

Abstract
The identity of the cells responsible for the initiation and maintenance of multiple myeloma (MM) remains unclear largely because of the difficulty growing MM cells in vitro and in vivo. MM cell lines and clinical specimens are characterized by malignant plasma cells that express the cell surface antigen syndecan-1 (CD138); however, CD138 expression is limited to terminally differentiated plasma cells during B-cell development. Moreover, circulating B cells that are clonally related to MM plasma cells have been reported in some patients with MM. We found that human MM cell lines contained small (< 5%) subpopulations that lacked CD138 expression and had greater clonogenic potential in vitro than corresponding CD138+ plasma cells. CD138− cells from clinical MM samples were similarly clonogenic both in vitro and in nonobese diabetic/severe combined immunodeficiency (NOD/SCID) mice, whereas CD138+ cells were not. Furthermore, CD138− cells from both cell lines and clinical samples phenotypically resembled postgerminal center B cells, and their clonogenic growth was inhibited by the anti-CD20 monoclonal antibody rituximab. These data suggest that MM “stem cells” are CD138− B cells with the ability to replicate and subsequently differentiate into malignant CD138+ plasma cells.
Introduction
Multiple myeloma (MM) is characterized by the accumulation of malignant plasma cells. Like their normal terminally differentiated counterparts, MM plasma cells appear to be relatively quiescent because the proliferative index and cloning efficiency of bone marrow samples from patients with MM is low.1,2 Therefore, it is unclear whether MM plasma cells have the proliferative capacity required for the initiation, maintenance, and progression of clinical disease. On the other hand, cells phenotypically resembling mature B cells and sharing immunoglobulin gene sequences and idiotype specificity with MM plasma cells have been found in the bone marrow and peripheral blood of patients with MM,3–8 but their role in the pathogenesis of the disease is unclear. Several investigators have proposed that these clonally related B cells represent the proliferating compartment or tumor “stem cells” in MM; however, this remains a matter of debate because these cells have not been functionally characterized.9–14
Syndecan-1 (CD138) is expressed by malignant plasma cells from the majority of MM cell lines and patient specimens.15–17 Furthermore, during normal B-cell development, CD138 expression is highly specific for terminally differentiated normal plasma cells because it is absent on highly proliferative normal plasma-blasts and all earlier B-cell stages.17–20 Therefore, we hypothesized that if clonogenic B cells represent putative MM stem cells, they should lack CD138 expression. Here, we demonstrate that highly clonogenic cells from both human MM cell lines and primary patient samples do not express CD138, but rather markers that are characteristic of B cells. In addition, these cells are inhibited in vitro by rituximab, a monoclonal antibody directed against the B-cell antigen CD20.
Patients, materials, and methods
Cell lines, patient samples, and cell culture
The human MM cell lines RPMI 8226 and NCI-H929 were obtained from the American Type Culture Collection (Manassas, VA). Cells were cultured in complete media (CM) consisting of RPMI 1640, 2 mM L-glutamine, 50 U/mL penicillin, 50 μg/mL streptomycin, and 10% fetal bovine serum (FBS). Clinical bone marrow and peripheral blood samples were obtained from patients with MM or healthy donors granting informed consent as approved by the Johns Hopkins Medical Institutes’ Institutional Review Board. Mononuclear cells were isolated by density centrifugation (Ficoll-Paque; Pharmacia, Piscataway, NJ). CD138+ or CD138− subsets were isolated from mononuclear cells using mouse antihuman CD138 antibodies coupled to magnetic microbeads (Miltenyi Biotec, Auburn, CA) followed by magnetic column selection or depletion, respectively (magnetic-activated cell sorting [MACS], Miltenyi Biotec). Resulting cells were additionally depleted of normal hematopoietic progenitors using mouse antihuman CD34 antibodies (Miltenyi Biotec). Secondary depletion of the CD138−/CD34+ cell population was performed by incubating with mouse antihuman CD45, CD19, CD22, or CD3 monoclonal antibodies coupled to magnetic microbeads and treated as described. Subsequent flow cytometric analysis demonstrated less than 5% contamination by relevant antigen-expressing cells. Clonogenic growth was evaluated by plating cells (1000 cells/mL for cell lines or 1 × 105 to 5 × 105 cells/mL for clinical specimens) in 1 mL 1.2% methylcellulose, 30% bovine serum albumin (BSA), 10−4 M 2-mercaptoethanol, and 2 mM L-glutamine. Methylcellulose cultures assessing clinical MM growth also contained 10% lymphocyte-conditioned media as a source of growth factors.21 Samples were plated in quadruplicate onto 35-mm2 tissue culture dishes and incubated at 37°C and 5% CO2. Colonies consisting of more than 40 cells were scored at 7 days for cell lines and 14 to 21 days for MM colonies from clinical samples. Serial replating was performed by washing plates 3 times with CM and resuspending cells in the original volume of methylcellulose as described.
