

Abstract
Bradykinin is a physiologically active peptide involved in vasodilation and smooth muscle contraction and is previously shown to be increased in gastrointestinal mucus during nematode challenge in sheep. Here, it is shown that bradykinin in the abomasum is positively correlated with both mast cells and globule leukocytes in the abomasum, and that all three of these parameters are negatively associated with numbers of adult Teladorsagia circumcincta during the challenge of immune sheep. It is suggested that bradykinin either stimulates the degranulation of mast cells, or is released during this degranulation process, or both. Multiple regression showed that almost 60% of the variation of in adult T. circumcincta could be explained by two variables, bradykinin and T. circumcincta—specific IgG1 in plasma. This provides further evidence that bradykinin may be a mechanism of protective immunity in sheep, although its involvement in asthma and other allergic disorders raises questions about its role in unwanted immunopathology.
1. Introduction
Helminth infections invariably invoke Th2-like immune responses in mammalian hosts, characterised by production of IgE, IgG1, mucosal mast cells, and eosinophils [1]. In addition, we have recently shown that the peptide bradykinin is significantly increased in abomasal mucus during helminth challenge in immune sheep [2]. Bradykinin is a potent vasodilator that, during inflammation, is cleaved from circulating kininogen and is normally associated with endothelial permeability and nonvascular smooth muscle contraction in conditions such as asthma [3, 4]. It was found that abomasal bradykinin was higher in sheep with lower total worm burdens, suggesting that it may be involved with protective immunity. However, it is not clear what role bradykinin plays and how it interacts with other immune mechanisms. The present communication reports associations between bradykinin and granulocytes and antibodies produced during helminth challenge in sheep.
2. Methods
2.1. Parasites and Animals
The experimental design of the animal trial is described in detail in Williams et al. [2]. Forty Merino sheep were housed indoors and challenged daily with 500 third-stage larvae each of Teladorsagia circumcincta and Trichostrongylus colubriformis. The sheep were from a line selectively bred for low faecal worm egg count (FEC) and had been raised on pastures contaminated with helminth larvae and thus had acquired a degree of protective immunity [5]. Total worm burdens were quantified after six weeks of continuous challenge.
2.2. Immunological Measurements
IgG1 specific to T. circumcincta and T. colubriformis L3 antigen was determined by ELISA four weeks after challenge began and was reported as ELISA units relative to a positive control (high-reading sera). At postmortem, tissue samples were taken from the abomasum and proximal third of the small intestine, fixed in 4% paraformaldehyde and the numbers of mast cells, globule leukocytes, and eosinophils enumerated by standard histological methods and expressed as cells per/mm2 of tissue. Mucus was snap-frozen in liquid nitrogen for bradykinin analysis. Bradykinin concentrations were measured in mucus after purification by solid-phase extraction, using a commercial ELISA kit according to the manufacturers instructions. Bradykinin levels were expressed per mg of protein in mucus samples. Further details are provided in Williams et al. [2, 6].
2.3. Statistical Analysis
Worm burden data was transformed (log10(N + 10)) before analysis. Other data was normally distributed and analysed untransformed. Simple and multiple regression was used to determine correlations and relationships between variables, using the STATISTICA program (version 10, StatSoft, Tulsa, OK, USA).
3. Results
Concentrations of bradykinin in the abomasum were positively correlated (r = 0.66, P < 0.001) with concentrations of bradykinin in the small intestine (Figure 1). However, the regression accounted for only 44% of the variation in abomasal bradykinin concentration (Table 1). Similarly, numbers of all three types of cell measured (eosinophils, mast cells, and globule leukocytes) were positively correlated between the two sites of infection (Table 1).
Figure 1.
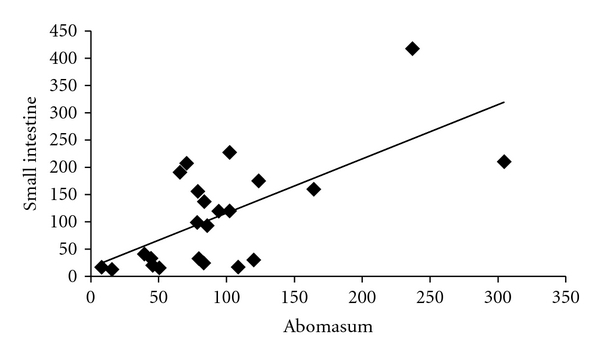
Relationship between concentrations of bradykinin (pg/mg protein) in abomasal mucus and the corresponding concentration in small intestinal mucus. r = 0.66, P < 0.001.
