

Abstract
Background
Delayed deterioration associated with vasospasm (DDAV) after aneurismal subarachnoid hemorrhage (SAH) is a major cause of morbidity. We have previously shown that myeloid cell depletion before experimental SAH in a murine model ameliorates DDAV. In this study, we address whether systemic administration of lipopolysaccharide (LPS) worsens DDAV in a myeloid cell-dependent fashion.
Methods
We challenged mice in our experimental SAH model with LPS before hemorrhage and evaluated the degree of vasospasm on day 6 with India ink angiography; behavioral deficits by rotorod, Y-maze and Barnes maze testing; microglial activation early after SAH by immunohistochemistry; and the brain levels of the chemokines CCL5 and KC at the time of vasospasm. Another group of animals were given the myeloid cell-depleting antibody against the neutrophil antigen Ly6G/C prior to LPS administration and SAH.
Results
LPS followed by SAH significantly worsens angiographic vasospasm as well as performance on the Barnes maze but not the Y-maze or rotorod tests. There was an increased activation of microglia in animals with LPS before SAH compared to SAH alone. Depletion of myeloid cells before LPS administration inhibited the development of vasospasm, improved the performance on behavioral tests and reduced microglial activation. The chemokines CCL5 and KC were incrementally elevated in SAH and LPS SAH but suppressed in animals with myeloid cell depletion.
Conclusions
LPS administration before SAH worsens DDAV through a myeloid cell-dependent mechanism supporting studies in humans which show that systemic inflammation increases the likelihood of developing DDAV.
Keywords: Subarachnoid hemorrhage, LPS, Delayed cerebral vasospasm, Innate immunity, Behavior
Introduction
Delayed neurological deterioration associated with vasospasm in aneurismal subarachnoid hemorrhage (SAH) is the major cause of morbidity and mortality in patients who survive the initial bleeding episode [1]. Clinical manifestations include encephalopathy, focal neurological signs and, in the most severe cases, stroke. This vexing disease presents itself with spasm of proximal arteries and delayed neurological deficits 4–12 days after the aneurysm rupture leaving a time window that could represent an opportunity to intervene before permanent brain damage occurs. Although the term-delayed cerebral ischemia (DCI) and delayed neurological ischemic deficits (DIND) have been used to describe the proposed cause of the injury, recent evidence suggests that mechanisms other than ischemia may be at play [2-4]. Unfortunately, focus on reversing vasoconstriction during the period of DCI has yielded disappointing results in patient outcomes [5]. A more appropriate term to describe the phenomenon may be delayed deterioration associated with vasospasm (DDAV) to denote the uncertainty in the cause of the brain injury.
Recently, there has been increased interest among some investigators in the role of early innate inflammation in both the vascular and cerebral manifestations of DDAV [6-8]. Myeloid cells, part of the innate immune response to infectious and non-infectious insults, are composed of neutrophils, monocytes and macrophages. Regulation of innate immune responses is complex and involve chemokine signals to attract cellular elements. Lipopolysaccharide A from Escherichia coli (LPS) is a known signaling molecule of the innate immune system mediated through the TLR4 receptor on the neutrophil and endothelial cell surface.
We have previously shown that the neutrophil percentage in the cerebrospinal fluid (CSF) early in the course of SAH can predict who will later develop DDAV[9]. Previous animal study has shown that early administration of modulators of innate inflammation can alter the course of the disease [8, 10-12]. Administration of an anti-CD11b antibody (against a major endothelial-signaling molecule for innate immune cells, ICAM) in an SAH model blocks vasospasm [10]. Direct administration of LPS into the CSF without SAH causes vasospasm [11]. More specifically, we have shown that myeloid cell depletion in a mouse model of DDAV ameliorates both the vascular and the behavioral effects [6].
The question remains as to whether systemic inflammatory signals in SAH patients precipitate or worsen DDAV. Recent studies in patients suggest that systemic inflammatory response syndrome (SIRS) in patients with SAH is associated with a greater risk of DDAV [13-15]. In this study, we investigate whether systemic administration of LPS worsens DDAV and whether this is mediated through myeloid cells.
Materials and Methods
All the experiments were conducted under the supervision of the Cleveland Clinic Institutional Animal Care and Use Committee (IACUC). Animals were randomized into three groups: (1) LPS administration followed by Sham surgery (LPS Sham), (2) LPS administration followed by SAH (LPS SAH), and (3) LPS administration followed by myeloid cell depletion followed by SAH (LPS SAH +Ly6G/C). All surgeries were done by one investigator (SS) who randomly assigned animals to each of the three treatment groups. Analysis of the perfusion experiments and all behavioral tests were done by a different investigator (SKM) blinded to the surgical assignments.
