

Abstract
Consciousness is subjective experience. During both sleep and anesthesia consciousness is common, evidenced by dreaming. A defining feature of dreaming is that, while conscious, we do not experience our environment – we are disconnected. Besides inducing behavioral unresponsiveness, a key goal of anesthesia is to prevent the experience of surgery (connected consciousness), by inducing either unconsciousness or disconnection of consciousness from the environment. Review of the isolated forearm technique demonstrates that consciousness, connectedness and responsiveness uncouple during anesthesia; in clinical conditions, a median 37% of patients demonstrate connected consciousness. We describe potential neurobiological constructs that can explain this phenomenon: during light anesthesia the subcortical mechanisms subserving spontaneous behavioral responsiveness are disabled but information integration within the corticothalamic network continues to produce consciousness, and unperturbed norepinephrinergic signaling maintains connectedness. These concepts emphasize the need for developing anesthetic regimens and depth of anesthesia monitors that specifically target mechanisms of consciousness, connectedness and responsiveness.
Introduction
“The fact that the body is lying down is no reason for supposing that the mind is at peace. Rest is… far from restful.” Seneca in approximately 60 A.D. 1
Consciousness is subjective experience2,3; it has been defined as “what abandons us every night when we fall into dreamless sleep and returns the next morning when we wake up or when we dream”2. This definition has relevance for anesthesia, as both patients and anesthesiologists assume that general anesthesia is associated with unconsciousness similar to a dreamless sleep. Debates on consciousness, sleep and anesthesia are often bedeviled by a plethora of confusing, often tautological, and partially overlapping synonyms and terms: e.g. “consciousness”, “awareness”, “responsiveness”, “perception”, “subjective experience”, “wakefulness”, “vigilance”, “arousal”, “hypnosis”, “sleep”, “sedation”… To simplify and clarify matters, in this paper we emphasize the distinction among three separate concepts that are relevant to anesthesia: consciousness, connectedness and responsiveness. Consciousness is subjective experience be it pure darkness, an engrossing movie, or intraoperative pain. Environmental connectedness describes the connection of consciousness to the external world allowing experience of external stimuli. Consciousness can be disconnected (e.g. Dreaming – where we are not conscious of our environment) or connected (e.g. Wakeful – where experiences can be triggered by environmental stimuli). The state of “anesthesia awareness” is therefore a state of environmentally connected consciousness. We shift to this terminology to help unbundle “anesthesia awareness” into its component parts (“connectedness” to the environment and “consciousness”). While “anesthesia awareness” is often used interchangeably with “anesthesia awareness with explicit recall”, here we use the term to mean experiencing the event of surgery and do not specify that the events are remembered. The concepts of consciousness or connectedness should not be confused with words describing the complexity of our behavioral interactions with the outside world – which we term responsiveness. Responsiveness can be further divided between behavior that is spontaneous or goal-directed (such as following a command).
We will provide examples that consciousness is not necessarily coupled to connectedness or spontaneous responsiveness during anesthesia; and illustrate that differing measures of consciousness, connectedness, and responsiveness can all provide information to guide anesthesia. A familiar illustration of these distinctions can be obtained by considering natural sleep (Table 1). During wakefulness we are typically conscious, connected to the environment and responsive. As we fall asleep, responsiveness and connectedness to the environment fade but only during early non-rapid eye movement (NREM) sleep (where slow wave activity is abundant) do we become unconscious. Consciousness is present in NREM sleep later in the night and it becomes vivid during dreams in rapid eye movement (REM) sleep, although we remain disconnected from and largely unresponsive to our environment. While in sleep connectedness and responsiveness are tightly coupled so connection to the environment rapidly leads to responsiveness, studies of the isolated forearm technique demonstrate that coupling between responsiveness and connectedness is sometimes lost during anesthesia - hence unresponsiveness provides inconsistent and sometimes unreliable information about the probability of unconsciousness or of connected consciousness.
Table 1.
Consciousness, connectedness and responsiveness in wake and sleep states.
| Consciousness | Connectedness | Responsiveness | |
|---|---|---|---|
| Awake | Yes | Yes | Yes |
| NREM Sleep | No | No | No |
| REM Sleep | Yes | No | No |
NREM = non-rapid eye movement sleep. This refers to slow wave sleep early in the night when subjects are often unconscious. REM = rapid eye movement sleep
The goals of this paper are (i) to define more clearly the component features of the anesthetic state that subserve the experience of surgery (consciousness and connectedness), (ii) to provide evidence that spontaneous responsiveness is not a good correlate of connected consciousness and (iii) to provide a structural framework for future enquiry in this area.
Experience under Anesthesia
Suppression of the experience of surgery is a primary aim of anesthesia; this may be achieved by suppressing consciousness or ensuring disconnection. Consciousness itself may not be a clinical problem if it is merely associated with dreaming (the rare “bad trip” may be an example of when this could be deleterious) provided that the patient’s consciousness is truly disconnected from the external world – i.e. in a state analogous to rapid-eye-movement sleep4. In addition to suppressing connected consciousness, analgesia or nociceptive blockade is also necessary to prevent central nervous system arousal, and cardiovascular and neurohumoral responses to surgery. Finally gross patient immobility is required to facilitate surgery. We argue that, in most cases, the triad of general anesthesia we should aim for is: lack of experience of surgery (unconsciousness or disconnected consciousness), nociceptive blockade and immobility for surgery.
As alluded to above (in the Experience Under Anesthesia section), we do not address the important issue of anesthesia and memory (implicit or explicit) for several reasons: Firstly excellent recent reviews are available on this subject5,6. Secondly we believe that, if possible, we should be seeking to ablate the experience of surgery during anesthesia and not merely induce amnesia of the events. We acknowledge that there exists a large body of opinion that follows a utilitarian approach, and insists that unconsciousness per se is not a requirement for the state of general anesthesia – but that amnesia plus immobility are the minimal necessary components of anesthesia7. This raises numerous philosophical, humanitarian, and neurobiological questions. From the patients’ perspective, most would likely choose to not experience an event – rather than experience it and not remember it. However lack of connected consciousness (versus purely amnesia) is a better goal for the delivery of anesthesia for the following practical reasons:
Preventing the experience of surgery represents the most secure way of inhibiting consciousness with recall. In essence we need a dosage buffer zone. The small doses of general anesthetic required to ablate memory in the unstimulated patient are nowhere near enough to reliably obtund behavioral and autonomic arousal induced by noxious stimuli. In contrast the larger doses that are required to induce unconsciousness, will result in amnesia of events as a secondary effect, as well as substantially suppressing the effects of noxious stimuli.
Memory is not essential for experience. We have all driven down a familiar road with no recollection of the events. One would hardly deny we were conscious during this period. Similarly when moderately drunk, a person is conscious and responsive but may not recall anything later on. Finally the evidence from the case of H.M., who following medial temporal lobe resection incurred profound deficits in memory formation yet clearly was conscious, further dissociates memory and consciousness.
Our interest in experience under anesthesia is not merely academic. Herein we review the known and potential mechanisms of consciousness, connectedness and responsiveness and advocate that study of each is required to provide a detailed understanding of anesthesia. This understanding will provide novel approaches for designing anesthetic regimens and monitoring technology. First we discuss some examples that illustrate that unresponsiveness ≠ unconsciousness.
Disconnected Consciousness: Dreaming in Sleep & Anesthesia
Dreams are a good example of how consciousness may be disconnected. While early in the night, subjects that are awoken from slow wave sleep (the deepest stage of NREM sleep) tend not to report anything “going through their mind”8 (suggesting they were unconscious; Table 1), if they are awakened later in the night or during REM sleep, they usually report some conscious experience and often vivid narrative dreams2–4. Indeed, consciousness is reported in approximately 80% of REM sleep awakenings, and in 23–74% of NREM sleep awakenings4,9, though vivid dreams are more common during REM sleep4. While environmental stimuli can be incorporated into dreams during sleep, review of the evidence suggests that this occurs rarely (often associated with more noxious stimulation4), as patients are disconnected from the environment4. For example, even when patients who slept with their eyes taped open were presented illuminated objects they did not incorporate these stimuli into dreams10. It is possible that noxious stimulation may be incorporated more readily into dreams as it produces arousal from sleep (overcoming the disconnection).
Strikingly, dreaming also occurs in at least 27% of patients anesthetized with propofol and 28% of patients undergoing desflurane anesthesia11. Given the amnesic effects of anesthetic agents, these data likely underestimate the true prevalence of dreaming since the reports required the recall of dreams. Importantly in our recent 300 patient study of anesthetic-related dreaming, experience of surgery was not described by any patient who reported dreams11. We ascribe this to a state of disconnected consciousness, similar to REM sleep, where experience is insulated from the environment.
Under anesthesia, REM-like electroencephalogram phenomena have been termed “beta arousals”12, characterized by loss of slow waves and spindles (waxing and waning alpha frequency oscillations representing thalamic oscillations, examples from sleep are shown in Figure 1A), and an increase in higher frequencies in the electroencephalogram. Because of the similarity to REM sleep13–16, also known as “paradoxical” sleep17, we call it “paradoxical” anesthesia (Figure 1B, 1C). These states probably arise from a relative failure by the anesthetic drug to prevent an increase in cholinergic excitation of the cortex (in the presence of histaminergic suppression18), perhaps via basal forebrain neural activity3,19. As administration of a muscarinic antagonist, scopolamine, prevents dreaming in patients under propofol-nitrous oxide anesthesia it is likely that anesthetic dreaming is driven through cholinergic neurotransmission19 (similar to dreaming in natural sleep13). Interestingly a cholinesterase inhibitor, physostigmine, also provokes return of consciousness in patients sedated with propofol; though this treatment does not always induce gross behavioral responsiveness20 (see review21). In summary, a high cholinergic tone in the cortex may underlie dreaming (a state of disconnected consciousness) in both REM sleep and anesthesia (Figure 2).
Figure 1.