Table 1.
Regression models for the relationship between granulocytes and bradykinin in the abomasum (A) and small intestine (S).
| Model | n | Equation | r 2 | P value |
|---|---|---|---|---|
| Eosinophils (E) | 40 | EA = (0.37 ± 0.14)ES + 11.5 ± 14.1 | 0.14 | 0.018 |
| Mast cells (MC) | 40 | MCA = (0.38 ± 0.12)MCS + 34.4 ± 16.8 | 0.15 | 0.016 |
| Globule leukocytes (GL) | 40 | GLA = (0.35 ± 0.08)GLS + 16.7 ± 5.8 | 0.18 | 0.013 |
| Bradykinin (BK) | 24 | BKA = (0.66 ± 0.16)BKS + 45.9 ± 11 | 0.44 | <0.001 |
The relationships between immune mechanisms and parasite burdens were reported in Williams et al. [2]. Briefly, bradykinin in the abomasum was negatively correlated (r = −0.62, P < 0.001) with adult T. circumcincta but not larval stages of T. circumcincta. In contrast, there was no relationship between intestinal bradykinin and numbers of T. colubriformis, nor was intestinal bradykinin correlated with numbers of granulocytes in the small intestine mucosa. Abomasal bradykinin was positively associated with both mast cells and globule leukocytes in the abomasum (Figure 2). Mast cells and globule leukocytes in the abomasum were also negatively correlated with numbers of adult T. circumcincta (Table 2), but multiple regression showed that the same amount of variation in T. circumcincta number was explained by bradykinin alone or when added to the model together with mast cells, or globule leukocytes (Table 2). In fact, neither mast cells or globule leukocytes were a significant variable when bradykinin was added to the model, suggesting that it is bradykinin rather than granulocytes which is responsible for the majority of the variation in T. circumcincta number.
Figure 2.
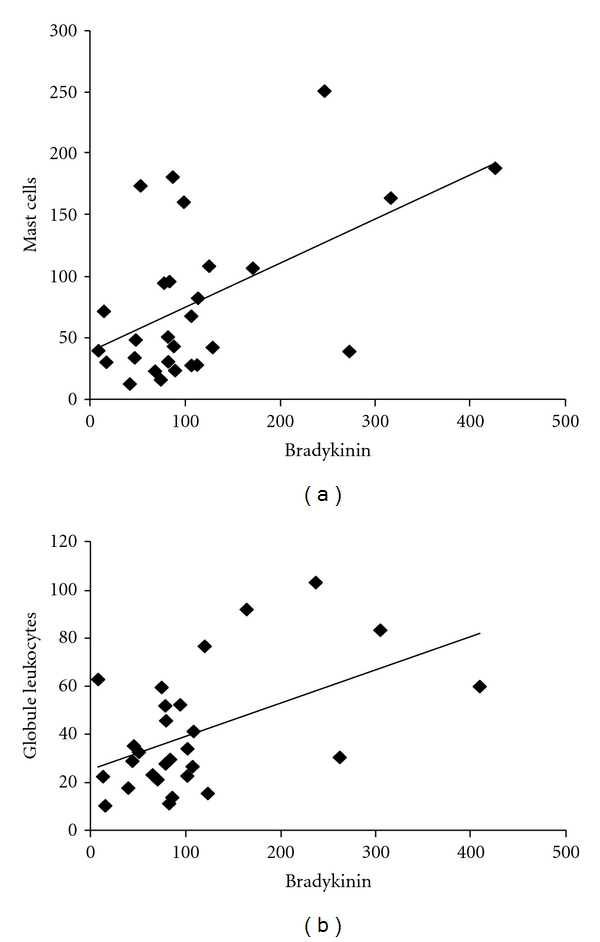
Relationships between the concentration of bradykinin in abomasal mucus and the numbers of mast cells (r = −0.53, P = 0.003, n = 28) and the numbers of globule leukocytes (r = −0.5, P = 0.006, n = 28) in the abomasum.
Table 2.
