

Abstract
(E)-β-caryophyllene (BCP) is a natural sequiterpene found in many essential oils of spice (best known for contributing to the spiciness of black pepper) and food plants with recognized anti-inflammatory properties. Recently it was shown that BCP is a natural agonist of endogenous cannabinoid 2 (CB2) receptors, which are expressed in immune cells and mediate anti-inflammatory effects. In this study we aimed to test the effects of BCP in a clinically relevant murine model of nephropathy (induced by the widely used antineoplastic drug cisplatin) in which the tubular injury is largely dependent on inflammation and oxidative/nitrative stress. β-caryophyllene dose-dependently ameliorated cisplatin-induced kidney dysfunction, morphological damage, and renal inflammatory response (chemokines MCP-1 and MIP-2, cytokines TNF-α and IL-1β, adhesion molecule ICAM-1, and neutrophil and macrophage infiltration). It also markedly mitigated oxidative/nitrative stress (NOX-2, NOX-4 expression, 4-HNE and 3-NT content) and cell death. The protective effects of BCP against biochemical and histological markers of nephropathy were absent in CB2 knockout mice. Thus, BCP may be an excellent therapeutic agent to prevent cisplatin-induced nephrotoxicity through a CB2 receptor dependent pathway. Given the excellent safety profile of BCP in humans it has tremendous therapeutic potential in multitude of diseases associated with inflammation and oxidative stress.
Keywords: nephropathy, cisplatin, (E)-β-caryophyllene, cannabinoid 2 receptor, inflammation, oxidative stress, cell death
Introduction
(E)-β-caryophyllene (BCP) is a natural sequiterpene found in many essential oils of spice (e.g. cinnamon, origanum, black pepper, basil and cloves) and food/medicinal plants[1, 2]. BCP is also often used in cosmetics and is an FDA-approved food additive and component of Cannabis sativa and Sativex, an approved drug in European countries and in Canada[3]. It has well-established anti-inflammatory[1] and antioxidant effects[4, 5] besides its antispasmodic[6], antiviral[7] and local anesthetic effect[8]. BCP effectively inhibited carrageen-induced paw edema and attenuated LPS-stimulated TNF-α and IL-1β expression in peripheral blood[1]. It has been shown to ameliorate dextrane sulphate sodium-induced colitis[9, 10] and inhibit gastric mucosal injuries induced by necrotizing agents such as absolute ethanol and 0.6 N HCl[11]. The wide availability, easy accessibility of BCP and its wide therapeutic window makes it an excellent candidate for therapeutic intervention.
Cisplatin is a small inorganic platinum agent with strong antineoplastic activity used in many cancer types, such as bladder, cervical, ovarian, lung and testicular cancer. Cisplatin forms highly reactive, charged, platinum complexes which bind to nucleophilic groups such as GC-rich sites in DNA, inducing intrastrand and interstrand DNA cross-links, as well as DNA-protein cross-links[12]. These cross-links arrest DNA synthesis and replication, and in rapidly diving cells (such as cancer cells) lead to DNA damage and consequently result in cell death. As most non-targeted chemotherapeutic agents, cisplatin also has several side effects. The dose-limiting side effect is its nephrotoxicity, and no effective therapy has been developed yet to prevent this complication of the therapy[13]. It is well established that oxidative/nitrative stress and inflammation play a key role in the pathomechanism of cisplatin-induced nephrotoxicity[14–21] which processes could serve as a good target for BCP.
In this study we aimed to investigate the possible therapeutic effect of BCP in a cisplatin-induced nephropathy model. Since BCP has been recently demonstrated selectively and potently activates the cannabinoid 2 (CB2) receptor[1], which are present in cells of the immune system and predominantly mediate anti-inflammatory effects[22–24], we also examined the possible involvement of CB2 receptors in the actions of BCP using CB2 knockout mice. This study may have very important clinical significance as (E)-β-caryophyllene (BCP) has been shown to exert antineoplastic effects[4] and potentiate the anticancer activity of the chemotherapeutic drug of paclitaxel[25] by mechanisms that remain unknown.
Material and methods
Animals and drug treatment
All animal experiments conformed to National Institutes of Health (NIH) guidelines and were approved by the Institutional Animal Care and Use Committee of the National Institute on Alcohol Abuse and Alcoholism (NIAAA; Bethesda, MD, USA). Six to 8-week-old male C57Bl/6J mice were obtained from The Jackson Laboratory (Bar Harbor, ME, USA). CB2 knockout mice (CB2−/−) were as described previously backcrossed to a C57Bl/6J background[26, 27]. All animals were kept in a temperature-controlled environment with a 12-h light–dark cycle and were allowed free access to food and water at all times, and were cared for in accordance with National Institutes of Health (NIH) guidelines. Mice were sacrificed 72 hours following a single injection of cisplatin (cis-diammineplatinum(II)-dichloride 25 mg/kg i.p.; Sigma) as described [16, 18, 28]. (E)-β-caryophyllene was purified by flash column chromatography on silica gel 60/0.015–0.04 mm (Macherey-Nagel), using n-hexan : ethylacetate (50:1) as decribed previously[1] from a ~80% pure natural commercial sample from Sigma-Aldrich (W225207) containing 20% other essential oil components from Syzygium aromaticum (cloves). The final purified product used in this study was >99% pure as determined by GC/MS. β-caryophyllene was dissolved in olive oil and administered at 1, 3 and 10 mg/kg, i.p. daily, starting 2 hours before the cisplatin administration.
Renal function monitoring
On the day of the sacrifice, blood was immediately collected and serum levels of creatinine and Blood Urea Nitrogen (BUN) were measured using VetTest 8000 blood chemistry analyzer (Idexx Lab)[29].
Histological examination
Following fixation of the kidneys with 10% formalin, renal tissues were sectioned and stained with periodic acid-Schiff (PAS) reagents for histological examination. Tubular damage in PAS-stained sections was examined under the microscope and scored based on the percentage of cortical tubules showing epithelial necrosis: 0 = normal; 1 = 1–25%; 2 = 26– 50%; 3 = 51–75%; 4 = >76%. Tubular necrosis was defined as the loss of the proximal tubular brush border, blebbing of apical membranes, tubular epithelial cell detachment from the basement membrane or intraluminal aggregation of cells and proteins as described[28]. For myeloperoxidase (MPO) or 4-hydroxy-2-nonenal (4-HNE) staining slides were deparaffinized, and hydrated in descending gradations of ethanol, followed by antigen retrieval procedure. Next, sections were incubated in 0.3% H2O2 in PBS to block endogenous peroxidase activity. The sections were then incubated with anti-MPO (Biocare Medical, Concord, CA) or anti-4-HNE (GENOX, Baltimore, MD) antibodies overnight at 4°C in a moist chamber. Biotinylated secondary antibodies and ABC reagent were added as per the kit’s instructions (Vector Laboratories, Burlingame, CA, USA). Color development was induced by incubation with DAB or by Vector Blue (Vector Laboratories) for 3–5 min, and in the case of MPO staining the sections were counter-stained with nuclear fast red as described [21]. Finally, the sections were dehydrated in ethanol and cleared in xylene and mounted. The specific staining was visualized and images were acquired using microscope IX-81 with 20X, 40X and 100x objectives (Olympus, Center Valley, PA). The morphometric examination was performed in a blinded manner by two independent investigators.