Electroencephalogram changes during non-rapid eye movement (NREM) to rapid eye movement-like (REM) transitions during sleep and general anesthesia. (A) During sleep, the change from NREM like activity to REM-like activity is accompanied by the loss of spindles and delta waves and a shift to higher frequency activity. (B) Midway through an operation REM-like activity was noted by loss of spindles and delta waves and a shift to higher frequency activity, in this snapshot, beta activity. (C) On emergence from anesthesia, the patient transitioned through both theta and gamma activity prior to waking, seemingly waking from a REM-like state (upper panel). This started at approximately 0.4 MAC of anesthesia, about 5 minutes from becoming responsive (lower panel). SWS = slow wave sleep. MAC = minimum alveolar concentration. SE = state entropy.
Figure 2.

Schematic representing changes in responsiveness, neuromodulators and corticothalamic network connectivity with escalating propofol dose. Panel A: We define four particular states: awake, positive response on the isolate forearm technique (IFT +ve), dreaming and unconscious. With increasing doses of propofol (Panel B) patients transition through these states, first entering a state of environmentally connected consciousness (ECC = IFT +ve), then disconnected consciousness (DC = dreaming) before becoming unconscious. Panel B: depicts propofol dose. Panel C: Putative parallel changes in neuromodulators underlying behavioral changes with escalating doses of propofol. Ach = acetylcholine. Norepi = norepinephrine. ECC is hypothesized to require active norepinephrinergic and cholinergic signaling. Panel D: changes in corticothalamic network connectivity. Put-Amy = Putamen and Amygdala connectivity. CT-VATT = corticothalamic-ventral attention network connectivity. CT-Total = total corticothalamic connectivity. ECC requires adequate CT-Total and CT-VATT signaling.
Connected consciousness is dissociable from spontaneous responsiveness
It is typical to awaken in the morning by transitioning through a period of REM sleep; we propose this favors the sequential activation of consciousness-connectedness-responsiveness. The value of this sequence is to ensure that conscious cognitive processing is active prior to connecting to the environment. Once connected, responsiveness rapidly follows – when wakeful we are conscious, connected and responsive.
There are some interesting variants of the sequence. At one extreme, subjects who sleepwalk are spontaneously responsive but are unconscious and do not follow commands (goal-directed behavior). At the other extreme is the sleep phenomenon of ‘lucid dreaming’. Subjects who are having a lucid dream are aware that they are asleep and can control their dreams. Remarkably they can communicate their experience to the environment through predefined eye or wrist movements13,22 (i.e. in a goal-directed manner).i Important examples of partially disconnected consciousness are hypnagogic and hypnopompic hallucinations at the transition between sleep and wakefulness, when subjects are conscious, connected to the environment, but remain immobile and incapable of moving due to brainstem mechanisms inducing REM sleep-like paralysis23. Nonetheless in the vast majority of cases of waking from natural sleep, conscious connection to the environment rapidly results in a state of spontaneous responsiveness.
Based on studies using the isolated forearm technique (IFT) (Table 2), and case reports of anesthetic awareness without change in patient variables24,25 (such as hemodynamic measures and respiratory rate) or spontaneous movement in non-paralyzed patients26,27, we provide evidence of connected consciousness that is dissociated from spontaneous responsiveness during anesthesia. In the IFT, anesthesia is followed by inflation of a cuff on the arm before neuromuscular blockade is induced. The cuff prevents paralysis of the hand allowing the patient to communicate to an observer through predefined hand movements, typically following a command like: “Mrs Jones, if you can hear me, squeeze my hand”. This is usually followed by further more complex commands such as: “Mrs Jones, if you are comfortable, squeeze my hand twice” – to detect the level of cognition in the patient. An advantage of the IFT is that it is not dependent on memory, indeed explicit recall of intraoperative events is usually absent (see reference28; Table 2). In most reports of positive IFT responses, the patient demonstrates almost normal cognitive function. Interestingly they show goal-directed responsiveness (i.e. they follow commands) but rarely show spontaneous responsiveness.
Table 2.
Isolated Forearm Technique responses during clinically relevant stimulation under anesthesia.
| Reference | Number of patients | Number of patients responding to IFT | Percentage of patients responding | Times when responsiveness noted | Number of responsive patients with explicit recall | Percentage of IFT responders, asked about pain, with pain |
|---|---|---|---|---|---|---|
| Tunstall, 1977111 | 12 | 4 | 33% | N/A | N/A | N/A |
|
| ||||||
| Tunstall & Sheikh, 1989112 | 113 | 47 | 42% | From 2 minutes until 5 minutes post-induction | 0% | N/A |
|
| ||||||
| Russell, 1986113 | E 30 | 2 | 7% (eye opening in 17%) | N/A | 0% | N/A |
| N20 25 | 11 | 44% (eye opening in 36%) | N/A | 4% | N/A | |
|
| ||||||
| Russell, 1993114 | 32 | 23 | 72% | N/A | 9% | 100% |
|
| ||||||
| King et al., 1993115 | 30 | 29 | 97% | Larygoscopy (33%), Skin incision (97%), 2 minutes after skin incision (20%) | 0% | 83% |
|
| ||||||
| Gaitini et al., 1995116 | K 25 | K 5 | K 24% | Within 13 minutes post-induction | 0% | N/A |
| T 25 | T 13 | T 52% | ||||
|
| ||||||
| Russell & Wang, 1997117 | 35 | 0 | 0% | N/A | 9% | N/A |
|
| ||||||
| St Pierre et al., 2000118 | E0.2 10 | E0.2 8 | E0.2 80% | Within 7.5 minutes post-induction | 6% | N/A |
| E0.3 10 | E0.3 7 | E0.3 70% | ||||
| E0.4 10 | E0.4 2 | E0.4 20% | ||||
|
| ||||||
| Russell & Wang, 200135 | 40 | 7 | 17.5% | 34–125 minutes | 0% | 14% |
|
| ||||||
| Schneider et al., 200234 | 20 | 8 | 40% | Within 3 minutes of intubation | 0% | N/A |
|
| ||||||
| Slavov et al., 2002119 | 41 | 10 | 24% | Within 1 minutes of intubation | 0% | N/A |
|
| ||||||
| Russell, 200633 | 12 | 12 | 100% | N/A | 44% | 8% |
|
| ||||||
| Andrade et al., 200836 | 184 | 2 | 1.1% | Within 17 minutes post-induction | 0% | 0% |
IFT = Isolated Forearm Technique. T = Thiopentone. E = Etomidate (superscript numbers refer to dose in mg/kg administered). K = Ketamine. F = Fentanyl. M = Midazolam. A = Alfentanil. P = Propofol. R = Remifentanil
In order to provide comprehensive review of the IFT literature one author (Dr. Sanders) performed a Medline search for “isolated forearm technique” identifying 18 relevant studies. Five studies were excluded28–32 as they did not test response to clinically relevant noxious external stimuli such as surgery or laryngoscopy (Table 2). Positive responses were observed in a median of 37% of patients (range 0 – 100%). The studies had heterogeneous methodology and employed a variety of anesthetic agents and techniques; therefore meta-analysis has not been performed. However even the more recent studies with modern anesthetic techniques show that a large proportion of patients will respond to the IFT during anesthesia and that IFT responses are not reliably detected by the electroencephalogram-based depth of anesthesia monitors33,34. For example, the Bispectral Index (BIS) cannot reliably distinguish between responders and non-responders to the IFT, both before and following laryngoscopy34 (i.e. the BIS values were the same in both groups). In one study, the stimulus of laryngoscopy produced a rise in BIS values in both groups from ~52 to ~70. Forty percent of the patients then became responsive to verbal command. One explanation for this finding is that both groups were conscious but only the responders were connected to the environment. Of course other explanations are possible including that the non-responders lacked motivation to respond or motor system impairment prevented response. Fortunately, the recent IFT studies suggest that intraoperative connected consciousness with pain is relatively infrequent (Table 2)33,35. We suspect that in the non-paralyzed patient, significant pain is usually stimulating enough to produce wakefulness; though clearly this is not helpful in the paralyzed patient.
Typically IFT patients with a positive response are in a state where they do not move their unparalyzed hand spontaneously rather they perform a goal-directed type of behavior. Interestingly a positive IFT response has been observed in a non-paralyzed pediatric patient undergoing orthopedic surgery36; although the patient did not make spontaneous movements or “wake up” they moved their hand to command. IFT positive responses may therefore share similarities with lucid dreaming where patients also do not move spontaneously, but clearly have higher order cognition and can perform goal-directed behavior The IFT data are supported by reports from neurolept anesthesia where patients report being “locked in”37 rather than unconscious during anesthesia24. Furthermore case reports describe external auditory and visual experiences under anesthesia. Indeed in one such case (without continued paralysis) a patient had an eye taped open for surgery and provided a description of visual scenes post-operatively yet did not move spontaneously during surgery27. In sum, these data suggest that connected consciousness may occur during anesthesia despite patients appearing spontaneously unresponsive.
The Utility of Connectedness as a Concept
Accepting that during general anesthesia the anesthesiologist’s primary role is to reduce the experience of surgery by suppressing connected consciousness, the most obvious and secure way of achieving this aim is to cause unconsciousness with deep enough anesthesia. However this may not be safely achievable due to the cardiovascular side effects of anesthesia especially given our limited ability to rely on autoregulation to protect end-organ function38. Furthermore it has been mooted that deep anesthesia may increase mortality or morbidity39 though this is controversial40–42. These are studies that use the BIS to measure depth of anesthesia, subsequently correlating sensitivity of the brain to anesthetics with mortality. It is unclear if the low BIS is a mediator or marker of subsequent mortality; many of these findings may be explained by reduced cerebral (and other end-organ) perfusion that also causes a decrease in BIS43. Remarkably the patients who have low BIS readings and suffer earlier mortality may also receive lower anesthetic doses40. Nonetheless given these concerns that unconsciousness (through increased depth of anesthesia) may be difficult to safely achieve, disconnecting the patient from the environment becomes an increasingly appealing goal. We will present a hypothesis that this can be achieved by additional suppression of norepinephrine signaling during anesthesia.
