easily converted through:
The relative fraction of H2/CO2-derived CH4 in the total generated CH4 was determined by:
where δma and δmc are the specific isotopic signatures of the carbon in CH4 produced solely from acetate and H2/CO2, respectively. The δ13C in the organic matter was analyzed at the Institute for Soil Science and Forest Nutrition (IBW) at the University of Göttingen, Göttingen, Germany, using an elemental analyzer coupled to a mass spectrometer. Measurements were done before and after acidification, the difference being due to carbonate (Nüsslein et al., 2003).
Slurry sampling
After incubations were completed, the tubes were opened in an anoxic glove box, and ∼1 g of slurry was sampled for nucleic acid extraction, immediately frozen in liquid nitrogen and stored at −80 °C until analysis.
Nucleic acid extraction, RNA purification and complementary DNA synthesis
Total nucleic acids were extracted by disrupting 0.5 g of soil or slurry sample in a Lysing Matrix E tube (MP Biomedicals) in the presence of phosphate buffer, 10% SDS solution and phenol using a FastPrep-24 bead beater (MP Biomedicals, Heidelberg, Germany). The process was repeated three times using fresh buffers and phenol, and the supernatant was collected. The extractant was then purified using standard phenol/chloroform/isoamyl alcohol and chloroform/isoamyl alcohol purification, and the supernatant was transferred to Non-Stick RNase-free Microfuge Tubes (Ambion, Applied Biosystems, Darmstadt, Germany). The nucleic acids were precipitated using 20% polyethylene glycol and 5 μg of glycogen (Ambion), washed once with ice-cold 75% EtOH and resuspended in low TE buffer.
In cases in which the extractant had colour, it was further purified using MicroSpin S-200 HR (GE Healthcare, Freiburg, Germany). Ten microliters of the extractant was diluted in 90 μl of low TE buffer to be used as DNA template. For complementary DNA synthesis from all RNA templates, 50 μl of nucleic acid extractant was digested with TURBO DNase (Ambion) and later purified using RNeasy MinElute Cleanup Kit (Qiagen, Hilden, Germany). Complete DNA removal was verified by failure to obtain PCR amplification product with the purified RNA template, using the conditions described below. Complementary DNA was synthesized using ImProm-II Reverse Transcriptase (Promega, Mannheim, Germany) using 100 ng purified RNA and 0.5 μg μl−1 of random hexamer primers, as described by the manufacturer. The resulting complementary DNA product was used directly as a template for consecutive PCR reactions. PCR amplifications were performed identically for both DNA and complementary DNA templates. For a detailed description of the procedure, see the Supplementary Information.
Primers and probes used in this study
Table 2 summarizes the primers and probes used in this study to monitor and quantify the archaeal populations. For sequence affiliation, oligonucleotide design and evaluation, and reconstruction of phylogenetic trees, the ARB software package was used (Ludwig et al., 2004). Analysis of 16S rRNA gene sequences and oligonucleotide design was done using the Silva database (Pruesse et al., 2007), whereas for analyses of mcrA sequences a specific ARB database was built. The mcrA database comprised 5200 translated nucleic acid sequences, which were obtained from EMBL (http://www.ebi.ac.uk/embl/), aligned using Promals (Pei and Grishin, 2007) and manually corrected. The mcrA ARB database is available at: http://www.mpi-marburg.mpg.de/downloads/conrad/. The qPCR primers and probe targeting the 16S rRNA gene of Methanocella were designed using Primrose (Ashelford et al., 2002).
Table 2. Primers and probes used in this study.
| Oligo. namea | Target | Oligo. sequence (5′-3′) | Positionb | GC (%) | Tmc | Amplicon size | Essay | Reference |
|---|---|---|---|---|---|---|---|---|
| ARCH109 - F | Archaea 16S rRNA gene | AHDGCTCAGTAACACRT | 109–125 | 42.2 | 56 | TRFLP/Sequencing | Miyashita et al. (2009) | |
| ARCH934 - R | GTGCTCCCCCGCCAATTCCT | 915–934 | 65 | 68 | 826 | Grosskopf et al. (1998) | ||
| U1406 - R | ACGGGCGGTGTGTRC | 1392–1406 | 70 | 63 | 1298 | qPCR std. | Amann et al. (1995) | |
| ARCH364 - F | CGGGGYGCASCAGGGGCGAA | 364–383 | 75–80 | 75 | 553 | qPCR | Burggraf et al. (1997) | |
| MSL812 - F | Methanosarcinales 16S rRNA gene | GTAAACGATRYTCGCTAGGT | 812–831 | 40–50 | 62 | |||
| MSL860 - P | AGGGAAGCCGTGAAGCGARCC | 860–880 | 62–67 | 70 | qPCR | Yu et al. (2005) | ||
| MSL1159 - R | GGTCCCCACAGWGTACC | 1143–1159 | 65 | 63 | 354 | |||
| Mcl282 – F | Methanocella 16S rRNA gene | ATCMGTACGGGTTGTGGG | 282–299 | 56–61 | 65 | |||
| Mcl609 – P | ATCCAGCGGCTTAACCGTTGGKCK | 609–632 | 54–63 | 72 | qPCR | This study | ||
| Mcl832 – R | CACCTAGCGRGCATCGTTTAC | 813–832 | 52–57 | 64 | 510 | |||
| 771F | Crenarchaeota 16S rRNA gene | ACGGTGAGGGATGAAAGCT | 753–771 | 53 | 63 | Ochsenreiter et al. (2003) | ||
| 957R | CGGCGTTGACTCCAATTG | 957–974 | 56 | 62 | 203 | qPCR | ||
| mlas-mod – F | Universal mcrA gene | GGYGGTGTMGGDTTCACMCARTA | 976–998a | 43–65 | 68 | Modified from: Steinberg and Regan (2008) | ||
| mcrA-rev - R | CGTTCATBGCGTAGTTVGGRTAGT | 1421–1444a | 42–54 | 66 | 469 | Sequencing/qPCR | Steinberg and Regan (2008) |
The following primer name suffixes are used: - F – forward primer, - R – reverse primer, - P – dual labelled probe.
Positions are based on the following: primers targeting the 16S rRNA gene - E.coli; primers targeting the mcrA gene - M. thermautotrophicus mcrA gene accession number: U10036 (following Steinberg and Regan, 2008);
Calculated using Nearest Neighbour method with OligoAnalyzer 3.1 (http://eu.idtdna.com/analyzer/Applications/OligoAnalyzer/).