Previously published India ink experiments in animals with SAH and Sham are presented for comparison (to limit the number of animals euthanized in the present study) [6]. These studies were also randomized and blinded in the same way. These experiments were not included in the statistical analysis.
Experimental SAH
We studied male C57BL6 mice (Jackson Labs, Maine) weighing 20–32 g, 10–12 weeks old (Table 1). Our murine model of SAH has been described [16]. In Brief, mice were anesthetized and placed in a prone position. An incision was made in the midline of the neck, the atlanto-occipital membrane was punctured, and a subarachnoid vein was transected. The bleeding was allowed to stop spontaneously, after which the incision was closed. Saline injection sham surgery involved the same procedure except that the atlanto-occipital membrane was entered with a 30 gauge needle and 50 μl of saline was instilled. No animals died as a result of the surgery, one animal in the LPS group died before surgery after LPS administration. All animals that had surgery survived all the post-hemorrhage testing.
Table 1.
Animal experiments
| India ink | Behavioral test | Immunostaining and brain chemokines | |
|---|---|---|---|
| Sham | – | – | 6 |
| LPS Sham | 10 | 10 | 6 |
| SAH | – | – | 6 |
| LPS SAH | 10 | 10 | 6 |
| LPS SAH +Ly6G/C | 10 | 10 | 6 |
This model employs a venous hemorrhage instead of an arterial hemorrhage as was done in other models of SAH. There is evidence that arterial blood is more spasmogenic than venous blood in humans [1, 17]. Mice are more prone to vasospasm than humans (as evidenced by 100% vasospasm in all the models), which makes the venous model viable. It is likely that the relative mildness of the vasospasm compared to arterial models is the reason behavioral testing in our model is informative.
LPS Administration
200 mg/kg of LPS in 150 μl diluent (Sigma Aldrich, Saint Louis, MO) was injected into the peritoneal cavity of experimental animals 24 h prior surgery. This dosage was chosen based on previous studies of LPS-induced fever and systemic inflammation [18]. Pilot experiments with control animal given higher dose of LPS showed unacceptable mortality within 24 h.
The rationale for administering LPS before SAH is that patients with SAH often arrive at the hospital with evidence of SIRS, suggesting that the onset of inflammation is very early. We felt that the administration of LPS at the time of SAH would lead to unacceptable mortality. For this reason, we chose to administer the LPS 24 h before surgery to allow the animals to recover from the immediate LPS dose effects before the stress of surgery.
Myeloid Cell Depletion
Animals were pretreated with the myeloid cell-depleting anti-ly6G/C antibody (RB6-8C5) at a dose of 13 mg/kg i.p. 24 h before surgery and immediately before surgery, or 50 μl of saline (saline control). The dose of RB6-8C5 was chosen based on our previous studies [6]. Pilot experiments with control animals given pretreatment with the irrelevant isotype-matched antibody, anti-β-Gal (GL113) before SAH yielded similar responses in blood vessel caliber to saline controls, but had increased rigors and lethargy. We therefore did not include this analysis and used saline control instead.
India Ink Assessment of Vessel Caliber
Animals were anesthetized with pentobarbital (6 mg/100 g i.p.); trans-cardiac perfusion was performed with 20 ml 4% paraformaldehyde followed by 10 ml of warmed 5% India ink in gelatin. Animals were decapitated, and their brains were removed, carefully preserving the vasculature. The circle of Willis vasculature was examined under the surgical microscope, and relevant pictures were captured (Leica, Wetzler, Germany) and analyzed with Adobe Photoshop CS2 (San Jose CA). The diameter of the middle cerebral artery (MCA) segment was measured 1 mm from the posterior wall of the carotid artery by a member of the research team not involved in the surgery and blinded to the intervention. All the measurements were made on the 6th day after the hemorrhage based on a previous study wwhich shows that DCI occurs around the 6th post-hemorrhage day in our model [6]. Ten mice were used for each of the three conditions (LPS Sham, LPS SAH, and myeloid cell-depleted LPS SAH) (Table 1).