Furthermore consciousness during anesthesia may not be a clinical problem if it is disconnected i.e. not associated with experience of surgery. A depth of anesthesia monitor that could accurately detect consciousness but were not sensitive to connectedness might prompt an unnecessary deepening of anesthesia if a patient is conscious but already disconnected from his/her environment and hence not experiencing surgery (e.g. the dreaming patient). A measure of connectedness would have utility to highlight the potential for connected consciousness and critically identify the experience of surgery. A full understanding of how and when consciousness becomes connected to the environment is essential to properly monitor if consciousness is associated with experience of surgery or is more akin to dreaming. Next we discuss the mechanisms of consciousness, connectedness and responsiveness and explore how understanding of each mechanism provides insight into how anesthesia can be improved and monitored more effectively.
Mechanisms of Consciousness
The Integrated Information Theory2, the Global Workspace Theory44–47, and Cognitive binding48 represent recent attempts at characterizing the neural basis of consciousness. All three focus on corticothalamic network function (Figure 3), supported by significant evidence stemming primarily from human lesion and neuroimaging studies3,47. Here we refer primarily to the Integrated Information Theory2 however a discussion of the other two theories is available in Appendix 1.
Figure 3.
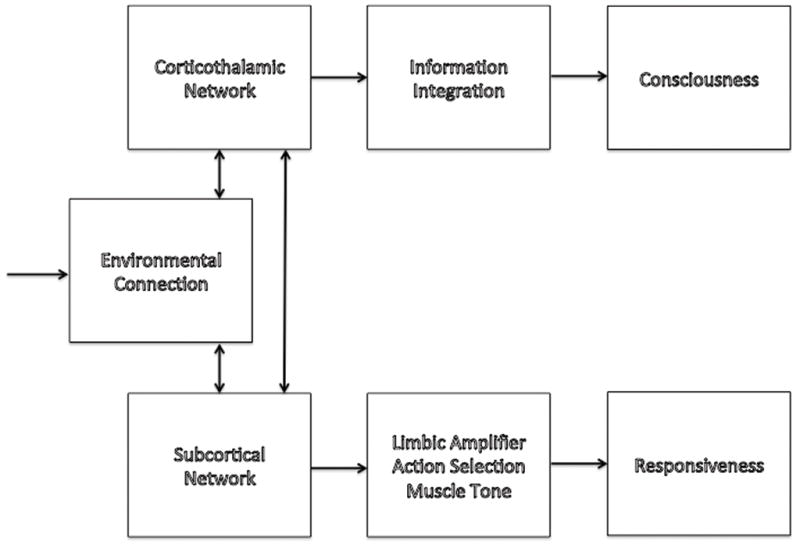
Schematic showing the relationship of consciousness, external connectedness and responsiveness with mediators where known.
The Integrated Information Theory of consciousness2 starts from two premises. First, every experience is one out of many: every experience, whether simple (pure darkness) or complex (a bustling street scene) represents a choice among a large repertoire of alternatives (think of all frames from all possible movies). For this reason, every experience is highly informative. Second, every experience is one: it cannot be decomposed into independent parts and it is thus integrated (for example, one cannot experience the left and right half of the visual field independently). From these premises, the theory says that the level of consciousness of a physical system is related to the repertoire of different states (information) that can be distinguished by the system as a whole (integration)”2,49. Thus, the corticothalamic system generates consciousness (Figure 3) because it can distinguish among a large repertoire of neural states (thanks to the functional specialization of different cortical areas and neurons) and it can do so as a single system (thanks to functional integration guaranteed by its dense intra- and inter-areal connections). Other brain structures such as the cerebellum lack the same pattern of connections, which is why, despite having even more neurons, they do not contribute to consciousness3. The theory accounts for the loss of consciousness during generalized seizures despite the hypersynchronous firing of neurons50 due to the resultant reduction of information (the repertoire of neural states shrinks). It also explains the fading of consciousness during slow wave sleep and certain anesthesia states, despite ongoing neural activity, as due an impairment of cortical integration and/or loss of information. Indeed a potentially important aspect of the Integrated Information Theory is that it is suggests experimental methods of obtaining semi-quantitative indices of consciousness based on evaluating the brain’s capacity for information integration.
The Integrated Information Theory2 predicts that in states associated with unconsciousness – such as in slow wave sleep early in the night, deep general anesthesia, and “vegetative” states – there is a breakdown in connectivity within the corticothalamic network. Direct cortical connectivity can be measured by transcranial magnetic stimulation (TMS) of the cortex to trigger a burst of local neuronal activity; a high density electroencephalogram can then be performed to assess the spread of the electrical activity. Consistent with predictions, during slow wave sleep and midazolam-induced hypnosis there is a breakdown of this connectivity, paralleling changes in consciousness51,52. Similar results have been found with functional magnetic resonance imaging approaches to assessing connectivity during slow wave sleep53 however connectivity is maintained in lighter sleep54. Perhaps of most interest is that connectivity during REM sleep, when consciousness is prevalent, is qualitatively similar to that found in wakefulness14. Thus during REM sleep, a state characterized by disconnected consciousness with unresponsiveness, there is intact corticothalamic integration; this emphasizes that responsiveness is not always an adequate measure of consciousness.
Consistent with this, it has been observed that propofol-induced unresponsiveness to verbal command occurs at lower doses than those required to suppress corticothalamic connectivity (Figure 2)55. At a propofol dose necessary for unresponsiveness, subcortical structures, in particular the putamen, become functionally disconnected. In this state the subject is unresponsive, but probably still conscious to some degree. Only at a deeper level of propofol, is reduced connectivity in corticothalamic networks observed56. Similar preservation of the corticothalamic connectivity under anesthesia has been observed in monkeys57. In summary, evidence suggests that slow wave sleep, deep anesthesia and coma reduce cortical connectivity, which is the key substrate of the integration of information51,52 – and that connectivity changes in parallel with consciousness. However evidence from light NREM sleep, REM sleep or lower doses of propofol shows that connectivity changes independently from arousal and responsiveness14: unresponsiveness ≠ unconsciousness.
Deep anesthesia may reduce consciousness also by reducing the available information within the corticothalamic network. This restriction of information may occur most obviously when the cortical neurons become bistable and oscillate, at around 1Hz, between depolarized “up” states, during which neurons can fire, and a hyperpolarized “down” state, during which neurons turn silent. The occurrence of down states vastly reduces the repertoire of states in the cortex and thereby the information available. Increasing anesthetic dosage increases the relative time the neurons are in the down state, which in turn appears to be associated with unconsciousness4. As the relative duration of the down states increases electroencephalographic changes occur with spindle and then delta wave activity, slow fluctuations in power, and eventually in burst suppression2,49,58. Recent evidence shows that spindles and delta waves are local (rather than global) phenomena, often occurring out of phase during NREM sleep suggesting that they may also inhibit the capacity for network integration in addition to reducing the available information59. Whether spindles and delta waves are also out of phase during anesthesia is unclear.
Changes in various neuromodulators may account for this increase in down states. Computer simulations suggest that acetylcholine plays a critical role in maintaining cortical network information integration by preventing down states during sleep60. During NREM sleep a reduction in cholinergic signaling leads to an increase in GABAergic tone within the cortex increasing the number of down states. This explains why consciousness occurs commonly in REM sleep but rarely in slow wave NREM sleep early in the night. Based on the clinical evidence19,20, it is likely that a similar cholinergic mechanism subserves disconnected consciousness (e.g. dreaming) during anesthesia.
Mechanism of Connectedness
As demonstrated by several studies, sensory stimuli can reach primary sensory cortices in both anesthesia49,61 and sleep4,62. Nevertheless, as illustrated most clearly by REM sleep, subjects may be vividly conscious of their dreams and yet completely ignore sensory signals4. Such disconnections in the face of vivid consciousness and despite the activation of sensory cortices suggests the presence of a “cortical gate” that “closes” to prevent the incorporation of sensory inputs into ongoing conscious processing. How such a cortical gate may be implemented remains unknown, but it appears likely that the opening and closing of the gate is ultimately controlled by the action of neuromodulators.
Although the complexity, overlapping function and the redundancy of the neuromodulatory systems makes it unlikely that connectedness can be solely ascribed to a single neurotransmission system, there is some evidence implicating specific neuromodulators. For example, a study of “narcoleptic” dogs suggested that during cataplexy, connectedness is maintained by active histaminergic signaling63 however the study could not definitively exclude a role for other pathways. Orexin, which is known to stabilize the “sleep-wake” switch64, could also conceivably contribute to a connection “switch”. However GABAergic anesthetics suppress both orexin (though halothane is an exception65) and histamine signaling18,66–69 and thus activation of these pathways is unlikely to account for maintained connected consciousness during anesthesia.
In contrast it is known that GABAergic anesthetics poorly suppress norepinephrinergic signaling18,66,68. We present a hypothesis that unperturbed norepinephrinergic neurotransmission is important in maintaining connectedness, due to its central role in controlling attention to external stimuli70. In particular, the cortical ventral attention network receives significant norepinephrinergic innervation from the locus ceruleus. Consistent with a role in orientating attention to environmental stimuli, areas of this network (especially the inferior parietal lobule) are deactivated during sleep when we are disconnected71. Furthermore alpha2 adrenergic agonists reduce attention to external stimuli via norepinephrinergic suppression70. Norepinephrine is well placed to control connectedness as it acts to increase the “signal-to-noise” ratio of cortical signaling following a sensory stimulus72–74; norepinephrine acts to reduce background neuronal activity while allowing evoked responses from sensory stimuli72–74. Suppression of norepinephrinergic signaling during sleep may explain why external stimuli rarely penetrate into dreams as the sensory “signal” is lost in cortical “noise”.