Renal myeloperoxidase activity assay
Myeloperoxidase [MPO, (EC1.11.1.7)] was measured by InnoZyme™ Myeloperoxidase Activity Kit (EMD Gibbstown, NJ) according to manufacturer’s instruction. Myeloperoxidase activities were expressed as fold change compared to the vehicle-treated control sample[28].
Renal 4-hydroxynonenal (4-HNE) content
4-HNE in the kidney tissues was determined using the kit (Cell Biolabs, San Diego)[18]. In brief, BSA or renal tissue extracts (10 μg/mL) are adsorbed on to a 96-well plate for 12hrs at 4°C. 4-HNE adducts present in the sample or standard are probed with anti-HNE antibody, followed by an HRP conjugated secondary antibody. The HNE-protein adducts content in an unknown sample is determined by comparing with a standard curve.
Renal poly(ADP-ribose) polymerase (PARP) activity and 3-nitrotyrosine (NT) content
PARP activity was determined by assay kit according to the manufacturer’s instructions (Trevigen, Gaithersburg, MD, USA)[30, 31]. NT was measured using the NT ELISA kit from Hycult Biotechnology (Cell Sciences, Canton, MA, USA) from tissue homogenates as described[31, 32]. Levels are presented as fold change compared to vehicle-treated control sample.
Detection of apoptosis by renal DNA fragmentation, and caspase 3/7 activity assays
For caspase assay of tissue lysate, caspase-3/7 activity of the lysate was measured using Apo-One Homogenous caspase-3/7 assay kit (Promega, Madison, WI) as described[33, 34]. An aliquot of caspase reagent was added to each well, mixed on a plate shaker for 1 h at room temperature with light protection, and the fluorescence emission was measured in Victor (Perkin Elmer). The DNA fragmentation assay was based on measuring the amount of mono- and oligonucleosomes in the cytoplasmic fraction of tissue extracts using a commercially available kit (Roche Diagnostics) according to the manufacturer’s instructions as described[35] and[27, 36].
Real-time PCR analyses
Total RNA was isolated from kidney homogenate using Trizol reagents (Invitrogen, Carlsbad, CA) according to manufacturer’s instruction. The isolated RNA was treated with RNase-free DNase (Ambion, Austin, TX) to remove traces of genomic DNA contamination. One microgram of total RNA of was reverse-transcribed to cDNA using the Super-Script II (Invitrogen, Carlsbad, CA). The target gene expression was quantified with Power Syber Green PCR Master Mix using ABI HT7900 Realtime PCR Instrument. Each amplified sample in all wells was analyzed for homogeneity using dissociation curve analysis. After denaturation at 95 °C for 2 min, 40 cycles were performed at 95 °C for 10 s, 60 °C for 30 s. Relative quantification was calculated using the comparative CT method (2−ΔΔCt method : ΔΔCt = Ct sample – Ct). Lower CT values and lower CT reflect a relatively higher amount of gene transcript. Statistical analyses were carried out for at least 6 to 8 replicate experimental samples in each set.
Primers used:
TNF-α: 5′AAGCCTGTAGCCCACGTCGTA3′ and 5′AGGTACAACCCATCGGCTGG3′
IL-1β: 5′AAAAAAGCCTCGTGCTGTCG3′ and 5′GTCGTTGCTTGGTTCTCCTTG3′
MCP-1: 5′TCAGCCAGATGCAGTTAACGC-3′ and 5′TCTGGACCCATTCCTTCTTGG-3′
MIP-2: 5′GGATGGCTTTCATGGAAGGAG-3′ and 5′TTGCTAAGCAAGGCACTGTGC-3′
ICAM: 5′AACTTTTCAGCTCCGGTCCTG 3′ and 5′ TCAGTGTGAATTGGACCTGCG3′
NOX2 (gp91phox): 5′GACCATTGCAAGTGAACACCC3′ and 5′AAATGAAGTGGACTCCACGCG3′
NOX4 (RENOX): 5′TCATTTGGCTGTCCCTAAACG3′ and 5′AAGGATGAGGCTGCAGTTGAG3′
F4/80: 5′TGTGTCGTGCTGTTCAGAACC3′ and 5′AGGAATCCCGCAATGATGG3′
Actin: 5′TGCACCACCAACTGCTTAG3′ and 5′GGATGCAGGGATGATGTTC3′.
Statistical analysis
Results are expressed as mean±SEM. Statistical significance among groups was determined by one-way ANOVA followed by Newman-Keuls post hoc analysis using GraphPad Prism 5 software (San Diego, CA). Probability values of P<0.05 were considered significant.
Results
β-caryophyllene (BCP) attenuates cisplatin-induced kidney injury in a CB2-receptor dependent manner
BCP pretreatment dose-dependently ameliorated kidney dysfunction as evidenced by the attenuation of the increased creatinine and blood urea nitrogen (BUN) values 72 hours after cisplatin administration reaching significance at 10 mg/kg (Figure 1). Furthermore, 10 mg/kg BCP also decreased tubular necrosis as determined by PAS staining of the kidney (Figure 2). CB2 knockout mice had increased susceptibility to cisplatin nephrotoxicity and BCP was not able to influence this increased kidney dysfunction (Figure 1) and the histological damage (Figure 2), suggesting that the protective action of BCP was mediated through CB2. BCP pretreatment alone had no effect in control mice on all variables studied.
Figure 1. β-caryophyllene pretreatment attenuates cisplatin-induced kidney dysfunction in CB2 receptor dependent manner.

Serum creatinine (A) ALT and (B) BUN levels in Control (vehicle-treated) mice pretreated with vehicle or β-caryophyllene (BCP) (n = 5–8/group) or in wild type and CB2 knockout mice treated with 25 mg/kg cisplatin pretreated with vehicle or BCP (1, 3 or 10 mg/kg, n = 5–12/group). Results are mean±S.E.M. *P<0.05 vs. Control with vehicle pretreatment, #P<0.05 vs. Cisplatin with vehicle pretreatment.
Figure 2. β-caryophyllene decreases cisplatin-induced histological injury.

(A) PAS staining of representative kidney sections of control mice treated with vehicle or BCP and wild type or CB2 knockout mice exposed to 25 mg/kg cisplatin treated with vehicle or BCP. Similar histological profiles were seen in three to five kidneys/group (400× magnification). (B) Quantification of the histological damage in kidney sections. Results are mean±S.E.M. of 20 representative field/group *P<0.05 vs. Control with vehicle pretreatment, #P<0.05 vs. Cisplatin with vehicle pretreatment.
β-caryophyllene (BCP) attenuates the inflammatory response in the kidney induced by cisplatin treatment
Cisplatin has greatly increased the mRNA expression of pro-inflammatory chemokines monocyte chemotactic protein-1 (MCP-1/CCL2), macrophage inflammatory protein-2 (MIP-2/CXCL2) and intercellular adhesion molecule 1 (ICAM-1/CD54) (Figure 3A–D) in the kidney as documented by real-time PCR. These responses were significantly attenuated by the pretreatment of 10 mg/kg BCP (Fig. 3A–C). Cisplatin enhanced leukocyte infiltration in the kidney demonstrated by myeloperoxidase (MPO) staining (Figure 4AB, brown staining in infiltrating immune cells) 72 hours after the injection of cisplatin. mRNA expressions of F4/80 (marker of macrophage infiltration; Figure 4C), as well as of pro-inflammatory cytokines TNF-α and IL-1β (Figure 5) were also significantly increased after cisplatin administration. BCP markedly reduced the inflammatory cell infiltration and pro-inflammatory response (Figures 3–5).