Behavioral Testing
Animals in the behavioral study underwent three batteries of tests in groups of 10 mice. All the tests were done at the same time of the day by a single handler (SKM) who was blinded to the treatment allocation. Groups of 5–10 animals randomized for treatment and blinded to the handler were tested together. This was repeated until 10 animals from each group were completed. Due to the effect of LPS injection, all mice were allowed 24 h after surgery to recover. On days 2 after the hemorrhage, the mice underwent rotarod testing to evaluate motor function and coordination. On the 3rd day post hemorrhage, the mice underwent Y-maze testing to evaluate spatial working (immediate) memory. On days 2 and 3, the animals also had training for the Barnes maze. On days 4 through 9, the mice were tested in a Barnes maze to test spatial learning, short-term and long-term memory.
In the rotarod test, time on the rod and maximum speed of rotation (RPM) were measured. Animals were placed in one of the four separated chambers with the spinning rod and a landing base. After a habituation for 30 s at low speed, the mice were tested with a continuously increasing rate of speed for 5 min or until the subject fell off the rod.
In the Y-maze, animals were placed in a random orientation in the center of a high-walled maze that has three equal-length passages in a Y configuration. The mice were recorded by video monitoring during 5 min of exploration and scored later. Percent alterations were calculated as the ratio of the number of times the animals chose to enter the three arms of the maze in succession to the total number of unique arms the animal chose to enter.
In the Barnes maze, the mice are placed on a round table with multiple open holes in the periphery one of which contains an under-mounted box that represents a safe haven for the mice. There are visual cues on the wall around the table for the animal to orient itself. Each animal had 2 days of training during which it was gently guided to the goal box. On subsequent days, animals were placed in the center of the table in a covered enclosure until ready for the trial. The cover was lifted and mice were allowed to find the goal box. The movements were recorded by video monitoring and scored later. The time it took to find the goal box (latency) was recorded.
Immunohistochemistry
Five groups of animals were tested in this experiment: (1) Sham, (2) SAH, (3) LPS Sham, (4) LPS SAH, and (5) LPS SAH +Ly6G/C. The monoclonal anti-microglial cell antibody (IBa1) (gift from Dr. Bruce Trapp) and the monoclonal anti-neutrophil antibody (7/4) (Serotec, Raleigh, NC) were used. 30-μm brain slices were incubated in primary antibody for 24 h using a free-floating technique. In brief, slices were microwaved for 2 min in citrate buffer followed by incubation in 500 μl of 3% goat serum and primary antibody at 20°C overnight in a 24-well plate. Secondary antibodies were resolved with either 3,3′-diaminobenzadine (DAB) (Sigma-Aldrich, Saint Louis, MO).
Chemokine Determination
Five groups of animals were tested in this experiment: (1) Sham, (2) SAH, (3) LPS Sham, (4) LPS SAH, and (5) LPS SAH +Ly6G/C. Brain inflammatory chemokines levels were determined by Dot–Blot analysis (R&D Systems, Minneapolis, MN) 24 h after surgery. Animals were anesthetized with pentobarbital (6 mg/100 g i.p.); trans-cardiac perfusion was performed with 20 ml 1% PBS. Animals were decapitated, and their brains were removed carefully and homogenized. After tissue protein extraction, the supernatant was incubated on the Dot–Blot membrane. Radioactive dots were visualized by radiography. Dot intensities were measured and normalized to a positive control, and analyzed with Image J (NIH, Bethesda, MD).
Statistics
Statistical analysis was performed with the aid of Graphpad Prism 5.0 (Graphpad Software Inc. San Diego, CA). Multiple comparisons were assessed using 1-way ANOVA. Comparisons between individual groups in experiments where 1-way ANOVA was significantly different were assessed using Tucky’s Multiple Comparison Test. Two-way ANOVA was used to evaluate the effects of SAH over time. A P-value of 0.05 was considered significant in all comparisons.
Results
Animals
Mice were grouped into three categories: (1) LPS administration followed by Sham surgery (LPS Sham), (2) LPS administration followed by SAH (LPS SAH), and (3) LPS administration followed by myeloid cell depletion followed by SAH (LPS SAH +Ly6G/C). Results from animals in a previous study, which underwent SAH and sham SAH, are shown in the vessel caliber study for comparison because the conditions of the experiment were identical (data published in [6]). Additional immunohistochemistry and chemokine analysis were done in Sham and SAH animals for this study.
LPS Followed by SAH Results in Vasospasm that is Reversed by Myeloid Cell Depletion
There was 1 preoperative death in the LPS injection group, which occured within 24 h after LPS injection before SAH. There was no operative mortality from the experimental SAH. After surgery, all the animals returned to feeding and were ambulating normally within 3 h.