Fluorescence-activated cell sorting and flow cytometry
Cell lines were initially depleted of necrotic cells by density centrifugation then stained with phycoerythrin (PE)–conjugated mouse antihuman CD138 antibodies (Beckman Coulter, Fullerton, CA) for 30 minutes at 4°C. Cells were washed then resuspended in phosphate-buffered saline (PBS) containing 5 μM propidium iodide (PI; Sigma, St Louis, MO) and analyzed on a FACSVantage SE flow cytometer (Becton Dickinson, Mountain View, CA). Cells were gated to exclude PI+ cells and sorted into CD138− and CD138+ fractions by gating on the lowest and highest 5% PE-expressing cells, respectively. CD138− cells had no detectable expression of CD138 when compared to an isotypic control antibody. Following sorting, the CD138+ and CD138− cell fractions were analyzed using a FACScan flow cytometer (Becton Dickinson) and found to be more than 98% pure. For phenotypic analyses of cell lines, cells were prepared as described, then stained with CD138-PE and fluorescein isothiocyanate (FITC)–conjugated mouse anti-human CD19, CD20, Ki67, κ or λ immunoglobulin light chains, or isotypic control antibodies (BD PharMingen, San Diego, CA). Cells were analyzed by gating on CD138+ or CD138− populations and subsequently evaluating FITC expression.
Detection of EBV
DNA was extracted using the QIAamp DNA Blood Mini Kit (Qiagen, Valencia, CA) according to the manufacturer’s instructions. The presence of Epstein-Barr virus (EBV) was detected by amplifying the BamHI W fragment using the following primers: 5′-CTT TAG AGG CGA ATG GGC GCC A-3′ and 5′-TCC AGG GCC TTC ACT TCG GTC T-3′. DNA (65–100 ng) and 200 nM of each primer were added to 45 μL Platinum PCR Supermix (Invitrogen, Carlsbad, CA). The DNA template was denatured and the DNA polymerase was activated by heating at 94°C for 2 minutes. Then 35 cycles of amplification were performed as follows: 94°C for 30 seconds, 55°C for 30 seconds, and 72°C for 1 minute. A final extension at 72°C for 10 minutes was also performed. The polymerase chain reaction (PCR) products were separated in a 1% agarose gel.
Transplantation of MM cells into NOD/SCID mice
Experiments using nonobese diabetic/severe combined immunodeficiency (NOD/SCID) mice were approved by the Johns Hopkins Medical Institutes’ Animal Care Committee. NOD/SCID mice were bred and maintained in the Johns Hopkins animal care facility. Six- to 10-week-old mice received 300 cGy irradiation (84 cGy/min using a 137Cs γ irradiator) 4 to 6 hours prior to intravenous injection via the dorsal tail vein22 of 1 to 10 × 106 CD138+ or CD138− cells isolated as described (see “Cell lines, patient samples, and cell culture”). In addition, cell fractions were depleted of T cells using magnetically conjugated murine antihuman CD3 antibodies (Miltenyi Biotec). Human MM cells were suspended in a total volume of 500 μL RPMI 1640 media. Mice were killed at 12 weeks after transplantation, and bone marrow was harvested from the long bones of the 2 posterior limbs by flushing with CM. Cells were evaluated by flow cytometry following cell surface staining with mouse antihuman CD45, CD138, and CD38 antibodies as well as intracellular staining with mouse antihuman κ and λ immunoglobulin light-chain antibodies and the Fix and Perm intracellular staining kit (Caltag Laboratories, Burlingame, CA). Human immunoglobulin was detected from the peripheral blood of killed animals using the antihuman immunoglobulin enzyme-linked immunosorbent assay (ELISA) kit according to the manufacturer’s directions (Southern Biotechnology, Birmingham, AL).
Treatment of clinical MM samples with rituximab
CD138/CD34-depleted cell fractions from bone marrow mononuclear cells (5 × 105) were incubated in CM containing 10 μg/mL rituximab (Idec, San Diego, CA) with or without 10% human serum as a source of complement for 24 hours at 37°C. Cells were washed in CM then evaluated for MM colony formation as described (see “Cell lines, patient samples, and cell culture”).
Statistical analysis
Results are presented as the mean ± SEM. Comparisons between groups were performed using a 2-tailed, paired Student t test or analysis of variance (ANOVA) as indicated.