Figure 3. β-caryophyllene attenuates cisplatin-induced acute pro-inflammatory chemokine response and adhesion molecule expression in the kidney.
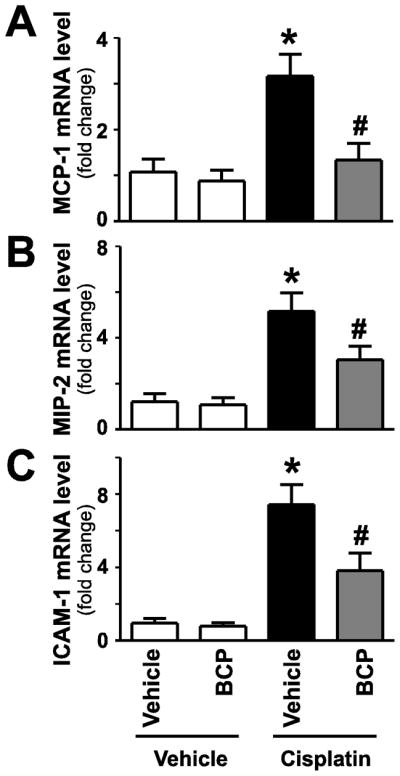
Cisplatin significantly increased renal mRNA expression of proinflammatory chemokines MCP-1 and MIP-2 (panels AB), and intercellular adhesion molecule 1 (ICAM-1, panel C) indicating enhanced inflammatory response 72 hours following its administration to mice. These were attenuated by treatment with BCP. Results are mean±S.E.M. of 8–16/group *P<0.05 vs. Control with vehicle pretreatment, #P<0.05 vs. Cisplatin with vehicle pretreatment.
Figure 4. β-caryophyllene attenuates cisplatin-induced leukocyte and macrophage infiltration.

Cisplatin significantly increased renal myeloperoxidase (MPO) activity/staining (panel A, B; an indicator of leukocyte infiltration), and mRNA expression of F4/80 (a marker of macrophages, panel C) indicating enhanced inflammatory response and cell infiltration. These were attenuated by BCP treatment (panels A–C; n=5–16/group). Panel A shows representative images of MPO staining (brown, 1000x magnification) from mice treated with cisplatin or cisplatin in combination with BCP, and Panel B shows quantifications from 8 representative fields with 400x magnification/group. Results are mean±S.E.M. *P<0.05 vs. Control with vehicle pretreatment, #P<0.05 vs. Cisplatin with vehicle pretreatment.
Figure 5. β-caryophyllene attenuates cisplatin-induced acute pro-inflammatory cytokine response in the kidney.
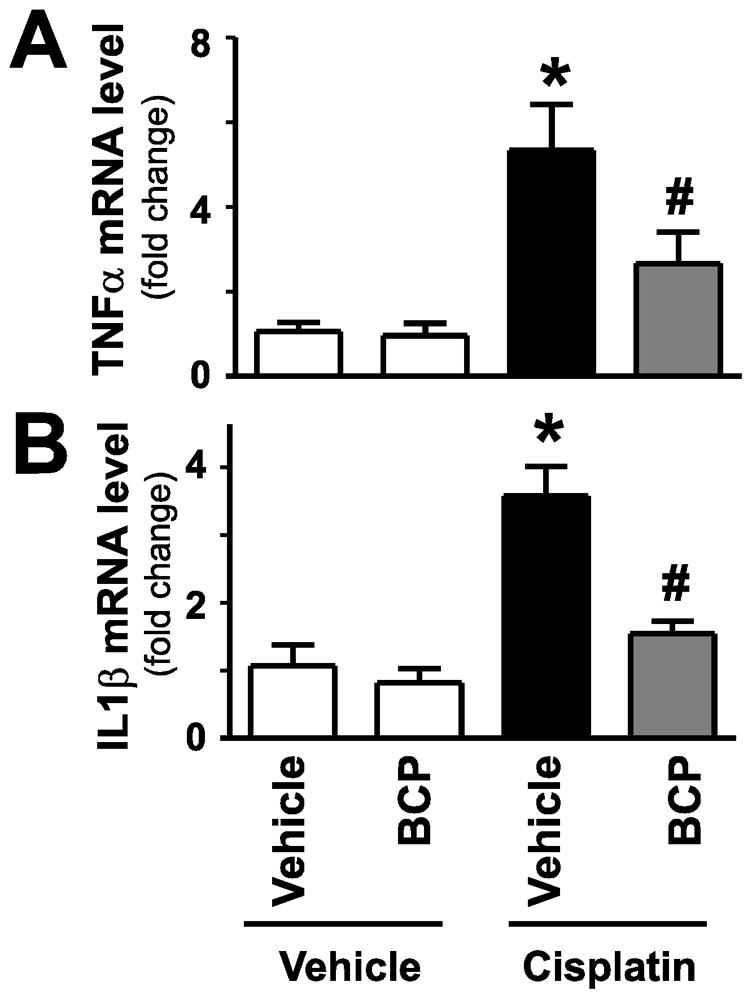
Cisplatin significantly increased renal mRNA expression of proinflammatory cytokines TNF-α and IL1β mRNA (panels A–B), indicating enhanced inflammatory response 72 hours following its administration to mice. These were attenuated by treatment with BCP. Results are mean±S.E.M. of 8–16/group. *P<0.05 vs. Control with vehicle pretreatment, #P<0.05 vs. Cisplatin with vehicle pretreatment.
β-caryophyllene (BCP) attenuates cisplatin-induced increased oxidative and nitrative stress
Reactive oxygen species (ROS) play a key role in the pathomechanism of cisplatin-induced kidney injury. Consistently with previous studies[16, 18, 28] cisplatin significantly increased renal mRNA expression of ROS generating NADPH oxidase enzyme isoforms NOX2 (gp91phox) and NOX4 (renox) (Figure 6AB) 3 days after cisplatin administration. Lipid peroxides are unstable indicators of oxidative stress in cells that decompose to form more complex and reactive compounds such as 4-hydroxynonenal (4-HNE), which has been shown to be capable of binding to proteins and forming stable adducts with possible signaling functions[37, 38]. Nitrotyrosine (3-NT) formation was initially considered a specific marker of in vivo peroxynitrite generation, but now it is rather used as a collective index of protein nitration, because other pathways have also been proposed to be involved in its formation (e.g., myeloperoxidase under certain inflammatory conditions[39–42]. Activated inflammatory cells are known to produce both superoxide via activation of NADPH oxidases and nitric oxide (NO) via activation of iNOS favoring peroxynitrite generation through a diffusion-limited reaction of NO and superoxide[43, 44]. Both reactive oxygen and nitrogen species may lead to increased lipid peroxidation[45]. In kidneys of cisplatin-treated mice we observed markedly increased immunohistochemical staining of 4-HNE (Figure 7A), which was also confirmed with quantitative assay from tissues (Figure 7B). Nitrotyrosine content was also markedly increased in the kidney after cisplatin treatment (Figure 7C). The cisplatin-induced enhanced oxidative and nitrative stress was significantly attenuated by BCP pretreatment (Figures 6–7).
Figure 6. β-caryophyllene attenuates cisplatin-induced increased expression of ROS generating NADPH oxidase isoforms NOX2 (gp91phox) and NOX4 (RENOX).