PCR amplification
PCR reactions were conducted in triplicates of 25 μl, which were pooled to minimize reaction variability. Each PCR reaction contained 5 μl GoTaq Flexi 5 × Green Buffer (Promega), 1.5 m MgCl2, 20 mg BSA (Roche Diagnostics, Mannheim, Germany), 0.75 U of GoTaq DNA polymerase (Promega) and 1 μl of template. For amplification of the 16S rRNA gene, 0.5 μ of each primer was used, whereas for amplification of mcrA 0.8 μ was used. The following program was used for the amplification of the 16S rRNA gene: 94 °C for 4 min, followed by 28 cycles of 94 °C for 1 min, 52 °C for 30 s and 72 °C for 45 s, and a single step of final elongation at 72 °C for 10 min. For the amplification of mcrA, the following program was used: 94 °C for 4 min, followed by five touchdown cycles of 94 °C for 30 s, 60–1 °C for 45 s and 72 °C for 30 s, followed by 30 cycles of 94 °C for 30 s, 55 °C for 30 s and 72 °C for 30 s, and a single step of final elongation at 72 °C for 10 min. Following amplification, the PCR products were purified using GeneElute PCR purification Kit (Sigma-Aldrich, Munich, Germany).
TRFLP analysis
For terminal restriction fragment length polymorphism (TRFLP) analysis, the forward primer was labeled with the fluorescent dye 6-carboxyfluorescein. Approximately 200 ng of purified PCR products was digested overnight at 37 °C with 20 U of the restriction enzyme TaqI and 2 μl of 10 × Buffer TaqI (Fermentas, St. Leon-Rot, Germany). Following digestion, samples were desalted using SigmaSpin Post-Reaction Clean-up Columns (Sigma); aliquots of 3 μl were mixed with 10 μl of HiDi Formamide (Applied Biosystems, Darmstadt, Germany) and 0.3 μl of the internal DNA standard MapMarker1000 (BioVentures, Murfreesboro, TN, USA). The samples were denatured at 94 °C for 2 min and loaded into an ABI 3100 automated gene sequencer (Applied Biosystems) for separation of the TRFs. TRFLP data were retrieved by comparison with the internal standard using GeneScan 3.71 software (Applied Biosystems).
Cloning and sequencing and phylogenetic analysis
Samples with high methanogenic potential from the following locations were used for cloning of both 16S rRNA and mcrA genes: Avdat natural field and Liman: BSC; Giessen: 0–7.5 cm; and Nifold Plain 2 0–10 cm. Libraries were constructed using purified PCR products and pGEM-T Easy cloning kit (Promega). Double-coverage sequencing services were provided by ADIS (Cologne, Germany) or GATC (Konstanz, Germany) using the primers T7f and M13r targeting flanking regions of the insert. A total of 250 clones containing 16S rRNA gene insert and 78 clones containing mcrA gene inserts were analyzed. All sequences were deposited into the GenBank (http://www.ncbi.nlm.nih.gov) and received the following accession numbers: HQ268968–HQ269217 and HQ269218–HQ269295 (16S rRNA and mcrA, respectively). Raw sequence data were analyzed using SeqMan 4.05 software (DNAStar, Madison, WI, USA), where the primers and vector data were trimmed. The 16S rRNA gene sequences were aligned using the SINA Webaligner (http://www.arb-silva.de/aligner/) against the Silva 102 database. mcrA sequences were converted to amino-acid sequences and aligned against the ARB database described above. Reconstruction of a maximum likelihood phylogenetic trees was conducted with RAxML 7.04 using rapid hill climbing algorithm and with GTRMIX and PROTMIX-JTT evolutionary models for 16S rRNA and mcrA genes, respectively (http://sco.h-its.org/exelixis/software.html; Stamatakis, 2006).
qPCR Assays
qPCR assays were used to quantify the 16S rRNA gene copies of the general archaeal, crenarchaeal, Methanosarcina and Methanocella populations. All qPCR reactions were performed on an iCycler thermocycler equipped with a MyiQ detection system (Bio-Rad, Munich, Germany) and the data were analyzed using iQ5 Optical System software (Bio-Rad). For all assays, standards containing known numbers of DNA copies of the target gene were used. Standards were serially diluted and used for construction of calibration curves in each reaction. The standard for the general archaeal and the Methanosarcina assays was prepared from a pure culture of Methanosarcina thermophila according to Lueders and colleagues (2004). The standards for Crenarchaeota and Methanocella assays were prepared from clones containing 16S rRNA genes affiliated with either Crenarchaeota or Methanocella as a plasmid insert. Total Archaea and Crenarchaeota assays were based on SYBR Green (Ambion). Each reaction was 25 μl in volume and contained the following mixture: 12.5 μl SYBR Green JumpStart Taq ReadyMix, 3 or 2 m MgCl2 (total Archaea and Crenarchaeota assays, respectively) and 0.8 ng μl−1 BSA, 0.66 or 0.44 μ of each primer (total Archaea and Crenarchaeota assays, respectively), and 5 μl of template. For the two assays, the program used was: 94 °C for 4 min, followed by 40 cycles of 94 °C for 30 s, 66 or 63 °C for 30 s (total Archaea and Crenarchaeota assays, respectively), 72 °C for 30 s and 85 °C for 10 s for signal reading. The assays targeting the 16S rRNA gene of Methanosarcina and Methanocella were based on dual-labeled probes. Each reaction was 25 μl in volume and contained the following mixture: 12.5 μl JumpStart Taq ReadyMix, 4 m MgCl2, 0.8 ng μl−1 BSA (Ambion), 0.5 μ of each primer, 0.2 μ of the dual-labeled probe and 5 μl of template. For the two assays, the program used was: 94 °C for 4 min, followed by 40 cycles of 94 °C for 30 s and 60 °C for 60 s for annealing, extension and signal reading.
Data transformation and statistical analysis
Affiliation of TRFs to their respective archaeal taxa was done using in silico prediction of the restriction sites in the sequence data. For principal component analysis, TRFLP data were transformed and standardized as described previously (Angel et al., 2010). Principal component analysis plots were computed using Canoco 4.53 and plotted using CanoDraw 4.12 (http://www.canoco.com).