Regression equations for the relationship between log10 adult T. circumcincta burden (Tcirc) and the concentrations of bradykinin in the abomasum (BK), numbers of mast cells (MC) and globule leukocytes (GL) in the abomasum, and concentrations of T. circumcincta-specific IgG1 in plasma (IgG).
| Model | N | R 2 | Equation | P-value | |
|---|---|---|---|---|---|
| BK | 28 | 0.38 | Tcirc = (−0.62 ± 0.15)BK + 336 ± 58.3 | 0.0004 | |
| MC | 40 | 0.25 | Tcirc = (−0.5 ± 0.17)MC + 3 ± 0.18 | 0.006 | |
| GL | 40 | 0.1 | Tcirc = (−0.31 ± 0.15)GL + 3 ± 0.21 | 0.051 | |
| BK + MC | 28 | 0.35 | BK | Tcirc = (−0.58 ± 0.13)BK | 0.003 |
| MC | Tcirc = (−0.23 ± 0.09)MC + 2 ± 0.04 | 0.19 | |||
| BK + GL | 28 | 0.38 | BK | Tcirc = (−0.56 ± 0.18)BK | 0.005 |
| GL | Tcirc = (−0.12 ± 0.18)GL + 3 ± 0.21 | 0.52 | |||
| IgG | 40 | 0.3 | Tcirc = (−0.56 ± 0.16)IgG + 4 ± 0.41 | <0.0001 | |
| BK + IgG | 28 | 0.58 | BK | Tcirc = (−0.51 ± 0.14)BK | 0.001 |
| IgG | Tcirc = (−0.44 ± 0.14)IgG + 4 ± 0.34 | 0.004 |
IgG1 specific for T. circumcincta was negatively correlated with adult T. circumcincta burdens (r = −0.57, P < 0.01), but not correlated with bradykinin. When bradykinin and IgG1 were included in a multiple regression analysis, they accounted for almost 60% of the variation in total adult T. circumcincta burdens, compared to 30% for IgG1 alone and 38% for bradykinin alone (Table 2).
4. Discussion
Bradykinin is clearly associated with immunity to T. circumcincta in sheep. The mechanisms whereby bradykinin production may contribute to nematode expulsion have not been elucidated. However, given bradykinin's role in vasodilation and airway bronchoconstriction in asthma, it may be that it contributes to mucus hypersecretion, plasma leakage into the gut, and increased peristalsis—similar to the role proposed for other inflammatory mediators such as leukotrienes that have been repeatedly been shown to be involved in immunity to gastrointestinal nematodes [2, 7, 8]. A similar relationship between bradykinin and T. colubriformis was not observed in this experiment, but that is likely a result of the experimental sheep or being extremely resistant to infection with this parasite, and thus, there was little variation to determine relationships between worm burdens and immune effector mechanisms [2].
Eosinophils, mast cells, and globule leukocytes are invariably increased in the gut mucosa during nematode infection in sheep and thought to be involved in protective immunity [1], and the positive correlations shown here between cell numbers in adjacent segments of the gut indicate that there appears to be a “generalised” nematode immunity, that is, sheep that have a heightened cellular response to one parasite also have this high response against a second parasite species. This suggests that sheep bred to be resistant to nematodes on the basis of exposure to one parasite species may also exhibit increased immunity when challenged with a heterologous species. The positive correlation between bradykinin concentrations in the abomasum and small intestine, also provides evidence for this concept.
The associations between bradykinin and mast cells/globule leukocytes suggest a close relationship between these two mechanisms. Consistent with this, mast-cell-derived heparin has recently been shown to initiate bradykinin formation in mouse experiments, leading to marked vascular permeability [9]. A similar process may operate in sheep, whereby mast cells stimulated to degranulate by nematode antigens lead to bradykinin formation which directly leads to plasma leakage into the gut. This leakage is thought to be an important component of protective immunity to gastrointestinal parasites, perhaps due to plasma leakage allowing antibodies to come into contact with nematodes at the mucosa/lumen interface. Consistent with this hypothesis, regression analysis in the current paper showed that bradykinin and IgG1 together accounted for a large proportion of the variation in adult worm number. This could be due to mast cell derived-bradykinin dilating blood vessels surrounding the gut mucosa, allowing IgG to come into direct contact with incoming larvae and facilitating their expulsion. Further experiments will be necessary to more rigorously test this hypothesis. In vitro studies would be particularly useful to determine the interplay among antigen-dependent activation of mast cells, bradykinin formation, and subsequent parasite growth inhibition. In addition, the contribution of other mechanisms may be important such as mucosal IgA, which was not measured here and has previously been reported to influence worm length and fecundity [10].