Inadequate suppression of norepinephrinergic signaling may explain connected consciousness evident in studies of the IFT despite clinical doses of anesthesia. Consistent with poor suppression of norepinephrine signaling, propofol unresponsiveness is not associated with reduced connectivity of the ventral attention network with other corticothalamic networks, though connectivity within the ventral attention network is diminished (perhaps due to the low level of external stimulation during the experimental study; Figure 2)75. Evidence for inadequate suppression of norepinephrine during GABAergic sedation is also forthcoming from a patient who underwent functional magnetic resonance image scanning during transition from wakefulness to sleep and wakefulness to sedation with midazolam or dexmedetomidine76. While dexmedetomidine and sleep produced remarkably similar effects (i.e. the scans showed few differences), activation was observed in the ventral attention network and thalamus during midazolam sedation relative to sleep. Therefore areas of the brain that receive significant norepinephrinergic innervation were not suppressed during midazolam sedation76; it is thus possible that during midazolam sedation, unlike in natural sleep, this patient remained connected to the environment. (However we should state that these data arise from a single subject and are prone to type 1 (false positive) error and hence further study is required.) Although much more experimental work is required to develop this hypothesis, it does suggest a plausible mechanism for why patients made spontaneously unresponsive by the administration of GABAergic drugs may remain connected to the environment. In turn this hypothesis implies that supplementation of a GABAergic anesthetic with an alpha2 agonist – as an adjunct to suppress norepinephrine signaling - may reduce connected consciousness under anesthesia. Nevertheless, we do not advocate that sole alpha2 agonist sedation/anesthesia disconnects the patient adequately for anesthesia. Indeed patients sedated with alpha2 agonists are remarkably rousable; this may be due to unperturbed excitatory neurotransmission (perhaps orexin signaling68) overcoming the norepinephrinergic suppression76.
In sum, given that (i) GABAergic anesthetics suppress histamine and orexin neurotransmission but do not perturb norepinephrine signaling; (ii) data from the IFT and case reports of anesthesia awareness demonstrate connectedness during anesthesia; and (iii) norepinephrinergic activity is suppressed during sleep when we are disconnected but is poorly suppressed during GABAergic anesthesia; (iv) norepinephrinergic signaling is known to play a role in enhancing cortical responses to external stimuli; it is likely that norepinephrine signaling plays an important role mediating connectedness and it follows that alpha2 agonists may therefore aid in the suppression of environmental connectedness under anesthesia.
During anesthesia, a “thalamic gate” has also been proposed to block ascending sensory information as thalamic hyperpolarization closes the “gateway” to the cortex77. The thalamus is most likely to block transfer of external information when hyperpolarized and enters a oscillatory burst-firing mode identified by electroencephalogram spindles78,79; this may be a biomarker of the “thalamic gate”. Abundant spindle activity can be seen in approximately 60% of patients intraoperatively80. However loss of spindle activity is a common response to surgical stimulation81. Therefore while the anesthetized thalamus does filter some information77, the thalamic “gate” is rarely absolutely closed (evidenced the activation of primary sensory cortices during anesthesia61).
Similar to the ventral attention network, the thalamus receives significant norepinephrinergic innervation82. When falling into natural sleep or with sedation with alpha2 agonist, norepinephrine signaling fades and in parallel there is reduced thalamic activity83,84 (evidenced by spindle activity in sleep and during dexmedetomidine sedation84). In keeping with the limited effect of GABAergic anesthetics on norepinephrinergic signaling, propofol and midazolam poorly suppresses thalamic activity at doses that prevent spontaneous unresponsiveness56,76. Thus continuing norepinephrinergic signaling during GABAergic sedation may maintain thalamic activity – leaving the “gate” open. Supplementation of anesthesia with an agent that suppresses norepinephrinergic activity, such as alpha2 adrenergic agonist, may reinforce thalamic hyperpolarization and help close the “thalamic gate” (again this must be confirmed in further studies).
Mechanisms of Unresponsiveness
Given the behavioral similarities between sleep and anesthesia it is unsurprising that anesthetics act, in part, on endogenous sleep pathways to reduce responsiveness79,85. In particular, suppression of histamine release from the tuberomammillary nucleus appears important for anesthesia18,67,79. Anesthetic effects in the brainstem also play an important role in producing unresponsiveness by reducing motor tone and inhibiting spontaneous motor responses86,87. Furthermore actions in the ventral horn of the spinal cord prevent reflex motor responses88. Nonetheless clinical data from the IFT show that anesthetics appear to differentially affect spontaneous and goal-directed responsiveness i.e. patients do not move spontaneously but will to verbal command. Next we explore the potential neurobiological explanations for this finding.
Based on neuroimaging studies55,56,75, corticothalamic connectivity (the substrate of consciousness) does not appear to play an important role in the mechanism of unresponsiveness (Figure 3). One interesting possibility is that reduced histamine release18, and potentially other as yet unidentified neurotransmitters, associated with anesthetic-induced spontaneous unresponsiveness does not lead to a breakdown in corticothalamic network connectivity but preferentially affects subcortical areas55. Several subcortical structures are innervated by the tuberomamillary nucleus, notably the amygdala and basal ganglia89, and are affected by low doses of anesthetics55,90. The subcortical regions affected play a role in learning, memory, motivation, and emotion, as well as motor output by action selection91. Propofol-induced spontaneous unresponsiveness is associated with reduced activity and connectivity of subcortical regions, in particular the putamen and amygdala55. A similar reduction of hippocampal-amygdala connectivity has been noted at subhypnotic levels with sevoflurane90. Reduced histamine release into subcortical regions maybe particularly important for producing spontaneous unresponsiveness by impairment of motivation/decision-making and action selection.
Amygdala activity and connectivity is highly susceptible to anesthetics55,90. The amygdala also plays an important role in decision making likely by ascribing value to an experience or action92. Inhibition of the amygdala during anesthesia may reduce the “value” of responding to stimuli.
The putamen plays a prominent role in the integrated basal ganglia circuit that controls motor output via selection of a particular action from competing options91. As its activity/connectivity is suppressed by propofol55 and increased by sleep deprivation93 it appears sensitive to arousal. Damage to the putamen is associated with loss of habitual behavior forcing patients into a goal-directed type of behavior91. It is likely that disconnection of the putamen during anesthesia anesthesia perturbs basal ganglia action selection, contributing to anesthesia-induced spontaneous unresponsiveness by acting in concert with anesthesia-mediated inhibition of descending and ventral horn control of motor responses86,87. In summary, the behavioral phenotype of spontaneous unresponsiveness may result from reduced ability to select a particular motor action out of a set of alternatives superimposed on descending motor inhibition from the brainstem resulting in a resting, “default” motor profile: unresponsiveness.
The loss of connectivity of the amygdala and putamen may also contribute to patient passivity as decision-making is impaired (loss of the “value” of responsiveness). Indeed due to basal ganglia effects, it is plausible that spontaneous motor responses may be impaired during the IFT, but may be overridden when patients follow predefined commands (goal-directed behavior).ii Anesthestic effects on the amygdala may also reduce the emotion and fear associated with pain, perhaps explaining why positive IFT responses are rarely associated with pain (Table 2). The amygdala may be seen as an “amplifier” that acts to disseminate warning signals throughout the brain92. During anesthesia this amplifier is turned down, reducing the impact of nociceptive signaling. Pain is also relatively rare in case reports of “anesthesia awareness”, rather feelings of weakness, paralysis and helplessness and hearing noises are typically reported26. Therefore it is clear that the subcortical anesthetic effects do not abrogate all the distress of connected consciousness during anesthesia.
Clinical implications and Future Directions
The most secure way of suppressing connected consciousness is to induce unconsciousness with deep enough anesthesia. However an alternate strategy is to reinforce the disconnection. Overall we advocate “balancing anesthesia” to ensure adequate suppression of a range of neurotransmitters that may be involved with consciousness and connectedness; and specifically enhanced suppression of norepinephrinergic signaling for a given “dose” of GABAergic anesthetic (a proposal that requires formal testing). Indeed we argue that achieving a depth of anesthesia that produces unconsciousness may be unnecessary provided the patient is disconnected from the environment.
Depth of Anesthesia monitors: Limitations and future directions
Present depth of anesthesia monitors have been developed based on the assumption that responsiveness and connected consciousness are causally linked and are not dissociable. However we have provided evidence that consciousness can occur in states of reduced responsiveness (e.g. REM sleep), even in states of reduced arousal. This will result in both false negative and false positive errors – which cannot be overcome with more sophisticated signal processing techniques. The numerous reports of failed detection of consciousness with recall94, dreaming11,95 and response on the isolated forearm technique (connected consciousness)33,34,96 by currently-used depth of anesthesia monitors are eloquent witness to this problem. To detect consciousness reliably, the electroencephalographic index should directly correspond with the actual neurobiological process required for consciousness – i.e. corticothalamic integration of information.
For detecting consciousness the use of transcranial magnetic stimulation and high density electroencephalogram under anesthesia appears particularly promising51 given that connectivity can measure neural correlates of consciousness independent from arousal level and responsiveness with good spatial resolution14,52. Furthermore this technology is not based on measures of behavioral responsiveness, but on objective evidence for corticothalamic integration of information. However before it can be routinely employed in the operating room this technology will have to significantly refined and simplified.
However, as mentioned above, a monitor that detected all conscious activity (including transcranial magnetic stimulation and high density electroencephalogram monitoring) could result in unnecessarily deep anesthesia – particularly when adjunctive nitrous oxide, opioids, or ketamine are used in the anesthesia. In order to be clinically useful we propose that, in addition to separating measures of responsiveness and consciousness, monitors need to specifically identify connected consciousness.
Research into the mechanism of connectedness is urgently needed to identify measures that are casually related to the ability to experience external stimuli. Evoked potentials offer a possible route to evaluating connectedness; although it is unclear which attributes of evoked potentials could lead to a reliable measure of connectedness. Evoked potential changes that correlate with IFT responsiveness (such as amplitude changes in mid latency auditory evoked potentials32) represent one approach to identify biomarkers of connected consciousness under anesthesia. The long latency components of an evoked response also offer promise as they are perturbed during anesthesia97–99, in vegetative state patients100 and inhibition of the long latency electrophysiological response to a visual stimulus in the brain prevents the experience of the external stimulus101. Indeed these long latency responses typically depend on backward (anterior to posterior) connectivity that are sensitive to anesthetic agents61,97–99,102,103. Using a mismatch negativity paradigm of auditory evoked potentials (where an “oddball” sound is intermittently mixed with a standardized repeated sound) long latency responses and fronto-temporal connectivity were found to be absent in vegetative state patients relative to controls100. However mismatch negativity responses can be similar in wakefulness and REM sleep104, therefore it is unclear how well these responses reflect connectedness. Further research is required to the correlation between evoked responses, connectedness and IFT responsiveness.