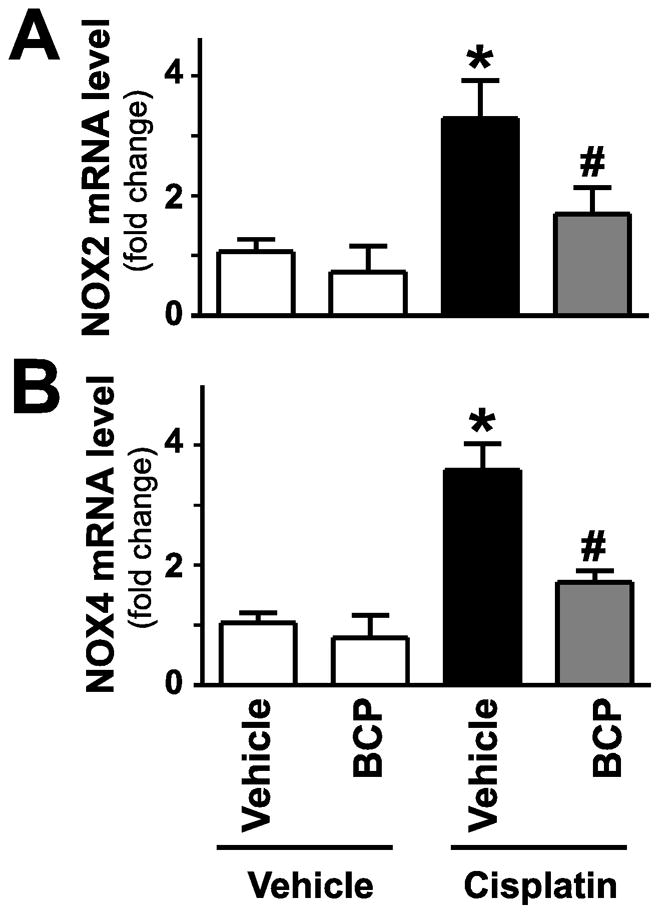
Cisplatin significantly increased renal mRNA expression of NOX2 and NOX4 (Panels A–B) 72 hours following its administration to mice. These were attenuated by treatment with BCP. Results are mean±S.E.M. of 8–16/group. *P<0.05 vs. Control with vehicle pretreatment, #P<0.05 vs. Cisplatin with vehicle pretreatment.
Figure 7. β-caryophyllene attenuates cisplatin-induced increased oxidative and nitrative stress.

Cisplatin significantly increased renal HNE protein adduct (panel A–B) and 3-NT levels in the kidneys (panels C), indicating enhanced oxidative/nitrative stress 72 hours following its administration to mice. These were attenuated by treatment with BCP. Panel A shows representative images of HNE staining (bluish grey, 400x magnification) from mice treated with cisplatin or cisplatin in combination with BCP. Results are mean±S.E.M of 6–10/group. *P<0.05 vs. Control with vehicle pretreatment, #P<0.05 vs. Cisplatin with vehicle pretreatment.
β-caryophyllene (BCP) attenuates cisplatin-induced enhanced cell death
Reactive oxygen and nitrogen species may activate mitochondrial and poly(ADP-ribose) polymerase (PARP)-dependent cell death pathways leading to both apoptotic or necrotic cell demise[39, 41, 46, 47], which were also implicated in the pathogenesis of cisplatin-induced nephropathy[16, 18, 21, 28]. Cisplatin increased renal tubular cell apoptosis as evidenced by the increased caspase 3/7 activity in the kidney tissue samples of mice (Figure 8A). It also caused a marked increase in DNA fragmentation, and PARP activation (Figure 8B,C). Intriguingly, these markers were all attenuated by 10 mg/kg BCP pretreatment.
Figure 8. β-caryophyllene attenuates cisplatin-induced enhanced cell death in the kidney.
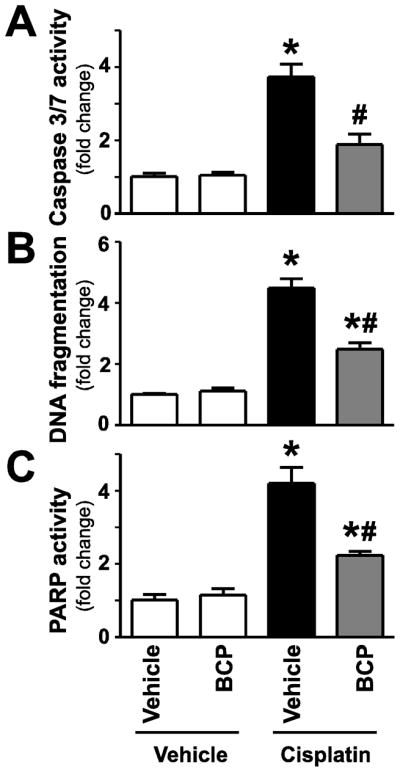
Cisplatin significantly increased DNA fragmentation, caspase 3/7 and PARP activity (Panels A–C) 72 hours following its administration to mice. These were attenuated by treatment with BCP. Results are mean±S.E.M. of 6–10/group *P<0.05 vs. Control with vehicle pretreatment, #P<0.05 vs. Cisplatin with vehicle pretreatment.
Discussion
In this study we provide evidence that β-caryophyllene (BCP) is able to markedly attenuate the cisplatin-induced decline in kidney function and ameliorate the observed histological damage, and that this protective effect is mediated through CB2 receptors since it could be completely abolished in CB2 receptor knockout mice. The protective effect involves marked attenuation of cisplatin-induced pro-inflammatory response and oxidative and nitrative stress.
The CB2 receptor is primarily considered to be expressed in immune or various immune-derived cells, such as various populations of T and B lymphocytes, leukocytes, monocytes/macrophages, dendritic cells, mast cells, microglia in the brain, and resident macrophages of the liver (Kupffer cells)[24, 48]. In these cell types synthetic CB2 agonists and/or endocannabinoids mostly mediate various immunosuppressive effects, which may limit inflammation and associated tissue injury in large number of pathological conditions[24]. Recent studies have also demonstrated existence of low level of CB2 receptors in gastrointestinal, cardiovascular, bone, various neuronal, liver tissues and/or cell types[24]. Despite the presence, CB2 receptors appear to play limited (if any) role in normal physiological regulation of these organ systems, consistently with the absence of apparent pathological alterations in normal CB2 receptor knockout mice[24]. However, under various pathological conditions, such as ischemic reperfusion injury of liver[26, 35, 36, 49], heart[50], and brain[51–53], other forms of liver damage[54, 55], colitis[56–58], and nephropathy[28, 59], among many others[24]), CB2 receptors may play a significant role in limiting the inflammatory response, oxidative/nitrative stress and consequent tissue injury, and ultimately organ dysfunction[24]. Under these conditions marked dysregulation of the cannabinoid receptors, endocannabinoids and/or their metabolizing enzymes have also been reported[22, 24]. In many of these tissue injury models CB2 knockout mice respond to insult with aggravated damage compared to their controls[24], supporting a potential protective role of the endocannabinoid system through CB2 receptors. The latter is also in agreement with our observation in the current nephropathy model.
