

Abstract
Compartmentalized redox faults are common to ageing diseases. Dietary constituents are catabolized to NAD(H) donating electrons producing proton-based bioenergy in coevolved, cross-species and cross-organ networks. Nicotinamide and NAD deficiency from poor diet or high expenditure causes pellagra, an ageing and dementing disorder with lost robustness to infection and stress. Nicotinamide and stress induce Nicotinamide-N-methyltransferase (NNMT) improving choline retention but consume methyl groups. High NNMT activity is linked to Parkinson's, cancers, and diseases of affluence. Optimising nicotinamide and choline/methyl group availability is important for brain development and increased during our evolution raising metabolic and methylome ceilings through dietary/metabolic symbiotic means but strict energy constraints remain and life-history tradeoffs are the rule. An optimal energy, NAD and methyl group supply, avoiding hypo and hyper-vitaminoses nicotinamide and choline, is important to healthy ageing and avoids utilising double-edged symbionts or uncontrolled autophagy or reversions to fermentation reactions in inflammatory and cancerous tissue that all redistribute NAD(P)(H), but incur high allostatic costs.
1. Introduction
“Progress engenders Leisure whereby Energy is directed toward advancement of the Mind in all parts of Society that in turn receives new Energy.” Turgot, 1750.
“In my theory there is no absolute tendency to Progress, except from favourable Circumstances” Darwin, 1838.
Mental and neurological disorders are poorly explained yet constitute a good proportion of the global burden of disease, now defined as the inability to adapt homeostatically in the face of social, physical, and emotional challenges [1–3]. Robust brain development in the first place must help and is defined by environmental factors and cell types that evolve in a series of networks to ensure the efficient flow of energy and information [4–6]. Energy supplies from food are a prerequisite for producing the necessary variety of cells and simultaneously act as a major selection force influencing their survival; yet despite its importance to brain evolution and development inequalities of diet, particularly over meat and calorific intake, remain a global issue, as do stress responses such as overeating [7–11]. Bidirectional circuits, from the energy supply to terminal fields, are selected from early exuberant developmental (but evolutionarily constrained) fields by environmental exposures but must have weak links as strong safety factors against every eventuality whether environmental, genetic or stochastic would be too expensive in energy terms. At risk circuits may include cholinergic, serotoninergic, and dopaminergic systems that have not scaled up as fast as the overall three-fold expansion of the brain in the human primate [12–20].
Redox energetic faults and aberrant mitochondrial dynamics with consequent oxidative stress, and loss of calcium, glutamate, proteosome, and inflammasome homeostasis are important proximate mechanisms of acute and chronic diseases and the physiological declines that are linked to ageing [21–31]. Some of these disease circuits may have particularly high energy requirements, for instance, those affecting higher intellectual and complex physical exploratory or reward functions using cholinergic or dopaminergic neurons: these for instance have mechanisms of autonomous pace-making with a high metabolic energy cost in terms of ATP to maintain tight control of intracellular calcium and complex redox needs involving cofactors, metals, and melanin [21, 32]. Confirmation of this fundamental and compartmentalized failure of the energy supply, often with impaired autophagy and mitophagy, comes from study of acute brain injury from hypoxia or hypoglycaemia or direct trauma, mitochondrial mutations linked with chronic diseases of ageing such as Alzheimer's (AD), Parkinson's (PD) and Huntington's Disease, and, Complex 1 toxins (such as 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)) or rotenone and physical tests of endurance such as polar and high altitude expeditions [33–40]. The root cause of these diseases may be exposed energy and informatic links and responses with inadequate redox and mitochondrial potentials affecting proton based power and available ATP. Levels of cellular ATP affect all processes from muscle contraction to repolarization of neuronal membranes, to synthesis of cellular building blocks, to protection against microbes and toxins, and maintenance of proteins, RNA, DNA, and chromatin [41–44].
Now is an appropriate time to look at the effects of energy and the interlinked nicotinamide and methyl-group under and oversupply and their relationships to information transfer and work outputs including those done to improve personal niche environments and evolutionary processes [45, 46]. This survey includes a historical episode when the nicotinamide, and tryptophan and hence NAD(P) supply (and potentially over 400 dependent reactions that influence every area of metabolism) failed as did the supply of methyl-groups. The forgotten case of the complex socioeconomic disease, Pellagra, may be a lesson about losing energy and redox “logic” at several hierarchical levels, and we intend to show that there may be current scope to adjust the dose of the macro- and micronutrients involved, across and within populations and according to individual need, aiming to improve robustness and resilience to a range of degenerative and proliferative diseases [47–49].
Background (1): Ageing Diseases, Evolution, and NADH —
Current theories on ageing, lifetime behaviour patterns, reproductive cycles, intergenerational transfer of resource and stress responses invoke energy sources, sensors, regulators and tradeoffs: as do wider theories on the astonishingly rapid metabolic evolution of big brains aided by the ecological, social, and cultural strategies used by man that affect both access to energy useful information and increase in net energy input and allocation of energy resources [50–58]. Thermodynamically open and phylogenomic models support the idea that NADH and nicotinamide adenine dinucleotide phosphate (reduced) (NADPH) have had a central role and are the common currency in linking circadian environments to mitochondrial metabolism and vice versa [59–66] (Figure 1). Sharing hydrogen and its carrier around equitably as the source of electrons is complex relative to the supply of the electron acceptor oxygen that is free, but when successful enables efficient energy flows to be achieved: there is some danger from free radical damage from oxygen metabolism, though a prime importance of this for ageing is now in some dispute with the spotlight now on NAD and nicotinamide [67]. Efficient energy conservation in turn allowed the evolution of complex ecosystems and the expansion of genetic and species diversity, including the development of socially interacting brains working vertically down, up, and across generations often through NAD-dependent neuroendocrine and neurotransmitter mechanisms [68–76].
Recent reviews on nicotinamide and NAD metabolism, evolution and function concentrate on its pivotal role in gene silencing, development and ageing, through recently appreciated NAD consumers, such as the sirtuins (SIRTs) [77–85], that evolved to work in large part as nutrient-sensing regulators, and poly (ADP ribose) polymerases (PARPs) [86, 87] and on the role of NAD(H) in Wallerian degeneration and neuronal death after a variety of insults [88, 89] (Figure 2). These reviews do not do justice to the importance of nutritional and symbiotic interfaces or the developmental dynamics of inducing NNMT, which partly regulates NAD(H) via controlling and detoxifying the supply of nicotinamide (vitamin B3), at the cost of losing S-adenosyl-methionine (SAM), the only metabolic methyl donor, or the related role of evolutionary history as the ultimate basis for understanding the proximate pathophysiology of a range of modern diseases [90–96].
Figure 1.
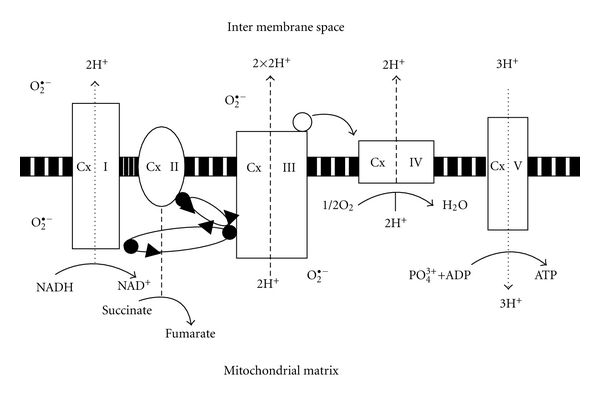
NAD(H)—the electron donor to complex I translocating protons across the membrane by a “steam engine” like mechanism producing ATP: the most important function of mitochondria alongside proton leaks for heat and energy dissipation and signals for autophagy and apoptosis. Many mutations that affect mitochondrial complex 1, or, microtubular function and kinases and autophagy/mitophagy and radical production contribute to rare forms of ageing diseases such as PD in a vicious cycle subverting normal quality controls that affect proteosomal homeostasis. In the case of Parkinsons', increasing NADH levels may drive excessive dopamine synthesis to toxic levels by enhancing the recycling of the cofactor for tyrosine hydroxylase.
Figure 2.
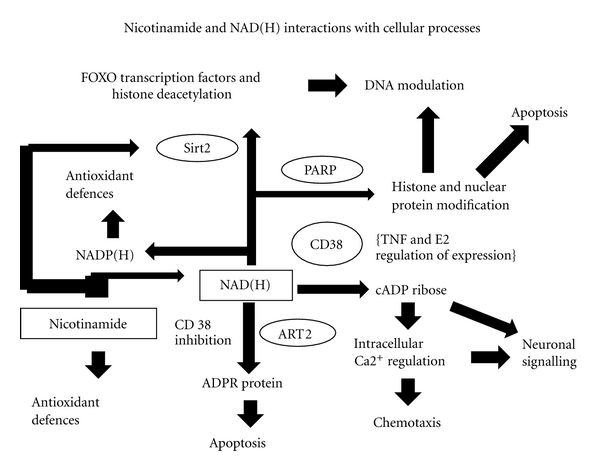
Direct redox regulation apart NAD-consuming pathways influences a prodigious number of pathways involved in internal metabolism and connections with the external world. Survival at the cellular level and of viable interactions with each other and diet and symbionts means that NAD and Nicotinamide are at the hub of the survival of superorganisms such as ourselves. Key: FoxO—forkhead transcription factors; Sirt2 and PARP in text; CD38 cluster differentiation 38; TNF—tumor necrosis factor; E2 Prostaglandin E2; cADP—cyclic adenosine di-phosphate; ART2—T-cell ADPribosyltransferase; ADPR protein—adenosine di-phosphate ribose protein.
Background (2): Symbiotic Energy Environments —
Flow of energy largely determines the structure of ecosystems via food webs and taste sensors with appetite and quality controls, that avoid toxins but allows self-medication, and involve ATP sensitive autophagy in hypothalamic neurones [97–99]. Diet with concurrent domestication of microbes as gut symbionts (or yeasts to ferment foodstuffs) that break down proteins or indigestible starches or manufacture vitamins, including nicotinamide, has evolved to supply and translocate energy and information via circadian pulses of NAD(H): particular symbionts, for example, buffer particular diets such as those low in meat and dairy consumption or high in plant based carbohydrates [100, 101]. Disturbance of these symbiotic relationships, such as by the over-use of antibiotics, is receiving considerable attention in several conditions that are increasing in incidence currently [102]. Swopping production of vitamins and some catabolic enzymes from personal metabolism to diet or symbionts appears to help find hydrogen-based energy [103]. Diets (that both depend on and have shaped cultural niche constructions, such as farming selected crops, domestication of animals for milk and meat, and cooking) and the entangled symbionts which form the microbiome in the gut are essential synergies for the evolution of human life and energy harvesting directly through NADH/ATP production [104, 105]. Diet and symbionts compensate for each other by acting as coupled recycling systems at many metabolic, immunologic, and behavioural levels, and both are implicated when energy balances go wrong as in the case of obesity, and when both are insufficient, gut cells may even break down their own components to obtain energy [106–111].
Failure of the supply of NAD(H) as a consequence of malnutrition or starvation triggers other attempts to compensate by autocarnivory to avoid apoptosis or necrotic cell death, particularly important for neurones and other cells that are rarely renewed after birth [112, 113]. Immune/inflammasome modulation with such malnutrition also occurs and allows the organism to tolerate microorganisms which are potentially deleterious but are also capable of supplying NAD(H) or its precursors [114]. Reversion to fermentation occurs in inflamed or cancerous tissue, favouring net NADH export (the Warburg phenomenon) and interacts with autophagic mechanisms with recruitment of somatic oncogene mutations and methyl-group sensitive epimutations to achieve this metabolic milieu [115–119]. All the above may be homeostatic short-term attempts to correct a poor local energy/redox environment whether by producing, redistributing, or wasting energy to heat. As another redistributive example, ectopic fat stores, which include uncoupling proton-wasting inflammatory sites with loss of mitochondrial potential, are part of the metabolic syndrome to which populations that have recently undergone a nutrition transition are especially prone [120–123].
2. Pellagra
An NAD(H) deficiency state that illustrates the complexity of these coevolved energy networks was observed during the largely forgotten epidemics of pellagra caused by economically determined suboptimal foraging of nicotinamide and tryptophan (tryptophan is converted to NAD and serotonin) and dietary sources of methyl-groups. Such a dietary deficiency may still be important in sections of modern populations and intracellular and microenvironmental deficiency states can also occur from high NAD expenditure with NAD(H) ratios also affected by any disorder that causes hypoxia or hypoglycemia [124, 125].
2.1. Clinical Features of Pellagra
Scientific detectives investigating ageing diseases and human evolution should revisit a fork in the intellectual road that happened exactly a century ago for real-life ecological rather than solely laboratory-based clues around the NADH-NAD consumer axis (that are often controversial due to confounding factors in experimental design that try too hard to separate physiological homeostasis and stress responses from ageing or are too genecentric given that most genes in some ecological context or other have survival value) [126–141]. Then the last major epidemic in man occurred (pellagra also occurs in several other carnivorous species such as dogs) with a systems failure of NAD(H) and therefore thermodynamic, NAD-Hub, and methylome homeostasis at all hierarchical levels. The epidemic started and ended socioculturally. Hypovitaminosis B3 caused a premature ageing condition with loss of robustness to chemical, physical, and emotional stress, leading to parkinsonism, dementia, metabolic disease, and cancer. It was conquered by dietary supplementation. Famously, pellagra caused dementia, a characteristic photosensitive dermatitis (Casal's necklace), diarrhea, and death. The cause was real world economics creating NAD(H) gradients across poor populations, and across sexes (as it was twice as common in women) in the cotton- and corn-dependent states of the southeastern USA that reached epidemic proportions. Too much corn and molasses and too little meat led to a diet deficient in nicotinamide and probably other vitamins such as riboflavin and choline but often adequate or even high in calories. Currently a similar situation exists in parts of Africa. In the USA, familial aggregations were common, suggesting genetic predisposing factors and widespread epigenetic reprogramming. A diet with too much corn and too little meat caused deficiencies of nicotinamide and the essential amino acid tryptophan, though deficiencies in other vitamins, such as choline, and other methyl donors contributed to the malaise. Mental symptoms were wider than dementia, in that depression, fatigue, psychomotor retardation, mania, obsessions, and a whole range of psychoses with auditory and visual hallucinations were well described, along with personality change and sociopathic and drug and alcohol addictive behaviours. Panic disorders were seen as was a general inability to deal with physical or mental stress. Poor brain development such as hydrocephalus or cerebral palsy was also common. Acute delirium or even coma occurred, with some patients having myoclonus and other extrapyramidal signs reminiscent of the spongiform encephalopathies. The dementias of pellagra included features akin to Lewy body, Alzheimer's, frontotemporal, vascular, and prion diseases. Parkinsonism was also common and a festinant gait was first described in pellagrins. Tremors of various descriptions, including asymmetric rest tremors, were noted and some patients had typical paralysis agitans. Pellagrins had a characteristic expressionless facies, so some signs of parkinsonism were present in most cases. Many features of pellagra closely resemble the nonmotor aspects of PD.