Another surrogate measure of connectedness – activity/connectivity of the ventral attention network – may already be measured. However it is possible that ventral attention network activation may occur following environmental connectedness, particularly following noxious stimulation, and so would merely inform the anesthesiologist that the patient is already connected. Nonetheless a measure such as this would inform the anesthesiologist that either (i) a greater dose of anesthetic/analgesic is required or (ii) increased suppression of norepinephrinergic signaling is required.
Emergence from Anesthesia and Delirium
Finally there are some implications for emergence from anesthesia. Unexpected connected consciousness during anesthesia can be viewed as “anesthesia awareness”. However manipulating the emergence sequence of consciousness-connectedness-responsiveness may also prove important for emergence from anesthesia and sedation. We have already discussed the value of sequential activation of consciousness-connectedness-responsiveness when waking from REM sleep to ensure that conscious cognitive processing is active prior to connecting to the environment. By contrast, awakening from a NREM like state can result in sleep inertia – a state of confusion on awakening. We suggest that a similar problem may underlie emergence delirium in anesthesia and some forms of delirium in the critically ill where conscious processing is impaired by increased GABAergic inhibitory tone within the corticothalamic network105. In these states connectedness can be considered to have occurred too rapidly – before the subject was sufficiently conscious to allow complex conscious cognitive interaction with the environment. Our proposal is consistent with the idea that connectedness is under norepinephrinergic control and that emergence from anesthesia is slowed by reducing norepinephrinergic signaling106. Emergence delirium can be considered as a state of excess noradrenaline with relatively low levels of other excitatory neuromodulators such as acetylcholine and histamine (possibly through increased GABAergic signaling105). This may account in part for why alpha2 adrenergic drugs may have utility in the treatment and prevention of delirium107,108, and why antimuscarinic (e.g. hyoscine) and GABAergic drugs (e.g. benzodiazepines) worsen delirium105.
Conclusions
Anesthesiologists must remain cognizant that unresponsiveness ≠ unconsciousness and that connected consciousness ≠ spontaneous responsiveness. Lack of spontaneous responsiveness does not inform us that the patient is not experiencing surgery. Furthermore we argue that amnesia of events under anesthesia is not a sufficient aim for anesthesiologists but rather a key aim of anesthesia is to prevent the experience of surgery. Calculation of the numbers needed to treat for “explicit recall of surgery” versus “lack of experience of surgery” illustrates that anesthesia may not be as successful as we think: the number of patients needed to treat to prevent explicit recall of surgery is approximately 1.002 (based on an incidence of “anesthetic awareness” of 0.2%109,110); the numbers needed to treat to prevent connected consciousness (based on the median responsiveness value from studies of the IFT of 37%) is 1.587.
Future depth of anesthesia monitors should focus on specific biomarkers of both consciousness and connectedness. We hypothesize that norepinephrinergic signaling influences the potential for connected consciousness and therefore supplementation of anesthesia with an alpha2 adrenergic agonist may have a beneficial role in preventing this connectedness. Advance in anesthesiology requires that the component features of anesthesia - consciousness, connectedness and responsiveness - be adequately unbundled.
Acknowledgments
Funding: RDS is supported by a Clinical Training Fellowship from the Medical Research Council, London, United Kingdom. GT by a Director’s Pioneer Award from the National Institutes of Health, Bethesda, Maryland, USA. SL is funded by the Belgian National fund for Scientific Research, Brussels, Belgium and European Commission (DECODER project). JW is supported by the University of Auckland and the Waikato District Health Board, Hamilton, New Zealand.
Appendix 1. Theories of Consciousness: Global Workspace Theory & Cognitive Binding
The Global Workspace Theory44, 45 suggests that different brain regions broadcast their information into a “global workspace” shared across the corticothalamic network and in particular bilateral frontoparietal regions. This has been likened to a stage (the global workspace), where a spotlight (consciousness) illuminates an actor speaking to many unconscious audience members (representing unconscious neural processes). Further actors are waiting in the wings to replace the one on stage, a decision made in part by attention. The theory suggests that the global workspace acts as a distribution centre for information. This theory has gained support from sleep, coma and anesthesia studies that have demonstrated reduced activity within frontoparietal regions (thought to represent the global workspace) with loss of consciousness47.
Cognitive binding attempts to tackle the binding problem, defined as the problem of binding together in a single unified percept the different features of an object48. Binding of information has been proposed at neural (convergent binding), systems (assembly binding) and global (synchronous binding) levels48. Indeed there is evidence that each of these mechanisms of binding play a role in different cognitive processes including perception. At a macroscopic level, electroencephalogram measured gamma oscillations are thought to play a role in synchronous cognitive binding integrating information across the corticothalamic network through co-incidence detection of neural firing patterns48, 120. In humans, anesthesia has been proposed to unbind cognitive processes by inhibiting gamma oscillations and their coherence48, 120.
Footnotes
It is unclear whether a person who is having a lucid dream is totally connected to the external world as, while they can communicate to the external world, it is unknown whether they will directly respond to spoken command. This needs to be investigated.
It is of interest that the studies of lucid dreaming and the IFT all predefined the commands. This may be an important factor in determining the ability of patients to respond in sleep or anesthesia.
Conflict of Interest: RDS has received honoraria for speaking on behalf of Hospira, Illinois, USA. Hospira, or any other industry, had no input into design of the talk or this manuscript.
References
- 1.Seneca L. Letters from a Stoic: Epistulae Morales ad Lucilium. New York: Penguin Books; 1969. [Google Scholar]
- 2.Tononi G. An information integration theory of consciousness. BMC Neurosci. 2004;5:42. doi: 10.1186/1471-2202-5-42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Tononi G, Laureys S. In: The Neurology of Consciousness: An Overview, The Neurology of Consciousness. 1. Laureys S, Tononi G, editors. London: Academic Press; 2009. pp. 375–412. [Google Scholar]
- 4.Nir Y, Tononi G. Dreaming and the brain: from phenomenology to neurophysiology. Trends Cogn Sci. 2010;14:88–100. doi: 10.1016/j.tics.2009.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Perouansky M, Pearce RA. How we recall (or don’t): the hippocampal memory machine and anesthetic amnesia. Can J Anaesth. 2010;57 :157–66. doi: 10.1007/s12630-010-9417-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Mashour GA, Orser B, Avidan MS. Intraoperative Awareness: From Neurobiology to Clinical Practice. Anesthesiology. 2011;114:1218–33. doi: 10.1097/ALN.0b013e31820fc9b6. [DOI] [PubMed] [Google Scholar]
- 7.Eger EI, 2nd, Sonner JM. Anaesthesia defined (gentlemen, this is no humbug) Best Pract Res Clin Anaesthesiol. 2006;20:23–9. doi: 10.1016/j.bpa.2005.07.011. [DOI] [PubMed] [Google Scholar]
- 8.Stickgold R, Malia A, Fosse R, Propper R, Hobson JA. Brain-mind states: I. Longitudinal field study of sleep/wake factors influencing mentation report length. Sleep. 2001;24:171–9. doi: 10.1093/sleep/24.2.171. [DOI] [PubMed] [Google Scholar]
- 9.Rechtshaffen A. The psychophysiology of mental activity during sleep. The Psychophysiology of Thinking: Studies of Covert Processes. 1973:153–205. [Google Scholar]
- 10.Rechtschaffen A, Foulkes D. Effect of Visual Stimuli on Dream Content. Perceptual and motor skills. 1965;20:1149–60. doi: 10.2466/pms.1965.20.3c.1149. [DOI] [PubMed] [Google Scholar]
- 11.Leslie K, Sleigh J, Paech MJ, Voss L, Lim CW, Sleigh C. Dreaming and electroencephalographic changes during anesthesia maintained with propofol or desflurane. Anesthesiology. 2009;111:547–55. doi: 10.1097/ALN.0b013e3181adf768. [DOI] [PubMed] [Google Scholar]
- 12.Aho AJ, Lyytikainen LP, Yli-Hankala A, Kamata K, Jantti V. Explaining Entropy responses after a noxious stimulus, with or without neuromuscular blocking agents, by means of the raw electroencephalographic and electromyographic characteristics. Br J Anaesth. 2011;106:69–76. doi: 10.1093/bja/aeq300. [DOI] [PubMed] [Google Scholar]