Results
Distinct phenotypic and functional cell populations are present in human MM cell lines
Recent studies have suggested that both human23 and murine24,25 MM cell lines can demonstrate heterogeneous expression of several MM-related cell surface antigens. Therefore, we analyzed the expression of CD138 in 2 distinct human MM cell lines, RPMI 8226 and NCI-H929. As expected, the majority of cells in both lines expressed high levels of CD138; however, each cell line contained a distinct population of CD138− cells that represented approximately 2% to 5% of total cells (Figure 1A). To examine the clonogenic capacity of these subpopulations, we isolated CD138+ and CD138− cells from both cell lines by FACS and examined each subpopulation for colony formation in methylcellulose. To examine self-renewal potential, colonies were scored weekly, collected, and then washed and serially replated. Initially, the RPMI 8226 CD138+ cells yielded 152 ± 17 colonies/1000 cells plated, and the CD138− cells 16 ± 6 colonies, whereas the NCI-H929 line generated 111 ± 23 colonies and 11 ± 3 colonies, respectively. Although the initial plating efficiency of CD138+ cells was approximately 10-fold higher than CD138− cells, CD138− cells from both cell lines underwent significantly greater clonogenic expansion than CD138+ cells during serial replating (Figure 1B).
Figure 1. Clonogenicity of MM cell lines by CD138 expression.

(A) Flow cytometric evaluation of CD138 expression by RPMI 8226 (λ light chain restricted) and NCI-H929 cells (κ light chain restricted). * denotes CD138− population. (B) Clonogenic expansion of CD138+ and CD138− RPMI 8226 and NCI-H929 cells during serial replating. Results are the mean ± SEM of 3 separate experiments for each cell line and represent the fold clonogenic expansion expressed as a ratio of number of colonies scored compared to the previous plating. P < .02 following the fifth and sixth transfer of both cell lines by Student t test. X-axis represents round of serial replating. (C) Flow cytometric evaluation comparing additional antigens expressed by CD138+ and CD138− RPMI 8226 and NCI-H929 cells.
CD138− cells from MM cell lines express B-cell antigens
We further characterized the phenotype of CD138− cells from both cell lines by flow cytometry and found that CD138− cells expressed CD19, CD20, as well as surface immunoglobulin with light-chain restriction that matched previously published findings for these 2 cell lines (Figure 1C).26 These antigens are normally expressed on mature B cells but not plasma cells, and accordingly, CD138+ cells expressed none of these markers (Figure 1C). Furthermore, CD138− cells expressed higher levels of the proliferation marker Ki67 than CD138+ cells (Figure 1C).
Clonogenic MM cells from clinical samples are CD138−
We examined the in vitro colony formation of CD138+ and CD138− cells from bone marrow specimens of 24 consecutive MM patients by isolating CD138+ and CD138− cell fractions and further depleting them of CD34+ hematopoietic progenitors that complicated the assessment of clonogenic growth (Table 1). We found that the CD138+/CD34− cells from all 24 patients were unable to form colonies in vitro, represented by 3 patients in Figure 2A. Conversely, the CD138−/CD34− cells generated colonies that consisted of morphologically mature plasma cells by 2 to 3 weeks, and in the 3 patients examined, colony formation occurred in a linear fashion with increasing numbers of plated cells (Figure 2A). Flow cytometric evaluation of pooled plasma cell colonies from the CD138−/CD34− cells demonstrated that the resulting CD138+ cells displayed intracellular immunoglobulin light-chain restriction that matched the patients’ MM (Figure 2B–C). Clonogenic growth could theoretically result from EBV transformation; however, analysis of these pooled plasma cell colonies from 3 patients revealed no EBV DNA sequences by PCR. Serial replating was performed on cells derived from colonies of 3 patients and demonstrated continued colony formation (Figure 2D).
Table 1.
In vitro clonogenic MM growth of CD138−/CD34− cells isolated from primary patient samples
| No. | Colony formation, no. (%) | No. of colonies/105 cells, mean (range) | |
|---|---|---|---|
|
MM
| |||
| BM | 24 | 21 (88) | 24 (4–43) |
|
| |||
| PB | 4 | 2 (50) | 7.5 (3–12) |
|
PCL
| |||
| BM | 2 | 2 (100) | 42 (17–66) |
|
| |||
| PB | 2 | 2 (100) | 30 (12–49) |
|
MGUS
| |||
| BM | 6 | 3 (50) | 5 (3–8) |
|
| |||
| Total | 32 | 26 (81) | ND |
P < .01 for comparison of no. of colonies/105 cells from bone marrow samples in MM, PCL, and MGUS groups by ANOVA.
BM indicates bone marrow; PB, peripheral blood; PCL, plasma cell leukemia or advanced MM; MGUS, monoclonal gammopathy of unknown significance; and ND, not determined.
Figure 2. Clonogenic growth of MM cells from clinical specimens.