Results
Methanogenic potential in upland soils
The methanogenic potential in upland soils was tested by incubating the samples under flooded conditions with anoxic gas headspace. Of the 27 soil samples tested, 22 produced at least some CH4 during the incubation (Table 1). We observed strong differences in the extent of the methanogenic potential of the soils, which were expressed in up to four orders of magnitude difference in daily production rates. Of the 22 samples, we classified 10 as having a strong methanogenic potential (highly methanogenic soils), meaning they had CH4 production rates in the hundreds or thousands nmol gdw−1 per day. In most incubations, CH4 was detected within 7–14 days, but a linear CH4 production began, on average, at day 21. Interestingly, CH4 production rates and lag times did not correlate with parameters such as precipitation, soil water content and organic matter, which would seem to be most important for supporting methanogenesis. For example, desert crust samples from the Negev Desert, Israel, had more than double the rate of potential CH4 production than the meadow samples. With respect to the desert samples, we observed that the BSC layer was the only truly active layer in the soil, whereas layers below it had a very low or no methanogenic potential. The only exception are the samples from the Liman (constructed desert minicatchment), whose soil periodically behaves like pond sediments when the Liman is flooded (after heavy rain), and also builds up much quicker than the native soil due to alluviation processes (as it is built to drain its surrounding area).
Stable isotope analysis of CH4 and CO2
In parallel to measuring potential CH4 production rates of the soils, we have also determined the isotopic signature (δ13C) of the carbon in the CH4 and CO2 being produced. This was done to estimate the proportion of each of the two main methanogenic pathways—hydrogenotrophic and aceticlastic—in our experiments. In addition, those samples that we classified as highly methanogenic were also incubated with CH3F to inhibit aceticlastic methanogenesis. This allowed us to specifically determine the isotopic signature of the CH4 produced through hydrogenotrophic methanogenesis (δmc).
The BSC samples from Israel showed a temporal trend during the incubation by which CH4 began with a light isotopic signature of the carbon (δ13C=approximately −57 on average) and became heavier with time (up to approximately −35 on average), whereas the isotopic signature of the carbon in the CO2 became slightly lighter over time starting at δ13C=approximately −9 and shifting toward approximately −15, on average (Table 3). A similar phenomenon with respect to CH4 but not to CO2 was observed for incubations of the samples from Utah and Germany, whereas the incubations from Australia showed a relatively stable isotopic signature in both CH4 and CO2 over time. Fractionation factors (ɛ) in all non-inhibited incubations were smaller than those in the inhibited incubations, but higher than those obtained for purely aceticlastic methanogenesis that typically range between ∼21 and 27 (Krzycki et al., 1987; Gelwicks et al., 1994), indicating a mixture of both pathways. To precisely determine the contribution of hydrogenotrophic and aceticlastic methanogenesis, knowledge of the exact isotopic signatures of the carbon in the CH4 produced in each pathway is required. One can, however, estimate the relative contribution of each pathway if the isotopic signature of the carbon in the CH4 (δ13C–CH4) produced in the inhibited incubations is used as the specific signature for CH4 produced from H2/CO2 alone (δmc) and the isotopic signature of the organic carbon in the soil is used to estimate the signature of CH4 produced from acetate. The measured δ13C of the organic carbon in the soil samples (which is the substrate for acetate) differed somewhat between sites but ranged from approximately −20.2 to −27.9. The fractionation factor for aceticlastic methanogenesis in pure cultures varies with respect to substrate concentration and can reach up to ɛ=∼25.6 when substrate is not limiting (Goevert and Conrad, 2009). As acetate concentrations were low in our incubations (relative to those used by Goevert and Conrad, 2009), one can expect a relatively high substrate turnover and assume a low fractionation factor (∼10). Using Equation 4 and these assumptions, the fractions of hydrogenotrophic methanogenesis in the system were estimated (Table 3). In all cases, except the samples from Australia, methanogenesis began with a dominance of hydrogenotrophic methanogenesis but later on dropped to about 10–25% of the CH4 being produced.
Table 3. Stable carbon isotope analysis of the methane and CO2.
|
δ13C-Corg
(‰) |
δ13C (‰) |
Apparent fractionation factor (ɛapp) |
δmaa (‰) |
Fraction of H2/CO2
methanogenesis |
|||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Non-acidified | Acidified | CH4 beginb | CH4 end | CO2 begin | CO2 end | ɛ begin | ɛ end | fH2 begin | fH2 end | ||
| Israel (Natural field) | −5.4 | −21.1 | −57.3±4.9 | −34.9±2.5 | −9.1±2.1 | −15.3±1.2 | 53±6 | 25±4 | −31.1 | 0.74 | 0.1 |
| Israel + CH3F | −66.3±3.2 | −68.7±0.6 | −7.5±1.67 | −12.2±1.1 | 61±3 | ||||||
| Utah (sample 1) | −26.4 | −26.6 | −74.4±6.8 | −44.4±3.4 | −24.8±1.8 | −24.9±0.6 | 54±10 | 27±4 | −36.6 | 1.00 | 0.25 |
| Utah + CH3F | −70.4±3.6 | −25.2±0.4 | 49±3 | ||||||||
| Australia (sample 2) | −20.6 | −20.4 | −69.6±4.4 | −65.4±2.7 | −20.0±0.1 | −16.8±0.7 | 34±7 | 46±6 | −30.4 | 0.72 | 0.67 |
| Australia + CH3F | −85.3±6.0 | −82.6±0.9 | −17.1±0.6 | −15.2±−6.2 | 76±3 | ||||||
| Germany (0–7.5 cm) | −28.3 | −28.2 | −80.1±2.9 | −51.9±11.0 | −22.2±0.8 | −22.7±0.8 | 63±4 | 30±11 | −38.2 | 0.95 | 0.27 |
| Germany + CH3F | −82.1±3.16 | −89.4±1.2 | −22.8±0.1 | −21.3±0.4 | 69±3 | ||||||
δma was calculated by assuming a ɛ=−10‰ fractionation from organic carbon (see text).
‘Begin' and ‘end' refer to the first third and last two thirds of the incubation period, respectively.
Molecular characterization of the methanogenic community
We characterized the archaeal community in the soil samples and incubated slurries through cloning and TRFLP profiling of the 16S rRNA and compared the preincubated with the post-incubated state of both inhibited and non-inhibited incubations. Surprisingly, the diversity of the methanogenic community in our samples was very low and nearly identical, although samples were obtained from different parts around the world. In all of the post-incubated samples, we identified only Methanosarcina- and Methanocella-related sequences along with a variety of crenarchaeal sequences, typically associated with aerated soils such as soil crenarchaeotic group 1.1b and occasionally sequences related to Thermoplasmatales (Figure 1). The latter two groups were detected as sole members of the archaeal communities in the samples before incubation, whereas none of the methanogens could be detected in our 16S rRNA clone libraries or TRFLP profiles. Inhibition with CH3F resulted in lower CH4 production rates but did not change the identity of the methanogenic community. Not only did we detect nothing but Methanosarcina and Methanocella as methanogenic members in our post-incubated samples, their within-group sequence diversity was itself very low. For example, the average sequence difference between Methanosarcina from Avdat BSC (Israel) and Giessen meadow soil (Germany) was only 1.8%±0.025 and that between the Methanocella members only 3.3%±0.1.