- 13.Hobson JA. REM sleep and dreaming: towards a theory of protoconsciousness. Nat Rev Neurosci. 2009;10:803–13. doi: 10.1038/nrn2716. [DOI] [PubMed] [Google Scholar]
- 14.Massimini M, Ferrarelli F, Murphy M, Huber R, Riedner B, Casarotto S, Tononi G. Cortical reactivity and effective connectivity during REM sleep in humans. Cogn Neurosci. 2010;1:176–183. doi: 10.1080/17588921003731578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Fuller PM, Saper CB, Lu J. The pontine REM switch: past and present. J Physiol. 2007;584:735–41. doi: 10.1113/jphysiol.2007.140160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Siegel JM. Clues to the functions of mammalian sleep. Nature. 2005;437:1264–71. doi: 10.1038/nature04285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Jouvet M, Michel F. New research on the structures responsible for the “paradoxical phase” of sleep. Journal de physiologie. 1960;52:130–1. [PubMed] [Google Scholar]
- 18.Nelson LE, Guo TZ, Lu J, Saper CB, Franks NP, Maze M. The sedative component of anesthesia is mediated by GABA(A) receptors in an endogenous sleep pathway. Nat Neurosci. 2002;5:979–84. doi: 10.1038/nn913. [DOI] [PubMed] [Google Scholar]
- 19.Toscano A, Pancaro C, Peduto VA. Scopolamine prevents dreams during general anesthesia. Anesthesiology. 2007;106:952–5. doi: 10.1097/01.anes.0000265154.24685.47. [DOI] [PubMed] [Google Scholar]
- 20.Meuret P, Backman SB, Bonhomme V, Plourde G, Fiset P. Physostigmine reverses propofol-induced unconsciousness and attenuation of the auditory steady state response and bispectral index in human volunteers. Anesthesiology. 2000;93:708–17. doi: 10.1097/00000542-200009000-00020. [DOI] [PubMed] [Google Scholar]
- 21.Backman SB, Fiset P, Plourde G. Cholinergic mechanisms mediating anesthetic induced altered states of consciousness. Prog Brain Res. 2004;145:197–206. doi: 10.1016/S0079-6123(03)45014-0. [DOI] [PubMed] [Google Scholar]
- 22.Voss U, Holzmann R, Tuin I, Hobson JA. Lucid dreaming: a state of consciousness with features of both waking and non-lucid dreaming. Sleep. 2009;32:1191–200. doi: 10.1093/sleep/32.9.1191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Cheyne JA, Newby-Clark IR, Rueffer SD. Relations among hypnagogic and hypnopompic experiences associated with sleep paralysis. Journal of sleep research. 1999;8:313–7. doi: 10.1046/j.1365-2869.1999.00165.x. [DOI] [PubMed] [Google Scholar]
- 24.Linnemann MU, Guldager H, Nielsen J, Ibsen M, Hansen RW. Psychomimetic reactions after neurolept and propofol anaesthesia. Acta Anaesthesiol Scand. 1993;37:29–32. doi: 10.1111/j.1399-6576.1993.tb03593.x. [DOI] [PubMed] [Google Scholar]
- 25.Moerman N, Bonke B, Oosting J. Awareness and recall during general anesthesia. Facts and feelings Anesthesiology. 1993;79:454–64. doi: 10.1097/00000542-199309000-00007. [DOI] [PubMed] [Google Scholar]
- 26.Ghoneim MM, Block RI, Haffarnan M, Mathews MJ. Awareness during anesthesia: risk factors, causes and sequelae: a review of reported cases in the literature. Anesth Analg. 2009;108:527–35. doi: 10.1213/ane.0b013e318193c634. [DOI] [PubMed] [Google Scholar]
- 27.Kino A, Kitamura R, Wakamatsu T, Hashiguchi M, Nakamura K. Awareness during anesthesia with sevoflurane: a case report. Masui. 2006;55:1250–2. [PubMed] [Google Scholar]
- 28.Kerssens C, Klein J, Bonke B. Awareness: Monitoring versus remembering what happened. Anesthesiology. 2003;99:570–5. doi: 10.1097/00000542-200309000-00011. [DOI] [PubMed] [Google Scholar]
- 29.Ge SJ, Zhuang XL, Wang YT, Wang ZD, Chen SL, Li HT. Performance of the rapidly extracted auditory evoked potentials index to detect the recovery and loss of wakefulness in anesthetized and paralyzed patients. Acta Anaesthesiol Scand. 2003;47:466–71. doi: 10.1034/j.1399-6576.2003.00090.x. [DOI] [PubMed] [Google Scholar]
- 30.Loveman E, Van Hooff JC, Smith DC. The auditory evoked response as an awareness monitor during anaesthesia. Br J Anaesth. 2001;86:513–8. doi: 10.1093/bja/86.4.513. [DOI] [PubMed] [Google Scholar]
- 31.Flaishon R, Windsor A, Sigl J, Sebel PS. Recovery of consciousness after thiopental or propofol. Bispectral index and isolated forearm technique. Anesthesiology. 1997;86:613–9. doi: 10.1097/00000542-199703000-00013. [DOI] [PubMed] [Google Scholar]
- 32.Bell SL, Smith DC, Allen R, Lutman ME. The auditory middle latency response, evoked using maximum length sequences and chirps, as an indicator of adequacy of anesthesia. Anesth Analg. 2006;102:495–8. doi: 10.1213/01.ane.0000189191.71449.48. [DOI] [PubMed] [Google Scholar]
- 33.Russell IF. The Narcotrend ‘depth of anaesthesia’ monitor cannot reliably detect consciousness during general anaesthesia: an investigation using the isolated forearm technique. Br J Anaesth. 2006;96:346–52. doi: 10.1093/bja/ael017. [DOI] [PubMed] [Google Scholar]
- 34.Schneider G, Wagner K, Reeker W, Hanel F, Werner C, Kochs E. Bispectral Index (BIS) may not predict awareness reaction to intubation in surgical patients. J Neurosurg Anesthesiol. 2002;14:7–11. doi: 10.1097/00008506-200201000-00002. [DOI] [PubMed] [Google Scholar]
- 35.Russell IF, Wang M. Absence of memory for intra-operative information during surgery with total intravenous anaesthesia. Br J Anaesth. 2001;86:196–202. doi: 10.1093/bja/86.2.196. [DOI] [PubMed] [Google Scholar]
- 36.Andrade J, Deeprose C, Barker I. Awareness and memory function during paediatric anaesthesia. Br J Anaesth. 2008;100:389–96. doi: 10.1093/bja/aem378. [DOI] [PubMed] [Google Scholar]
- 37.Laureys S, Pellas F, Van Eeckhout P, Ghorbel S, Schnakers C, Perrin F, Berre J, Faymonville ME, Pantke KH, Damas F, Lamy M, Moonen G, Goldman S. The locked-in syndrome : what is it like to be conscious but paralyzed and voiceless? Prog Brain Res. 2005;150:495–511. doi: 10.1016/S0079-6123(05)50034-7. [DOI] [PubMed] [Google Scholar]
- 38.Sanders RD, Degos V, Young WL. Cerebral perfusion under pressure: is the autoregulatory ‘plateau’ a level playing field for all? Anaesthesia. 2011 doi: 10.1111/j.1365-2044.2011.06915.x. [Epub ahead of print] [DOI] [PubMed] [Google Scholar]
- 39.Monk TG, Saini V, Weldon BC, Sigl JC. Anesthetic management and one-year mortality after noncardiac surgery. Anesthesia and analgesia. 2005;100:4–10. doi: 10.1213/01.ANE.0000147519.82841.5E. [DOI] [PubMed] [Google Scholar]
- 40.Kertai MD, Pal N, Palanca BJ, Lin N, Searleman SA, Zhang L, Burnside BA, Finkel KJ, Avidan MS. Association of perioperative risk factors and cumulative duration of low bispectral index with intermediate-term mortality after cardiac surgery in the B-Unaware Trial. Anesthesiology. 2010;112:1116–27. doi: 10.1097/ALN.0b013e3181d5e0a3. [DOI] [PubMed] [Google Scholar]
- 41.Kertai MD, Palanca BJ, Pal N, Burnside BA, Zhang L, Sadiq F, Finkel KJ, Avidan MS. Bispectral index monitoring, duration of bispectral index below 45, patient risk factors, and intermediate-term mortality after noncardiac surgery in the B-Unaware Trial. Anesthesiology. 2011;114:545–56. doi: 10.1097/ALN.0b013e31820c2b57. [DOI] [PubMed] [Google Scholar]
- 42.Lindholm ML, Traff S, Granath F, Greenwald SD, Ekbom A, Lennmarken C, Sandin RH. Mortality within 2 years after surgery in relation to low intraoperative bispectral index values and preexisting malignant disease. Anesthesia and analgesia. 2009;108:508–12. doi: 10.1213/ane.0b013e31818f603c. [DOI] [PubMed] [Google Scholar]
- 43.Leslie K, Short TG. Low bispectral index values and death: the unresolved causality dilemma. Anesthesia and analgesia. 2011;113:660–3. doi: 10.1213/ANE.0b013e31822401cc. [DOI] [PubMed] [Google Scholar]
- 44.Baars BJ, Ramsoy TZ, Laureys S. Brain, conscious experience and the observing self. Trends Neurosci. 2003;26:671–5. doi: 10.1016/j.tins.2003.09.015. [DOI] [PubMed] [Google Scholar]
- 45.Baars BJ, Franklin S. How conscious experience and working memory interact. Trends Cogn Sci. 2003;7:166–172. doi: 10.1016/s1364-6613(03)00056-1. [DOI] [PubMed] [Google Scholar]
- 46.Dehaene S, Naccache L. Towards a cognitive neuroscience of consciousness: basic evidence and a workspace framework. Cognition. 2001;79:1–37. doi: 10.1016/s0010-0277(00)00123-2. [DOI] [PubMed] [Google Scholar]
- 47.Boveroux P, Bonhomme V, Boly M, Vanhaudenhuyse A, Maquet P, Laureys S. Brain function in physiologically, pharmacologically, and pathologically altered states of consciousness. Int Anesthesiol Clin. 2008;46:131–46. doi: 10.1097/AIA.0b013e318181a8b3. [DOI] [PubMed] [Google Scholar]
- 48.Mashour GA. Consciousness unbound: toward a paradigm of general anesthesia. Anesthesiology. 2004;100:428–33. doi: 10.1097/00000542-200402000-00035. [DOI] [PubMed] [Google Scholar]
- 49.Alkire MT, Hudetz AG, Tononi G. Consciousness and anesthesia. Science. 2008;322:876–80. doi: 10.1126/science.1149213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Boly M, Massimini M, Tononi G. Theoretical approaches to the diagnosis of altered states of consciousness. Prog Brain Res. 2009;177:383–98. doi: 10.1016/S0079-6123(09)17727-0. [DOI] [PubMed] [Google Scholar]