With regard to day 6 vessel caliber, the three groups were significantly different (1-way ANOVA: F(2, 12) = 23.22, P < 0.001). In mice subjected to LPS injection 24 h before experimental SAH, there were significant narrowings of the MCA compared to LPS Sham (0.060 mm ± 0.005 vs. 0.118 mm ± 0.005 [mean ± SEM], P < 0.001) (Fig. 1a).
Fig. 1.
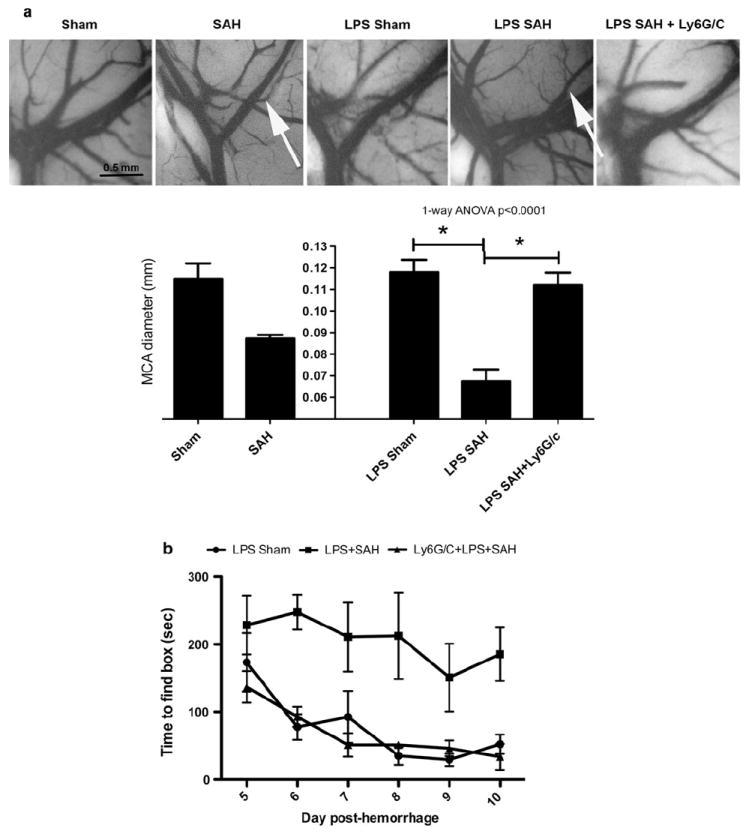
a India ink angiograms of mice on the 6th day. Middle cerebral artery (MCA) angiography showing no narrowing (the bar graph below shows the mean MCA diameter) in LPS sham animals. LPS SAH mice show significant narrowing of the MCA (white arrows) showing DDAV. In SAH mice administered with the myeloid cell-depleting antibody against Ly6G/C coincident with LPS, there is the absence of vasospasm. *The sham and SAH comparators were previously published and are included for comparison [8]. b Behavioral deficits seen on Barnes maze testing in mice with SAH after LPS administration. It takes significantly longer for mice with SAH after LPS to find the goal box than for animals that have had LPS followed by sham surgery (2-way ANOVA P < 0.001). Mice that have myeloid cell depletion concurrent with LPS administration and SAH have times to find goal boxes that are significantly shorter than LPS SAH (2-way ANOVA P < 0.001) and are similar to sham animals
Behavioral testing was done in ten mice per group and included rotorod testing to determine if there were physical deficits that could confound Y-maze and Barnes maze cognitive tests. Rotorod testing of mice after experimental SAH showed no evidence of motor deficits or in-coordination 3 days post hemorrhage in any group either by assessment of time on the rod or maximum RPMs. Y-maze testing in SAH and sham revealed no significant differences in percent alternations in the three experimental groups. Barnes maze testing done every day after a two-day education period showed that the three groups were significantly different. LPS/SAH animals took significantly longer to find the goal box (latency) than LPS/Sham animals (ANOVA P < 0.001) (Fig. 1b).
The diameter of the MCA was significantly larger in myeloid cell-depleted LPS/SAH animals than in LPS/SAH animals at the 6th day after hemorrhage (0.112 mm ± 0.005 vs. 0.067 ± 0.005 [mean ± SEM], P = 0.005) (Fig. 1a). Myeloid cell-depleted LPS SAH mice showed no difference from control or non-depleted SAH mice in rotarod testing and Y maze testing (data not shown). The LPS/SAH mice showed significant delayed time to find the goal box from day 5 to 10 after hemorrhage compared with myeloid cell-depleted LPS/SAH mice (ANOVA P < 0.001) (Fig. 1b). Myeloid cell-depleted LPS SAH mice showed no difference from LPS Sham in Barnes maze testing from day 5 to 10 after hemorrhage. These data support the hypothesis that Ly6G/C+ cell depletion protects from delayed behavioral deficits after SAH in LPS injected animals.