The neurological manifestation did not stop there because other degenerative conditions, such as an amyotrophic lateral sclerosis-like picture, were described, with fasciculation of the tongue and upper and lower motor neuron signs. Cerebellar syndromes occurred and vertigo was frequent. Headaches, sensory and pain syndromes, epilepsy, and involuntary movements were noted as well as sleep disturbances. Cord lesions were also seen, as was optic atrophy, so there were multiple sclerosis (MS), like variants.
Non-neurological effects included the diagnostic photo-sensitive and ageing dermatitis along with diarrhea from multiple gut infections. Other infections including tuberculosis (TB) were common, suggesting a widespread effect on the immune system. There were also profound metabolic effects on the endocrine and thermoregulatory systems. For instance, endocrine dysfunction in thyroid and cortisol and reproductive pathways were all clearly described. Cancer rates were increased but masked by a high premature death rate.
Pellagra without dermatitis was well accepted and a frequently overlooked diagnosis. Young men from affected states had inordinate difficulty in passing the intellectual tests that involved reading, writing and numeracy skills and the physical fitness tests required by the military, suggesting that subclinical disease was rife. Indeed poor diet and health have been implicated in the Southern states both having a tradition of violence (particularly toward the stealing of nicotinamide-rich livestock) and losing the American Civil War [142].
The classic pathological sign was chromatolysis of the high energy requiring pyramidal cells in the cortical gray matter but Purkinje cells also degenerated and there was a widespread cell loss in basal and sympathetic ganglia. Demyelination was commonly seen in the spinal cord, particularly in the posterior and lateral columns. Much was made of pigmentary change, glial overgrowth, vascular change, amyloid deposition, cytoskeletal and mitochondrial abnormalities and signs of inflammation and atrophy in all organs. A systems failure with the degenerative pathology of precocious senility was proposed by several pathologists in the late 19th century, and clinicians commented on patients as “wrecks of humanity”.
Pellagra is a wide systems failure in that an ecologic dietary stress triggers developmental and degenerative effects affecting behaviour and a vicious cycle which amplified the original economic stress. In addition, healthy members of society were phobic about pellagrins and showed them little if any empathy (unlike most other disabilities) and preferred explanations, such as a poor gene pool or that they were infectious, rather than blaming poverty. This picture, we suggest, may represent some aspects of human evolution gone wrong given that not only individuals but also their social relationships were all “wrecked”.
2.2. Is Subclinical Pellagra Relevant to Contemporary Disease?
Many poor individuals and groups, even in rich communities, have low amounts of vitamins in their diet. This assumes that the recommended daily allowance (RDA) for vitamins is correct for good health, as opposed to simply avoiding deficiency states. Consequently, subpellagrous nicotinamide deficiency may have lifetime roles in a range of behavioural traits, neuropsychiatric diseases, and dementias. Dietary choline deficiency exacerbates nicotinamide deficiency because metabolism of the two agents is linked; N-methylnicotinamide blocks choline export [143, 144]. Currently, trials of nicotinamide are underway in AD, stroke and several other neurological conditions based on the following [144–182].
The brain uses a lot of energy proportionate to its weight and glucose and oxygen status and NADH modulates memory retrieval.
In prospective studies, diets of AD patients were found to be deficient in nicotinamide premorbidly.
Nicotinamide reduces the accumulation of amyloid by an effect on secretases and affects the toxic effects of amyloid on astrocytes by inhibiting PARP and the NAD synthase nicotinamide mononucleotide adenylyl transferase (NMNAT) that has protein-protein chaperone activity and also affects Tau-induced neurodegeneration by promoting clearance of hyperphosphorylated Tau oligomers.
In transgenic mice, nicotinamide restores cognitive deficits with effects on both dendrites and axons and reduces a phosphospecies of tau which is linked to microtubule depolymerisation, an effect also seen following the inhibition of SIRT1, and increases acetylated alpha-tubulin which is linked to microtubule and tau stability.
In models of AD, overexpression of SIRT1 or PARPs reduces memory loss and neurodegeneration.
Loss of SIRT1 is associated with the accumulation of amyloid-beta and hyperphosphorylated tau in the cortex of AD brains (thought to be compensatory changes to restore glucose/ketone metabolism).
Recovery of learning and memory has been associated with chromatin remodeling that happens with agents that affect SIRT and methyl metabolism and epigenetic mechanisms seem to be important in AD.
Energy paths involving cAMP and insulin-like growth factors are involved in memory consolidation and enhancement.
Dementia is being linked with both being under and over weight stress and exercise levels and the metabolic syndrome.
Nicotinamide reduces ischaemic and traumatic brain damage directly and via enhancement of the synthesis of NAD: both are common additional factors in many forms of dementia.
Redox active methylene blue affects dementia pathology and its clinical manifestations.
SIRT I regulates Notch signaling that is involved with mechanisms of action of mutations in the PSEN1 gene and the proliferation of neural progenitors in the subventricular zone important in the development and evolution of large cortical mantles and essential for cell specification and tissue patterning (and is involved in human cancer).
Nicotinamide and SIRTs are involved with promoting DNA repair particularly in oxidative stress circumstances.
Poor diet and chronic diarrhea affecting brain function may also interact with tradeoffs at genetic and antagonistic pleiotropic levels. Two copies of the e4 allele of apolipoprotein E lower the incidence of childhood diarrhea and poor cognitive development but increase the risk of AD later and appear to be within the reach of natural selection as do other genes affecting human longevity.
Lysosomal proteolysis and autophagy require presenilin 1 and are disrupted by Alzheimer mutations and PD-related mutations.
The neurology of HIV/AIDS, which includes premature ageing, dementia, and parkinsonism has clinical, pathological and biochemical similarities with pellagra [183]: many at-risk groups are nicotinamide deficient given that monophagic corn-based diets are now common in Africa. Both AIDS sufferers and pellagrins get many parasitic infections which may be dangerous homeostatic responses that boost NAD(H) in the host when dietary sources of energy are poor. Tuberculosis, for instance, excretes nicotinamide, is inhibited by nicotinamide and related compounds such as Isoniazid and treatment can precipitate pellagra and many gut infections/parasites break down otherwise indigestible cellulose, ultimately producing NAD(H) [184]. The HIV disease marker CD38 is a NADase that regulates intracellular NAD(H), implying disturbed nicotinamide-NAD(H) metabolism [185]. SIRTs regulate the HIV transactivator Tat, promoting viral transcription and, through the NF-kappaB complex, induce T-cell activation particularly in dendritic cells and there are other links between NAD and the immune system [186, 187]. In view of the inhibitory effect of nicotinamide on SIRT activation and other actions, nicotinamide ought to be protective in AIDS and other chronic infections such as TB (that were particularly common in pellagrins) as an important missing host factor that affects virulence acting much like a vaccine [188]. Interaction between symbionts, such as the tuberculosis bacillus, happens by modulation of immune responses via tryptophan-NAD and mTOR pathways in dendritic and Th17 cells that have been under significant evolutionary pressures and span immune and energy pathways, even using ATP as a danger signal [189–193]. The tryptophan oxidation pathway is particularly interesting in the current context as it is both the de novo synthetic pathway for NAD (nicotinamide is like choline a semivitamin) and the tolerogenic pathway both for microbes and cancer and contains some potential neurotoxins and is disturbed in many degenerative diseases: enough NAD from vitamin B3 sources means less toleration of symbionts and at the extreme immune intolerance and allergic disorders [194, 195].
3. Hypervitaminosis B3 and Choline
Macronutrients and micronutrients, which include nicotinamide, choline, and other methyl sources, may all obey Bertrand's rule in that they have an inverted “U” shaped dose-response curve with toxicity at either extreme, as was suggested by Waaler regarding increased mortality at either extreme of weight/height ratio curves [196]. Affluent populations are exposed to diets with a historically high calorific content from refined sugars and fats relative to energy expenditure. In these populations, energy expenditure is relatively low because of limited exercise and the fact that body heat is maintained through living in artificial environments supported by the use of fossil fuels. Damage is often assumed to occur from the effects of an obesogenic environment rather than the more direct effect of disturbed NAD+ : NADH ratios on a variety of dependent insulin, cortisol and other pathways (also affected by other bad habits such as smoking and alcohol and stress) with obesity being an attempt to sequester energy and actively metabolise nicotinamide (more in keeping with its biological buffering role against starvation, even if it comes at a price) [197–199]. The relationship of the dose of the hydrogen source (caloric intake) with the dose of the suppliers of the carrier (NAD), derived ultimately from nicotinamide and tryptophan, that must be important for high quality biologically useful energy, has never been investigated in detail. Recently the possibility of both too little and too much choline in diet, modified by gut flora and genetic factors, has been raised in the context of vascular disease [200–202].
Extra choline and other methyl-group sources in diet from vegetables, meat, milk, and eggs during our evolution just like other key molecules such as nicotinamide would have enabled the development of cholinergic, glutaminergic, and monoaminergic systems that have actively evolved in primates and through, for instance, the NADH-dependent recycling of an essential cofactor, tetrahydrobiopterin, remain dependent on a good NADH supply for adequate learning and social and reward behaviour involved in sustenance and in desiring advance information, often mediated by dopamine [203–207]. Oxytocin/Vasopressin pathways are also important for the social interactions that have been so important to our evolution as are social substances such as alcohol which are also NAD-dependent, but all these systems may leave us prone to overindulgence as circumstances have improved [208–211].
4. An Evolutionary Perspective: Nutritional and Cognitive Transitions
An evolutionary perspective is warranted, because ageing of the cerebral cortex and basal ganglia is more prominent in humans than other primates, who also do not get AD or PD naturally (even though toxins such as MPTP produce a PD-like phenotype in the laboratory) and as living to old ages appears to have arrived late in our evolution and the ageing process may even finally stop [212–215]. High exposure to nicotinamide and choline and other dietary changes that boosted the overall supply of NAD(P)H may have been important ingredients of the change in diet of primates from a grass eating or herbivorous and fruit eating one toward one containing more starch from tubers and vegetables on to meat/marrow/brain eating. This triggered the 3-fold growth over 3 million years of the modern human brain and a high degree of control over the energy environment [19, 210–220].
This changed diet towards high quality energy (not just extra calories from “junk” food, because it combines high calorific content with key vitamins such as nicotinamide and several methyl donors (choline/folate/B12) and essential amino acids such as tryptophan and phenylalanine and lipid components such as Omega-3 fatty acids from fish [221, 222]). This change first happened in NAD(H) source rich patches of the less arid parts of Africa with repeated genetic and informatic bottle-necking as our species followed energy trails out of Africa and was facilitated by the original invention of tools for butchery and later by hunting in groups and by the sharing and storing of meat [221]. Significant brain power is necessary for these group behaviours where individuals specialize and complement each other and read and anticipate minds of other hunters and prey and animal assistants with special sensory skills, and, learn to avoid toxic fauna and find natural medication now in a more seasonal and unpredictable environment. This brain power enables future-orientated thinking and delayed reward discounting, so that habitats or relationships were enhanced and not destroyed for short-term gain [223–227].
Preadaptations such as endothermy and earlier modifications of aerobic mitochondrial function enabling proton leaks and an ample blood flow to the brain to keep it cool for diurnal and persistent hunting may have helped [228]. Hominids, from Homo Erectus on, clearly went for acquiring multiple high energy sources. This enabled the resultant evolutionary advances via a positive feedback loop in which increasing energy sources that were easier to chew allowed the development of larger brains (but small teeth, masseters and guts), which in turn increased needs, which led to the search for more and greater energy sources as niche construction and included the use of exosomatic energy. However, our brain size may have peaked and it is smaller than the more carnivorous Neanderthals. In several species, domestication can lead to striking reductions in both cortical and striatal volumes surprisingly quickly, showing that this is a fluid process [229]. The invention of farming of grain and particularly meat and milk suppliers, followed by corresponding nutrigenomic innovations, that facilitated adaptations to new NADH-rich environmental opportunity, affecting lactose (milk is a valuable source of calories, pathogen-free fluid, and nicotinamide) and alcohol tolerance, amylase activity and nicotinamide methylation, happened on multiple occasions and spread fast [227–230]. This suggests very strong selection pressures working through both cultural and genetic inheritance, as does the rapid evolution and diversity of genes involved with mitochondrial function and glycolytic pathways [231]. This pressure seems to be every bit as important as mutational changes in genes directly affecting brain function [232–235].
Bipedal man is thought to have adapted fast, allowing time and energy to be freed up along with sedentism that in itself reduced energy expenditure from personal and child transport and by having a roof over their heads [236]. The resultant leisure led to the advancement of interactive minds and a distributed intelligence through multigenerational and multidirectional social structures with critical mass—important for divisions of labour—and a series of information revolutions that continue to this day (the “grandmother hypothesis” favours informatic and energy transfer but grandchildren may equally facilitate vertical transfers of certain resources such as new language or technological skills and have been an underrated factor in social cohesion and longevity).
Along with the social transmission of food preferences often mediated by acetylcholine release, which reduced the fear of novel foods, these information revolutions improved decisions over the trading of and even inheritance patterns of energy sources and other materials [237, 238]. Intolerance of a restricted diet based on “fall-back” foods may also explain our love of flavour and the difficulty of finding single foods that are particularly protective against diseases rather than well-balanced omnivorous mixtures. The preference for a broad diet and the danger inherent in monophagy are evidenced by the many phenotypes seen with pellagra [239].
All in turn allowed advanced niche construction and the development of an energy-rich environment, which was enhanced by cooking that releases bioavailable nicotinamide unlike some other vitamins that are destroyed by cooking. Combined with other uses of fire, this enabled a more ecologically diverse and productive environment, such as increasing the number of available herbivores, so that less energy was expended on hunting but with greater certainty of success. Early man, through some degree of social independence and a consciousness, could move to brand new pastures or construct pastures new through planning, imagination, and innovation [240, 241].
5. More Recent History: The Neolithic and the Columbian Exchange
The history of maize, the crop that triggered pellagra, is a good example of coevolution because it is now dependent upon us for its sexual reproduction and we are very dependent on it as an energy source. The initial cultural spread of maize is one of the best examples of diffusion of innovation, which sometimes went wrong from lack of attention to detail. When it was imported to the southeastern states around 2000BC and later Europe from central Mexico as part of the “Columbian exchange”, it was improperly cooked and not combined with beans and squash or adequate meat. Maize is popular as it is a C4 plant with very efficient photosynthesis particularly when water is at a premium. New breeds underpin a large section of modern agribusiness. Together with other technoinnovations they have raised production from 1 Kg per hour on a Kenyan farm to 1000 Kg per hour on farms in Iowa [242].
Maintaining and enhancing the supply of high quality components, such as meat and milk, during the original agricultural revolution/Neolithic Demographic transition and over recent centuries has not always been easy and has caused tradeoffs between high population growth and health [243–246]. At times, micronutrient shortage such as lack of nicotinamide would have been more of a problem than shortage of calories and may be a mirror image of the much more recent contemporary demographic transition in Western industrialized societies. Population growth may have had a large part to play in the chronic malnutrition that has stunted growth and led populations to be prone to acute infection (which stresses the NADH axis); it has coincided with the coevolution of chronic infections (e.g., TB that excretes nicotinamide) and malaria (that affects NADH status, as do malaria resistance genes such as those causing G-6PD deficiency).