Figure 1.

Maximum likelihood phylogenetic tree based on aligned partial archaeal 16S rRNA gene sequences. Sequences were aligned against the SILVA 102 database using the SINA aligner, and the tree was calculated with RAxML 7.04 using rapid hill climbing algorithm and GTRMIX evolutionary model. Bootstrap values above 50% (out of 100 trials) are displayed next to the nodes. Shaded clusters contain sequences that were only detected in the preincubated soil samples, whereas sequences from the post-incubated slurries cluster both into the shaded clusters and into the shaded clusters with diagonal lines.
TRFLP analysis
We used TRFLP profiling to decipher relative changes in the archaeal community from in situ to post-incubated state and with respect to the effect of inhibition with CH3F. A total of 67 replicated TRFLP profiles, representing the soil samples before incubation and the inhibited and uninhibited slurry incubations, are summarized in Figure 2. As expected, all preincubated samples (green circles) had similar 16S rRNA profiles on both DNA and RNA levels and clustered very tightly with respect to the primary principal component, where methanogens showed the highest loadings. Most post-incubated samples (red and blue circles) were found on the right to the ordinate, indicating methanogenic activity, and their approximate CH4 production rates can be deduced from the fitted isolines (LOESS fit). Those samples that did not demonstrate potential methanogenic activity remained close to the preincubated samples in these plots. Methanosarcina was more abundant than Methanocella in most methanogenic samples on the DNA level and seemed to have been overwhelmingly dominant on the RNA level. It seems that the inhibition with CH3F, while reducing methanogenic activity, did not have an effect on the composition of the methanogenic community.
Figure 2.

Principal component analysis plots of the archaeal community as deciphered from the 16S rRNA gene TRFLP profiles. Circles indicate individual samples. The relative abundance of each of the archaeal types can be estimated from the perpendicular projection of each sample to the individual vectors, whereas the length of each vector indicates the variance (range of relative abundance values) of its respective archaeal type. CH3F was used to inhibit aceticlastic methanogenesis. Fitted methane production rates are shown as isolines; (a) DNA TRFLP profiles and (b) rRNA TRFLP profiles.
Molecular characterization using the mcrA marker gene
Our inability to detect methanogens in the 16S rRNA gene TRFLP profiles and clone libraries of the preincubated soil samples is most likely the result of their low relative abundance compared with the populations of Crenarchaeota and Thermoplasmatales. Any community member that comprises 1% or less of the community would be undetected in TRFLP profiles (due to the method's threshold and standardization procedures) and also in the clone libraries (considering our sequencing efforts). We therefore performed an analysis of the methanogenic community in both the preincubated soil samples and the post-incubated slurries using the mcrA functional gene. Although we were unable to amplify the gene in all our samples (most likely due to the low abundance of methanogens in the samples before incubation), we did obtain PCR products from at least one sample at each site. The phylogenetic affiliation of these sequences is presented in Figure 3. The majority of the mcrA sequences belonged to either Methanosarcina or Methanocella, confirming our observation from the analysis of the 16S rRNA genes and transcripts of the post-incubated slurries. We did, however, detect several sequences belonging to Methanobrevibacter and Methanobacterium, which were not detected in the post-incubated samples using the 16S rRNA gene analysis.
Figure 3.

Maximum likelihood phylogenetic tree based on aligned partial amino-acid sequences of the mcrA. Amino-acid composition was deduced from DNA sequences, and the tree was calculated with RAxML 7.04 using rapid hill climbing algorithm and PROTMIX-JTT evolutionary model. Bootstrap values above 50% (out of 100 trials) are displayed next to the nodes. Shaded clusters contain sequences that were only detected in the preincubated soil samples, whereas sequences from the post-incubated slurries cluster both into the shaded clusters and into the shaded clusters with diagonal lines clusters.
Quantification of the methanogenic community
To determine the quantitative effect of our incubations on the different members of the archaeal community, we used qPCR and quantified the total archaeal, the crenarchaeal communities and the two methanogens that were identified in the phylogenetic analysis. Quantification was performed on the soil samples before incubation and the post-incubated slurries. Copy numbers of the archaeal 16S rRNA gene in the highly methanogenic samples rose between 0.5 and 2 orders of magnitude in the post-incubated slurries compared with the preincubated soils and were moderately correlated to CH4 production rates (Figure 4a). Both methanogen types were undetected in the preincubated soil samples and non-methanogenic samples (that is, they were below 103 copies) but reached up to 109 copies gdw−1 after the incubation. Copy numbers of Methanosarcina and Methanocella 16S rRNA gene were within one order of magnitude from each other. As both methanogen types contain three copies of the 16S rRNA gene in their genomes, the actual number of methanogenic cells is a third of the gene copy numbers displayed in the figure. Copy numbers of both methanogens were positively correlated to CH4 production rates, but Methanosarcina was better correlated (Figures 4c and d). In contrast, gene copy numbers of Crenarchaeota did not change in most samples between the pre- and post-incubated states and showed no correlation to CH4 production rates (Figure 4b).
Figure 4.

Archaeal 16S rRNA gene copy numbers are quantified using qPCR plotted against potential methane production rates. Both preincubated soil samples as well as post-incubated slurries are shown in these plots. The preincubated samples as well as the slurries that showed no methanogenic potential are positioned at 0 on the x axis.
Discussion
When the domain Archaea was first described by Woese and Fox (1977), it was thought to be an ancient lineage of prokaryotes (hence the name), which evolved early in the evolutionary history in a world which was hotter and whose atmosphere and hydrosphere were much more reduced. This notion emerged from the fact that at that time the known archaea were predominantly extremophiles or anaerobes (Woese et al., 1990; Takai and Horikoshi, 1999). In the past two decades though, environmental surveys using molecular screening tools have detected numerous archaeal sequences in virtually all mesothermic environments, including aerated soils, ocean water and freshwater (DeLong, 1992; Bintrim et al., 1997; Schleper et al., 1997). It is mainly based on these studies that Archaea are now considered as cosmopolitan as Bacteria, adapted to their ecological niches and perform a variety of metabolic activities (DeLong, 1998; Schleper et al., 2005; Leininger et al., 2006; Auguet et al., 2010).