(A) MM colony formation of CD138+ and CD138− cells from 3 clinical samples plated at various densities. (B) Flow cytometric evaluation of colonies derived from CD138−/CD34− cells. The top 2 panels show the initial clinical marrow specimen; the middle panels, the CD138-depleted cells prior to plating; and the bottom panels, the pooled colonies after 21 days from a representative MM patient. (C) Flow cytometric evaluation of pooled colonies from 3 additional patients. (D) Colony formation of CD138− clinical MM samples during serial replating.
The bone marrow samples were examined from patients with various stages of MM (stage I, 5 patients; stage II, 8 patients; stage III, 11 patients), and we found MM colony formation in 21 of 24 (88%; Table 1). In contrast, we observed no plasma cell colonies in 5 consecutive normal bone marrow specimens (data not shown). In addition, we obtained MM colonies from peripheral blood samples in 2 of 4 patients using the same methodology (Table 1). We also observed malignant plasma cell colony growth from the blood and bone marrow of patients with advanced MM and circulating plasma cells as well as 3 of 6 bone marrow samples from patients with monoclonal gammopathy of unknown significance (Table 1). Furthermore, the cloning efficiency positively correlated with disease stage (Table 1; P < .01).
As a second measure of clonogenic growth, we examined the ability of CD138+ or CD138− cells to be engrafted in immunodeficient mice. CD138+/CD34− and CD138−/CD34− cells were isolated from bone marrow mononuclear cells of 4 patients and injected intravenously into NOD/SCID mice. Human engraftment was not detected in any of the mice injected with CD138+ cells (data not shown). However, CD138− cells from a single patient were engrafted in 3 of 3 animals as evidenced by the presence of human CD138+ cells with light-chain restriction matching the original patient sample within the murine bone marrow (Figure 3A). Analysis of the peripheral blood also demonstrated the presence of light chain–restricted immunoglobulin that matched the patient’s M protein (Figure 3B).
Figure 3. Engraftment of MM CD138− cells in NOD/SCID mice.
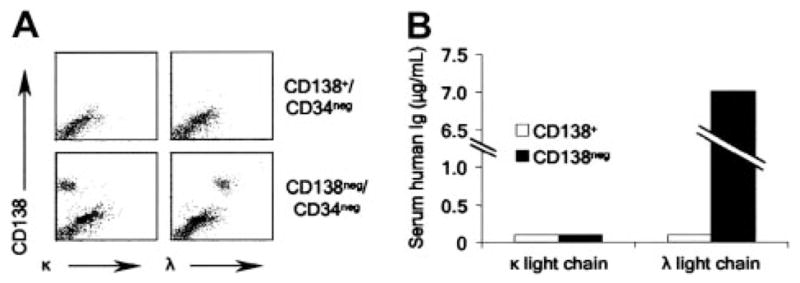
(A) Flow cytometric evaluation of murine bone marrow from animals injected with CD138−/CD34− or CD138+/CD34− cells from a single patient. (B) Serum human immunoglobulin levels measured by ELISA in mice injected with CD138−/CD34− or CD138+/CD34− cells from a single patient.
Clonogenic MM progenitors from clinical samples express B-cell antigens
To further define the phenotype of clonogenic MM cells from primary patient samples, we analyzed CD138−/CD34− cell populations from bone marrow samples of 12 patients with MM by depleting additional cell populations using monoclonal antibodies directed against cell surface antigens. CD45 is present on the majority of human lymphoid cells but absent on normal, and most malignant, plasma cells,18,27,28 and we found that CD45 depletion significantly decreased in vitro MM colony formation (Figure 4). Similarly, antibodies directed against the B-cell antigens CD19 and CD22 significantly decreased clonogenic MM growth, whereas antibody against the T-cell antigen CD3 had little effect on colony formation (Figure 4).
Figure 4. MM colony formation of CD138−/CD34− cells following antibody-mediated cell depletion.
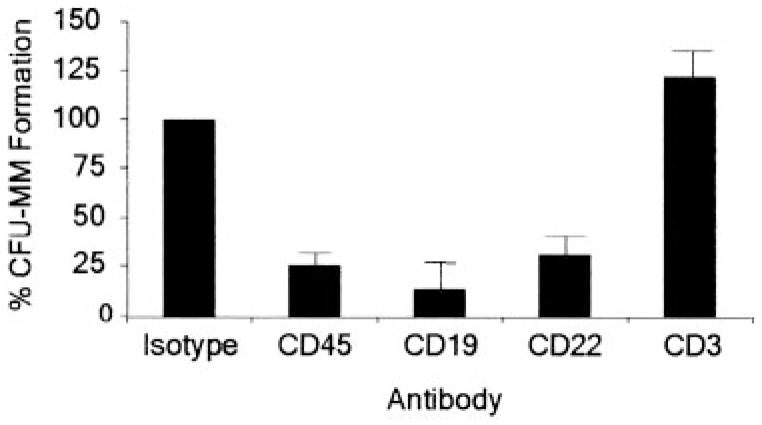
Results are presented as a percentage of the untreated control group that was depleted of CD34+ and CD138+ cells. P < .01 for CD45+, CD19+, and CD22+ cell depletions calculated by Student t test comparing each group with the untreated control (n = 12). CFU indicates colony-forming units.