6. Affluence
A doubling of life span has occurred in many economies over the last 200 years, largely from the reduction in what were then common infectious diseases. Better diet has increased resistance to acute infections that compete for NAD resources as do their toxins. In the case of the so-called “infectious chronic unconquerables”, such as malaria and TB, there appears to be an immunological stand-off in which a degree of tolerance may have been traded for an increased supply of an essential nutrient. Many apparently healthy carriers, which were present in the dietary deficient population, literally disappeared as dietary standards improved in Europe and the USA. Thus, with the increasing quality of diet, symbionts become no longer necessary to supplement diet and the immune system can revert from controlling the population size of a mutualist parasite to trying to kill every invader. Recent information on the role of NAD modulating innate immunity suggests complex energy-centred interactions between neuronal and immunological systems. Contextually useful organisms (as seen in cases of pellagra) depend on ecological factors such as diet rather than geography or phylogenetic considerations, and on a good diet with changed prebiotic composition may no longer be advantageous. With the improved western diet, the immunological reaction to the formerly “useful” and actively welcomed biotic antigens concerned may be “confused” and may extend through molecular mimicry and nutrition-related signals to other antigens, thus setting off abnormal inflammatory and autoimmune reactions including to foods and pollens (that contain NAD(P)H oxidases). This is a different explanation to the more restricted hygiene hypothesis where lack of exposure rather than actively shunning “old friends” is postulated to cause immunological hyperreactivity [247–251].
Here we suggest the possibility that a hypervitaminosis B3 state, with both nicotinamide and N-methylnicotinamide having an optimal range, resulting from a western diet overrich in meat and vitamin supplements working alongside calorific excesses may be implicated in other modern disease phenomena. Meat eating is preferred where it is available, though during pregnancy it is a common dietary aversion, suggesting that it can be toxic in early development as does the presence of the detoxification enzyme, NNMT and evidence of acute toxicity with nicotinamide overdose and worries about red meat being a risk factor for some cancers [252–254]. Nicotinamide exposure can be high, in excess of five times the recommended daily amount (15 mg/day) where diet has both cereals and “high energy” drinks supplemented with the vitamin and where there is an abundance of cheap meat and milk. This chronic overload can be envisioned as working through unbalancing NAD(H)-dependent systems, including SIRTs and PARPs or dehydrogenase pathways (for instance in cortisol metabolism), and through the direct effects of nicotinamide or N-methylated metabolites. NAD+ is the substrate for SIRTs and nicotinamide is an inhibitor, so dietary levels of NAD(H) precursors could act as both positive and negative influences on such systems. In the case of nicotinamide, dietary inputs are modulated in part by genetic determination of NNMT levels, with 24% of the population being high expressors. Nongenetic factors also modulate expression of the enzyme. These include nicotinamide itself, stress and the demands imposed on NAD(H) availability by growth, tissue repair and exercise. Interestingly, NNMT and other components of NAD(H) pathways are markedly induced in a variety of currently common cancers as well as in the metabolic syndrome, obesity, PD and autism [255–262].
An animal model of toxicity from excess dietary nicotinamide has been reported. In this model, dopaminergic neurons were damaged and locomotor activity was reduced. In addition, this model responded to L-DOPA [263]. In other models, nicotinamide and novel derivatives such as N-Nicotinoyl dopamine can reduce melanin as happens in PD [264, 265].
One consequence of NNMT induction and other methyl-transferase reactions is a depletion of methyl stores and S-adenosylmethionine (SAM) that is ultimately dependent on an adequate dietary supply from expensive vegetables and meat rather than cheaper cereals such as corn (banned in 19th century France as unfit for human consumption, thus eliminating pellagra at a stroke: a lesson for modern Africa) [266]. Depletion of available methyl groups, whether from a poor diet or one with excessive nicotinamide, may be a common cause of epigenetic phenomena given the marked influence methylation has on gene expression in pluripotent and differentiated cells that maintain cellular identity [265]. The methylome is an important mediator of information and modifies gene transcription and translation including riboswitches affecting RNA metabolism and in toxicology pathways in concert with NAD and ATP and all appears to be involved in a number of diseases seen within the pellagra phenotype such as cancer and motor neurone disease [267–272]. A major role for the methylome is well accepted for many cancers and their related epimutations sometimes working in concert with DNA sequence mutations in tumour suppressor and DNA repair genes and may well be modifiable by diet [273–277]. There may be several mechanisms whereby such metabolic programming can be inherited and affect offspring [278].
7. A Deep History of Hydrogen and Hydrogen Symbioses: Origin of Cooperative Species [279–281]
Life without hydrogen is impossible, whether free or bound with NAD(P) as energy or with oxygen as a solvent. The sun burns hydrogen in a fusion reaction and emits light energy. Early life chemo-litho-autotrophs used redox gradients including hydrogen as the electron donor and then the earliest animals developed symbiotic relationships with bacteria that produced hydrogen and the invention of photosynthesis split water to produce hydrogen as NADPH and oxygen (earlier forms split H2S). Hydrogen can escape from the earth and led to the great oxidation events and an oxygen atmosphere that along with important symbiotic acquisitions involving chloroplasts, hydrogenosomes, and mitochondria allowed the evolution of complex plants and the rise of an animal kingdom that eats hydrogen sources or has gut symbionts that produce hydrogen equivalents from otherwise indigestible foods. Too much hydrogen escape from splitting of water by radiation in the upper atmosphere creates “runaway greenhouse” effects that are incompatible with life and may have happened on other planets and could happen here. Early mammalian inventions that would increase hydrogen and nicotinamide sources include placental and milk feeding. Healthy energy ecosystems reduce poverty and underpin healthy interactive economies: and even though modern economies appear to move away from a primary reliance on natural capital and capturing solar energy, we all need to eat well to stay healthy and need to appreciate the real source of our prosperity and the need to avoid food crises worldwide.
8. Longevity and Nutritional Balance
The NAD(H) energy path and NAD consuming hubs are involved in many key cellular processes and in the regulation of food intake and the phenomenon of caloric restriction and hormesis (that which does not kill you, whether diet, toxin, emotional, acute infectious or traumatic stress, makes you stronger) which are implicated in many diseases of ageing [282–302]. These hubs are amenable to therapeutic intervention and may unwittingly have contributed by a better overall diet and energy budgets to recent increases in longevity in affluent countries (Figure 2). Age-related changes in NAD/NADH ratios have been noted and a world where NAD is central postulated [303, 304]. Placebo effects that are so prominent in man also have been linked to energy pathways and are particularly effective when the energy future is perceived as healthy, allowing energy to be invested in healing rather than fire fighting [305]. Endocannaboids that depend upon the organisms energy status may translate food availability into fundamental choices about development which affect lifespan, and some addictions that were common in pellagrins such as Nicotine are also closely linked to energy pathways. Caloric restriction hints at the need for a nutritional stoichiometry to balance energy demand and supply throughout life. It is associated with telomeric stability and increased SIRT 1 activity stabilising PGC-1 and results in increased mitochondrial biogenesis, metabolic function, and the deacetylation and inactivation p53. This balance may be more about NAD+ : NADH ratios than calories per se, which may explain the perplexing and contradictory role of NAD-consumer agonists (such as resveratrol) and antagonists in disease models. SIRT1 knockouts show widespread p53 activation and shortened life expectancy [306]. Increased longevity as a result of caloric restriction and related genetic models fits with abundant evidence that nicotinamide is involved in neuroprotection [307–318]. The damaging effects of too low a dose are a matter of historical record. Evidence is emerging on excessive intake of the vitamin [319, 320]. Nicotinamide's actions may be double-edged as although it is efficacious in a wide range of current insults and pathologies too much may be as harmful as too little.
Links between nicotinamide, acetyl and methyl metabolism and epigenetic phenomena are strong [319, 320]. Other recently studied famines such as the Dutch hunger winter of 1944-45 and the Chinese famine of 1958–61 had marked long-term and transgenerational effects on health such as the risk of the metabolic syndrome but also psychiatric and antisocial behaviours working through epigenetic mechanisms: those affected would have been NAD(H)-deficient with an element of pellagra [321, 322]. Optimising choline and other aspects of methyl metabolism in embryonic life improves brain development and strikingly reduces age-related declines, perhaps through reducing epigenetic instability. Epigenomes, which may be marked by dietary, emotional, and intellectual experiences working through neuroendocrine stress pathways that themselves are redox and energy-regulated and implicated with many “modern” diseases, can be assimilated into the germline and are unstable, creating somatic epimutations during mitoses, some of which may be resistant to erasure. Surviving epigenetic imprints may be the missing transgenerational elements, along with nonshared environments (merging nature and nurture), causing both familial clustering and sporadic cases in diseases such as AD and PD. Twin studies show low concordance in AD and PD, thereby demonstrating the importance of environmental dynamics and chance epigenetic drift over lifetimes [323].
9. Development to Decline: Managing Metabolic Expectations
Within the context of lifetime availability of NAD(H), where in adult life the net free energy or the availability of methyl groups is greater or lesser than that in early life or expected a priori by the genome of the individual, an imbalance between (epi)genetic-primed protein inductions and nicotinamide may occur if circumstance overcome tastes, habits, and addictions that attempt to stabilise intake. This imbalance is more nuanced than might be expected and not simply due to the fact that modern diets are mismatched with genetic “thrifty” gene hangovers from the Pleistocene [324]. Our evolutionary history of extreme fluctuations in meat intake from low to high as hunter-gatherers to patchy and still in transition is likely to be more relevant. Nutrigenomic and informatic adaptations and changes in tradeoffs resulting from knowing that the energy future is often more likely to be benign have occurred over the last 100,000 years and during individuals lifetimes. Low levels of nicotinamide and choline in early life adversely affect neonatal brain development and later degeneration [325]. Low NAD(H) levels in the foetal/neonatal period could result in a phenotype which could be reinforced by low levels through life, giving rise to poor brain development and function. High early levels could lead to good brain development (“luxury” phenotype) but a drop in the supply later in life, even to average levels when NNMT, the catabolic enzyme, has been induced, could trigger intracellular pellagra. If the NAD(H) and nicotinamide supply is superabundant throughout, life toxicity might result from excessive N-methyl-nicotinamide, a MPTP lookalike (Figure 3). This is plausible because nicotinamide and redox status have marked morphogenic effects that instruct internally secreted morphogens which regulate all phases of neuronal development and in effect brain evolution [326–328]. The effects can become pathological when unexpected energy circumstances are encountered, as seen with metabolic syndrome, and we propose neurological disease [329].
Figure 3.
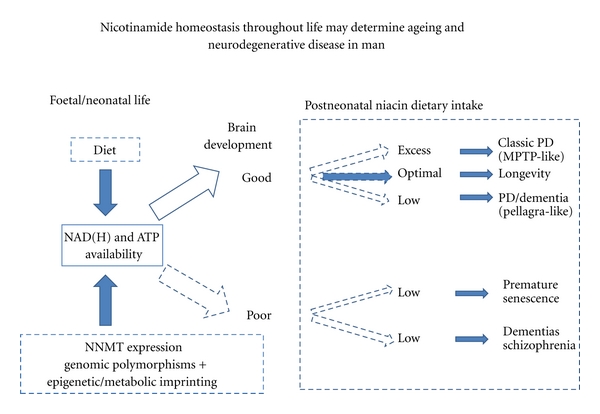
NAD(H) availability is key to good brain evolution and development. An inherent weak link is poor supply or a potential imbalance between induction of NNMT and nicotinamide and choline intake in early and later life with a risk of late degenerations such as Alzheimer's and Parkinson's disease. Overinduction of NNMT by nicotinamide could paradoxically cause intracellular pellagra perhaps explaining why nicotinamide can appear to help short-term even though nicotinamide may be a long-term toxin. In addition, poor diet or induction of NNMT disturbs the methylome and the epigenome creating cancer promoting epimutations.
10. Metabolic Fields
Accumulating evidence points to the probability that ageing diseases such as AD, PD, cancers, and metabolic diseases originate in mitochondrial depolarisations and uncouplings with disturbed electron-proton flows as “protonopathies” and have consequent effects on ATP [330–333]. A unifying explanation imagines abnormally spreading metabolic fields, if the challenges of balancing NADH- dependent energy supply and demand and the need for optimal methyl group availability that arises during growth, reproduction, exercise, and tissue repair are not met; most cancers, for instance are demethylated and have changed their energy metabolism profile even in precancerous stages [334]. These challenges could happen at the same stage of life in diverse species, explaining similar incidences of cancer and degeneration despite large variations in numbers of cells/body size and longevity that are hard to explain on a somatic mutational or purely immunological basis which may be secondary or compounding phenomena [335]. Reactions to stabilize metabolism by excreting NAD(P)H (essentially a waste product not a source of energy in cells using anaerobic metabolism) or other reducing equivalents (e.g., lactate), such as autocarnivory/autophagy/apoptosis and the Warburg effect (such that both the oxic and, of course, the anoxic portions of the tumor are both fermenting) could spread like a wave: as do their pathological correlates, whether degenerations, infections, or cancers. NAD metabolizing ectoenzymes and salvage pathways appear to be important in shaping tumor-host interactions with mitochondrial pathways affecting whether or not cancer cells live or are primed for death [255, 336–344]. NNMT activity affects proliferation of cancer cells, affects radiation sensitivity, and alters free radical production; nicotinamide affects genomic instability and cancer cells eat tryptophan [340, 345, 346]. These heavily involved pathways have opened therapeutic possibilities for several tumours including glioblastoma where the importance of mitochondrial dynamics is increasingly recognised [347]. In cancer cells many germline and somatic mutations continue to evolve as cancer spreads, for example, in p53 and relevant receptors, such as those for steroids and growth factor signals. Thus glucose uptake and mitochondrial and isocitrate and phosphoglycerate dehydrogenases and pyruvate kinases are affected, allowing multiple switches from oxidative metabolism to net NAD(P)H production affecting both energetic and redox buffering/antioxidant capacity [262, 348–355]. The paradox here if solely seen from the cancers point of view, rather than viewing this as a pseudo-symbiosis, is that the growing tissue is wasting energy and the argument that it gets ATP faster through glycolysis is weak [356]. Cancers cannot always be eliminated and often recur (though occasionally spontaneously remit) or a second cancer develops, suggesting that the basic problem is not eliminated by simply killing cancer and surrounding normal cells (which may however temporarily improve the local energy environment by releasing ATP and its precursors).