LPS Exacerbates Microglia Activation and Brain Inflammatory Chemokines, which is Reversed by Myeloid Cell Depletion
Microglia in LPS SAH mice showed impressive activation defined by thickened processes and enlarged cell bodies (ameboid morphology) compare to the SAH animals 1 day after SAH. Minimal microglia activation was seen in Sham and LPS/SAH myeloid-depleted animals (Fig. 2). Although in the SAH mice, the majority of microglia were activated, the highest density of activated microglia were found near the basal cistern where the blood clot was concentrated.
Fig. 2.
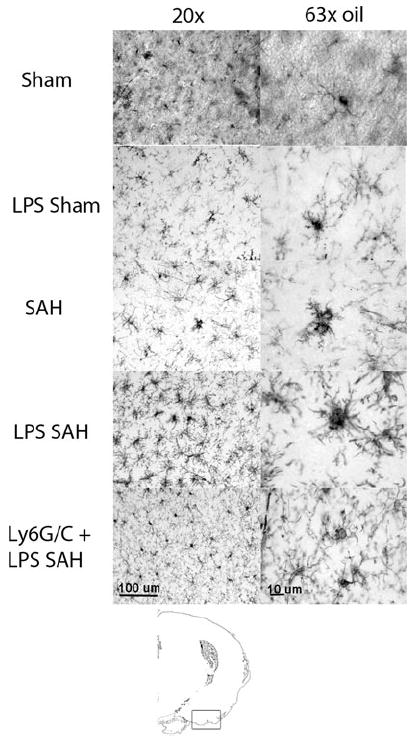
Immunohistochemistry of microglia 1 day after SAH at two magnifications (antibody, Iba1). Sham animals show rest appearing microglia with multiple long fine cell projections. LPS Sham mice show slightly more retracted processes (a sign of activation). SAH animals show ameboid morphology where the process retraction is severe, and the cells take on the appearance of macrophages. LPS SAH animals show even more densely packed projections and rounding of the cell bodies. Finally, in animals that are given a myeloid cell-depleting antibody again Ly6G/C, the microglia animals again exhibit a resting phenotype seen in the Sham animals. The diagram at the bottom shows the area of the brain imaged
The inflammatory chemokines keratinocyte-derived chemokine (KC) and CCL5 were elevated at day 6 after hemorrhage. LPS SAH mice had higher KC and CCL5 levels compared with LPS Sham for KC (96.1% of positive control ±1 vs. 57.0% ±4.1, P = 0.008) and CCL5 (61.7% ± 3.3 vs. 92.7% ± 1.5, P = 0.001), reversible by myeloid depletion for KC (96.1% ± 1.0 vs. 14.3% ± 4.7, P < 0.0001) and CCL5 (61.7% ± 3.3 vs. 11.7% ± 3.3, P < 0.001) (Fig. 3).
Fig. 3.
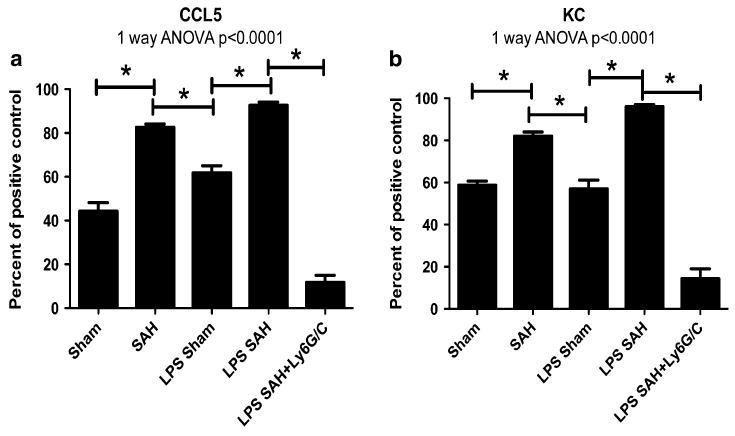
Levels of CCL5 and KC chemokines in the brain. Levels of CCL5 (a) and KC (b) increase with SAH compared with Sham in both non-LPS and LPS-treated mice. LPS treatment before SAH increases the level significantly from SAH. Myeloid cell depletion significantly decreases level
Discussion
In this study, we have shown that systemic administration of LPS worsens DCI after experimental murine SAH. In Addition, immunohistochemistry of the brain suggests that in addition to worsening vascular disease, there is extensive microglial activation suggesting direct brain inflammation. Both these findings are associated with behavioral abnormalities on Barnes maze testing analogous to the cognitive dysfunction seen in patients with SAH [19, 20]. Interestingly, there were no abnormalities seen on the Y maze test, suggesting that not all cognitive functions are equally affected. Levels of the chemokines KC and CCL5 suggest that neutrophils are part of the primary pathophysiology of DDAV.