Although the exact role of the endocannabinoid system and CB1/2 receptor in cancer is still a controversial and largely unexplored issue[22, 24], there are encouraging reports in support of the potential use of various cannabinoid ligands not only as palliative therapy, but also because of their ability to inhibit the growth and metastasis formation of certain types of tumors such as gliomas, tumors of immune origin, and melanomas[24, 60–63]. The recent recognition of the importance of microenvironment and local inflammation in cancer progression, growth and metastasis formation, may also provide additional rational for exploring the possibility of targeting CB2 receptors in certain types of tumors[24].
We have recently shown that activation of the CB2 receptor can attenuate the inflammation, oxidative/nitrative stress, and cell death in cisplatin-induced nephropathy and proposed this receptor as a potential target to prevent this devastating complication of chemotherapy[28]. Consistently with this observation, a recent study has provided additional support on the protective role of this receptor in a model of diabetic nephropathy[59]. While traditional cannabinoid ligands such as delta-9-tetrahydrocannabinol (THC; an agonist of both cannabinoid 1 and 2 receptors) with central CB1 receptor activity (CB1 receptors in CNS mediate the known psychoactive effects of cannabis) are not acceptable to pharmacological development despite their known anti-inflammatory properties most likely via CB2 receptors expressed in immune cells[22, 24], BCP seems to be an excellent candidate as it is readily available as a natural compound found in many food plants and it is without any known side effect[64].
Cisplatin has been shown to cause renal injury via multiple intercalating pathways with central role of inflammation and oxidative and nitrative stress[13–18, 20, 21], and activation of the CB2 receptor can interfere with this complex pathomechanism at multiple levels[24]. The proinflammatory cytokine TNF-α, for example plays a key role in the pathomechanism of cisplatin-induced kidney injury[15]. HU-910, a highly specific CB2-receptor agonist, was able to attenuate TNF-α production in Kuppfer cells (the resident macrophages of the liver)[36]. As we have seen a significant increase in the marker of macrophage infiltration (F4/80) after cisplatin administration, decreased activation of these cells by BCP could be a possible mechanism of the compound’s beneficial effect. Furthermore, activation of CB2 receptors was also able to attenuate TNF-α-induced ICAM-1 and VCAM-1 expression on endothelial cells and TNF-α-induced NF-κB activation[65] which may also contribute to the anti-inflammatory property of BCP. Moreover, BCP attenuate the cisplatin-induced increased mRNA expression of proinflammatory chemokines (which attract inflammatory cells to the site of injury) MCP-1 and MIP-2. These results are in agreement with recent findings in a diabetic nephopathy model[59], in which activation of CB2 receptors could ameliorate albuminuria, podocyte protein downregulation, and glomerular monocyte infiltration by interfering with the deleterious effects of MCP-1 signaling[59]. Consistently BCP also markedly attenuated the cisplatin-induced inflammatory cell infiltration and associated oxidative and nitrative stress.
In addition to proximal tubular cells cisplatin is also directly toxic to endothelial cells[66], vascular injury and ischemia-reperfusion can play a very important role in its pathogenesis. CB2 receptor activation has been shown to attenuate very effectively hepatic[26, 36, 49], myocardial[67], and other forms of ischemia-reperfusion injury (reviewed in[24, 53]) via attenuation of endothelial cell activation, chemokine signaling, inflammatory cell infiltration and/or activation of these immune cells[24]. These protective effects of CB2 receptor activation may also be important during vascular injury[65, 68, 69]. Therefore it is quite reasonable to speculate that BCP can also have an indirect effect on kidney injury via attenuating the ensuing damage from renal ischemia.
From a therapeutic point of view it is important to understand if BCP itself is responsible for the protective effects observed in our kidney injury model or its putative CB2 receptor active metabolites may also play a role. To date the metabolism of BCP is still largely unexplored. The potential metabolites β-caryophyllene oxide and the corresponding diol do not interact with CB2 receptors (unpublished data) similarly to epoxide[1], acetate and glycol metabolites reported from rabbits[70, 71]. It is therefore likely that BCP is the major CB2 agonist and that the pharmacokinetics of this compound largely determines its pharmacological efficacy.
Collectively, we demonstrate that pretreatment with the natural product BCP can ameliorate the deleterious effect of cisplatin on kidney injury through a CB2 receptor dependent pathway. Our results combined with the known antineoplastic and chemotherapeutic potentiating effect of this agent make this compound an excellent candidate for clinical drug development. Given the excellent safety profile of BCP in humans it has tremendous therapeutic potential in multitude of diseases associated with inflammation and oxidative stress, which warrants further exploration.
Acknowledgments
This study was supported by the Intramural Research Program of NIH/NIAAA (to P.P.) and HL072889 (to A.H.B). Dr. Béla Horváth is the recipient of a Hungarian Scientific Research Fund Fellowship (OTKA-NKTH-EU MB08-80238). Authors are indebted to Dr. George Kunos for continuous support. Dr. Pacher dedicates this study to his beloved mother Iren Bolfert, who died from complications of chemotherapy.
Footnotes
Disclosures: No conflict of interest to disclose.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Gertsch J, Leonti M, Raduner S, Racz I, Chen JZ, Xie XQ, Altmann KH, Karsak M, Zimmer A. Beta-caryophyllene is a dietary cannabinoid. Proc Natl Acad Sci U S A. 2008;105:9099–9104. doi: 10.1073/pnas.0803601105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Gertsch J. Anti-inflammatory cannabinoids in diet: Towards a better understanding of CB(2) receptor action? Commun Integr Biol. 2008;1:26–28. doi: 10.4161/cib.1.1.6568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Sibbald B. Conditional okay for cannabis prescription drug. CMAJ. 2005;172:1672. doi: 10.1503/cmaj.050628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kubo I, Chaudhuri SK, Kubo Y, Sanchez Y, Ogura T, Saito T, Ishikawa H, Haraguchi H. Cytotoxic and antioxidative sesquiterpenoids from Heterotheca inuloides. Planta Med. 1996;62:427–430. doi: 10.1055/s-2006-957932. [DOI] [PubMed] [Google Scholar]