Foetal implants develop PD pathology, illustrating the point that the niche microenvironment is the key [357–363]. Stem cell implants in PD and other conditions survive and connect but form strange lesions and connections suggesting that lack of stem cells is not the real deficit. Rather stem-cell-based therapy from the haematological transplant to those for other organs involves the efflux of NAD and ATP into the extracellular space or autocrine induction of NAD release activating purinergic and calcium pathways affecting growth-supporting and antiapoptotic activities relevant to their therapeutic effects [364]. Parabiotic fusions of young environments rejuvenate cells, even those expressing procancerous somatic mutations or destined for autophagy [365] and changing the ageing milieu, alter neurogenesis and improves cognitive function. Stem cell fates toward both normal and abnormal differentiation or dedifferentiation depend on their microenvironment and stochastic factors, and even cells that are already cancerous can be induced to behave normally [366–369]. Protein folding and the function of amyloid/synuclein/prion proteins and ionic and neurotransmitter gradients are exquisitely sensitive to energy landscapes, as are NAD(H)-consumer linked DNA repairs. Consequently, all these phenomena, when abnormal, may be secondary and late effects of abnormal NAD(H) ratios and methylation status affecting epigenomic phenotypic plasticity.
11. Regeneration and Degeneration on Purpose
In complex organisms stem-cell-driven regeneration, which could compensate for long-term damage that is beyond repair, is suppressed particularly in the CNS, whilst dying cells from traumatic injury releasing NADH equivalents may be a potent stimulus for regeneration in other tissues and simpler organisms [370]. This apparent design failure, which is actively pursued by complex organisms, may be rooted in energy allocation models, that is, in tradeoffs that favour building spare capacity during development. Spare capacity when young, which can be coopted as an exaptation for early in life innovative thinking about the environment, may be evolutionarily more advantageous than maintaining, at high cost, regenerative machinery permanently on stand-by that may never get used.
The brain functions within the constraints of an energy (ATP) budget, with high levels of energy used to process information from the environment via action potentials and less energy intensive metabolic processes used to recycle neurotransmitters [371–374]. There is evidence that this budget evolved to permit other tradeoffs allowing neuronal energy costs to remain low while still allowing the organism to gather all required information. In environments where certain sensory information is not needed, evolution has led to the loss of superfluous sensory systems. This responsive and adaptable evolutionary process can also lead to the addition of sensory systems as required by changing environments. Such additions and deletions develop as a result of an advantageous cost/benefit analysis by the evolving organism responding to changing environments and to learn for specific tasks as well as for general intelligence [375].
Poor development from poor early energy circumstances and loss of reserve capacity would be expected to cause late problems, as suspected with AD. Nevertheless, continual regeneration and selection of circuits that are using energy are constantly occurring by altering mitochondrial and SIRT function, and bioelectric free energy and proton fields and fluxes, that influence positional memory and regeneration of even complex structures such as the spinal cord (giving hope that improving the NAD(H) and methyl environment is never too late) [7, 376–378].
12. Energy Circuits Can Breakdown
We live in a complex habitat and an energy and informatic subsistence environment largely of our own making that has driven our brain and behaviour patterns to ecological rationality, in order to cope with being far from thermodynamic equilibrium, with strong positive feed-forward decision-making and negative feedback homeostatic mechanisms that drive progress. Less fortunately disease phenotypes from the cancerous to the neuropsychiatric happen when it goes wrong, as happened in the case of pellagra. Pellagra like most famines was not wholly caused by natural disasters but man-made collapses at a series of hierarchical levels from the social to the subcellular. Ever since Homo Erectus, our genus has been addicted to acquiring more and more energy, first as NADH and then harnessing all forms of energy, with a push from coping with poor energy environments and a pull from using excess energy to best advantage through the use of our collective imagination and each other: much current intellectual effort is currently expended on discovering new sources of energy especially those based on hydrogen [379, 380] (Table 1). Many memes/ideas are generated and their survival may often have been dependent upon whether they contain energy-useful ideas, with logic systems working at biological and behavioral levels building on simpler circuits such as the iconic transcriptional regulation of the lac operon in response to changing energy sources. This requires a high quality diet and gut symbionts forming superorganisms with extended metabolisms that are not simply divided by skin or skull from the world but are complementary systems that still have fault lines that can breakdown. Self, as usually defined by the immunologists, is misleading, and more modern views on metagenomes are closer to the mark and, like the psychologist William James, define self as “all a man can call his” and includes his sources of energy and information [381].
Table 1.
Three worlds that interact are largely human thermodynamic and cognitive niche constructions that are all heritable making us codirectors of our own and our collaborators evolution. There is a cost to this cumulative evolution that may include hypo- and hypervitaminosis, energy, and informatic states.
| Thermodynamic and material | Personal | Information and society |
|---|---|---|
| Omnivorous diet | Conscious perception | Divisions of labour |
| Trading symbionts | Empathy | Trading |
| Shelter/clothes | Mental time travel | Information stores |
| Know-how | ||
| Language | ||
| Nutrigenomics | ||
| Immunogenomics | ||
| Neuro/endocrine homeostasis |
Surplus energy for innovation of tools and technology and further harnessing of energy.
Despite consuming a high proportion of the body's energy, brains are efficient (Watson, IBM's latest AI answer machine runs on 100 Kw compared with 100 w for a brain) and lose functions no longer used, suggesting strict constraints on bioenergy allowing the brain to continually evolve along with the changing environment, with considerable potential for human enhancement if the environment improves [382]. Enrichment of the cell's microenvironmental energy ecosystem and a “use it or lose it” strategy leading to energy flow may be key to improvements in the longevity of many cell types and in prompting neural stem cells, “hungry for action”, to divide. Maintaining optimal NADH : NAD+ ratios and methylation status through dynamic energy budgets in every compartment is a continual challenge which is awe-inspiring in its complexity. Nevertheless, such considerations are necessary for every single cell if they are to avoid death or cancerous change and may be aided by quantum effects on the distribution of energy [383]. High energy circuits and electrical waves, such as gamma oscillations, are needed for intellectual hippocampal and motor programmes that require strong functional performance of mitochondria and particularly need the pyramidal neurones that take a big hit in pellagra [384, 385]. These circuits are synchronous with circadian NAD(H)-dependent informatic rhythms between individuals and their diets, symbionts and social relationships and beliefs, some mediated by hormones such as oxytocin and neurotransmitters such as dopamine. Sleep gives time for synaptic normalization after dealing with ever-changing environments and unsustainable consumption of energy with saturated abilities to learn as environmentally cued memory and free-will decision making, over and above reflex preactivation of volition, require particularly high energy levels: these are functions hit early in cortical and basal ganglia degenerations some of which compensate by the patients developing a sweet tooth or otherwise modify energy intake or expenditure as part of the symptoms or attempts at treatment [386–396].
These environmentally coupled systems, largely of our own making to keep energy flowing and balancing excitatory and inhibitory circuits, can get decoupled with energy or informatic under or overload (“rewired and running hot”) and may be hit preferentially in PD and AD and other diseases peculiar to Homo sapiens, such as depression and psychoses where nicotinamide aberrations have long been suspected [358, 359]. We urgently need to improve our abilities to measure or image suitable NADH supplies in a series of human habitats on the outsides of our Russian Doll Redox arrangement and NAD(P)H/NAD(P)+ ratios in every internal compartment, cell type and organelle as early warning systems to intervene and avoid pathology even getting started.
13. Conclusion
Classical pellagra is an archetypal energy and NAD supply-side premature ageing disease, with excessive use of symbionts and degenerative pathology, that has been cured but we have never checked to see whether subpellagrous NADH deficiency has been eliminated. Sub pellagrous NAD deficiency may be a man-made socioeconomic “place” disease that rears its head episodically as a “time” disease triggering age-related infections, degenerations, and cancers [397]. This type of socioeconomic illness does not naturally trigger an empathetic response reducing the chances of resolving the situation [398]. Interactions with other vitamins may be important either with deficits or excesses as we have implied for choline, and for example, the combination of low vitamin D intake with genetic predisposing defects, and, NAD supplies, and, altered relationships with microorganisms triggering autoimmune demyelination may be important in the pathogenesis of multiple sclerosis [399, 400]. Intracellular pellagra may also be caused by other mechanisms such as high NAD expenditure after physical, emotional, or chemical toxic stress or acute infection or inadvertent removal of key symbionts, such as after antibiotic use, or mutations that impair autophagosome functions such that at some times or for some people the dose may need to be higher than usually recommended. Stress from loss of relative status is a powerful driver of ill health and primates including man revert to a high glucose and low nicotinamide diet or addictive behaviours that as with stress also induce NNMT that will further lower nicotinamide levels [401, 402].
One should not, on the other hand, expose populations to “too much of a good thing” with excessive supplies of nicotinamide or choline or calories and thereby perturb NAD : NADH ratios in favour of NADH, creating a mirror image of pellagra, perhaps with an equally broad phenotype [403]. Modern disease patterns may be contemporary mirror images of the Neolithic meat transition when with less meat fertility rose but so did disease reducing longevity, whereas now as meat eating is on the increase longevity is increasing with fertility dropping and different chronic diseases emerging, and others such as TB rapidly decreasing.
Poor worldwide distribution of NAD(H) supplies makes little sense if oversupply is dangerous, particularly if undersupply is not only dangerous to those affected but leads to new symbionts or drug resistant microbes aided and abetted by the host in metabolic desperation that can then infect others who are more affluent and cause potentially epidemic pathology [403]. Our current behaviour that does not deal with this urgently “altruistically” may need the insight that as group sizes were enlarged we moved away from our hunter-gatherer roots of an egalitarian norm and a sharing of nicotinamide and methyl group rich meat. “Stinginess” may date from when the natural meat supply deteriorated and meat and the considerable wherewithal to produce it became treated as property. Fighting and migrating were preferred over starving, and trading may have promoted specialisation, competition, and inequality including of access to meat [404]. Resource allocation both of food and of medicines is culturally driven with a tension as to whether those that earn the most or those with the greatest need are given preference. The former is the usual modern default option but is a cultural “homo-economicus” evolutionary trap that needs to be consciously and socially corrected [405–407]. Fairer access to high quality energy sources allows all people the choice of moderation and a Hippocratic style diet and exercise regimen, and, access to parallel drug developments which have hormetic and preconditioning effects: that test the energy axis, induce autophagy and unfolded protein targeting and endoplasmic reticulum stress responses to recycle protein debris, thus preventing and “vaccinating” against disease [408–410]. New treatments that quickly supply energy, or save energy by inducing artificial hibernation, if employed after an unexpected insult such as trauma, and allow natural healing to take place, may also work best [411].
Humans have been exploring ways of preventing cancer and degeneration for decades whereas evolution has coped with episodes of NAD(H) deficiency and excess for several billion years. Now it may be time to learn lessons from the experts and find a Goldilocks energy-methylome-informatic-economy that matches supply and demand throughout life. Fewer energy tradeoffs or faulty signals may avoid lost robustness, shorter lives and loss of a fitness trait such as intellectual or physical capacity as seen with the degenerative AD and PD or the proliferative cancers, inflammatory and allergic diseases.