We have shown that cells from the innate immune system are implicated in the development of DDAV [6]. Previous studies based on the myeloid cell recruitment molecules ICAM and CD11/18 support this mechanism [10, 21, 22]. The next logical question is whether activation of the peripheral inflammatory system worsens DDAV. Although there is evidence that intrathecal LPS can precipitate DDAV without hemorrhage, peripheral activation of innate immune cells in this study suggests that the systemic innate immune system affects brain damage [11]. Our study supports the finding by Dhar and colleagues and Tam and colleagues that activation of the systemic innate immune system makes DDAV more likely and more severe [13, 14].
The concept that inflammation is an important mediator of DDAV has developed over the last 10–15 years, but remains controversial. Because the location of the blood clot is correlated with the blood vessels involved in DDAV, blood breakdown products have been implicated as part of the pathogenesis of DDAV. There are now studies that link inflammation to the red blood cell breakdown product bilirubin oxidation products (BOXs) and the haptoglobin α2 genotype [9, 23, 24].
LPS is a selective TLR4-receptor agonist that is in part responsible for the vasodilatory shock seen in the systemic inflammatory response syndrome (SIRS). TLR4 is a receptor located predominantly on cells from the myeloid hematopoetic cell lineage and is associated with activation of innate immune responses. Although LPS alone is not responsible for SIRS, it is accepted to act as an agent for innate immune activation in murine models.
There is growing evidence that vascular narrowing and ischemia from vasospasm may not be solely responsible for the deficits experienced by patients [4]. In this model, the mice reliably develop vasospasm on day 6, but do not develop behavioral deficits until a few days later. Interestingly, LPS worsens both vasospasm and cognitive behavioral testing, suggesting that regardless of whether the damage is ischemic or primarily inflammatory, LPS modulates the effect. This is in contrast to human clinical trials where the vascular and behavioral effects dissociate with treatment [2].
Chemokines are small chemoattractant cytokines that are produced in response to damage or infection to call inflammatory cells into the area. KC is the murine homolog of CXCL8 (IL-8) and CXCL1 (Gro-α) and is important in attracting neutrophils to sites of damage. CCL5 (RANTES) is important for attracting secondary inflammatory agents such as T-cells and monocytes. Typically, the presence of the attracted cell works as a feedback mechanism to down-regulate the production of the chemokine. Therefore, the behavior of the chemokines CCR5 and KC in this study may give a clue about the source of the inflammation signal in SAH. If the primary signal for inflammation in SAH is blood or blood breakdown products, then one would expect that the LPS would have a modest effect on chemokine production but that the absence of neutrophils after depletion in mice would increase the chemokine level (particularly KC) precipitously since the primary down-regulatory signal from myeloid cells would be absent [25]. This is not what we actually see. In fact, the chemokine levels are much lower, suggesting decreased inflammatory signals. This correlates well with the histology data showing less microglial activation. This suggests that myeloid cells are the instigators of inflammation in this system. Further studies are necessary to elucidate the precise signals that begin the inflammatory cascade after SAH.
In conclusion, we believe that this study helps clarify the role of inflammation in DDAV, but further work is necessary to determine the exact role of innate immunity in DDAV. It is possible that treatment strategies could be developed around immune modulation of the myeloid cells in DDAV.
Acknowledgments
The authors would like to thank James Bena for the statistical assistance. This work was supported by a grant from the NIH (K08-NS051350 JJP) and the Cleveland Clinic Cerebrovascular Center (Institutional Support JJP).
Footnotes
Conflict of interest The authors declared that they have no conflicts of interest.
Contributor Information
Saksith Smithason, Cerebrovascular Center, Cleveland Clinic, NC30, 9500 Euclid Ave., Cleveland, OH 44195, USA; Neuroinflammation Research Center, Cleveland Clinic, NC30, 9500 Euclid Ave., Cleveland, OH 44195, USA.