- 51.Ferrarelli F, Massimini M, Sarasso S, Casali A, Riedner BA, Angelini G, Tononi G, Pearce RA. Breakdown in cortical effective connectivity during midazolam-induced loss of consciousness. Proc Natl Acad Sci U S A. 2010;107:2681–6. doi: 10.1073/pnas.0913008107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Massimini M, Ferrarelli F, Huber R, Esser SK, Singh H, Tononi G. Breakdown of cortical effective connectivity during sleep. Science. 2005;309:2228–32. doi: 10.1126/science.1117256. [DOI] [PubMed] [Google Scholar]
- 53.Horovitz SG, Braun AR, Carr WS, Picchioni D, Balkin TJ, Fukunaga M, Duyn JH. Decoupling of the brain’s default mode network during deep sleep. Proc Natl Acad Sci U S A. 2009;106:11376–81. doi: 10.1073/pnas.0901435106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Larson-Prior LJ, Zempel JM, Nolan TS, Prior FW, Snyder AZ, Raichle ME. Cortical network functional connectivity in the descent to sleep. Proc Natl Acad Sci U S A. 2009;106:4489–94. doi: 10.1073/pnas.0900924106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Mhuircheartaigh RN, Rosenorn-Lanng D, Wise R, Jbabdi S, Rogers R, Tracey I. Cortical and subcortical connectivity changes during decreasing levels of consciousness in humans: a functional magnetic resonance imaging study using propofol. J Neurosci. 2010;30:9095–102. doi: 10.1523/JNEUROSCI.5516-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Boveroux P, Vanhaudenhuyse A, Bruno MA, Noirhomme Q, Lauwick S, Luxen A, Degueldre C, Plenevaux A, Schnakers C, Phillips C, Brichant JF, Bonhomme V, Maquet P, Greicius MD, Laureys S, Boly M. Breakdown of within-and between-network resting state functional magnetic resonance imaging connectivity during propofol-induced loss of consciousness. Anesthesiology. 2010;113:1038–53. doi: 10.1097/ALN.0b013e3181f697f5. [DOI] [PubMed] [Google Scholar]
- 57.Vincent JL, Patel GH, Fox MD, Snyder AZ, Baker JT, Van Essen DC, Zempel JM, Snyder LH, Corbetta M, Raichle ME. Intrinsic functional architecture in the anaesthetized monkey brain. Nature. 2007;447:83–6. doi: 10.1038/nature05758. [DOI] [PubMed] [Google Scholar]
- 58.MacKay EC, Sleigh JW, Voss LJ, Barnard JP. Episodic waveforms in the electroencephalogram during general anaesthesia: a study of patterns of response to noxious stimuli. Anaesth Intensive Care. 2010;38:102–12. doi: 10.1177/0310057X1003800118. [DOI] [PubMed] [Google Scholar]
- 59.Nir Y, Staba RJ, Andrillon T, Vyazovskiy VV, Cirelli C, Fried I, Tononi G. Regional slow waves and spindles in human sleep. Neuron. 2011;70:153–69. doi: 10.1016/j.neuron.2011.02.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Esser SK, Hill S, Tononi G. Breakdown of effective connectivity during slow wave sleep: investigating the mechanism underlying a cortical gate using large-scale modeling. J Neurophysiol. 2009;102:2096–111. doi: 10.1152/jn.00059.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Hudetz AG. Are we unconscious during general anesthesia? Int Anesthesiol Clin. 2008;46:25–42. doi: 10.1097/AIA.0b013e3181755db5. [DOI] [PubMed] [Google Scholar]
- 62.Wehrle R, Kaufmann C, Wetter TC, Holsboer F, Auer DP, Pollmacher T, Czisch M. Functional microstates within human REM sleep: first evidence from fMRI of a thalamocortical network specific for phasic REM periods. Eur J Neurosci. 2007;25:863–71. doi: 10.1111/j.1460-9568.2007.05314.x. [DOI] [PubMed] [Google Scholar]
- 63.John J, Wu MF, Boehmer LN, Siegel JM. Cataplexy-active neurons in the hypothalamus: implications for the role of histamine in sleep and waking behavior. Neuron. 2004;42:619–34. doi: 10.1016/s0896-6273(04)00247-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Saper CB, Scammell TE, Lu J. Hypothalamic regulation of sleep and circadian rhythms. Nature. 2005;437:1257–63. doi: 10.1038/nature04284. [DOI] [PubMed] [Google Scholar]
- 65.Gompf H, Chen J, Sun Y, Yanagisawa M, Aston-Jones G, Kelz MB. Halothane-induced hypnosis is not accompanied by inactivation of orexinergic output in rodents. Anesthesiology. 2009;111:1001–9. doi: 10.1097/ALN.0b013e3181b764b3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Lu J, Nelson LE, Franks N, Maze M, Chamberlin NL, Saper CB. Role of endogenous sleep-wake and analgesic systems in anesthesia. J Comp Neurol. 2008;508:648–62. doi: 10.1002/cne.21685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Nelson LE, Lu J, Guo T, Saper CB, Franks NP, Maze M. The alpha2-adrenoceptor agonist dexmedetomidine converges on an endogenous sleep-promoting pathway to exert its sedative effects. Anesthesiology. 2003;98:428–36. doi: 10.1097/00000542-200302000-00024. [DOI] [PubMed] [Google Scholar]
- 68.Zecharia AY, Nelson LE, Gent TC, Schumacher M, Jurd R, Rudolph U, Brickley SG, Maze M, Franks NP. The involvement of hypothalamic sleep pathways in general anesthesia: testing the hypothesis using the GABAA receptor beta3N265M knock-in mouse. J Neurosci. 2009;29:2177–87. doi: 10.1523/JNEUROSCI.4997-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Kelz MB, Sun Y, Chen J, Cheng Meng Q, Moore JT, Veasey SC, Dixon S, Thornton M, Funato H, Yanagisawa M. An essential role for orexins in emergence from general anesthesia. Proc Natl Acad Sci U S A. 2008;105:1309–14. doi: 10.1073/pnas.0707146105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Coull JT, Buchel C, Friston KJ, Frith CD. Noradrenergically mediated plasticity in a human attentional neuronal network. Neuroimage. 1999;10:705–15. doi: 10.1006/nimg.1999.0513. [DOI] [PubMed] [Google Scholar]
- 71.Maquet P, Ruby P, Maudoux A, Albouy G, Sterpenich V, Dang-Vu T, Desseilles M, Boly M, Perrin F, Peigneux P, Laureys S. Human cognition during REM sleep and the activity profile within frontal and parietal cortices: a reappraisal of functional neuroimaging data. Prog Brain Res. 2005;150:219–27. doi: 10.1016/S0079-6123(05)50016-5. [DOI] [PubMed] [Google Scholar]
- 72.Sara SJ. The locus coeruleus and noradrenergic modulation of cognition. Nature reviews Neuroscience. 2009;10:211–23. doi: 10.1038/nrn2573. [DOI] [PubMed] [Google Scholar]
- 73.Berridge CW, Waterhouse BD. The locus coeruleus-noradrenergic system: modulation of behavioral state and state-dependent cognitive processes. Brain research Brain Res Rev. 2003;42:33–84. doi: 10.1016/s0165-0173(03)00143-7. [DOI] [PubMed] [Google Scholar]
- 74.Hasselmo ME, Linster C, Patil M, Ma D, Cekic M. Noradrenergic suppression of synaptic transmission may influence cortical signal-to-noise ratio. J Neurophys. 1997;77:3326–39. doi: 10.1152/jn.1997.77.6.3326. [DOI] [PubMed] [Google Scholar]
- 75.Schrouff J, Perlbarg V, Boly M, Marrelec G, Boveroux P, Vanhaudenhuyse A, Bruno MA, Laureys S, Phillips C, Pelegrini-Issac M, Maquet P, Benali H. Brain functional integration decreases during propofol-induced loss of consciousness. NeuroImage. 2011;57:198–205. doi: 10.1016/j.neuroimage.2011.04.020. [DOI] [PubMed] [Google Scholar]
- 76.Sanders RD, Maze M. Contribution of sedative-hypnotic agents to delirium via modulation of the sleep pathway. Can J Anaesth. 2011;58:149–156. doi: 10.1007/s12630-010-9421-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Angel A. The G. L. Brown lecture. Adventures in anaesthesia. Exp Physiol. 1991;76:1–38. doi: 10.1113/expphysiol.1991.sp003471. [DOI] [PubMed] [Google Scholar]
- 78.Le Masson G, Renaud-Le Masson S, Debay D, Bal T. Feedback inhibition controls spike transfer in hybrid thalamic circuits. Nature. 2002;417:854–8. doi: 10.1038/nature00825. [DOI] [PubMed] [Google Scholar]
- 79.Franks NP. General anaesthesia: from molecular targets to neuronal pathways of sleep and arousal. Nat Rev Neurosci. 2008;9:370–86. doi: 10.1038/nrn2372. [DOI] [PubMed] [Google Scholar]
- 80.McKay ID, Voss LJ, Sleigh JW, Barnard JP, Johannsen EK. Pharmacokinetic-pharmacodynamic modeling the hypnotic effect of sevoflurane using the spectral entropy of the electroencephalogram. Anesth Analg. 2006;102:91–7. doi: 10.1213/01.ane.0000184825.65124.24. [DOI] [PubMed] [Google Scholar]
- 81.Sleigh JW, Leslie K, Voss L. The effect of skin incision on the electroencephalogram during general anesthesia maintained with propofol or desflurane. J Clin Monit Comput. 2010;24:307–18. doi: 10.1007/s10877-010-9251-3. [DOI] [PubMed] [Google Scholar]
- 82.Krout KE, Belzer RE, Loewy AD. Brainstem projections to midline and intralaminar thalamic nuclei of the rat. J Comp Neurol. 2002;448:53–101. doi: 10.1002/cne.10236. [DOI] [PubMed] [Google Scholar]
- 83.Bonhomme V, Maquet P, Phillips C, Plenevaux A, Hans P, Luxen A, Lamy M, Laureys S. The effect of clonidine infusion on distribution of regional cerebral blood flow in volunteers. Anesth Analg. 2008;106:899–909. doi: 10.1213/ane.0b013e3181619685. [DOI] [PubMed] [Google Scholar]
- 84.Huupponen E, Maksimow A, Lapinlampi P, Sarkela M, Saastamoinen A, Snapir A, Scheinin H, Scheinin M, Merilainen P, Himanen SL, Jaaskelainen S. Electroencephalogram spindle activity during dexmedetomidine sedation and physiological sleep. Acta Anaesthesiol Scand. 2008;52:289–94. doi: 10.1111/j.1399-6576.2007.01537.x. [DOI] [PubMed] [Google Scholar]