References
- 1.Collins PY, Patel V, Joestl SS, et al. Grand challenges in global mental health. Nature. 2011;475(7354):27–30. doi: 10.1038/475027a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Huber M, Knottnerus JA, Green L, et al. Health: how should we define it? British Medical Journal. 2011;343:235–237. [Google Scholar]
- 3.McEwen BS. Interacting mediators of allostasis and allostatic load: towards an understanding of resilience in aging. Metabolism. 2003;52(2):10–16. doi: 10.1016/s0026-0495(03)00295-6. [DOI] [PubMed] [Google Scholar]
- 4.Chandra FA, Buzi G, Doyle JC. Glycolytic oscillations and limits on robust efficiency. Science. 2011;333(6039):187–192. doi: 10.1126/science.1200705. [DOI] [PubMed] [Google Scholar]
- 5.Bray D. Wetware: A Computer in Every Living Cell. London, UK: Yale University Press; 2009. [Google Scholar]
- 6.Nurse P. Life, logic and information. Nature. 2008;454(7203):424–426. doi: 10.1038/454424a. [DOI] [PubMed] [Google Scholar]
- 7.Baquero F, Alvarez-Ortega C, Martinez JL. Ecology and evolution of antibiotic resistance. Environmental Microbiology Reports. 2009;1(6):469–476. doi: 10.1111/j.1758-2229.2009.00053.x. [DOI] [PubMed] [Google Scholar]
- 8.Grant PR, Grant BR. Evolution of character displacement in Darwin’s finches. Science. 2006;313(5784):224–226. doi: 10.1126/science.1128374. [DOI] [PubMed] [Google Scholar]
- 9.Ffrench-Constant RH, Daborn PJ, Le Goff G. The genetics and genomics of insecticide resistance. Trends in Genetics. 2004;20(3):163–170. doi: 10.1016/j.tig.2004.01.003. [DOI] [PubMed] [Google Scholar]
- 10.Offer A, Pechey R, Ulijaszek S. Obesity under affluence varies by welfare regimes: the effect of fast food, insecurity, and inequality. Economics and Human Biology. 2010;8(3):297–308. doi: 10.1016/j.ehb.2010.07.002. [DOI] [PubMed] [Google Scholar]
- 11.Tiffin R, Salois M. Inequalities in diet and nutrition. In: Proceedings of the The Summer Meeting of the Nutrition Society. 70th Anniversary Conference on “From Plough through Practice to Policy” Symposium 1: Food Chain and Health; July 2011; University of Reading; [Google Scholar]
- 12.Finlay BL, Hinz F, Darlington RB. Mapping behavioural evolution onto brain evolution: the strategic roles of conserved organization in individuals and species. Philosophical Transactions of the Royal Society B. 2011;366(1574):2111–2123. doi: 10.1098/rstb.2010.0344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Matthew T, Tickle C. Generation of pattern and form in the developing limb. International Journal of Developmental Biology. 2009;53(5-6):805–812. doi: 10.1387/ijdb.072499mt. [DOI] [PubMed] [Google Scholar]
- 14.Hoppeler H, Weibel ER. Limits for oxygen and substrate transport in mammals. Journal of Experimental Biology. 1998;201(8):1051–1064. doi: 10.1242/jeb.201.8.1051. [DOI] [PubMed] [Google Scholar]
- 15.Alexander R. A theory of mixed chains applied to safety factors in biological systems. Journal of Theoretical Biology. 1997;184(3):247–252. doi: 10.1006/jtbi.1996.0270. [DOI] [PubMed] [Google Scholar]
- 16.Innocenti GM. Development and evolution. Two determinants of cortical connectivity. Progress in Brain Research. 2011;189:65–75. doi: 10.1016/B978-0-444-53884-0.00018-X. [DOI] [PubMed] [Google Scholar]
- 17.Rajan A, Perrimon N. Drosophila as a model for interorgan communication: lessons from studies on energy homeostasis. Developmental Cell. 2011;21(1):29–31. doi: 10.1016/j.devcel.2011.06.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Schlaepfer MA, Runge MC, Sherman PW. Ecological and evolutionary traps. Trends in Ecology and Evolution. 2002;17(10):474–480. [Google Scholar]
- 19.Raghanti MA, Simic G, Watson S, Stimpson CD, Hof PR, Sherwood CC. Compartative analysis of the nucleus basalis of Meynert among primates. Neuroscience. 2011;184:1–15. doi: 10.1016/j.neuroscience.2011.04.008. [DOI] [PubMed] [Google Scholar]
- 20.Raghanti MA, Stimpson CD, Marcinkiewicz JL, Erwin JM, Hof PR, Sherwood CC. Cortical dopaminergic innervation among humans, chimpanzees, and macaque monkeys: a comparative study. Neuroscience. 2008;155(1):203–220. doi: 10.1016/j.neuroscience.2008.05.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Adams JD, Chang ML, Klaidman L. Parkinson’s disease—redox mechanisms. Current Medicinal Chemistry. 2001;8(7):809–814. doi: 10.2174/0929867013372995. [DOI] [PubMed] [Google Scholar]
- 22.Kapogiannis D, Mattson MP. Disrupted energy metabolism and neuronal circuit dysfunction in cognitive impairment and Alzheimer’s disease. The Lancet Neurology. 2011;10(2):187–198. doi: 10.1016/S1474-4422(10)70277-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Schrack JA, Simonsick EM, Ferrucci L. The energetic pathway to mobility loss: an emerging new framework for longitudinal studies on aging. Journal of the American Geriatrics Society. 2010;58(2, supplement):S329–S336. doi: 10.1111/j.1532-5415.2010.02913.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Fadini GP, Ceolotto G, Pagnin E, De Kreutzenberg S, Avogaro A. At the crossroads of longevity and metabolism: the metabolic syndrome and lifespan determinant pathways. Aging Cell. 2011;10(1):10–17. doi: 10.1111/j.1474-9726.2010.00642.x. [DOI] [PubMed] [Google Scholar]
- 25.Cunnane S, Nugent S, Roy M, et al. Brain fuel metabolism, aging, and Alzheimer’s disease. Nutrition. 2011;27(1):3–20. doi: 10.1016/j.nut.2010.07.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lazarov O, Mattson MP, Peterson DA, Pimplikar SW, van Praag H. When neurogenesis encounters aging and disease. Trends in Neurosciences. 2010;33(12):569–579. doi: 10.1016/j.tins.2010.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Van Leeuwen IM, Vera J, Wolkenhauer O. Dynamic energy budget approaches for modelling organismal ageing. Philosophical Transactions of the Royal Society B. 2010;365(1557):3443–3454. doi: 10.1098/rstb.2010.0071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kelly DP. Cell biology: ageing theories unified. Nature. 2011;470(7334):342–343. doi: 10.1038/nature09896. [DOI] [PubMed] [Google Scholar]
- 29.Kowald A, Kirkwood TB. Evolution of the mitochondrial fusion-fission cycle and its role in aging. Proceedings of the National Academy of Sciences of the United States of America. 2011;108(25):10237–10242. doi: 10.1073/pnas.1101604108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Leloup C, Casteilla L, Carrière A, et al. Balancing mitochondrial redox signaling: a key point in metabolic regulation. Antioxidants and Redox Signaling. 2011;14(3):519–530. doi: 10.1089/ars.2010.3424. [DOI] [PubMed] [Google Scholar]
- 31.Perluigi M, Di Domenico F, Giorgi A, et al. Redox proteomics in aging rat brain: involvement of mitochondrial reduced glutathione status and mitochondrial protein oxidation in the aging process. Journal of Neuroscience Research. 2010;88(16):3498–3507. doi: 10.1002/jnr.22500. [DOI] [PubMed] [Google Scholar]
- 32.Surmeier DJ, Guzman JN, Sanchez-Padilla J, Schumacker PT. The role of calcium and mitochondrial oxidant stress in the loss of substantia nigra pars compacta dopaminergic neurons in Parkinson's disease. Neuroscience. 2011;198:221–231. doi: 10.1016/j.neuroscience.2011.08.045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Batlevi Y, La Spada AR. Mitochondrial autophagy in neural function, neurodegenerative disease, neuron cell death, and aging. Neurobiology of Disease. 2011;43(1):46–51. doi: 10.1016/j.nbd.2010.09.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Luce K, Weil AC, Osiewacz HD. Mitochondrial protein quality control systems in aging and disease. Advances in Experimental Medicine and Biology. 2010;694:108–125. doi: 10.1007/978-1-4419-7002-2_9. [DOI] [PubMed] [Google Scholar]
- 35.Wallace DC. Bioenergetics and the epigenome: interface between the environment and genes in common diseases. Developmental Disabilities Research Reviews. 2010;16(2):114–119. doi: 10.1002/ddrr.113. [DOI] [PubMed] [Google Scholar]
- 36.Roth J, Szulc AL, Danoff A. Energy, evolution, and human diseases: an overview. American Journal of Clinical Nutrition. 2011;93(4):875S–883S. doi: 10.3945/ajcn.110.001909. [DOI] [PubMed] [Google Scholar]
- 37.Green DR, Galluzzi L, Kroemer G. Mitochondira and the autophagy-inflammation-cell death axis in organismal aging. Science. 2011;333(6046):1109–1112. doi: 10.1126/science.1201940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bland ML, Birnbaum MJ. Adapting to energetic stress. Science. 2011;332(6036):1387–1388. doi: 10.1126/science.1208444. [DOI] [PubMed] [Google Scholar]
- 39.Speakman JR, Król E. Limits to sustained energy intake. XIII. Recent progress and future perspectives. Journal of Experimental Biology. 2011;214(2):230–241. doi: 10.1242/jeb.048603. [DOI] [PubMed] [Google Scholar]
- 40.Noakes TD. Time to move beyond a brainless exercise physiology: the evidence for complex regulation of human exercise performance. Applied Physiology, Nutrition and Metabolism. 2011;36(1):23–35. doi: 10.1139/H10-082. [DOI] [PubMed] [Google Scholar]
- 41.Lane N. Power, Sex, Suicide: Mitochondria and the Meaning of Life. New York, NY, USA: OUP; 2005. [Google Scholar]
- 42.Sies H, Bricelius R, Wefers H, Müller A, Cadenas E. Cellular redox changes and response to drugs and toxic agents. Toxicological Sciences. 1983;3(4):200–208. doi: 10.1016/s0272-0590(83)80126-2. [DOI] [PubMed] [Google Scholar]
- 43.Mailloux RJ, Bériault R, Lemire J, et al. The tricarboxylic acid cycle, an ancient metabolic network with a novel twist. Plos ONE. 2007;2(8, article e690) doi: 10.1371/journal.pone.0000690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Buler M, Aatsinki SM, Skoumal R, Hakkola J. Energy sensing factors PGC-1α and SIRT1 modulate PXR expression and function. Biochemical Pharmacology. 2011;82(12):2008–2015. doi: 10.1016/j.bcp.2011.09.006. [DOI] [PubMed] [Google Scholar]
- 45.Schoener TW. The newest synthesis: understanding the interplay of evolutionary and ecological dynamics. Science. 2011;331(6016):426–429. doi: 10.1126/science.1193954. [DOI] [PubMed] [Google Scholar]
- 46.Rankin DJ. The social side of Homo economicus. Trends in Ecology and Evolution. 2011;26(1):1–3. doi: 10.1016/j.tree.2010.10.005. [DOI] [PubMed] [Google Scholar]
- 47.Williams AC, Ramsden DB. Pellagra: a clue as to why energy failure causes diseases? Medical Hypotheses. 2007;69(3):618–628. doi: 10.1016/j.mehy.2007.01.029. [DOI] [PubMed] [Google Scholar]
- 48.Marmot M, Friel S, Bell R, Houweling TA, Taylor S. Closing the gap in a generation: health equity through action on the social determinants of health. The Lancet. 2008;372(9650):1661–1669. doi: 10.1016/S0140-6736(08)61690-6. [DOI] [PubMed] [Google Scholar]
- 49.Rose G. Sick individuals and sick populations. International Journal of Epidemiology. 1985;30(3):427–432. doi: 10.1093/ije/30.3.427. [DOI] [PubMed] [Google Scholar]
- 50.Di Domenico F, Perluigi M, Butterfield DA, Cornelius C, Calabrese V. Oxidative damage in rat brain during aging: interplay between energy and metabolic key target proteins. Neurochemical Research. 2010;35(12):2184–2192. doi: 10.1007/s11064-010-0295-z. [DOI] [PubMed] [Google Scholar]
- 51.Paul L. Diet, nutrition and telomere length. Journal of Nutritional Biochemistry. 2011;22(10):895–901. doi: 10.1016/j.jnutbio.2010.12.001. [DOI] [PubMed] [Google Scholar]
- 52.Drenos F, Kirkwood TB. Modelling the disposable soma theory of ageing. Mechanisms of Ageing and Development. 2005;126(1):99–103. doi: 10.1016/j.mad.2004.09.026. [DOI] [PubMed] [Google Scholar]
- 53.Flatt T, Schmidt PS. Integrating evolutionary and molecular genetics of aging. Biochimica et Biophysica Acta. 2009;1790(10):951–962. doi: 10.1016/j.bbagen.2009.07.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Fontana L, Partridge L, Longo VD. Extending healthy life span-from yeast to humans. Science. 2010;328(5976):321–326. doi: 10.1126/science.1172539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Krebs JR. The gourmet ape: evolution and human food preferences. American Journal of Clinical Nutrition. 2009;90(3):707–711. doi: 10.3945/ajcn.2009.27462B. [DOI] [PubMed] [Google Scholar]
- 56.Navarrete A, van Schaik CP, Isler K. Energertics and the evolution of human brain size. Nature. 2011;480:91–93. doi: 10.1038/nature10629. [DOI] [PubMed] [Google Scholar]
- 57.Kaplan H, Hill K, Lancaster J, Hurtado AM. A theory of human life history evolution: diet, intelligence, and longevity. Evolutionary Anthropology. 2000;9(4):156–185. [Google Scholar]
- 58.Sol D. Opinion piece. Revisiting the cognitive buffer hypothesis for the evolution of large brains. Biology Letters. 2009;5(1):130–133. doi: 10.1098/rsbl.2008.0621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Ma H, Sorokin A, Mazein A, et al. The Edinburgh human metabolic network reconstruction and its functional analysis. Molecular Systems Biology. 2007;3, article 135 doi: 10.1038/msb4100177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Zhao J, Yu H, Luo JH, Cao ZW, Li YX. Hierarchical modularity of nested bow-ties in metabolic networks. BMC Bioinformatics. 2006;7, article 386 doi: 10.1186/1471-2105-7-386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Csete M, Doyle J. Bow ties, metabolism and disease. Trends in Biotechnology. 2004;22(9):446–450. doi: 10.1016/j.tibtech.2004.07.007. [DOI] [PubMed] [Google Scholar]
- 62.Sahar S, Nin V, Barbosa MT, Chini EN, Sassone-Corsi P. Altered behavioural and metabolic circadian rhythms in mice with disrupted NAD+ oscillation. Aging. 2001;3(8):794–802. doi: 10.18632/aging.100368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Schneider ED, Sagan D. Into the Cool—Energy Flow, Thermodynamics, and Life. Illinois, Ill, USA: The University of Chicago Press; 2005. [Google Scholar]
- 64.Lampl M. Cellular life histories and bow tie biology. American Journal of Human Biology. 2005;17(1):66–80. doi: 10.1002/ajhb.20094. [DOI] [PubMed] [Google Scholar]
- 65.Goodman M, Sternera KN. Phylogenomic evidence of adaptive evolution in the ancestry of humans. Proceedings of the National Academy of Sciences of the United States of America. 2010;107(2):8918–8923. doi: 10.1073/pnas.0914626107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Aita T, Husimi Y. Biophysical connection between evolutionary dynamics and thermodynamics in in vitro evolution. Journal of Theoretical Biology. 2012;294:122–129. doi: 10.1016/j.jtbi.2011.10.036. [DOI] [PubMed] [Google Scholar]
- 67.Speakman JR, Selman C. The free-radical damage theory: accumulating evidence against a simple link of oxidative stress to ageing and lifespan. BioEssays. 2011;33(4):255–259. doi: 10.1002/bies.201000132. [DOI] [PubMed] [Google Scholar]
- 68.Margulis L, Sagan D. Acquiring Genomes: A Theory of the Origins of Species. New York, NY, USA: Basic Books; 2002. [Google Scholar]