Because the humanized monoclonal antibody rituximab has activity against most B-cell malignancies,29 we examined its effect on clonogenic MM growth. CD138−/CD34− cells from clinical MM bone marrow samples were incubated with rituximab prior to plating in methylcellulose. Rituximab significantly (P < 0.01) inhibited clonogenic MM growth compared to untreated controls (Figure 5). Furthermore, the addition of human serum as a source of complement further enhanced the activity of rituximab against MM progenitors in vitro (Figure 5).
Figure 5. Effect of rituximab on clonogenic MM progenitors in vitro.

MM colony formation following treatment of bone marrow mononuclear cells depleted of CD34+ and CD138+ cells with rituximab or 10% human serum (compl.). Results are presented as a percentage of the untreated control group. P < .01 for the rituximab and rituximab plus complement groups compared to the untreated (control) group by Student t test (n = 3).
Discussion
The identity of the MM cell of origin has remained controversial, at least in part because only a minority of MM cells are clonogenic in vivo and in vitro.2 Despite being one of the first cancers grown in semisolid clonogenic assays,2,30 MM growth in vitro has been difficult to reproduce.31 There have also been several reports of MM growth in immunodeficient mice, but again, growth has generally been inconsistent or nonquantitative.32–35 CD138 is the most specific marker for normal and MM plasma cells. However, normal CD138+ plasma cells appear to be terminally differentiated and unable to proliferate,20 and there have been few studies using this marker to study the proliferative capacity of MM cells. Although reports have described the engraftment of immunodeficient mice with CD138+ cells from clinical MM specimens, these samples are derived from cases with advanced disease or plasma cell leukemia in which aberrant expression of cell surface antigens may be expected.32,35 Furthermore, the quiescent, but most invasive, cells from a murine model of MM are CD138−, even though the majority of the tumor cells are CD138+.24
The low clonogenicity of many cancers, including MM, could be explained by all cancer cells’ having the ability to proliferate but only at a low rate. However, in many cancers it appears that only a small subset of cells retains the capacity to self-renew and proliferate.36–38 Chronic myeloid leukemia (CML) perhaps best illustrates a malignancy arising from cancer stem cells. CML is characterized by increased numbers of blood and bone marrow cells that display variable degrees of myeloid differentiation. Although these mature cells form the bulk of the tumor mass, they lack significant self-renewal potential and are unable to propagate the disease. In contrast, the neoplastic progenitors responsible for the initiation and maintenance of CML phenotypically resemble normal hematopoietic stem cells.39 Furthermore, rare cancer stem cells that are phenotypically distinct from the differentiated progeny that characterize a particular disease have been described in acute myeloid leukemia,40 acute lymphocytic leukemia,41 myelodysplastic syndrome,42 and breast cancer.43
Our data suggest that MM is another example in which cancer stem cells are a rare cell population that is distinct from the differentiated cells that comprise the bulk of the disease. We found that highly clonogenic MM cells lack CD138 expression typical of plasma cells and, unlike plasmablasts, express CD20 and surface immunoglobulin.18,44 Given that MM cells exhibit somatic hyper-mutation of their immunoglobulin genes without intraclonal variation,5,45 our data suggest that MM arises at the level of postgerminal center B cells, similar to most B-cell non-Hodgkin lymphomas.46 It is likely that eliminating the terminally differentiated CD138+ plasma cells enriched for clonogenic MM progenitors and allowed their detection in most patients with MM. The pooled MM colonies consisted entirely of CD138+ cells with monoclonal light-chain restriction that matched the patients’ M protein. Assaying for immunoglobulin light-chain expression should be the most specific way to confirm that the colonies arose from the MM clone because it quantifies the number of cells expressing the clonotypic light chain. Other methods, such as PCR-based sequence analysis of the complementary-determining region 3 of the immunoglobulin heavy-chain gene, are less useful in this regard because their high sensitivity may detect small contaminating cell populations.