Here, we have shown that methanogens are also ubiquitous in mesophilic, aerated soils around the world. Through simple anoxic incubations with only water but no additional substrate added, nearly all the soil samples produced CH4 from degradation of soil organic matter. Many of our samples had very low methanogenic potential and long lag times, but some samples reacted very quickly and produced ample amounts of CH4 within a few days. Particularly intriguing is the fact that in the arid samples from Israel it was the topmost part of the soil—the BSC—which held nearly all of the methanogenic potential of the soil. Although it is true that BSCs contain most of the microbial biomass and activity in desert soils (Garcia-Pichel and Belnap, 2003), it is also the layer most exposed to oxygen. In fact, as BSCs are mostly formed by photosynthetic microorganisms, they can turn hyperoxic at parts during daytime when moist (Garcia-Pichel and Belnap, 1996). The existence of soils with such high methanogenic potential and fast response indicates that they are also active, at least at certain times, under natural conditions.
It is difficult to suggest from the data what determines the difference in methanogenic potential in the different soil samples. Important factors that might reduce the methanogenic potential are low quantity and quality of available organic matter (Wagner et al., 2005; Guérin et al., 2008), low soil pH (Phelps and Zeikus, 1984) and the availability of alternative electron acceptors (Peters and Conrad, 1996; Yao et al., 1999). Desert soils are typically neutral/alkaline, well buffered and are poor in nitrogen, iron and manganese, which could serve as alternative electron acceptors (Knight, 1991). Sulfate may be available in desert soils containing gypsum, but we found no evidence for substantial sulfate reduction in our incubations (data not shown). It is unclear to which extent the abiological characteristics of desert soil can explain the relatively high methanogenic potentials of some of our desert samples. However, biological activity in the BSCs that typically form on the surface of most desert soils may support methanogenesis through the supply of fresh organic matter from photosynthesis by cyanobacteria during daytime and the depletion of oxygen through respiration activity during nighttime (Garcia-Pichel and Belnap, 1996; Lange, 2003; Angel et al., 2011).
Similar to many natural environments, we found a mixture of both aceticlastic and hydrogenotrophic methanogenesis in our incubations (Conrad, 1999), but the specific proportion of each pathway varied between samples and even within certain samples as incubation proceeded. All incubations began with CH4 being produced mostly or entirely out of hydrogen and CO2, but in nearly all cases shifted to being predominantly aceticlastic as the incubation proceeded. This pattern is known from studies on methanogenesis in rice field soils (Roy et al., 1997), and is explained by the temporal change in thermodynamic conditions and differential expression of microbial activity upon flooding (Glissmann and Conrad, 2002; Conrad, 2002). The ratio of aceticlastic methanogenesis in these samples stabilized around 60–70%, which is very close to the ratio found in many natural environments in general (Conrad, 1999) and anoxically incubated upland soils in particular (Peters and Conrad, 1996). Methanogenesis in the samples from Australia, on the other hand, remained predominantly hydrogenotrophic throughout the incubation.
Although methanogenic potential thus seems to be a global trait of upland soils (albeit of different characteristics), we have recovered a remarkably low diversity of methanogens in our samples. Of the 30 known methanogenic genera, we found only 2 closely related genera in our post-incubated samples belonging to Methanosarcina and Methanocella, both types are of high ecological importance. Methanosarcina are, along with Methanosaeta, the only methanogens capable of performing aceticlastic methanogenesis and are therefore predominant methanogens in most natural environments (Liu and Whitman, 2008). Methanocella is a newly discovered genus, also globally distributed but colonizes rice roots in particular (Grosskopf et al., 1998). Methanocella was also shown to be of high importance as one of the primary CH4 producers in rice fields, probably feeding on fermentation products of root exudates (Lu and Conrad, 2005). Although phylogenetically closely related to Methanosarcina and similar in their cellular structure, Methanocella species can only perform hydrogenotrophic methanogenesis (Sakai et al., 2010).
In our samples, it appears that Methanosarcina was the dominant methanogens. Certainly, this stems from our analysis of methanogenic pathways, where in the latter part of the incubations aceticlastic methanogenesis accounted for roughly two-thirds of the newly formed CH4, but also generally speaking Methanosarcina had a higher relative abundance in the TRFLP profiles and their absolute abundance correlated better with CH4 production rates. Similarly, Methanosarcina and Methanocella were also the methanogenic types that were found by Nicol et al. (2003) in a pasture soil in Scotland and by Poplawski et al. (2007) in a barley field in Sweden, thus reinforcing our notion that these types are universal upland soil methanogens. In cases where heavy grazing was involved, the authors have also found Methanosarcina as dominant methanogens, but along with it other types that could be directly associated to rumen microflora (Gattinger et al., 2007; Radl et al., 2007). To our knowledge, there is only one molecular study explicitly targeting archaea in desert soils (Soule et al., 2009), detecting only Crenarchaeota of the group 1.1b in the native soil.
The phylogenetic analysis of the preincubated soils using mcrA revealed also the presence of a few other methanogens that are closely affiliated with Methanobrevibacter and Methanobacterium. None of these methanogens could be enriched in our incubations and it remains unclear whether these were viable cells. Nevertheless, their presence does imply that the diversity of the methanogenic community in these aerated soils is potentially wider.
The formation of biogenic CH4 when aerated soils are incubated anoxically testifies not only for the presence of active methanogens but also for the presence of many other anaerobic microorganisms. As CH4 is only an end product in a cascade of anaerobic degradation processes, other viable anaerobic microorganisms from fermenters to acetogens and syntrophs need to be present in the soil for it to be formed (Garcia et al., 2000; Stams and Plugge, 2009). The presence and potential activity of such anaerobes in upland soils have indeed been shown in the past (Kusel and Drake, 1994; Degelmann et al., 2009). It thus indicates that upland soils host not just methanogens but entire consortia of anaerobes capable, at least potentially, to carry a full anaerobic degradation pathway.
Methanogens of the type Methanosarcina and Methanocella appear to be autochthonous and universal members of the biome in aerated soils. Despite being exposed to constant oxygen flux when the soil is dry, a core population of these methanogens is able to survive and become rapidly active when incubated anoxically with water. The existence of a unique and viable upland soil methanogenic population is an indication for a natural niche for these microorganisms, yet their activity in field conditions still remains to be shown.