- 69.Valente TW. Social Networks and Health—Models, Methods and Applications. London, UK: Oxford University Press; 2010. [Google Scholar]
- 70.Smil V. Energy in Nature and Society: General Energetics of Complex Systems. Cambridge, UK: MIT Press; 2008. [Google Scholar]
- 71.David LA, Alm EJ. Rapid evolutionary innovation during an Archaean genetic expansion. Nature. 2011;469:93–96. doi: 10.1038/nature09649. [DOI] [PubMed] [Google Scholar]
- 72.Kramer KL. The evolution of hman parental care and recruitment of juvenile help. Trends in Ecology and Evolution. 2011;26(10):533–540. doi: 10.1016/j.tree.2011.06.002. [DOI] [PubMed] [Google Scholar]
- 73.Santos FC, Pinheiro FL, Lenaerts T, Pacheco JM. The role of diversity in the evolution of cooperation. doi: 10.1016/j.jtbi.2011.09.003. Journal of Theoretical Biology. In press. [DOI] [PubMed] [Google Scholar]
- 74.Soares MC, Bshary R, Fusani L, et al. Hormonal mechanisms of cooperative behaviour. Philosophical Transactions of the Royal Society B. 2010;365(1553):2737–2750. doi: 10.1098/rstb.2010.0151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Melis AP, Semmann D. How is human cooperation different? Philosophical Transactions of the Royal Society B. 2010;365(1553):2663–2674. doi: 10.1098/rstb.2010.0157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Bergmüller R, Schürch R, Hamilton IM. Evolutionary causes and consequences of consistent individual variation in cooperative behaviour. Philosophical Transactions of the Royal Society B. 2010;365(1553):2751–2764. doi: 10.1098/rstb.2010.0124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Vinciguerra M, Fulco M, Ladurner A, Sartorelli V, Rosenthal N. SirT1 in muscle physiology and disease: lessons from mouse models. DMM Disease Models and Mechanisms. 2010;3(5-6):298–303. doi: 10.1242/dmm.004655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Wang Y, Liang Y, Vanhoutte PM. SIRT1 and AMPK in regulating mammalian senescene: a critical review and a working model. FEBS Letters. 2010;585(7):986–994. doi: 10.1016/j.febslet.2010.11.047. [DOI] [PubMed] [Google Scholar]
- 79.Finkel T, Deng CX, Mostoslavsky R. Recent progress in the biology and physiology of sirtuins. Nature. 2009;460(7255):587–591. doi: 10.1038/nature08197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Milner J. Cellular regulation of SIRT1. Current Pharmaceutical Design. 2009;15(1):39–44. doi: 10.2174/138161209787185841. [DOI] [PubMed] [Google Scholar]
- 81.Brooks CL, Gu W. How does SIRT1 affect metabolism, senescence and cancer? Nature Reviews Cancer. 2009;9(2):123–128. doi: 10.1038/nrc2562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Feige JN, Johan A. Transcriptional targets of sirtuins in the coordination of mammalian physiology. Current Opinion in Cell Biology. 2008;20(3):303–309. doi: 10.1016/j.ceb.2008.03.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Cantó C, Auwerx J. PGC-1α, SIRT1 and AMPK, an energy sensing network that controls energy expenditure. Current Opinion in Lipidology. 2009;20(2):98–105. doi: 10.1097/MOL.0b013e328328d0a4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Bass J, Takahashi JS. Circadian integration of metabolism and energetics. Science. 2010;330(6009):1349–1354. doi: 10.1126/science.1195027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Vassilopoulos A, Fritz KS, Petersen DR, Gius D. The human sirtuin family: evolutinary divergences and functions. Human Genomics. 2011;5(5):485–496. doi: 10.1186/1479-7364-5-5-485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Kauppinen TM, Swanson RA. The role of poly(ADP-ribose) polymerase-1 in CNS disease. Neuroscience. 2007;145(4):1267–1272. doi: 10.1016/j.neuroscience.2006.09.034. [DOI] [PubMed] [Google Scholar]
- 87.Abd Elmageed ZY, Naura AS, Errami Y, Zerfaoui M. The Poly(ADP-ribose) polymerases (PARPs): new roles in intracellular transport. Cellular Signalling. 2012;24(1):1–8. doi: 10.1016/j.cellsig.2011.07.019. [DOI] [PubMed] [Google Scholar]
- 88.Avery MA, Sheehan AE, Kerr KS, Wang J, Freeman MR. Wlds requires Nmnatl enzymatic activity and N16- VCP interactions to suppress Wallerian degeneration. Journal of Cell Biology. 2009;184(4):501–513. doi: 10.1083/jcb.200808042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Yahata N, Yuasa S, Araki T. Nicotinamide mononucleotide adenylyltransferase expression in mitochondrial matrix delays Wallerian degeneration. Journal of Neuroscience. 2009;29(19):6276–6284. doi: 10.1523/JNEUROSCI.4304-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Williams AC, Ramsden DB. Nicotinamide homeostasis: a xenobiotic pathway that is key to development and degenerative diseases. Medical Hypotheses. 2005;65(2):353–362. doi: 10.1016/j.mehy.2005.01.042. [DOI] [PubMed] [Google Scholar]
- 91.Stearns SC, Nesse RM, Govindaraju DR, Ellison PT. Evolution in health and medicine Sackler colloquim: evolutionary perspectives on health and medicine. Proceedings of the National Academy of Sciences of the United States of America. 2010;107(supplement 1):1691–1695. doi: 10.1073/pnas.0914475107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Liow LH, Van Valen L, Stenseth NC. Red Queen: from populations to taxa and communities. Trends in Ecology and Evolution. 2011;26(7):349–358. doi: 10.1016/j.tree.2011.03.016. [DOI] [PubMed] [Google Scholar]
- 93.Roberts I. The Energy Glut. The Politics of Fatness in an Overheating World. London, UK: Zed Books Ltd.; 2010. [Google Scholar]
- 94.Peng Y, Sartini D, Pozzi V, Wilk D, Emanuelli M, Yee VC. Structural basis of substrate recognition in human nicotinamide N-methyltransferase. Biochemistry. 2011;50(36):7800–7808. doi: 10.1021/bi2007614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Parsons RB, Aravindan S, Kadampeswaran A, et al. The expression of nicotinamide N-methyltranferas increases ATP synthesis and protects SH-SY5Y neuroblastoma cells against the toxicity of complex I inhibitors. Biochemical Journal. 2011;436(1):145–155. doi: 10.1042/BJ20101685. [DOI] [PubMed] [Google Scholar]
- 96.Parsons RB, Smith SW, Waring RH, Williams AC, Ramsden DB. High expression of nicotinamide N-methyltransferase in patients with idiopathic Parkinson’s disease. Neuroscience Letters. 2003;342(1-2):13–16. doi: 10.1016/s0304-3940(03)00218-0. [DOI] [PubMed] [Google Scholar]
- 97.Watts AG, Donovan CM. Sweet talk in the brain: glucosensing, neural networks, and hypoglycemic counterregulation. Frontiers in Neuroendocrinology. 2010;31(1):32–43. doi: 10.1016/j.yfrne.2009.10.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Chen X, Gabitto M, Peng Y, Ryba NJP, Zukeer CS. A gustopic map of tase qualities in the mammalian brain. Science. 2011;333(6047):1262–1266. doi: 10.1126/science.1204076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Kaushik S, Rodriguez-Navarro JA, Arias E, et al. Autophagy in hypothalamic agrp neurons regulates food intake and energy balance. Cell Metabolism. 2011;14(2):173–183. doi: 10.1016/j.cmet.2011.06.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Douglas AE. The Symbiotic Habit. New York, NY, USA: Princeton University Press; 2010. [Google Scholar]
- 101.Gophna U. The guts of dietary habits. Science. 2011;334(6052):45–46. doi: 10.1126/science.1213799. [DOI] [PubMed] [Google Scholar]
- 102.Blaser M. Antibiotic overuse: stop the killing of beneficial bacteria. Nature. 2011;476(7361):393–394. doi: 10.1038/476393a. [DOI] [PubMed] [Google Scholar]
- 103.Pester M, Brune A. Hydrogen is the central free intermediate during lignocellulose degradation by termite gut symbionts. ISME Journal. 2007;1(6):551–565. doi: 10.1038/ismej.2007.62. [DOI] [PubMed] [Google Scholar]
- 104.Rosenberg E, Zilber-Rosenberg I. Symbiosis and development: the hologenome concept. Birth Defects Research C. 2011;93(1):56–66. doi: 10.1002/bdrc.20196. [DOI] [PubMed] [Google Scholar]
- 105.Bocquet-Appel J-R, Bar-Yosef O, editors. The Neolithic Demographic Transition and its Consequences. New York, NY, USA: Springer; 2008. [Google Scholar]
- 106.Murphy EF, Cotter PD, Healy S, et al. Composition and energy harvesting capacity of the gut microbiota: relationship to diet, obesity and time in mouse models. Gut. 2010;59(12):1635–1642. doi: 10.1136/gut.2010.215665. [DOI] [PubMed] [Google Scholar]
- 107.Muegge BD, Kuczynski J, Knights D, et al. Diet drives convergence in gut microbiome functions across mannalian phylogeny and within humans. Science. 2011;332(6032):970–971. doi: 10.1126/science.1198719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Armstrong JB, Schindler DE. Excess digestive capacity in predators reflects a life of feast and famine. Nature. 2011;476(7358):84–88. doi: 10.1038/nature10240. [DOI] [PubMed] [Google Scholar]
- 109.Turnbaugh PJ, Gordon JI. The core gut microbiome, energy balance and obesity. Journal of Physiology. 2009;587(17):4153–4158. doi: 10.1113/jphysiol.2009.174136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Donohoe DR, Garge N, Zhang X, et al. The microbiome and butyrate regulate energy metabolism and autophagy in the mammalian colon. Cell Metabolism. 2011;13(5):517–526. doi: 10.1016/j.cmet.2011.02.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Hardie DG. Why starving cells eat themselves. Science. 2011;331(6016):410–411. doi: 10.1126/science.1201691. [DOI] [PubMed] [Google Scholar]
- 112.Rabinowitz JD, White E. Autophagy and metabolism. Science. 2010;330(6009):1344–1348. doi: 10.1126/science.1193497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Cheung ZH, Ip NY. Autophagy deregulation in neurodegenerative diseases—recent advances and future perspectives. Journal of Neurochemistry. 2011;118(3):317–325. doi: 10.1111/j.1471-4159.2011.07314.x. [DOI] [PubMed] [Google Scholar]
- 114.Wallace TC, Guarner F, Madsen K, et al. Human gut microbiota and its relationship to health and disease. Nutrition Reviews. 2011;69(7):392–403. doi: 10.1111/j.1753-4887.2011.00402.x. [DOI] [PubMed] [Google Scholar]
- 115.Levine AJ, Puzio-Kuter AM. The control of the metabolic switch in cancers by oncogenes and tumor suppressor genes. Science. 2010;330(6009):1340–1344. doi: 10.1126/science.1193494. [DOI] [PubMed] [Google Scholar]
- 116.Richardson AD, Scott DA. Reversing the Warburg effect through stromal autophagy. Cell Cycle. 2011;10(17):2830–2831. [PubMed] [Google Scholar]
- 117.Cairns RA, Harris IS, Mak TW, et al. Regulation of cancer metabolism. Nature Reviews Cancer. 2011;11:85–95. doi: 10.1038/nrc2981. [DOI] [PubMed] [Google Scholar]
- 118.Koppenol WH, Bounds PL, Dang CV. Otto Warburg's contributions to current concepts of cancer metabolism. Nature Reviews Cancer. 2011;11(5):325–337. doi: 10.1038/nrc3038. [DOI] [PubMed] [Google Scholar]
- 119.Marie SK, Shinjo SM. Metabolism and brain cancer. Clinics. 2011;66(supplement 1):33–43. doi: 10.1590/S1807-59322011001300005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Celi FS. Brown adipose tissue—when it pays to be inefficient. The New England Journal of Medicine. 2009;360(15):1553–1556. doi: 10.1056/NEJMe0900466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Fromme T, Klingenspor M. Uncoupling protein 1 expression and high-fat diets. American Journal of Physiology. 2011;300(1):R1–R8. doi: 10.1152/ajpregu.00411.2010. [DOI] [PubMed] [Google Scholar]
- 122.Jastroch M, Divakaruni AS, Mookerjee S, Treberg JR, Brand MD. Mitochondiral proton and electron leaks. Biochemistry. 2010;47:53–67. doi: 10.1042/bse0470053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Onat A. Metabolic syndrome: nature, therapeutic solutions and options. Expert Opinion on Pharmacotherapy. 2011;12(12):1887–1900. doi: 10.1517/14656566.2011.585462. [DOI] [PubMed] [Google Scholar]
- 124.Passamonti L, Crockett MJ, Apergis-Schoute AM, et al. Effects of acute tryptophan depletion on prefrontal-amygdala connectivity while viewing facial signals of aggression. Biological Psychiatry. 2012;71(1):36–43. doi: 10.1016/j.biopsych.2011.07.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Seal AJ, Creeke PI, Dibari F, et al. Low and deficient niacin status and pellagra are endemic in postwar Angola. American Journal of Clinical Nutrition. 2007;85(1):218–224. doi: 10.1093/ajcn/85.1.218. [DOI] [PubMed] [Google Scholar]
- 126.Etheridge EW. The Butterfly Castle: A Social History of Pellegra in the south. Connecticut, Conn, USA: Greenwood; 1972. [Google Scholar]
- 127.Kraut A. Goldberger’s War: The Life and Work of a Public Health Crusader. New York, NY, USA: Hill and Wang; 2003. [Google Scholar]
- 128.Cornell University. A Plague of Corn: The Social History of Pellagra. London, UK: Cornell University Press; 1973. [Google Scholar]
- 129.De Raadt LE. Pellagra in the Oto-Neurology and Rhino-Laryngology. Leiden, The Netherlands: Universitaire Pers Leiden; 1947. [Google Scholar]
- 130.Marie A. Pellagra. South Carolina, SC, USA: The State Company; 1910. [Google Scholar]
- 131.Terris M. Goldberger on Pellagra. New York Medical College, Louisiana State University; 1964. [Google Scholar]
- 132.Perdue EM. Pellagra in the United States. Kansas, Kan, USA: Burton; 1916. [Google Scholar]
- 133.Piro A, Tagarelli G, Lagonia P, Tagarelli A, Quattrone A. Casimir Funk: his discovery of the vitamins and their deficiency disorders. Annals of Nutrition and Metabolism. 2010;57(2):85–88. doi: 10.1159/000319165. [DOI] [PubMed] [Google Scholar]
- 134.Lanska DJ. Chapter 30 historical aspects of the major neurological vitamin deficiency disorders: the water-soluble B vitamins. Handbook of Clinical Neurology. 2009;95:445–476. doi: 10.1016/S0072-9752(08)02130-1. [DOI] [PubMed] [Google Scholar]
- 135.Weathers AL, Lewis SL. Rare and unusual....or are they? Less commonly diagnosed encephalopathies associated with systemic disease. Semin Neurol. 2009;29(2):136–153. doi: 10.1055/s-0029-1213734. [DOI] [PubMed] [Google Scholar]
- 136.Rajakumar K. Pellagra in the United States: a historical perspective. Southern Medical Journal. 2000;93(3):272–277. [PubMed] [Google Scholar]
- 137.Semba RD. Theophile roussel and the elimination of pellagra from 19th century France. Nutrition. 2000;16(3):231–233. doi: 10.1016/s0899-9007(99)00273-7. [DOI] [PubMed] [Google Scholar]
- 138.Wan P, Moat S, Anstey A. Pellagra: a review with emphasis on photosensitivity. British Journal of Dermatology. 2011;164(6):1188–1200. doi: 10.1111/j.1365-2133.2010.10163.x. [DOI] [PubMed] [Google Scholar]
- 139.Lombard DB, Pletcher SD. A valuable background check. Nature. 2011;477(7365):p. 410. doi: 10.1038/477410a. [DOI] [PubMed] [Google Scholar]
- 140.Burnett C, Valentini S, Cabreiro F, et al. Absence of effects of Sir2 overexpression on lifespan in C. elegans and Drosophila . Nature. 2011;477(7365):482–485. doi: 10.1038/nature10296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Couzin-Frankel J. Aging genes: the sirtuin story unravels. Science. 2011;334(6060):1194–1198. doi: 10.1126/science.334.6060.1194. [DOI] [PubMed] [Google Scholar]