Our finding that rituximab inhibits in vitro clonogenic MM growth is in contrast to a recent report describing the lack of clinical efficacy of 4 to 8 weeks of this antibody in the majority of patients with MM when assessed at 3 to 6 months.47 Standard clinical parameters used to assess response (ie, bone marrow plasmacytosis and monoclonal immunoglobulin level) primarily reflect the effects of treatment on MM plasma cells not MM progenitors. Accordingly, responses were seen only in patients whose MM plasma cells expressed CD20.47 Because both normal and MM plasma cells, although terminally differentiated, are long-lived,1,48 therapies that selectively target MM stem cells may not demonstrate an immediate clinical effect using standard response criteria until the remaining plasma cells gradually undergo spontaneous apoptosis. Therefore, potentially useful therapies with activity primarily against MM stem cells could be prematurely abandoned. Conversely, therapies that target MM plasma cells may lead to rapid clinical improvement but are unlikely to be curative unless MM stem cells are also eliminated.
Acknowledgments
Supported in part by National Institutes of Health grants P01CA70970 and P01CA15396. W.M. is a fellow of the Leukemia Lymphoma Society.
References
- 1.Drewinko B, Alexanian R, Boyer H, Barlogie B, Rubinow SI. The growth fraction of human myeloma cells. Blood. 1981;57:333–338. [PubMed] [Google Scholar]
- 2.Hamburger A, Salmon SE. Primary bioassay of human myeloma stem cells. J Clin Invest. 1977;60:846–854. doi: 10.1172/JCI108839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Pilarski LM, Jensen GS. Monoclonal circulating B cells in multiple myeloma: a continuously differentiating, possibly invasive, population as defined by expression of CD45 isoforms and adhesion molecules. Hematol Oncol Clin North Am. 1992;6:297–322. [PubMed] [Google Scholar]
- 4.Billadeau D, Ahmann G, Greipp P, Van Ness B. The bone marrow of multiple myeloma patients contains B cell populations at different stages of differentiation that are clonally related to the malignant plasma cell. J Exp Med. 1993;178:1023–1031. doi: 10.1084/jem.178.3.1023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bakkus MH, Van RI, Van Camp B, Thielemans K. Evidence that the clonogenic cell in multiple myeloma originates from a pre-switched but somatically mutated B cell. Br J Haematol. 1994;87:68–74. doi: 10.1111/j.1365-2141.1994.tb04872.x. [DOI] [PubMed] [Google Scholar]
- 6.Berenson JR, Vescio RA, Hong CH, et al. Multiple myeloma clones are derived from a cell late in B lymphoid development. Curr Top Microbiol Immunol. 1995;194:25–33. doi: 10.1007/978-3-642-79275-5_4. [DOI] [PubMed] [Google Scholar]
- 7.Rasmussen T, Kastrup J, Knudsen LM, Johnsen HE. High numbers of clonal CD19+ cells in the peripheral blood of a patient with multiple myeloma. Br J Haematol. 1999;105:265–267. [PubMed] [Google Scholar]
- 8.Szczepek AJ, Seeberger K, Wizniak J, et al. A high frequency of circulating B cells share clonotypic Ig heavy-chain VDJ rearrangements with autologous bone marrow plasma cells in multiple myeloma, as measured by single-cell and in situ reverse transcriptase-polymerase chain reaction. Blood. 1998;92:2844–2855. [PubMed] [Google Scholar]
- 9.Epstein J. Myeloma stem cell phenotype: implications for treatment. Hematol Oncol Clin North Am. 1997;11:43–49. doi: 10.1016/s0889-8588(05)70414-7. [DOI] [PubMed] [Google Scholar]
- 10.Berenson JR, Vescio RA, Said J. Multiple myeloma: the cells of origin—a two-way street. Leukemia. 1998;12:121–127. doi: 10.1038/sj.leu.2400947. [DOI] [PubMed] [Google Scholar]
- 11.Mitterer M, Oduncu F, Lanthaler AJ, et al. The relationship between monoclonal myeloma precursor B cells in the peripheral blood stem cell harvests and the clinical response of multiple myeloma patients. Br J Haematol. 1999;106:737–743. doi: 10.1046/j.1365-2141.1999.01603.x. [DOI] [PubMed] [Google Scholar]