Acknowledgments
We thank Dr Claudia Kammann of Giessen FACE Experiment for providing the samples from Giessen (Germany); Professor Andreas Brune for sampling in Australia and Professor JC Ensign for sampling in Utah. The Zuckerberg Institute for Water Research of Ben-Gurion University of the Negev, Israel, for allowing the use of their facilities; the Arava Institute for Environmental Studies, Israel, for providing boarding services and administrative assistance; The Institute for Soil Science and Forest Nutrition (IBW) at the University of Göttingen, Germany, for determining the isotopic signature of the soil organic carbon; The Analytical Chemical Laboratory of the Philipps University, Marburg, for the CN analyses; and the Chair of the Physical Geography, Aachen, for analysis of the soil texture. RA was supported by a fellowship of the Max Planck Society.
Footnotes
Supplementary Information accompanies the paper on The ISME Journal website (http://www.nature.com/ismej)
Supplementary Material
References
- Amann R, Ludwig W, Schleifer K. Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiol Rev. 1995;59:143–169. doi: 10.1128/mr.59.1.143-169.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Angel R, Conrad R. In situ measurement of methane fluxes and analysis of transcribed particulate methane monooxygenase in desert soils. Environ Microbiol. 2009;11:2598–2610. doi: 10.1111/j.1462-2920.2009.01984.x. [DOI] [PubMed] [Google Scholar]
- Angel R, Matthies D, Conrad R. Activation of methanogenesis in arid biological soil crusts despite the presence of oxygen. PLoS One. 2011;6:e20453. doi: 10.1371/journal.pone.0020453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Angel R, Soares MIM, Ungar ED, Gillor O. Biogeography of soil archaea and bacteria along a steep precipitation gradient. ISME J. 2010;4:553–563. doi: 10.1038/ismej.2009.136. [DOI] [PubMed] [Google Scholar]
- Ashelford KE, Weightman AJ, Fry JC. PRIMROSE: a computer program for generating and estimating the phylogenetic range of 16S rRNA oligonucleotide probes and primers in conjunction with the RDP-II database. Nucleic Acids Res. 2002;30:3481–3489. doi: 10.1093/nar/gkf450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Auguet J, Barberan A, Casamayor EO. Global ecological patterns in uncultured Archaea. ISME J. 2010;4:182–190. doi: 10.1038/ismej.2009.109. [DOI] [PubMed] [Google Scholar]
- Belnap J, Buedel B, Lange OL.2003Biological soil crusts: characteristics and distributionIn: Belnap J, Lange OL (eds). Biological Soil Crusts Springer: Berlin, Germany; 3–30. [Google Scholar]
- Bintrim S, Donohue T, Handelsman J, Roberts G, Goodman R. Molecular phylogeny of archaea from soil. Proc Natl Acad Sci USA. 1997;94:277–282. doi: 10.1073/pnas.94.1.277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burggraf S, Huber H, Stetter KO. Reclassification of the crenarchaeal orders and families in accordance with 16S rRNA sequence data. Int J Syst Bacteriol. 1997;47:657–660. doi: 10.1099/00207713-47-3-657. [DOI] [PubMed] [Google Scholar]
- Conrad R. Contribution of hydrogen to methane production and control of hydrogen concentrations in methanogenic soils and sediments. FEMS Microbiol Ecol. 1999;28:193–202. [Google Scholar]
- Conrad R. Control of microbial methane production in wetland rice fields [Review] Nutr Cycl Agroecosyst. 2002;64:59–69. [Google Scholar]
- Conrad R. Quantification of methanogenic pathways using stable carbon isotopic signatures: a review and a proposal. Org Geochem. 2005;36:739–752. [Google Scholar]
- Conrad R, Claus P, Casper P. Characterization of stable isotope fractionation during methane production in the sediment of a eutrophic lake, Lake Dagow, Germany. Limnol Oceanogr. 2009;54:457–471. [Google Scholar]
- Degelmann DM, Kolb S, Dumont M, Murrell JC, Drake HL. Enterobacteriaceae facilitate the anaerobic degradation of glucose by a forest soil. FEMS Microbiol Ecol. 2009;68:312–319. doi: 10.1111/j.1574-6941.2009.00681.x. [DOI] [PubMed] [Google Scholar]
- DeLong EF. Archaea in coastal marine environments. Proc Natl Acad Sci. USA. 1992;89:5685–5689. doi: 10.1073/pnas.89.12.5685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeLong EF. Everything in moderation: archaea as ‘non-extremophiles'. Curr Opin Genet Dev. 1998;8:649–654. doi: 10.1016/s0959-437x(98)80032-4. [DOI] [PubMed] [Google Scholar]
- Fetzer S, Bak F, Conrad R. Sensitivity of methanogenic bacteria from paddy soil to oxygen and desiccation. FEMS Microbiol Ecol. 1993;12:107–115. [Google Scholar]
- Fetzer S, Conrad R. Effect of redox potential on methanogenesis by Methanosarcina barkeri. Arch Microbiol. 1993;160:108–113. [Google Scholar]
- Garcia J, Patel BKC, Ollivier B. Taxonomic, phylogenetic, and ecological diversity of methanogenic Archaea. Anaerobe. 2000;6:205–226. doi: 10.1006/anae.2000.0345. [DOI] [PubMed] [Google Scholar]
- Garcia-Pichel F.2002Desert environments: biological soil crustsIn: Bitton G (ed). Encyclopedia of Environmental Microbiology 6 Volume Set Wiley-Interscience: New York, NY, USA [Google Scholar]
- Garcia-Pichel F, Belnap J. Microenvironments and microscale productivity of cyanobacterial desert crusts. J Phycol. 1996;32:774–782. [Google Scholar]
- Garcia-Pichel F, Belnap J.2003Small-scale environments and distribution of biological soil crustIn: Belnap J, Lange OL (eds). Biological Soil Crusts Springer: Berlin, Germany [Google Scholar]
- Gattinger A, Höfle MG, Schloter M, Embacher A, Böhme F, Munch JC, et al. Traditional cattle manure application determines abundance, diversity and activity of methanogenic Archaea in arable European soil. Environ Microbiol. 2007;9:612–624. doi: 10.1111/j.1462-2920.2006.01181.x. [DOI] [PubMed] [Google Scholar]