- 5.Singh G, Marimuthu P, de Heluani CS, Catalan CA. Antioxidant and biocidal activities of Carum nigrum (seed) essential oil, oleoresin, and their selected components. J Agric Food Chem. 2006;54:174–181. doi: 10.1021/jf0518610. [DOI] [PubMed] [Google Scholar]
- 6.Leonhardt V, Leal-Cardoso JH, Lahlou S, Albuquerque AA, Porto RS, Celedonio NR, Oliveira AC, Pereira RF, Silva LP, Garcia-Teofilo TM, Silva AP, Magalhaes PJ, Duarte GP, Coelho-de-Souza AN. Antispasmodic effects of essential oil of Pterodon polygalaeflorus and its main constituent beta-caryophyllene on rat isolated ileum. Fundam Clin Pharmacol. 2010;24:749–758. doi: 10.1111/j.1472-8206.2009.00800.x. [DOI] [PubMed] [Google Scholar]
- 7.Astani A, Reichling J, Schnitzler P. Screening for Antiviral Activities of Isolated Compounds from Essential Oils. Evid Based Complement Alternat Med. 2011 doi: 10.1093/ecam/nep187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ghelardini C, Galeotti N, Di Cesare Mannelli L, Mazzanti G, Bartolini A. Local anaesthetic activity of beta-caryophyllene. Farmaco. 2001;56:387–389. doi: 10.1016/s0014-827x(01)01092-8. [DOI] [PubMed] [Google Scholar]
- 9.Cho JY, Chang HJ, Lee SK, Kim HJ, Hwang JK, Chun HS. Amelioration of dextran sulfate sodium-induced colitis in mice by oral administration of beta-caryophyllene, a sesquiterpene. Life Sci. 2007;80:932–939. doi: 10.1016/j.lfs.2006.11.038. [DOI] [PubMed] [Google Scholar]
- 10.Bento AF, Marcon R, Dutra RC, Claudino RF, Cola M, Leite DF, Calixto JB. beta-Caryophyllene inhibits dextran sulfate sodium-induced colitis in mice through CB2 receptor activation and PPARgamma pathway. Am J Pathol. 2011;178:1153–1166. doi: 10.1016/j.ajpath.2010.11.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Tambe Y, Tsujiuchi H, Honda G, Ikeshiro Y, Tanaka S. Gastric cytoprotection of the non-steroidal anti-inflammatory sesquiterpene, beta-caryophyllene. Planta Med. 1996;62:469–470. doi: 10.1055/s-2006-957942. [DOI] [PubMed] [Google Scholar]
- 12.Zwelling LA, Anderson T, Kohn KW. DNA-protein and DNA interstrand cross-linking by cis- and trans-platinum(II) diamminedichloride in L1210 mouse leukemia cells and relation to cytotoxicity. Cancer Res. 1979;39:365–369. [PubMed] [Google Scholar]
- 13.Pabla N, Dong Z. Cisplatin nephrotoxicity: mechanisms and renoprotective strategies. Kidney Int. 2008;73:994–1007. doi: 10.1038/sj.ki.5002786. [DOI] [PubMed] [Google Scholar]
- 14.Davis CA, Nick HS, Agarwal A. Manganese superoxide dismutase attenuates Cisplatin-induced renal injury: importance of superoxide. J Am Soc Nephrol. 2001;12:2683–2690. doi: 10.1681/ASN.V12122683. [DOI] [PubMed] [Google Scholar]
- 15.Ramesh G, Reeves WB. TNF-alpha mediates chemokine and cytokine expression and renal injury in cisplatin nephrotoxicity. J Clin Invest. 2002;110:835–842. doi: 10.1172/JCI15606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Pan H, Mukhopadhyay P, Rajesh M, Patel V, Mukhopadhyay B, Gao B, Hasko G, Pacher P. Cannabidiol attenuates cisplatin-induced nephrotoxicity by decreasing oxidative/nitrosative stress, inflammation, and cell death. J Pharmacol Exp Ther. 2009;328:708–714. doi: 10.1124/jpet.108.147181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Mukhopadhyay P, Pan H, Rajesh M, Batkai S, Patel V, Harvey-White J, Mukhopadhyay B, Hasko G, Gao B, Mackie K, Pacher P. CB1 cannabinoid receptors promote oxidative/nitrosative stress, inflammation and cell death in a murine nephropathy model. Br J Pharmacol. 2010;160:657–668. doi: 10.1111/j.1476-5381.2010.00769.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Mukhopadhyay P, Horvath B, Kechrid M, Tanchian G, Rajesh M, Naura AS, Boulares AH, Pacher P. Poly(ADP-ribose) polymerase-1 is a key mediator of cisplatin-induced kidney inflammation and injury. Free Radic Biol Med. 2011;51:1774–1788. doi: 10.1016/j.freeradbiomed.2011.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Mukhopadhyay P, Horvath B, Zsengeller Z, Zielonka J, Tanchian G, Holovac E, Kechrid M, Patel V, Stillman IE, Parikh SM, Joseph J, Kalyanaraman B, Pacher P. Mitochondrial-targeted antioxidants represent a promising approach for prevention of cisplatin-induced nephropathy. Free Radic Biol Med. 2011 doi: 10.1016/j.freeradbiomed.2011.11.001. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Chirino YI, Trujillo J, Sanchez-Gonzalez DJ, Martinez-Martinez CM, Cruz C, Bobadilla NA, Pedraza-Chaverri J. Selective iNOS inhibition reduces renal damage induced by cisplatin. Toxicol Lett. 2008;176:48–57. doi: 10.1016/j.toxlet.2007.10.006. [DOI] [PubMed] [Google Scholar]
- 21.Guerrero-Beltran CE, Mukhopadhyay P, Horvath B, Rajesh M, Tapia E, Garcia-Torres I, Pedraza-Chaverri J, Pacher P. Sulforaphane, a natural constituent of broccoli, prevents cell death and inflammation in nephropathy. J Nutr Biochem. 2011 doi: 10.1016/j.jnutbio.2011.02.004. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Pacher P, Batkai S, Kunos G. The endocannabinoid system as an emerging target of pharmacotherapy. Pharmacol Rev. 2006;58:389–462. doi: 10.1124/pr.58.3.2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Di Marzo V, Bifulco M, De Petrocellis L. The endocannabinoid system and its therapeutic exploitation. Nat Rev Drug Discov. 2004;3:771–784. doi: 10.1038/nrd1495. [DOI] [PubMed] [Google Scholar]
- 24.Pacher P, Mechoulam R. Is lipid signaling through cannabinoid 2 receptors part of a protective system? Prog Lipid Res. 2011;50:193–211. doi: 10.1016/j.plipres.2011.01.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Legault J, Pichette A. Potentiating effect of beta-caryophyllene on anticancer activity of alpha-humulene, isocaryophyllene and paclitaxel. J Pharm Pharmacol. 2007;59:1643–1647. doi: 10.1211/jpp.59.12.0005. [DOI] [PubMed] [Google Scholar]