- 12.Davies FE, Rawstron AC, Owen RG, Morgan GJ. Controversies surrounding the clonogenic origin of multiple myeloma. Br J Haematol. 2000;110:240–241. doi: 10.1046/j.1365-2141.2000.02072-6.x. [DOI] [PubMed] [Google Scholar]
- 13.Rawstron AC, Barrans SL, Blythe D, et al. In multiple myeloma, only a single stage of neoplastic plasma cell differentiation can be identified by VLA-5 and CD45 expression. Br J Haematol. 2001;113:794–802. doi: 10.1046/j.1365-2141.2001.02730.x. [DOI] [PubMed] [Google Scholar]
- 14.Joshua D, Pope B, Brown R, et al. Phenotyping primitive plasma cells. Br J Haematol. 2002;117:252–253. doi: 10.1046/j.1365-2141.2002.3406_5.x. [DOI] [PubMed] [Google Scholar]
- 15.Chilosi M, Adami F, Lestani M, et al. CD138/syndecan-1: a useful immunohistochemical marker of normal and neoplastic plasma cells on routine trephine bone marrow biopsies. Mod Pathol. 1999;12:1101–1106. [PubMed] [Google Scholar]
- 16.Wei A, Juneja S. Bone marrow immunohistology of plasma cell neoplasms. J Clin Pathol. 2003;56:406–411. doi: 10.1136/jcp.56.6.406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Wijdenes J, Vooijs WC, Clement C, et al. A plasmocyte selective monoclonal antibody (B-B4) recognizes syndecan-1. Br J Haematol. 1996;94:318–323. doi: 10.1046/j.1365-2141.1996.d01-1811.x. [DOI] [PubMed] [Google Scholar]
- 18.Jego G, Robillard N, Puthier D, et al. Reactive plasmacytoses are expansions of plasmablasts retaining the capacity to differentiate into plasma cells. Blood. 1999;94:701–712. [PubMed] [Google Scholar]
- 19.Jego G, Bataille R, Pellat-Deceunynck C. Inter-leukin-6 is a growth factor for nonmalignant human plasmablasts. Blood. 2001;97:1817–1822. doi: 10.1182/blood.v97.6.1817. [DOI] [PubMed] [Google Scholar]
- 20.Calame KL. Plasma cells: finding new light at the end of B cell development. Nat Immunol. 2001;2:1103–1108. doi: 10.1038/ni1201-1103. [DOI] [PubMed] [Google Scholar]
- 21.Bedi A, Zehnbauer BA, Barber JP, Sharkis SJ, Jones RJ. Inhibition of apoptosis by BCR-ABL in chronic myeloid leukemia. Blood. 1994;83:2038–2044. [PubMed] [Google Scholar]
- 22.Lumkul R, Gorin NC, Malehorn MT, et al. Human AML cells in NOD/SCID mice: engraftment potential and gene expression. Leukemia. 2002;16:1818–1826. doi: 10.1038/sj.leu.2402632. [DOI] [PubMed] [Google Scholar]
- 23.Mahmoud MS, Ishikawa H, Fujii R, Kawano MM. Induction of CD45 expression and proliferation in U-266 myeloma cell line by interleukin-6. Blood. 1998;92:3887–3897. [PubMed] [Google Scholar]
- 24.Asosingh K, De Raeve H, Van Riet I, Van Camp B, Vanderkerken K. Multiple myeloma tumor progression in the 5T2MM murine model is a multistage and dynamic process of differentiation, proliferation, invasion, and apoptosis. Blood. 2003;101:3136–3141. doi: 10.1182/blood-2002-10-3000. [DOI] [PubMed] [Google Scholar]
- 25.Asosingh K, Willems A, Van Riet I, Van Camp B, Vanderkerken K. Delayed in vivo disease progression is associated with high proportions of CD45+ myeloma cells in the 5T2MM murine model. Cancer Res. 2003;63:3019–3020. [PubMed] [Google Scholar]
- 26.Gooding RP, Bybee A, Cooke F, et al. Phenotypic and molecular analysis of six human cell lines derived from patients with plasma cell dyscrasia. Br J Haematol. 1999;106:669–681. doi: 10.1046/j.1365-2141.1999.01602.x. [DOI] [PubMed] [Google Scholar]
- 27.Terstappen LW, Johnsen S, Segers-Nolten IM, Loken MR. Identification and characterization of plasma cells in normal human bone marrow by high-resolution flow cytometry. Blood. 1990;76:1739–1747. [PubMed] [Google Scholar]
- 28.Witzig TE, Kimlinger TK, Ahmann GJ, Katzmann JA, Greipp PR. Detection of myeloma cells in the peripheral blood by flow cytometry. Cytometry. 1996;26:113–120. doi: 10.1002/(SICI)1097-0320(19960615)26:2<113::AID-CYTO3>3.0.CO;2-H. [DOI] [PubMed] [Google Scholar]
- 29.Johnson P, Glennie M. The mechanisms of action of rituximab in the elimination of tumor cells. Semin Oncol. 2003;30:3–8. doi: 10.1053/sonc.2003.50025. [DOI] [PubMed] [Google Scholar]