- 85.Franks NP, Zecharia AY. Sleep and general anesthesia. Can J Anaesth. 2011;58:139–48. doi: 10.1007/s12630-010-9420-3. [DOI] [PubMed] [Google Scholar]
- 86.Sukhotinsky I, Hopkins DA, Lu J, Saper CB, Devor M. Movement suppression during anesthesia: neural projections from the mesopontine tegmentum to areas involved in motor control. J Comp Neurol. 2005;489:425–48. doi: 10.1002/cne.20636. [DOI] [PubMed] [Google Scholar]
- 87.Devor M, Zalkind V. Reversible analgesia, atonia, and loss of consciousness on bilateral intracerebral microinjection of pentobarbital. Pain. 2001;94:101–12. doi: 10.1016/S0304-3959(01)00345-1. [DOI] [PubMed] [Google Scholar]
- 88.Antognini JF, Schwartz K. Exaggerated anesthetic requirements in the preferentially anesthetized brain. Anesthesiology. 1993;79:1244–9. doi: 10.1097/00000542-199312000-00015. [DOI] [PubMed] [Google Scholar]
- 89.Panula P, Pirvola U, Auvinen S, Airaksinen MS. Histamine-immunoreactive nerve fibers in the rat brain. Neuroscience. 1989;28:585–610. doi: 10.1016/0306-4522(89)90007-9. [DOI] [PubMed] [Google Scholar]
- 90.Alkire MT, Gruver R, Miller J, McReynolds JR, Hahn EL, Cahill L. Neuroimaging analysis of an anesthetic gas that blocks human emotional memory. Proc Natl Acad Sci U S A. 2008;105:1722–7. doi: 10.1073/pnas.0711651105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Redgrave P, Rodriguez M, Smith Y, Rodriguez-Oroz MC, Lehericy S, Bergman H, Agid Y, DeLong MR, Obeso JA. Goal-directed and habitual control in the basal ganglia: implications for Parkinson’s disease. Nat Rev Neurosci. 2010;11:760–72. doi: 10.1038/nrn2915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Morrison SE, Salzman CD. Re-valuing the amygdala. Curr Opin Neurobiol. 2010;20:221–30. doi: 10.1016/j.conb.2010.02.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Bell-McGinty S, Habeck C, Hilton HJ, Rakitin B, Scarmeas N, Zarahn E, Flynn J, DeLaPaz R, Basner R, Stern Y. Identification and differential vulnerability of a neural network in sleep deprivation. Cereb Cortex. 2004;14:496–502. doi: 10.1093/cercor/bhh011. [DOI] [PubMed] [Google Scholar]
- 94.Rampersad SE, Mulroy MF. A case of awareness despite an “adequate depth of anesthesia” as indicated by a Bispectral Index monitor. Anesth Analg. 2005;100:1363–4. doi: 10.1213/01.ANE.0000148121.84560.8D. [DOI] [PubMed] [Google Scholar]
- 95.Samuelsson P, Brudin L, Sandin RH. BIS does not predict dreams reported after anaesthesia. Acta Anaesthesiol Scand. 2008;52:810–4. doi: 10.1111/j.1399-6576.2008.01633.x. [DOI] [PubMed] [Google Scholar]
- 96.Voss L, Sleigh J. Monitoring consciousness: the current status of EEG-based depth of anaesthesia monitors. Best Pract Res Clin Anaesthesiol. 2007;21:313–25. doi: 10.1016/j.bpa.2007.04.003. [DOI] [PubMed] [Google Scholar]
- 97.Imas OA, Ropella KM, Ward BD, Wood JD, Hudetz AG. Volatile anesthetics disrupt frontal-posterior recurrent information transfer at gamma frequencies in rat. Neurosci Lett. 2005;387:145–50. doi: 10.1016/j.neulet.2005.06.018. [DOI] [PubMed] [Google Scholar]
- 98.Hudetz AG. Feedback suppression in anesthesia. Is it reversible? Conscious Cogn. 2009;18:1079–81. doi: 10.1016/j.concog.2009.08.004. [DOI] [PubMed] [Google Scholar]
- 99.Hudetz AG, Vizuete JA, Imas OA. Desflurane selectively suppresses long-latency cortical neuronal response to flash in the rat. Anesthesiology. 2009;111:231–9. doi: 10.1097/ALN.0b013e3181ab671e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Boly M, Garrido MI, Gosseries O, Bruno MA, Boveroux P, Schnakers C, Massimini M, Litvak V, Laureys S, Friston K. Preserved feedforward but impaired top-down processes in the vegetative state. Science. 2011;332:858–62. doi: 10.1126/science.1202043. [DOI] [PubMed] [Google Scholar]
- 101.Ro T, Breitmeyer B, Burton P, Singhal NS, Lane D. Feedback contributions to visual awareness in human occipital cortex. Curr Biol. 2003;13:1038–41. doi: 10.1016/s0960-9822(03)00337-3. [DOI] [PubMed] [Google Scholar]
- 102.Lee U, Kim S, Noh GJ, Choi BM, Hwang E, Mashour GA. The directionality and functional organization of frontoparietal connectivity during consciousness and anesthesia in humans. Conscious Cogn. 2009;18:1069–78. doi: 10.1016/j.concog.2009.04.004. [DOI] [PubMed] [Google Scholar]
- 103.Lee U, Mashour GA, Kim S, Noh GJ, Choi BM. Propofol induction reduces the capacity for neural information integration: implications for the mechanism of consciousness and general anesthesia. Conscious Cogn. 2009;18:56–64. doi: 10.1016/j.concog.2008.10.005. [DOI] [PubMed] [Google Scholar]
- 104.Atienza M, Cantero JL, Gomez CM. The mismatch negativity component reveals the sensory memory during REM sleep in humans. Neuro lett. 1997;237:21–4. doi: 10.1016/s0304-3940(97)00798-2. [DOI] [PubMed] [Google Scholar]
- 105.Sanders RD. Hypothesis for the pathophysiology of delirium: Role of baseline brain network connectivity and changes in inhibitory tone. Med Hypotheses. 2011;77:140–3. doi: 10.1016/j.mehy.2011.03.048. [DOI] [PubMed] [Google Scholar]
- 106.Friedman EB, Sun Y, Moore JT, Hung HT, Meng QC, Perera P, Joiner WJ, Thomas SA, Eckenhoff RG, Sehgal A, Kelz MB. A conserved behavioral state barrier impedes transitions between anesthetic-induced unconsciousness and wakefulness: evidence for neural inertia. PLoS One. 2010;5:e11903. doi: 10.1371/journal.pone.0011903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Pandharipande PP, Pun BT, Herr DL, Maze M, Girard TD, Miller RR, Shintani AK, Thompson JL, Jackson JC, Deppen SA, Stiles RA, Dittus RS, Bernard GR, Ely EW. Effect of sedation with dexmedetomidine vs lorazepam on acute brain dysfunction in mechanically ventilated patients: the MENDS randomized controlled trial. JAMA. 2007;298:2644–53. doi: 10.1001/jama.298.22.2644. [DOI] [PubMed] [Google Scholar]
- 108.Pandharipande PP, Sanders RD, Girard TD, McGrane S, Thompson JL, Shintani AK, Herr DL, Maze M, Ely EW. Effect of dexmedetomidine versus lorazepam on outcome in patients with sepsis: an a priori-designed analysis of the MENDS randomized controlled trial. Crit Care. 2010;14:R38. doi: 10.1186/cc8916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Sandin RH, Enlund G, Samuelsson P, Lennmarken C. Awareness during anaesthesia: a prospective case study. Lancet. 2000;355:707–11. doi: 10.1016/S0140-6736(99)11010-9. [DOI] [PubMed] [Google Scholar]
- 110.Sebel PS, Bowdle TA, Ghoneim MM, Rampil IJ, Padilla RE, Gan TJ, Domino KB. The incidence of awareness during anesthesia: a multicenter United States study. Anesth Analg. 2004;99:833–9. doi: 10.1213/01.ANE.0000130261.90896.6C. [DOI] [PubMed] [Google Scholar]
- 111.Tunstall ME. Detecting wakefulness during general anaesthesia for caesarean section. Br Med J. 1977;1:1321. doi: 10.1136/bmj.1.6072.1321-a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Tunstall ME, Sheikh A. Comparison of 1.5% enflurane with 1. 25% isoflurane in oxygen for caesarean section: avoidance of awareness without nitrous oxide. Br J Anaesth. 1989;62:138–43. doi: 10.1093/bja/62.2.138. [DOI] [PubMed] [Google Scholar]
- 113.Russell IF. Comparison of wakefulness with two anaesthetic regimens. Total i.v. v. balanced anaesthesia. Br J Anaesth. 1986;58:965–8. doi: 10.1093/bja/58.9.965. [DOI] [PubMed] [Google Scholar]
- 114.Russell IF. Midazolam-alfentanil: an anaesthetic? An investigation using the isolated forearm technique. Br J Anaesth. 1993;70:42–6. doi: 10.1093/bja/70.1.42. [DOI] [PubMed] [Google Scholar]
- 115.King H, Ashley S, Brathwaite D, Decayette J, Wooten DJ. Adequacy of general anesthesia for cesarean section. Anesth Analg. 1993;77:84–8. doi: 10.1213/00000539-199307000-00017. [DOI] [PubMed] [Google Scholar]
- 116.Gaitini L, Vaida S, Collins G, Somri M, Sabo E. Awareness detection during caesarean section under general anaesthesia using EEG spectrum analysis. Can J Anaesth. 1995;42:377–81. doi: 10.1007/BF03015480. [DOI] [PubMed] [Google Scholar]
- 117.Russell IF, Wang M. Absence of memory for intraoperative information during surgery under adequate general anaesthesia. Br J Anaesth. 1997;78:3–9. doi: 10.1093/bja/78.1.3. [DOI] [PubMed] [Google Scholar]
- 118.St Pierre M, Landsleitner B, Schwilden H, Schuettler J. Awareness during laryngoscopy and intubation: quantitating incidence following induction of balanced anesthesia with etomidate and cisatracurium as detected with the isolated forearm technique. J Clin Anesth. 2000;12:104–8. doi: 10.1016/s0952-8180(00)00127-6. [DOI] [PubMed] [Google Scholar]
- 119.Slavov V, Motamed C, Massou N, Rebufat Y, Duvaldestin P. Systolic blood pressure, not BIS, is associated with movement during laryngoscopy and intubation. Can J Anaesth. 2002;49:918–21. doi: 10.1007/BF03016874. [DOI] [PubMed] [Google Scholar]
- 120.John ER, Prichep LS. The anesthetic cascade: A theory of how anesthesia suppresses consciousness. Anesthesiology. 2005;102:447–71. doi: 10.1097/00000542-200502000-00030. [DOI] [PubMed] [Google Scholar]