- 142.Nisbett RE. Violence and U.S. regional culture. American Psychologist. 1993;48(4):441–449. doi: 10.1037//0003-066x.48.4.441. [DOI] [PubMed] [Google Scholar]
- 143.Lamoureux JA, Meck WH, Williams CL. Prenatal choline availability alters the context sensitivity of Pavlovian conditioning in adult rats. Learning and Memory. 2008;15(12):866–875. doi: 10.1101/lm.1058708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Meck WH, Williams CL, Cermak JM, Blusztajn JK. Developmental periods of choline sensitivity provide an ontogenetic mechanism for regulating memory capacity and age-related dementia. Frontiers in Integrative Neuroscience. 2007;1:7–12. doi: 10.3389/neuro.07.007.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Rafalski VA, Brunet A. Energy metabolism in adult neural stem cell fate. Progress in Neurobiology. 2010;93(2):182–203. doi: 10.1016/j.pneurobio.2010.10.007. [DOI] [PubMed] [Google Scholar]
- 146.Morris MC, Evans DA, Bienias JL, et al. Dietary niacin and the risk of incident Alzheimer's disease and of cognitive decline. Journal of Neurology, Neurosurgery and Psychiatry. 2004;75(8):1093–1099. doi: 10.1136/jnnp.2003.025858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Green KN, Steffan JS, Martinez-Coria H, et al. Nicotinamide restores cognition in Alzheimer’s disease transgenic mice via a mechanism involving sirtuin inhibition and selective reduction of Thr231-phosphotau. Journal of Neuroscience. 2008;28(45):11500–11510. doi: 10.1523/JNEUROSCI.3203-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Chan A, Tchantchou F, Rogers EJ, Shea TB. Dietary deficiency increases presenilin expression, gamma-secretase activity, and Abeta levels: potentiation by ApoE genotype and alleviation by S-adenosyl methionine. Journal of Neurochemistry. 2009;110(3):831–836. doi: 10.1111/j.1471-4159.2009.06177.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Chen J, Zhou Y, Mueller-Steiner S, et al. SIRT1 protects against microglia-dependent amyloid-β toxicity through inhibiting NF-κB signaling. Journal of Biological Chemistry. 2005;280(48):40364–40374. doi: 10.1074/jbc.M509329200. [DOI] [PubMed] [Google Scholar]
- 150.Adamczyk A, Jeśko H, Strosznajder RP. Alzheimer’s disease related peptides affected cholinergic receptor mediated poly(ADP-ribose) polymerase activity in the hippocampus. Folia Neuropathologica. 2005;43(3):139–142. [PubMed] [Google Scholar]
- 151.Longpre F, Garneau P, Christen Y, Ramassamy C. Protection by EGb 761 against beta-amyloid-induced neurotoxicity: involvement of NF-kappaB, SIRT1, and MAPKs pathways and inhibition of amyloid fibril formation. Free Radical Biology & Medicine. 2006;41(12):1781–1794. doi: 10.1016/j.freeradbiomed.2006.08.015. [DOI] [PubMed] [Google Scholar]
- 152.Julien C, Tremblay C, Émond V, et al. Sirtuin 1 reduction parallels the accumulation of tau in alzheimer disease. Journal of Neuropathology and Experimental Neurology. 2009;68(1):48–58. doi: 10.1097/NEN.0b013e3181922348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Kim D, Nguyen MD, Dobbin MM, et al. SIRT1 deacetylase protects against neurodegeneration in models for Alzheimer’s disease and amyotrophic lateral sclerosis. The EMBO Journal. 2007;26(13):3169–3179. doi: 10.1038/sj.emboj.7601758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Fischer A, Sananbenesi F, Wang XY, Dobbin M, Tsai LH. Recovery of learning and memory is associated with chromatin remodelling. Nature. 2007;447(7141):178–182. doi: 10.1038/nature05772. [DOI] [PubMed] [Google Scholar]
- 155.Gao J, Wang WY, Mao YW, et al. A novel pathway regulates memory and plasticity via SIRT1 and miR-134. Nature. 2010;466(7310):1105–1109. doi: 10.1038/nature09271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Winder R, Borrill J. Fuels for memory: the role of oxygen and glucose in memory enhancement. Psychopharmacology. 1998;136(4):349–356. doi: 10.1007/s002130050577. [DOI] [PubMed] [Google Scholar]
- 157.Anstey KJ, Cherbuin N, Budge M, Young J. Body mass index in midlife and late-life as a risk factor for dementia: a meta-analysis of prospective studies. Obesity Reviews. 2011;12(501):e426–e437. doi: 10.1111/j.1467-789X.2010.00825.x. [DOI] [PubMed] [Google Scholar]
- 158.Filley CM, Anderson CA. Dementia: five new things. Neurology. 2011;76(7, supplement 2):S26–S30. doi: 10.1212/WNL.0b013e31820c35e1. [DOI] [PubMed] [Google Scholar]
- 159.García-Lara JM, Aguilar-Navarro S, Gutiérrez-Robledo LM, Ávila-Funes JA. The metabolic syndrome, diabetes, and Alzheimer's disease. Revista de Investigacion Clinica. 2010;62(4):343–349. [PubMed] [Google Scholar]
- 160.Veeraraghavalu K, Choi SH, Zhang X, Sisodia SS. Presenilin 1 mutants impair the self-renewal and differentiation of adult murine subventricular zone-neuronal progenitors via cell-autonomous mechanisms involving notch signaling. Journal of Neuroscience. 2010;30(20):6903–6915. doi: 10.1523/JNEUROSCI.0527-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Lee JH, Yu WH, Kumar A, et al. Lysosomal proteolysis and autophagy require presenilin 1 and are disrupted by Alzheimer-related PS1 mutations. Cell. 2010;141(7):1146–1158. doi: 10.1016/j.cell.2010.05.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Abeti R, Abramov AY, Duchen MR. β-amyloid activates PARP causing astrocytic metabolic failure and neuronal death. Brain. 2011;134(6):1658–1672. doi: 10.1093/brain/awr104. [DOI] [PubMed] [Google Scholar]
- 163.Fuso A, Scarpa S. One-carbon metabolism and Alzheimer’s disease: is it all a methylation matter? Neurobiology of Aging. 2011;32(7):1192–1195. doi: 10.1016/j.neurobiolaging.2011.01.012. [DOI] [PubMed] [Google Scholar]
- 164.Koh P-O. Nicotinamide attenuates the ischemic brain injury-induced decrease of Akt activation and Bad phosphorylation. Neuroscience Letters. 2011;498(2):105–109. doi: 10.1016/j.neulet.2011.05.003. [DOI] [PubMed] [Google Scholar]
- 165.Wen Y, Parrish JZ, He R, Zhai RG, Kim MD. Nmnat exerts neuroprotective effects in dendrites and axons. Molecular and Cellular Neuroscience. 2011;48(1):1–8. doi: 10.1016/j.mcn.2011.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Haar CV, Anderson GD, Hoane MR. Continuous nicotinamide administration improves behavioral recovery and reduces lesion size following bilateral frontal controlled cortical impact injury. Behavioural Brain Research. 2011;224(2):311–317. doi: 10.1016/j.bbr.2011.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Middleton LE, Manini TM, Simonsick EM, et al. Activity energy expenditure and incident cognitive impairment in older adults. Archives of Internal Medicine. 2011;171(14):1251–1257. doi: 10.1001/archinternmed.2011.277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Colman RJ, Anderson RM, Johnson SC, et al. Caloric restriction delays disease onset and mortality in rhesus monkeys. Science. 2009;325(5937):201–204. doi: 10.1126/science.1173635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Hathorn T, Snyder-Keller A, Messer A. Nicotinamide improves motor deficits and upregulates PGC-1α and BDNF gene expression in a mouse model of Huntington’s disease. Neurobiology of Disease. 2011;41(1):43–50. doi: 10.1016/j.nbd.2010.08.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Kirkland JB. Niacin status impacts chromatin structure. Journal of Nutrition. 2009;139(12):2397–2401. doi: 10.3945/jn.109.111757. [DOI] [PubMed] [Google Scholar]
- 171.Klawitter V, Morales P, Bustamante D, Goiny M, Herrera-Marschitz M. Plasticity of the central nervous system (CNS) following perinatal asphyxia: does nicotinamide provide neuroprotection? Amino Acids. 2006;31(4):377–384. doi: 10.1007/s00726-006-0372-4. [DOI] [PubMed] [Google Scholar]
- 172.Liu D, Gharavi R, Pitta M, Gleichmann M, Mattson MP. Nicotinamide prevents NAD+ depletion and protects neurons against excitotoxicity and cerebral ischemia: NAD+ consumption by sirt1 may endanger energetically compromised neurons. NeuroMolecular Medicine. 2009;11(1):28–42. doi: 10.1007/s12017-009-8058-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Maiese K. Triple play: promoting neurovascular longevity with nicotinamide, WNT, and erythropoietin in diabetes mellitus. Biomedicine and Pharmacotherapy. 2008;62(4):218–232. doi: 10.1016/j.biopha.2008.01.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Kaneko S, Wang J, Kaneko M, et al. Protecting axonal degeneration by increasing nicotinamide adenine dinucleotide levels in experimental autoimmune encephalomyelitis models. Journal of Neuroscience. 2006;26(38):9794–9804. doi: 10.1523/JNEUROSCI.2116-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Du H, Guo L, Yan S, Sosunov AA, McKhann GM, Yan SS. Early deficits in synaptic mitochondria in an Alzheimer’s disease mouse model. Proceedings of the National Academy of Sciences of the United States of America. 2010;107(43):18670–18675. doi: 10.1073/pnas.1006586107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Drenos F, Kirkwood TBL. Selection on alleles affecting human longevity and late-life disease: the example of apolipoprotein E. Plos ONE. 2010;5(4) doi: 10.1371/journal.pone.0010022. Article ID e10022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Mastroeni D, Grover A, Delvaux E, Whiteside C, Coleman PD, Rogers J. Epigenetic mechanisms in Alzheimer's disease. Neurobiology of Aging. 2011;32(7):1161–1180. doi: 10.1016/j.neurobiolaging.2010.08.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Milosavljevic A. Emerging patterns of epigenomic variation. Trends in Genetics. 2011;27(6):242–250. doi: 10.1016/j.tig.2011.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Ali YO, Ruan K, Zhai RG. NMNAT suppresses Tau-induced neurodegeneration by promoting clearance of hyperphosphorylated Tau oligomers in a Drosophila model of tauopathy. doi: 10.1093/hmg/ddr449. Human Molecular Genetics. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Zhang LQ, Heruth DP, Ye SQ. Nicotinamide phosphoribosyltransferase in human diseases. Journal of Bioanalysis and Biomedicine. 2011;3(1):13–25. doi: 10.4172/1948-593X.1000038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Garten A, Petzold S, Schuster S, Körner A, Kratzsch J, Kiess W. Nampt and its potential role in inflammation and type 2 diabetes. Handbook of Experimental Pharmacology. 2011;203:147–164. doi: 10.1007/978-3-642-17214-4_7. [DOI] [PubMed] [Google Scholar]
- 182.Wang P, Guan YF, Du H, Zhai QW, Su DF, Miao CY. Induction of autophagy contributes to the neuroprotection of nicotinamide phosphoribosyltransferase in cerebral ischemia. Autophagy. 2012;8(1) doi: 10.4161/auto.8.1.18274. [DOI] [PubMed] [Google Scholar]
- 183.Murray MF. Tryptophan depletion and HIV infection: a metabolic link to pathogenesis. Lancet Infectious Diseases. 2003;3(10):644–652. doi: 10.1016/s1473-3099(03)00773-4. [DOI] [PubMed] [Google Scholar]
- 184.Rozwarski DA, Grant GA, Barton DH, Jacobs WR, Jr., Sacchettini JC. Modification of the NADH of the isoniazid target (InhA) from Mycobacterium tuberculosis. Science. 1998;279(5347):98–102. doi: 10.1126/science.279.5347.98. [DOI] [PubMed] [Google Scholar]
- 185.Aksoy P, White TA, Thompson M, Chini EN. Regulation of intracellular levels of NAD: a novel role for CD38. Biochemical and Biophysical Research Communications. 2006;345(4):1386–1392. doi: 10.1016/j.bbrc.2006.05.042. [DOI] [PubMed] [Google Scholar]
- 186.Zhang HS, Wu MR. SIRT1 regulates Tat-induced HIV-1 transactivation through activating AMP-activated protein kinase. Virus Research. 2009;146(1-2):51–57. doi: 10.1016/j.virusres.2009.08.005. [DOI] [PubMed] [Google Scholar]
- 187.Grahnert A, Grahnert A, Klein C, Schilling E, Wehrhahn J, Hauschildt S. NAD+: a modulator of immune functions. Innate Immunity. 2011;17(2):212–223. doi: 10.1177/1753425910361989. [DOI] [PubMed] [Google Scholar]
- 188.Moëll A, Skog O, Åhlin E, Korsgren O, Frisk G. Antiviral effect of nicotinamide on enterovirus-infected human islets in vitro: effect on virus replication and chemokine secretion. Journal of Medical Virology. 2009;81(6):1082–1087. doi: 10.1002/jmv.21476. [DOI] [PubMed] [Google Scholar]
- 189.Orabona C, Grohmann U. Indoleamine 2,3-dioxygenase and regulatory function: tryptophan starvation and beyond. Methods in Molecular Biology. 2011;677:269–280. doi: 10.1007/978-1-60761-869-0_19. [DOI] [PubMed] [Google Scholar]
- 190.Tracey KJ. Cell biology. Ancient neurons regulate immunity. Science. 2011;332:673–674. doi: 10.1126/science.1206353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Hansen JD, Vojtech LN, Laing KJ. Sensing disease and danger: a survey of vertebrate PRRs and their origins. Developmental and Comparative Immunology. 2011;35(9):886–897. doi: 10.1016/j.dci.2011.01.008. [DOI] [PubMed] [Google Scholar]
- 192.Burnstock G, Ulrich H. Purinergic signaling in embryonic and stem cell development. Cellular and Molecular Life Sciences. 2011;68(8):1369–1394. doi: 10.1007/s00018-010-0614-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Février M, Dorgham K, Rebollo A. CD4+T cell depletion in human immunodeficiency virus (HIV) infection: role of apoptosis. Viruses. 2011;3(5):586–612. doi: 10.3390/v3050586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Xu H, Zhang GX, Ciric B, Rostami A. IDO: a double-edged sword for TH1/TH2 regulation. Immunology Letters. 2008;121(1):1–6. doi: 10.1016/j.imlet.2008.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Stone TW, Forrest CM, Darlington LG. Kynurenine pathway inhibition as a therapeutic strategy for neuroprotection. doi: 10.1111/j.1742-4658.2012.08487.x. The FEBS Journal. In press. [DOI] [PubMed] [Google Scholar]
- 196.Raubenheimer D, Lee KP, Simpson SJ. Does Bertrand’s rule apply to macronutrients? Proceedings of the Royal Society B. 2005;272(1579):2429–2434. doi: 10.1098/rspb.2005.3271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.de Kreutzenberg SV, Ceolotto G, Papparella I, et al. Downregulation of the longevity-associated protein SIRT1 in insulin resistance and metabolic syndrome. Potential biochemical mechanisms. Diabetes. 2010;59(4):1006–1015. doi: 10.2337/db09-1187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Hollis G, Huber R. 11β-hydroxysteroid dehydrogenase type 1 inhibition in type 2 diabetes mellitus. Diabetes, Obesity and Metabolism. 2011;13(1):1–6. doi: 10.1111/j.1463-1326.2010.01305.x. [DOI] [PubMed] [Google Scholar]
- 199.Pang J, Gong H, Xi C, Fan W, Dai Y, Zhang T-M. Poly(ADP-ribose) polymerase 1 is involved in glucose toxicity through SIRT1 modulation in HepG2 hepatocytes. Journal of Cellular Biochemistry. 2011;112(1):299–306. doi: 10.1002/jcb.22919. [DOI] [PubMed] [Google Scholar]