- 30.Millar BC, Bell JB, Lakhani A, et al. A simple method for culturing myeloma cells from human bone marrow aspirates and peripheral blood in vitro. Br J Haematol. 1988;69:197–203. doi: 10.1111/j.1365-2141.1988.tb07622.x. [DOI] [PubMed] [Google Scholar]
- 31.Barker HF, Ball J, Drew M, Franklin IM. Multiple myeloma: the biology of malignant plasma cells. Blood Rev. 1993;7:19–23. doi: 10.1016/0268-960x(93)90020-5. [DOI] [PubMed] [Google Scholar]
- 32.Rebouissou C, Wijdenes J, Autissier P, et al. A gp130 interleukin-6 transducer-dependent SCID model of human multiple myeloma. Blood. 1998;91:4727–4737. [PubMed] [Google Scholar]
- 33.Yaccoby S, Epstein J. The proliferative potential of myeloma plasma cells manifest in the SCID-hu host. Blood. 1999;94:3576–3582. [PubMed] [Google Scholar]
- 34.Pilarski LM, Hipperson G, Seeberger K, et al. Myeloma progenitors in the blood of patients with aggressive or minimal disease: engraftment and self-renewal of primary human myeloma in the bone marrow of NOD SCID mice. Blood. 2000;95:1056–1065. [PubMed] [Google Scholar]
- 35.Reme T, Gueydon E, Jacquet C, Klein B, Brochier J. Growth and immortalization of human myeloma cells in immunodeficient severe combined immunodeficiency mice: a preclinical model. Br J Haematol. 2001;114:406–413. doi: 10.1046/j.1365-2141.2001.02947.x. [DOI] [PubMed] [Google Scholar]
- 36.Reya T, Morrison SJ, Clarke MF, Weissman IL. Stem cells, cancer, and cancer stem cells. Nature. 2001;414:105–111. doi: 10.1038/35102167. [DOI] [PubMed] [Google Scholar]
- 37.Jones RJ. Leukemic stem cells—where have they gone wrong? Blood. 2001;97:3681–3682. [Google Scholar]
- 38.Dick JE. Breast cancer stem cells revealed. Proc Natl Acad Sci U S A. 2003;100:3547–3549. doi: 10.1073/pnas.0830967100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Bedi A, Zehnbauer BA, Collector MI, et al. BCR-ABL gene rearrangement and expression of primitive hematopoietic progenitors in chronic myeloid leukemia. Blood. 1993;81:2898–2902. [PubMed] [Google Scholar]
- 40.Lapidot T, Sirard C, Vormoor J, et al. A cell initiating human acute myeloid leukaemia after transplantation into SCID mice. Nature. 1994;367:645–648. doi: 10.1038/367645a0. [DOI] [PubMed] [Google Scholar]
- 41.George AA, Franklin J, Kerkof K, et al. Detection of leukemic cells in the CD34+ CD38(−) bone marrow progenitor population in children with acute lymphoblastic leukemia. Blood. 2001;97:3925–3930. doi: 10.1182/blood.v97.12.3925. [DOI] [PubMed] [Google Scholar]
- 42.Nilsson L, Astrand-Grundstrom I, Arvidsson I, et al. Isolation and characterization of hematopoietic progenitor/stem cells in 5q-deleted myelodysplastic syndromes: evidence for involvement at the hematopoietic stem cell level. Blood. 2000;96:2012–2021. [PubMed] [Google Scholar]
- 43.Al Hajj M, Wicha MS, Benito-Hernandez A, Morrison SJ, Clarke MF. Prospective identification of tumorigenic breast cancer cells. Proc Natl Acad Sci U S A. 2003;100:3983–3988. doi: 10.1073/pnas.0530291100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Tarte K, De Vos J, Thykjaer T, et al. Generation of polyclonal plasmablasts from peripheral blood B cells: a normal counterpart of malignant plasma-blasts. Blood. 2002;100:1113–1122. [PubMed] [Google Scholar]
- 45.Sahota SS, Leo R, Hamblin TJ, Stevenson FK. Myeloma VL and VH gene sequences reveal a complementary imprint of antigen selection in tumor cells. Blood. 1997;89:219–226. [PubMed] [Google Scholar]
- 46.Kuppers R, Klein U, Hansmann ML, Rajewsky K. Cellular origin of human B-cell lymphomas. N Engl J Med. 1999;341:1520–1529. doi: 10.1056/NEJM199911113412007. [DOI] [PubMed] [Google Scholar]
- 47.Treon SP, Pilarski LM, Belch AR, et al. CD20-directed serotherapy in patients with multiple myeloma: biologic considerations and therapeutic applications. J Immunother. 2002;25:72–81. doi: 10.1097/00002371-200201000-00008. [DOI] [PubMed] [Google Scholar]
- 48.Slifka MK, Antia R, Whitmire JK, Ahmed R. Humoral immunity due to long-lived plasma cells. Immunity. 1998;8:363–72. doi: 10.1016/s1074-7613(00)80541-5. [DOI] [PubMed] [Google Scholar]