- 26.Batkai S, Osei-Hyiaman D, Pan H, El-Assal O, Rajesh M, Mukhopadhyay P, Hong F, Harvey-White J, Jafri A, Hasko G, Huffman JW, Gao B, Kunos G, Pacher P. Cannabinoid-2 receptor mediates protection against hepatic ischemia/reperfusion injury. FASEB J. 2007;21:1788–1800. doi: 10.1096/fj.06-7451com. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Mukhopadhyay P, Rajesh M, Horvath B, Batkai S, Park O, Tanchian G, Gao RY, Patel V, Wink DA, Liaudet L, Hasko G, Mechoulam R, Pacher P. Cannabidiol protects against hepatic ischemia/reperfusion injury by attenuating inflammatory signaling and response, oxidative/nitrative stress, and cell death. Free Radic Biol Med. 2011;50:1368–1381. doi: 10.1016/j.freeradbiomed.2011.02.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Mukhopadhyay P, Rajesh M, Pan H, Patel V, Mukhopadhyay B, Batkai S, Gao B, Hasko G, Pacher P. Cannabinoid-2 receptor limits inflammation, oxidative/nitrosative stress, and cell death in nephropathy. Free Radic Biol Med. 2010;48:457–467. doi: 10.1016/j.freeradbiomed.2009.11.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Chandra R, Federici S, Nemeth ZH, Horvath B, Pacher P, Hasko G, Deitch EA, Spolarics Z. Female X-chromosome mosaicism for NOX2 deficiency presents unique inflammatory phenotype and improves outcome in polymicrobial sepsis. J Immunol. 2011;186:6465–6473. doi: 10.4049/jimmunol.1100205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Rajesh M, Mukhopadhyay P, Batkai S, Patel V, Saito K, Matsumoto S, Kashiwaya Y, Horvath B, Mukhopadhyay B, Becker L, Hasko G, Liaudet L, Wink DA, Veves A, Mechoulam R, Pacher P. Cannabidiol attenuates cardiac dysfunction, oxidative stress, fibrosis, and inflammatory and cell death signaling pathways in diabetic cardiomyopathy. J Am Coll Cardiol. 2010;56:2115–2125. doi: 10.1016/j.jacc.2010.07.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Mukhopadhyay P, Horvath B, Rajesh M, Matsumoto S, Saito K, Batkai S, Patel V, Tanchian G, Gao RY, Cravatt BF, Hasko G, Pacher P. Fatty acid amide hydrolase is a key regulator of endocannabinoid-induced myocardial tissue injury. Free Radic Biol Med. 2011;50:179–195. doi: 10.1016/j.freeradbiomed.2010.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.El-Remessy AB, Rajesh M, Mukhopadhyay P, Horvath B, Patel V, Al-Gayyar MM, Pillai BA, Pacher P. Cannabinoid 1 receptor activation contributes to vascular inflammation and cell death in a mouse model of diabetic retinopathy and a human retinal cell line. Diabetologia. 2011;54:1567–1578. doi: 10.1007/s00125-011-2061-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Gao RY, Mukhopadhyay P, Mohanraj R, Wang H, Horvath B, Yin S, Pacher P. Resveratrol attenuates azidothymidine-induced cardiotoxicity by decreasing mitochondrial reactive oxygen species generation in human cardiomyocytes. Mol Med Report. 2011;4:151–155. doi: 10.3892/mmr.2010.390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Mukhopadhyay P, Rajesh M, Batkai S, Patel V, Kashiwaya Y, Liaudet L, Evgenov OV, Mackie K, Hasko G, Pacher P. CB1 cannabinoid receptors promote oxidative stress and cell death in murine models of doxorubicin-induced cardiomyopathy and in human cardiomyocytes. Cardiovasc Res. 2010;85:773–784. doi: 10.1093/cvr/cvp369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Batkai S, Mukhopadhyay P, Horvath B, Rajesh M, Gao RY, Mahadevan A, Amere M, Battista N, Lichtman AH, Gauson LA, Maccarrone M, Pertwee RG, Pacher P. Delta(8) -Tetrahydrocannabivarin protects against hepatic ischemia/reperfusion injury by attenuating oxidative stress and inflammatory response involving CB(2) receptors. Br J Pharmacol. 2011 doi: 10.1111/j.1476-5381.2011.01410.x. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Horvath B, Magid L, Mukhopadhyay P, Batkai S, Rajesh M, Park O, Tanchian G, Gao RY, Goodfellow CE, Glass M, Mechoulam R, Pacher P. A new cannabinoid 2 receptor agonist HU-910 attenuates oxidative stress, inflammation, and cell death associated with hepatic ischemia/reperfusion injury. Br J Pharmacol. 2011 doi: 10.1111/j.1476-5381.2011.01381.x. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Forman HJ, Fukuto JM, Miller T, Zhang H, Rinna A, Levy S. The chemistry of cell signaling by reactive oxygen and nitrogen species and 4-hydroxynonenal. Arch Biochem Biophys. 2008;477:183–195. doi: 10.1016/j.abb.2008.06.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Forman HJ, Maiorino M, Ursini F. Signaling functions of reactive oxygen species. Biochemistry. 2010;49:835–842. doi: 10.1021/bi9020378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Pacher P, Beckman JS, Liaudet L. Nitric oxide and peroxynitrite in health and disease. Physiol Rev. 2007;87:315–424. doi: 10.1152/physrev.00029.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Eiserich JP, Hristova M, Cross CE, Jones AD, Freeman BA, Halliwell B, van der Vliet A. Formation of nitric oxide-derived inflammatory oxidants by myeloperoxidase in neutrophils. Nature. 1998;391:393–397. doi: 10.1038/34923. [DOI] [PubMed] [Google Scholar]
- 41.Szabo C, Ischiropoulos H, Radi R. Peroxynitrite: biochemistry, pathophysiology and development of therapeutics. Nat Rev Drug Discov. 2007;6:662–680. doi: 10.1038/nrd2222. [DOI] [PubMed] [Google Scholar]
- 42.Beckman JS, Koppenol WH. Nitric oxide, superoxide, and peroxynitrite: the good, the bad, and ugly. Am J Physiol. 1996;271:C1424–1437. doi: 10.1152/ajpcell.1996.271.5.C1424. [DOI] [PubMed] [Google Scholar]
- 43.Forman HJ, Torres M. Reactive oxygen species and cell signaling: respiratory burst in macrophage signaling. Am J Respir Crit Care Med. 2002;166:S4–8. doi: 10.1164/rccm.2206007. [DOI] [PubMed] [Google Scholar]
- 44.Ischiropoulos H, Zhu L, Beckman JS. Peroxynitrite formation from macrophage-derived nitric oxide. Arch Biochem Biophys. 1992;298:446–451. doi: 10.1016/0003-9861(92)90433-w. [DOI] [PubMed] [Google Scholar]
- 45.Radi R, Beckman JS, Bush KM, Freeman BA. Peroxynitrite-induced membrane lipid peroxidation: the cytotoxic potential of superoxide and nitric oxide. Arch Biochem Biophys. 1991;288:481–487. doi: 10.1016/0003-9861(91)90224-7. [DOI] [PubMed] [Google Scholar]
- 46.Radi R, Cassina A, Hodara R, Quijano C, Castro L. Peroxynitrite reactions and formation in mitochondria. Free Radic Biol Med. 2002;33:1451–1464. doi: 10.1016/s0891-5849(02)01111-5. [DOI] [PubMed] [Google Scholar]
- 47.Szabo C. DNA strand breakage and activation of poly-ADP ribosyltransferase: a cytotoxic pathway triggered by peroxynitrite. Free Radic Biol Med. 1996;21:855–869. doi: 10.1016/0891-5849(96)00170-0. [DOI] [PubMed] [Google Scholar]
- 48.Howlett AC, Barth F, Bonner TI, Cabral G, Casellas P, Devane WA, Felder CC, Herkenham M, Mackie K, Martin BR, Mechoulam R, Pertwee RG. International Union of Pharmacology. XXVII. Classification of cannabinoid receptors. Pharmacol Rev. 2002;54:161–202. doi: 10.1124/pr.54.2.161. [DOI] [PubMed] [Google Scholar]
- 49.Rajesh M, Pan H, Mukhopadhyay P, Batkai S, Osei-Hyiaman D, Hasko G, Liaudet L, Gao B, Pacher P. Cannabinoid-2 receptor agonist HU-308 protects against hepatic ischemia/reperfusion injury by attenuating oxidative stress, inflammatory response, and apoptosis. J Leukoc Biol. 2007;82:1382–1389. doi: 10.1189/jlb.0307180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Defer N, Wan J, Souktani R, Escoubet B, Perier M, Caramelle P, Manin S, Deveaux V, Bourin MC, Zimmer A, Lotersztajn S, Pecker F, Pavoine C. The cannabinoid receptor type 2 promotes cardiac myocyte and fibroblast survival and protects against ischemia/reperfusion-induced cardiomyopathy. FASEB J. 2009;23:2120–2130. doi: 10.1096/fj.09-129478. [DOI] [PubMed] [Google Scholar]