- Gelwicks JT, Risatti JB, Hayes JM. Carbon isotope effects associated with aceticlastic methanogenesis. Appl Environ Microbiol. 1994;60:467–472. doi: 10.1128/aem.60.2.467-472.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glissmann K, Conrad R. Saccharolytic activity and its role as a limiting step in methane formation during the anaerobic degradation of rice straw in rice paddy soil. Biol Fertil Soils. 2002;35:62–67. [Google Scholar]
- Goevert D, Conrad R. Effect of substrate concentration on carbon isotope fractionation during acetoclastic methanogenesis by Methanosarcina barkeri, M. acetivorans and in rice field soil. Appl Environ Microbiol. 2009;75:2605–2612. doi: 10.1128/AEM.02680-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grosskopf R, Janssen PH, Liesack W. Diversity and structure of the methanogenic community in anoxic rice paddy soil microcosms as examined by cultivation and direct 16S rRNA gene sequence retrieval. Appl Environ Microbiol. 1998;64:960–969. doi: 10.1128/aem.64.3.960-969.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guérin F, Abril G, de Junet A, Bonnet M-P. Anaerobic decomposition of tropical soils and plant material: implication for the CO2 and CH4 budget of the Petit Saut Reservoir. Appl Geochem. 2008;23:2272–2283. [Google Scholar]
- Hedderich R, Whitman WB.2006Physiology and biochemistry of the methane-producing archaeaIn: Dworkin M, Falkow S, Rosenberg E, Schleifer K, Stackebrandt E (eds). The Prokaryotes Springer: New York [Google Scholar]
- Janssen P, Frenzel P. Inhibition of methanogenesis by methyl fluoride: studies of pure and defined mixed cultures of anaerobic bacteria and archaea. Appl Environ Microbiol. 1997;63:4552–4557. doi: 10.1128/aem.63.11.4552-4557.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiener A, Leisinger T. Oxygen sensitivity of methanogenic bacteria. Syst Appl Microbiol. 1983;4:305–312. doi: 10.1016/S0723-2020(83)80017-4. [DOI] [PubMed] [Google Scholar]
- Knight WG.1991Chemistry of arid region soilIn: Skujins J (ed). Semiarid Lands and Deserts: Soil Resource and Reclamation Marcel Dekker INC: New York; 111–171. [Google Scholar]
- Krzycki JA, Kenealy WR, DeNiro MJ, Zeikus JG. Stable carbon isotope fractionation by Methanosarcina barkeri during methanogenesis from acetate, methanol, or carbon dioxide-hydrogen. Appl Environ Microbiol. 1987;53:2597–2599. doi: 10.1128/aem.53.10.2597-2599.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kusel K, Drake HL. Acetate synthesis in soil from a Bavarian beech forest. Appl Environ Microbiol. 1994;60:1370–1373. doi: 10.1128/aem.60.4.1370-1373.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lange OL.2003Photosynthesis of soil-crust biota as dependent on environmental factorsIn: Belnap J, Lange OL (eds). Biological Soil Crusts Springer: Berlin, Germany; 363–383. [Google Scholar]
- Leininger S, Urich T, Schloter M, Schwark L, Qi J, Nicol GW, et al. Archaea predominate among ammonia-oxidizing prokaryotes in soils. Nature. 2006;442:806–809. doi: 10.1038/nature04983. [DOI] [PubMed] [Google Scholar]
- Liu C-T, Miyaki T, Aono T, Oyaizu H. Evaluation of methanogenic strains and their ability to endure aeration and water stress. Curr Microbiol. 2008;56:214–218. doi: 10.1007/s00284-007-9059-7. [DOI] [PubMed] [Google Scholar]
- Liu Y, Whitman W. Metabolic, phylogenetic, and ecological diversity of the methanogenic archaea. Ann NY Acad Sci. 2008;1125:171–189. doi: 10.1196/annals.1419.019. [DOI] [PubMed] [Google Scholar]
- Lu Y, Conrad R. In situ stable isotope probing of methanogenic archaea in the rice rhizosphere. Science. 2005;309:1088–1090. doi: 10.1126/science.1113435. [DOI] [PubMed] [Google Scholar]
- Ludwig W, Strunk O, Westram R, Richter L, Meier H, Yadhukumar, et al. ARB: a software environment for sequence data. Nucleic Acids Res. 2004;32:1363–1371. doi: 10.1093/nar/gkh293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lueders T, Manefield M, Friedrich MW. Enhanced sensitivity of DNA- and rRNA-based stable isotope probing by fractionation and quantitative analysis of isopycnic centrifugation gradients. Environ Microbiol. 2004;6:73–78. doi: 10.1046/j.1462-2920.2003.00536.x. [DOI] [PubMed] [Google Scholar]
- Luton PE, Wayne JM, Sharp RJ, Riley PW. The mcrA gene as an alternative to 16S rRNA in the phylogenetic analysis of methanogen populations in landfill. Microbiology. 2002;148:3521–3530. doi: 10.1099/00221287-148-11-3521. [DOI] [PubMed] [Google Scholar]
- Ma K, Lu Y. Regulation of microbial methane production and oxidation by intermittent drainage in rice field soil. FEMS Microbiol Ecol. 2011;75:446–456. doi: 10.1111/j.1574-6941.2010.01018.x. [DOI] [PubMed] [Google Scholar]
- Miyashita A, Mochimaru H, Kazama H, Ohashi A, Yamaguchi T, Nunoura T, et al. Development of 16S rRNA gene-targeted primers for detection of archaeal anaerobic methanotrophs (ANMEs) FEMS Microbiol Lett. 2009;297:31–37. doi: 10.1111/j.1574-6968.2009.01648.x. [DOI] [PubMed] [Google Scholar]
- Nicol GW, Glover LA, Prosser JI. Molecular analysis of methanogenic archaeal communities in managed and natural upland pasture soils. Global Change Biol. 2003;9:1451–1457. [Google Scholar]
- Nüsslein B, Eckert W, Conrad R. Stable isotope biogeochemistry of methane formation in profundal sediments of lake kinneret (Israel) Limnol Oceanogr. 2003;48:1439–1446. [Google Scholar]
- Ochsenreiter T, Selezi D, Quaiser A, Bonch-Osmolovskaya L, Schleper C. Diversity and abundance of Crenarchaeota in terrestrial habitats studied by 16S RNA surveys and real time PCR. Environ Microbiol. 2003;5:787–797. doi: 10.1046/j.1462-2920.2003.00476.x. [DOI] [PubMed] [Google Scholar]
- Oremland R.1988Biogeochemistry of methanogenic bacteriaIn: Zehnder AJ (ed). Biology of Anaerobic Microorganisms John Wiley & Sons Ltd: New York, NY, USA [Google Scholar]
- Pei J, Grishin NV. PROMALS: towards accurate multiple sequence alignments of distantly related proteins. Bioinformatics. 2007;23:802–808. doi: 10.1093/bioinformatics/btm017. [DOI] [PubMed] [Google Scholar]