Shari Korday Moore, Neuroinflammation Research Center, Cleveland Clinic, NC30, 9500 Euclid Ave., Cleveland, OH 44195, USA.
J. Javier Provencio, Email: provenj@ccf.org, Cerebrovascular Center, Cleveland Clinic, NC30, 9500 Euclid Ave., Cleveland, OH 44195, USA; Neuroinflammation Research Center, Cleveland Clinic, NC30, 9500 Euclid Ave., Cleveland, OH 44195, USA.
References
- 1.van Gijn J, Rinkel GJ. Subarachnoid haemorrhage: diagnosis, causes and management. Brain. 2001;124:249–78. doi: 10.1093/brain/124.2.249. [DOI] [PubMed] [Google Scholar]
- 2.Etminan N, Vergouwen MD, Ilodigwe D, Macdonald RL. Effect of pharmaceutical treatment on vasospasm, delayed cerebral ischemia, and clinical outcome in patients with aneurysmal subarachnoid hemorrhage: a systematic review and meta-analysis. J Cereb Blood Flow Metab. 2011;31(6):1443–51. doi: 10.1038/jcbfm.2011.7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Stein SC, Levine JM, Nagpal S, LeRoux PD. Vasospasm as the sole cause of cerebral ischemia: how strong is the evidence? Neurosurg Focus. 2006;21:E2. doi: 10.3171/foc.2006.21.3.2. [DOI] [PubMed] [Google Scholar]
- 4.Vergouwen MD, Ilodigwe D, Macdonald RL. Cerebral infarction after subarachnoid hemorrhage contributes to poor outcome by vasospasm-dependent and -independent effects. Stroke. 2011;42:924–9. doi: 10.1161/STROKEAHA.110.597914. [DOI] [PubMed] [Google Scholar]
- 5.Provencio JJ, Vora N. Subarachnoid hemorrhage and inflammation: bench to bedside and back. Semin Neurol. 2005;25:435–44. doi: 10.1055/s-2005-923537. [DOI] [PubMed] [Google Scholar]
- 6.Provencio JJ, Altay T, Smithason S, Moore SK, Ransohoff RM. Depletion of Ly6G/C(+) cells ameliorates delayed cerebral vasospasm in subarachnoid hemorrhage. J Neuroimmunol. 2010;232(1–2):94–100. doi: 10.1016/j.jneuroim.2010.10.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Clatterbuck RE, Gailloud P, Ogata L, et al. Prevention of cerebral vasospasm by a humanized anti-CD11/CD18 monoclonal antibody administered after experimental subarachnoid hemorrhage in nonhuman primates. J Neurosurg. 2003;99:376–82. doi: 10.3171/jns.2003.99.2.0376. [DOI] [PubMed] [Google Scholar]
- 8.Zubkov AY, Tibbs RE, Aoki K, Zhang JH. Prevention of vasospasm in penetrating arteries with MAPK inhibitors in dog double-hemorrhage model. Surg Neurol. 2000;54:221–7. doi: 10.1016/s0090-3019(00)00290-1. [DOI] [PubMed] [Google Scholar]
- 9.Provencio JJ, Fu X, Siu A, Rasmussen PA, Hazen SL, Ransohoff RM. CSF neutrophils are implicated in the development of vasospasm in subarachnoid hemorrhage. Neurocrit Care. 2010;12:244–51. doi: 10.1007/s12028-009-9308-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Pradilla G, Wang PP, Legnani FG, Ogata L, Dietsch GN, Tamargo RJ. Prevention of vasospasm by anti-CD11/CD18 monoclonal antibody therapy following subarachnoid hemorrhage in rabbits. J Neurosurg. 2004;101:88–92. doi: 10.3171/jns.2004.101.1.0088. [DOI] [PubMed] [Google Scholar]
- 11.Recinos PF, Pradilla G, Thai Q-A, Perez M, Hdeib AM, Tamargo RJ. Controlled release of lipopolysaccharide in the subarachnoid space of rabbits induces chronic vasospasm in the absence of blood. Surg Neurol. 2006;66:463–9. doi: 10.1016/j.surneu.2006.04.010. [DOI] [PubMed] [Google Scholar]
- 12.Zhou C, Yamaguchi M, Kusaka G, Schonholz C, Nanda A, Zhang JH. Caspase inhibitors prevent endothelial apoptosis and cerebral vasospasm in dog model of experimental subarachnoid hemorrhage. J Cereb Blood Flow Metab. 2004;24:419–31. doi: 10.1097/00004647-200404000-00007. [DOI] [PubMed] [Google Scholar]