- 51.Zhang M, Martin BR, Adler MW, Razdan RK, Jallo JI, Tuma RF. Cannabinoid CB(2) receptor activation decreases cerebral infarction in a mouse focal ischemia/reperfusion model. J Cereb Blood Flow Metab. 2007;27:1387–1396. doi: 10.1038/sj.jcbfm.9600447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Zhang M, Adler MW, Abood ME, Ganea D, Jallo J, Tuma RF. CB2 receptor activation attenuates microcirculatory dysfunction during cerebral ischemic/reperfusion injury. Microvasc Res. 2009;78:86–94. doi: 10.1016/j.mvr.2009.03.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Pacher P, Hasko G. Endocannabinoids and cannabinoid receptors in ischaemia-reperfusion injury and preconditioning. Br J Pharmacol. 2008;153:252–262. doi: 10.1038/sj.bjp.0707582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Louvet A, Teixeira-Clerc F, Chobert MN, Deveaux V, Pavoine C, Zimmer A, Pecker F, Mallat A, Lotersztajn S. Cannabinoid CB2 receptors protect against alcoholic liver disease by regulating Kupffer cell polarization in mice. Hepatology. 2011;54:1217–1226. doi: 10.1002/hep.24524. [DOI] [PubMed] [Google Scholar]
- 55.Teixeira-Clerc F, Belot MP, Manin S, Deveaux V, Cadoudal T, Chobert MN, Louvet A, Zimmer A, Tordjmann T, Mallat A, Lotersztajn S. Beneficial paracrine effects of cannabinoid receptor 2 on liver injury and regeneration. Hepatology. 2010;52:1046–1059. doi: 10.1002/hep.23779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Storr MA, Keenan CM, Zhang H, Patel KD, Makriyannis A, Sharkey KA. Activation of the cannabinoid 2 receptor (CB2) protects against experimental colitis. Inflamm Bowel Dis. 2009;15:1678–1685. doi: 10.1002/ibd.20960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Wright KL, Duncan M, Sharkey KA. Cannabinoid CB2 receptors in the gastrointestinal tract: a regulatory system in states of inflammation. Br J Pharmacol. 2008;153:263–270. doi: 10.1038/sj.bjp.0707486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Izzo AA, Sharkey KA. Cannabinoids and the gut: new developments and emerging concepts. Pharmacol Ther. 2010;126:21–38. doi: 10.1016/j.pharmthera.2009.12.005. [DOI] [PubMed] [Google Scholar]
- 59.Barutta F, Piscitelli F, Pinach S, Bruno G, Gambino R, Rastaldi MP, Salvidio G, Di Marzo V, Cavallo Perin P, Gruden G. Protective role of cannabinoid receptor type 2 in a mouse model of diabetic nephropathy. Diabetes. 2011;60:2386–2396. doi: 10.2337/db10-1809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Aguado T, Carracedo A, Julien B, Velasco G, Milman G, Mechoulam R, Alvarez L, Guzman M, Galve-Roperh I. Cannabinoids induce glioma stem-like cell differentiation and inhibit gliomagenesis. J Biol Chem. 2007;282:6854–6862. doi: 10.1074/jbc.M608900200. [DOI] [PubMed] [Google Scholar]
- 61.Blazquez C, Carracedo A, Barrado L, Real PJ, Fernandez-Luna JL, Velasco G, Malumbres M, Guzman M. Cannabinoid receptors as novel targets for the treatment of melanoma. FASEB J. 2006;20:2633–2635. doi: 10.1096/fj.06-6638fje. [DOI] [PubMed] [Google Scholar]
- 62.Grimaldi C, Capasso A. The endocannabinoid system in the cancer therapy: an overview. Curr Med Chem. 2011;18:1575–1583. doi: 10.2174/092986711795471374. [DOI] [PubMed] [Google Scholar]
- 63.Gustafsson K, Wang X, Severa D, Eriksson M, Kimby E, Merup M, Christensson B, Flygare J, Sander B. Expression of cannabinoid receptors type 1 and type 2 in non-Hodgkin lymphoma: growth inhibition by receptor activation. Int J Cancer. 2008;123:1025–1033. doi: 10.1002/ijc.23584. [DOI] [PubMed] [Google Scholar]
- 64.Gertsch J, Pertwee RG, Di Marzo V. Phytocannabinoids beyond the Cannabis plant - do they exist? Br J Pharmacol. 2010;160:523–529. doi: 10.1111/j.1476-5381.2010.00745.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Rajesh M, Mukhopadhyay P, Batkai S, Hasko G, Liaudet L, Huffman JW, Csiszar A, Ungvari Z, Mackie K, Chatterjee S, Pacher P. CB2-receptor stimulation attenuates TNF-alpha-induced human endothelial cell activation, transendothelial migration of monocytes, and monocyte-endothelial adhesion. Am J Physiol Heart Circ Physiol. 2007;293:H2210–2218. doi: 10.1152/ajpheart.00688.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Dursun B, He Z, Somerset H, Oh DJ, Faubel S, Edelstein CL. Caspases and calpain are independent mediators of cisplatin-induced endothelial cell necrosis. Am J Physiol Renal physiol. 2006;291:F578–587. doi: 10.1152/ajprenal.00455.2005. [DOI] [PubMed] [Google Scholar]
- 67.Montecucco F, Lenglet S, Braunersreuther V, Burger F, Pelli G, Bertolotto M, Mach F, Steffens S. CB(2) cannabinoid receptor activation is cardioprotective in a mouse model of ischemia/reperfusion. J Mol Cell Cardiol. 2009;46:612–620. doi: 10.1016/j.yjmcc.2008.12.014. [DOI] [PubMed] [Google Scholar]
- 68.Steffens S, Veillard NR, Arnaud C, Pelli G, Burger F, Staub C, Karsak M, Zimmer A, Frossard JL, Mach F. Low dose oral cannabinoid therapy reduces progression of atherosclerosis in mice. Nature. 2005;434:782–786. doi: 10.1038/nature03389. [DOI] [PubMed] [Google Scholar]
- 69.Montecucco F, Di Marzo V, da Silva RF, Vuilleumier N, Capettini L, Lenglet S, Pagano S, Piscitelli F, Quintao S, Bertolotto M, Pelli G, Galan K, Pilet L, Kuzmanovic K, Burger F, Pane B, Spinella G, Braunersreuther V, Gayet-Ageron A, Pende A, Viviani GL, Palombo D, Dallegri F, Roux-Lombard P, Santos RA, Stergiopulos N, Steffens S, Mach F. The activation of the cannabinoid receptor type 2 reduces neutrophilic protease-mediated vulnerability in atherosclerotic plaques. Eur Heart J. 2011 doi: 10.1093/eurheartj/ehr449. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Asakawa Y, Taira Z, Takemoto T, Ishida T, Kido M, Ichikawa Y. X-ray crystal structure analysis of 14-hydroxycaryophyllene oxide, a new metabolite of (−)-caryophyllene, in rabbits. J Pharm Sci. 1981;70:710–711. doi: 10.1002/jps.2600700642. [DOI] [PubMed] [Google Scholar]
- 71.Asakawa Y, Ishida T, Toyota M, Takemoto T. Terpenoid biotransformation in mammals. IV Biotransformation of (+)-longifolene, (−)-caryophyllene, (−)-caryophyllene oxide, (−)-cyclocolorenone, (+)-nootkatone, (−)-elemol, (−)-abietic acid and (+)-dehydroabietic acid in rabbits. Xenobiotica. 1986;16:753–767. doi: 10.3109/00498258609043566. [DOI] [PubMed] [Google Scholar]