- Peters V, Conrad R. Methanogenic and other strictly anaerobic bacteria in desert soil and other oxic soils. Appl Environ Microbiol. 1995;61:1673–1676. doi: 10.1128/aem.61.4.1673-1676.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peters V, Conrad R. Sequential reduction processes and initiation of CH4 production upon flooding of oxic upland soils. Soil Biol Biochem. 1996;28:371–382. [Google Scholar]
- Phelps TJ, Zeikus JG. Influence of pH on terminal carbon metabolism in anoxic sediments from a mildly acidic lake. Appl Environ Microbiol. 1984;48:1088–1095. doi: 10.1128/aem.48.6.1088-1095.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poplawski AB, Mårtensson L, Wartiainen I, Rasmussen U. Archaeal diversity and community structure in a Swedish barley field: specificity of the Ek510r/(EURY498) 16S rDNA primer. J Microbiol Methods. 2007;69:161–173. doi: 10.1016/j.mimet.2006.12.018. [DOI] [PubMed] [Google Scholar]
- Pruesse E, Quast C, Knittel K, Fuchs BM, Ludwig W, Peplies J, et al. SILVA: a comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007;35:7188–7196. doi: 10.1093/nar/gkm864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Radl V, Gattinger A, Chronakova A, Nemcova A, Cuhel J, Simek M, et al. Effects of cattle husbandry on abundance and activity of methanogenic archaea in upland soils. ISME J. 2007;1:443–452. doi: 10.1038/ismej.2007.60. [DOI] [PubMed] [Google Scholar]
- Roy R, Klüber HD, Conrad R. Early initiation of methane production in anoxic rice soil despite the presence of oxidants. FEMS Microbiol Ecol. 1997;24:311–320. [Google Scholar]
- Sakai S, Conrad R, Liesack W, Imachi H. Methanocella arvoryzae sp. nov., a hydrogenotrophic methanogen, isolated from Italian rice field soil. Int J Syst Evol Microbiol. 2010;60 (Pt 12:2918–2923. doi: 10.1099/ijs.0.020883-0. [DOI] [PubMed] [Google Scholar]
- Schleper C, Holben W, Klenk H. Recovery of crenarchaeotal ribosomal DNA sequences from freshwater-lake sediments. Appl Environ Microbiol. 1997;63:321–323. doi: 10.1128/aem.63.1.321-323.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schleper C, Jurgens G, Jonuscheit M. Genomic studies of uncultivated archaea. Nat Rev Micro. 2005;3:479–488. doi: 10.1038/nrmicro1159. [DOI] [PubMed] [Google Scholar]
- Soule T, Anderson IJ, Johnson SL, Bates ST, Garcia-Pichel F. Archaeal populations in biological soil crusts from arid lands in North America. Soil Biol Biochem. 2009;41:2069–2074. [Google Scholar]
- Stamatakis A. Raxml-Vi-Hpc: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics. 2006;22:2688–2690. doi: 10.1093/bioinformatics/btl446. [DOI] [PubMed] [Google Scholar]
- Stams AJM, Plugge CM. Electron transfer in syntrophic communities of anaerobic bacteria and archaea. Nat Rev Micro. 2009;7:568–577. doi: 10.1038/nrmicro2166. [DOI] [PubMed] [Google Scholar]
- Steinberg LM, Regan JM. Phylogenetic comparison of the methanogenic communities from an acidic, oligotrophic fen and an anaerobic digester treating municipal wastewater sludge. Appl Environ Microbiol. 2008;74:6663–6671. doi: 10.1128/AEM.00553-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takai K, Horikoshi K. Genetic diversity of archaea in deep-sea hydrothermal vent environments. Genetics. 1999;152:1285–1297. doi: 10.1093/genetics/152.4.1285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Teh YA, Silver WL, Conrad ME. Oxygen effects on methane production and oxidation in humid tropical forest soils. Global Change Biol. 2005;11:1283–1297. [Google Scholar]
- Thauer RK, Jungermann K, Decker K. Energy conservation in chemotrophic anaerobic bacteria. Microbiol Mol Biol Rev. 1977;41:100–180. doi: 10.1128/br.41.1.100-180.1977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thauer RK, Zinkhan DM, Spormann AM. Biochemistry of acetate catabolism in anaerobic chemotrophic bacteria. Ann Rev Microbiol. 1989;43:43–67. doi: 10.1146/annurev.mi.43.100189.000355. [DOI] [PubMed] [Google Scholar]
- Ueki A, Ono K, Tsuchiya A, Ueki K. Survival of methanogens in air-dried paddy field soil and their heat tolerance. Water Sci Tech. 1997;36:517–522. [Google Scholar]
- Wagner D, Lipski A, Embacher A, Gattinger A. Methane fluxes in permafrost habitats of the Lena Delta: effects of microbial community structure and organic matter quality. Environ Microbiol. 2005;7:1582–1592. doi: 10.1111/j.1462-2920.2005.00849.x. [DOI] [PubMed] [Google Scholar]
- West A, Schmidt S. Endogenous methanogenesis stimulates oxidation of atmospheric CH4 in Alpine tundra soil. Microb Ecol. 2002;43:408–415. doi: 10.1007/s00248-001-1049-x. [DOI] [PubMed] [Google Scholar]
- Whitman WB, Bowen T, Boone D.2006The methanogenic bacteriaIn: Dworkin M, Falkow S, Rosenberg E, Schleifer K, Stackebrandt E (eds). The Prokaryotes Springer: New York [Google Scholar]
- Woese CR, Fox GE. Phylogenetic structure of the prokaryotic domain: the primary kingdoms. Proc Natl Acad Sci USA. 1977;74:5088–5090. doi: 10.1073/pnas.74.11.5088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woese CR, Kandler O, Wheelis ML. Towards a natural system of organisms: proposal for the domains archaea, bacteria, and eucarya. Proc Natl Acad Sci USA. 1990;87:4576–4579. doi: 10.1073/pnas.87.12.4576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yao H, Conrad R, Wassmann R, Neue HU. Effect of soil characteristics on sequential reduction and methane production in sixteen rice paddy soils from china, the Philippines, and Italy. Biogeochemistry. 1999;47:269–295. [Google Scholar]
- Yu Y, Lee C, Kim J, Hwang S. Group-specific primer and probe sets to detect methanogenic communities using quantitative real-time polymerase chain reaction. Biotechnol Bioeng. 2005;89:670–679. doi: 10.1002/bit.20347. [DOI] [PubMed] [Google Scholar]
- Yuan Y, Conrad R, Lu Y. Responses of methanogenic archaeal community to oxygen exposure in rice field soil. Environ Microbiol Rep. 2009;1:347–335. doi: 10.1111/j.1758-2229.2009.00036.x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.