- 13.Dhar R, Diringer MN. The burden of the systemic inflammatory response predicts vasospasm and outcome after subarachnoid hemorrhage. Neurocrit Care. 2008;8:404–12. doi: 10.1007/s12028-008-9054-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Tam AK, Ilodigwe D, Mocco J, et al. Impact of systemic inflammatory response syndrome on vasospasm, cerebral infarction, and outcome after subarachnoid hemorrhage: exploratory analysis of CONSCIOUS-1 database. Neurocrit Care. 2010;13:182–9. doi: 10.1007/s12028-010-9402-x. [DOI] [PubMed] [Google Scholar]
- 15.Yoshimoto Y, Tanaka Y, Hoya K. Acute systemic inflammatory response syndrome in subarachnoid hemorrhage. Stroke. 2001;32:1989–93. doi: 10.1161/hs0901.095646. [DOI] [PubMed] [Google Scholar]
- 16.Altay T, Smithason S, Volokh N, Rasmussen PA, Ransohoff RM, Provencio JJ. A novel method for subarachnoid hemorrhage to induce vasospasm in mice. J Neurosci Methods. 2009;183:136–40. doi: 10.1016/j.jneumeth.2009.06.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Goergen SK, Barrie D, Sacharias N, Waugh JR. Perimesencephalic subarachnoid haemorrhage: negative angiography and favourable prognosis. Australas Radiol. 1993;37:156–60. doi: 10.1111/j.1440-1673.1993.tb00040.x. [DOI] [PubMed] [Google Scholar]
- 18.Hang CH, Shi JX, Tian J, Li JS, Wu W, Yin HX. Effect of systemic LPS injection on cortical NF-kappaB activity and inflammatory response following traumatic brain injury in rats. Brain Res. 2004;1026:23–32. doi: 10.1016/j.brainres.2004.07.090. [DOI] [PubMed] [Google Scholar]
- 19.Powell J, Kitchen N, Heslin J, Greenwood R. Psychosocial outcomes at 18 months after good neurological recovery from aneurysmal subarachnoid haemorrhage. J Neurol Neurosurg Psychiatry. 2004;75:1119–24. doi: 10.1136/jnnp.2002.000414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Noble AJ, Baisch S, Mendelow AD, Allen L, Kane P, Schenk T. Posttraumatic stress disorder explains reduced quality of life in subarachnoid hemorrhage patients in both the short and long term. Neurosurgery. 2008;63:1095–104. doi: 10.1227/01.NEU.0000327580.91345.78. [DOI] [PubMed] [Google Scholar]
- 21.Clatterbuck RE, Oshiro EM, Hoffman PA, Dietsch GN, Pardoll DM, Tamargo RJ. Inhibition of vasospasm with lymphocyte function-associated antigen-1 monoclonal antibody in a femoral artery model in rats. J Neurosurg. 2002;97:676–82. doi: 10.3171/jns.2002.97.3.0676. [DOI] [PubMed] [Google Scholar]
- 22.Aihara Y, Kasuya H, Onda H, Hori T, Takeda J. Quantitative analysis of gene expressions related to inflammation in canine spastic artery after subarachnoid hemorrhage. Stroke. 2001;32:212–7. doi: 10.1161/01.str.32.1.212. [DOI] [PubMed] [Google Scholar]
- 23.Pyne-Geithman GJ, Morgan CJ, Wagner K, et al. Bilirubin production and oxidation in CSF of patients with cerebral vasospasm after subarachnoid hemorrhage. J Cereb Blood Flow Metab. 2005;25:1070–7. doi: 10.1038/sj.jcbfm.9600101. [DOI] [PubMed] [Google Scholar]
- 24.Chaichana KL, Levy AP, Miller-Lotan R, Shakur S, Tamargo RJ. Haptoglobin 2–2 genotype determines chronic vasospasm after experimental subarachnoid hemorrhage. Stroke. 2007;38:3266–71. doi: 10.1161/STROKEAHA.107.490003. [DOI] [PubMed] [Google Scholar]
- 25.Ellis TN, Beaman BL. Murine polymorphonuclear neutrophils produce interferon-gamma in response to pulmonary infection with Nocardia asteroides. J Leukoc Biol. 2002;72:373–81. [PubMed] [Google Scholar]