A further aspect to be considered is whether increased levels of bradykinin may have unwanted pathological side effects. It has been noted several times that sheep that are highly resistant to helminth parasites can suffer an increase in diarrhoea during natural grazing, and it has been speculated that this may be due to a hypersensitive mucosal immune response leading to larval rejection whilst also causing unwanted immunopathology. Given bradykinin's role in allergic disorders such as asthma, further studies should investigate this relationship in order to determine whether bradykinin may be suitable as a phenotypic marker for nematode resistance in sheep. However, it has previously been reported with these experimental animals that bradykinin levels were not associated with faecal softening during parasite challenge [2].
In conclusion, bradykinin is associated with the mast-cell response to nematode larval challenge in immune sheep and may function together with well-known immune mechanisms such as IgG1 to facilitate nematode expulsion.
Acknowledgments
The author is grateful to P. Vercoe, D. Palmer, I. Williams, and J. Karlsson for their contribution to this work. This work was supported by the Australian Government and Australian woolgrowers through Australian Wool Innovation.
References
- 1.Balic A, Bowles VM, Meeusen ENT. The immunobiology of gastrointestinal nematode infections in ruminants. Advances in Parasitology. 2000;45:181–241. doi: 10.1016/s0065-308x(00)45005-0. [DOI] [PubMed] [Google Scholar]
- 2.Williams AR, Karlsson LJE, Palmer DG, et al. Relationships between faecal dry matter, worm burdens and inflammatory mediators and cells in parasite-resistant Merino rams. Veterinary Parasitology. 2010;171(3-4):263–272. doi: 10.1016/j.vetpar.2010.03.031. [DOI] [PubMed] [Google Scholar]
- 3.Kaplan AP, Joseph K. The bradykinin-forming cascade and its role in hereditary angioedema. Annals of Allergy, Asthma and Immunology. 2010;104(3):193–204. doi: 10.1016/j.anai.2010.01.007. [DOI] [PubMed] [Google Scholar]
- 4.Bertram CM, Misso NL, Fogel-Petrovic M, et al. Expression of kinin receptors on eosinophils: comparison of asthmatic patients and healthy subjects. Journal of Leukocyte Biology. 2009;85(3):544–552. doi: 10.1189/jlb.0508283. [DOI] [PubMed] [Google Scholar]
- 5.Karlsson LJE, Greeff JC. Selection response in fecal worm egg counts in the Rylington Merino parasite resistant flock. Australian Journal of Experimental Agriculture. 2006;46(6-7):809–811. [Google Scholar]
- 6.Williams AR, Palmer DG, Williams IH, Vercoe PE, Emery DL, Karlsson LJE. Relationships between immune indicators of parasitic gastroenteritis, nematode burdens and faecal dry matter in sheep. Animal Production Science. 2010;50(3):219–227. [Google Scholar]
- 7.Gray GD, Barger IA, Le Jambre LF, Douch PGC. Parasitological and immunological responses of genetically resistant Merino sheep on pastures contaminated with parasitic nematodes. International Journal for Parasitology. 1992;22(4):417–425. doi: 10.1016/0020-7519(92)90142-8. [DOI] [PubMed] [Google Scholar]
- 8.Douch PGC, Morum PE, Rabel B. Secretion of anti-parasite substances and leukotrienes from ovine gastrointestinal tissues and isolated mucosal mast cells. International Journal for Parasitology. 1996;26(2):205–211. doi: 10.1016/0020-7519(95)00120-4. [DOI] [PubMed] [Google Scholar]
- 9.Oschatz C, Lecher B, Jansen T, et al. Mast cells increase vascular permeability by heparin-initiated bradykinin formation in vivo. Immunity. 2011;34(2):258–268. doi: 10.1016/j.immuni.2011.02.008. [DOI] [PubMed] [Google Scholar]
- 10.Stear MJ, Strain S, Bishop SC. Mechanisms underlying resistance to nematode infection. International Journal for Parasitology. 1999;29(1):51–56. doi: 10.1016/s0020-7519(98)00179-9. [DOI] [PubMed] [Google Scholar]