- 200.Zeisel SH. Nutritional genomics: defining the dietary requirement and effects of choline. Journal of Nutrition. 2011;141(3):531–534. doi: 10.3945/jn.110.130369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.West AA, Caudill MA. Genetic variation: impact on folate (and choline) bioefficacy. International Journal for Vitamin and Nutrition Research. 2010;80(4-5):319–329. doi: 10.1024/0300-9831/a000040. [DOI] [PubMed] [Google Scholar]
- 202.Wang Z, Klipfell E, Bennett BJ, et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature. 2011;472(7341):57–65. doi: 10.1038/nature09922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Yamamoto K, Vernier P. The evolution of dopamine systems in chordates. Frontiers in Neuroanatomy. 2011;5(21) doi: 10.3389/fnana.2011.00021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Edwards S, Self DW. Monogamy: dopamine ties the knot. Nature Neuroscience. 2006;9(1):7–8. doi: 10.1038/nn0106-7. [DOI] [PubMed] [Google Scholar]
- 205.Bromberg-Martin ES, Hikosaka O. Midbrain dopamine neurons signal preference for advance information about upcoming rewards. Neuron. 2009;63(1):119–126. doi: 10.1016/j.neuron.2009.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Fliessbach K, Weber B, Trautner P, et al. Social comparison affects reward-related brain activity in the human ventral striatum. Science. 2007;318(5854):1305–1308. doi: 10.1126/science.1145876. [DOI] [PubMed] [Google Scholar]
- 207.Previc FH. Dopamine and the origins of human intelligence. Brain and Cognition. 1999;41(3):299–350. doi: 10.1006/brcg.1999.1129. [DOI] [PubMed] [Google Scholar]
- 208.Rodrigues SM, Saslow LR, Garcia N, John OP, Keltner D. Oxytocin receptor genetic variation relates to empathy and stress reactivity in humans. Proceedings of the National Academy of Sciences of the United States of America. 2009;106(50):21437–21441. doi: 10.1073/pnas.0909579106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Higashida H, Lopatina O, Yoshihara T, et al. Oxytocin signal and social behaviour: comparison amoung adult and infant oxytocin, oxytocin receptor and CD38 gene knockout mice. Journal of Neuroendocrinology. 2010;22(5):373–379. doi: 10.1111/j.1365-2826.2010.01976.x. [DOI] [PubMed] [Google Scholar]
- 210.Flinn MV, Nepomnaschy PA, Muehlenbein MP, Ponzi D. Evolutionary functions of early social modulation of hypothalamic-pituitary-adrenal axis development in humans. Neuroscience and Biobehavioral Reviews. 2011;35(7):1611–1629. doi: 10.1016/j.neubiorev.2011.01.005. [DOI] [PubMed] [Google Scholar]
- 211.Higashida H. Biological basis for social recognition: CD38 controls oxytocin secretion. Tanpakushitsu Kakusan Koso. 2007;52(14):1832–1839. [PubMed] [Google Scholar]
- 212.Sherwood CC, Gordon AD, Allen JS, et al. Aging of the cerebral cortex differs between humans and chimpanzees. Proceedings of the National Academy of Sciences of the United States of America. 2011;108(32):13029–13034. doi: 10.1073/pnas.1016709108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Trinkaus E. Late Pleistocene adult mortality patterns and modern human establishment. Proceedings of the National Academy of Sciences of the United States of America. 2011;108(4):1267–1271. doi: 10.1073/pnas.1018700108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Shahrestani P, Mueller LD, Rose MR. Does aging stop? Current Aging Science. 2009;2(1):3–11. doi: 10.2174/1874609810902010003. [DOI] [PubMed] [Google Scholar]
- 215.Bronikowski AM, Altmann J, Brockman DK, et al. Aging in the natural world: comparative data reveal similar mortality patterns across primates. Science. 2011;331(6022):1325–1328. doi: 10.1126/science.1201571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Stiner MC, Barkai R, Gopher A. Cooperative hunting and meat sharing 400–200 kya at Qesem Cave, Israel. Proceedings of the National Academy of Sciences of the United States of America. 2009;106(32):13207–13212. doi: 10.1073/pnas.0900564106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Northcutt RG. Evolving large and complex brains. Science. 2011;332(6032):926–927. doi: 10.1126/science.1206915. [DOI] [PubMed] [Google Scholar]
- 218.Sharma Y, Xu T, Graf WM, et al. Comparative anatomy of the locus coeruleus in humans and nonhuman primates. Journal of Comparative Neurology. 2010;518(7):963–971. doi: 10.1002/cne.22249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Raghanti MA, Stimpson CD, Marcinkiewicz JL, Erwin JM, Hof PR, Sherwood CC. Cortical dopaminergic innervation among humans, chimpanzees, and macaque monkeys: a comparative study. Neuroscience. 2008;155(1):203–220. doi: 10.1016/j.neuroscience.2008.05.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Ungar PS, Sponheimer M. The diets of early hominins. Science. 2011;334(6053):190–193. doi: 10.1126/science.1207701. [DOI] [PubMed] [Google Scholar]
- 221.Vinot N, Jouin M, Lhomme-Duchadeuil A, et al. Omega-3 fatty acids from fish oil lower anxiety, improve cognitive functions and reduce spontaneous locomotor activity in a non-human primate. Plos ONE. 2011;6(6) doi: 10.1371/journal.pone.0020491. Article ID e20491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222.Robinson GE. Entomology: royal aspirations. Nature. 2011;473(7348):454–455. doi: 10.1038/473454a. [DOI] [PubMed] [Google Scholar]
- 223.Pontzer H, Kamilar JM. Great ranging associated with greater reproductive investment in mammals. Proceedings of the National Academy of Sciences of the United States of America. 2009;106(1):192–196. doi: 10.1073/pnas.0806105106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Bar M. Wait for the second marshmallow? Future-oriented thinking and delayed reward discounting in the brain. Neuron. 2010;66(1):4–5. doi: 10.1016/j.neuron.2010.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Krach S, Paulus FM, Bodden M, Kircher T. The rewarding nature of social interactions. Frontiers in Neuroscience. 2010;4(22) doi: 10.3389/fnbeh.2010.00022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226.Sterelny K. From hominins to humans: how sapiens became behaviourally modern. Philosophical Transactions of the Royal Society of London B. 2011;366(1566):809–822. doi: 10.1098/rstb.2010.0301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Rendell L, Fogarty L, Laland KN. Runaway cultural niche construction. Philosophical transactions of the Royal Society of London B. 2011;366(1566):823–835. doi: 10.1098/rstb.2010.0256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.Cabanac M. Selective brain cooling in humans: “Fancy” or fact? The FASEB Journal. 1993;7(12):1143–1147. doi: 10.1096/fasebj.7.12.8375612. [DOI] [PubMed] [Google Scholar]
- 229.Ponce de León MS, Golovanova L, Doronichev V, et al. Neanderthal brain size at birth provides insights into the evolution of human life history. Proceedings of the National Academy of Sciences of the United States of America. 2008;105(37):13764–13768. doi: 10.1073/pnas.0803917105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Richards MP, Trinkaus E. Out of Africa: modern human origins special feature: isotopic evidence for the diets of European Neanderthals and early modern humans. Proceedings of the National Academy of Sciences of the United States of America. 2009;106:16034–16039. doi: 10.1073/pnas.0903821106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 231.Campbell MC, Tishkoff SA. The evolution of human genetic and phenotypic variation in Africa. Current Biology. 2010;20(4):R166–R173. doi: 10.1016/j.cub.2009.11.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Konopka G, Bomar JM, Winden K, et al. Human-specific transcriptional regulation of CNS development genes by FOXP2. Nature. 2009;462(7270):213–217. doi: 10.1038/nature08549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.Schwander T, Leimar O. Genes as leaders and followers in evolution. Trends in Ecology and Evolution. 2011;26(3):143–151. doi: 10.1016/j.tree.2010.12.010. [DOI] [PubMed] [Google Scholar]
- 234.Evans PD, Gilbert SL, Mekel-Bobrov N, et al. Microcephalin, a gene regulating brain size, continues to evolve adaptively in humans. Science. 2005;309(5741):1717–1720. doi: 10.1126/science.1113722. [DOI] [PubMed] [Google Scholar]
- 235.Mekel-Bobrov N, Gilbert SL, Evans PD, et al. Ongoing adaptive evolution of ASPM, a brain size determinant in Homo sapiens. Science. 2005;309(5741):1720–1722. doi: 10.1126/science.1116815. [DOI] [PubMed] [Google Scholar]
- 236.Cerling TE, Wynn JG, Andanje SA, et al. Woody cover and hominin environments in the past 6-million years. Nature. 2011;476(7358):51–56. doi: 10.1038/nature10306. [DOI] [PubMed] [Google Scholar]
- 237.Norenzayan A. Explaining human behavioral diversity. Science. 2011;332(6033):1041–1042. doi: 10.1126/science.1207050. [DOI] [PubMed] [Google Scholar]
- 238.Gold PE, Countryman RA, Dukala D, Chang Q. Acetylcholine release in the hippocampus and prelimbic cortex during acquisition of a socially transmitted food preference. Neurobiology of Learning and Memory. 2011;96(3):498–503. doi: 10.1016/j.nlm.2011.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 239.Sherman PW, Billing J. Darwinian gastronomy: why we use spices. BioScience. 1999;49(6):453–463. [Google Scholar]
- 240.Carmody RN, Wrangham RW. The energetic significance of cooking. Journal of Human Evolution. 2009;57(4):379–391. doi: 10.1016/j.jhevol.2009.02.011. [DOI] [PubMed] [Google Scholar]
- 241.Mellars P. Fire ecology, animal populations and man: a study of some ecological relationships in prehistory. The Proceedings of the Prehistoric Society. 1976;42:15–45. [Google Scholar]
- 242.Ryan B, Gross NC. The diffusion of hybrid seed corn in two iowa communities. Rural Sociology. 1943;8(1):15–24. [Google Scholar]
- 243.Mummert A, Esche E, Robinson J, Armelagos GJ. Stature and robusticity during the agricultural transition: evidence from the bioarchaeological record. Economics and Human Biology. 2011;9(3):284–301. doi: 10.1016/j.ehb.2011.03.004. [DOI] [PubMed] [Google Scholar]
- 244.Fogal RW. The Escape from Hunger and Premature Death, 1700–2100 Europe, America and the Third World. Cambridge, UK: Cambridge University Press; 2004. [Google Scholar]
- 245.Bocquet-Appel JP. When the world's population took off: the springboard of the neolithic demographic transition. Science. 2011;333(6042):560–561. doi: 10.1126/science.1208880. [DOI] [PubMed] [Google Scholar]
- 246.Smalley J, Blake M. Sweet beginnings: stalk sugar and the domestication of maize. Current Anthropology. 2003;44(5):675–703. [Google Scholar]
- 247.Lin SJ, Guarente L. Nicotinamide adenine dinucleotide, a metabolic regulator of transcription, longevity and disease. Current Opinion in Cell Biology. 2003;15(2):241–246. doi: 10.1016/s0955-0674(03)00006-1. [DOI] [PubMed] [Google Scholar]
- 248.Smillie CS, Smith MB, Friedman J, Cordero OX, David LA, Alm EJ. Ecology drives a global network of gene exchange connecting the human microbiome. Nature. 2011;480(7376):241–244. doi: 10.1038/nature10571. [DOI] [PubMed] [Google Scholar]
- 249.Wang XL, Takai T, Kamijo S, Gunawan H, Ogawa H, Okumura K. NADPH oxidase activity in allergenic pollen grains of different plant species. Biochemical and Biophysical Research Communications. 2009;387(3):430–434. doi: 10.1016/j.bbrc.2009.07.020. [DOI] [PubMed] [Google Scholar]
- 250.Lomax AR, Calder PC. Prebiotics, immune function, infection and inflammation: a review of the evidence. British Journal of Nutrition. 2009;101(5):633–658. doi: 10.1017/S0007114508055608. [DOI] [PubMed] [Google Scholar]
- 251.Rook GA. Hygiene and other early childhood influences on the subsequent function of the immune system. Digestive Diseases. 2011;29(2):144–153. doi: 10.1159/000323877. [DOI] [PubMed] [Google Scholar]
- 252.Palacios OM, Carmona JJ, Michan S, et al. Diet and exercise signals regulate SIRT3 and activate AMPK and PGC-1alpha in skeletal muscle. Aging. 2009;1(9):771–783. doi: 10.18632/aging.100075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253.Fessler DM. Reproductive immunosupression and diet. An evolutionary perpective on pregnancy sickness and meat consumption. Current Anthropology. 2002;42(1):19–61. doi: 10.1086/324128. [DOI] [PubMed] [Google Scholar]
- 254.Daul AM, Beuhler MC. Niacin toxicity resulting from urine drug test evasion scheme. Journal of Emergency Medicine. 2011;41(3):e65–e68. doi: 10.1016/j.jemermed.2009.11.029. [DOI] [PubMed] [Google Scholar]
- 255.Tang S-W, Yang T-C, Lin W-C, et al. Nicotinamide N-methyltransferase induces cellular invasion through activating matrix metalloproteinase-2 expression in clear cell renal cell carcinoma cells. Carcinogenesis. 2011;32(2):138–145. doi: 10.1093/carcin/bgq225. [DOI] [PubMed] [Google Scholar]
- 256.Emanuelli M, Santarelli A, Sartini D, et al. Nicotinamide N-Methyltransferase upregulation correlates with tumour differentiation in oral squamous cell carcinoma. Histology and Histopathology. 2010;25(1):15–20. doi: 10.14670/HH-25.15. [DOI] [PubMed] [Google Scholar]
- 257.Yap IK, Angley M, Veselkov KA, Holmes E, Lindon JC, Nicholson JK. Urinary metabolic phenotyping differentiates children with autism from their unaffected siblings and age-matched controls. Journal of Proteome Research. 2010;9(6):2996–3004. doi: 10.1021/pr901188e. [DOI] [PubMed] [Google Scholar]
- 258.Riederer M, Erwa W, Zimmermann R, Frank S, Zechner R. Adipose tissue as a source of nicotinamide N-methyltransferase and homocysteine. Atherosclerosis. 2009;204(2):412–417. doi: 10.1016/j.atherosclerosis.2008.09.015. [DOI] [PubMed] [Google Scholar]
- 259.van Driel LM, Smedts HP, Helbing WA, et al. Eight-fold increased risk for congenital heart defects in children carrying the nicotinamide N-methyltransferase polymorphism and exposed to medicines and low nicotinamide. European Heart Journal. 2008;29(11):1424–1431. doi: 10.1093/eurheartj/ehn170. [DOI] [PubMed] [Google Scholar]
- 260.Zhang L, Miyaki K, Araki J, Nakayama T, Muramatsu M. The relation between nicotinamide N-methyltransferase gene polymorphism and plasma homocysteine concentration in healthy Japanese men. Thrombosis Research. 2007;121(1):55–58. doi: 10.1016/j.thromres.2007.02.012. [DOI] [PubMed] [Google Scholar]
- 261.Parsons RB, Smith M-L, Williams AC, Waring RH, Ramsden DB. Expression of nicotinamide N-methyltransferase (E.C. 2.1.1.1) in the Parkinsonian brain. Journal of Neuropathology and Experimental Neurology. 2002;61(2):111–124. doi: 10.1093/jnen/61.2.111. [DOI] [PubMed] [Google Scholar]
- 262.Pozzi V, Mazzotta M, Lo Muzio L, et al. Inhibiting proliferation in KB cancer cells by RNA interference-medicated knockdown of nicotinamide n-methyitransferase expression. International Journal of Immunopathology and Pharmacology. 2011;24:69–77. doi: 10.1177/039463201102400109. [DOI] [PubMed] [Google Scholar]
- 263.Lee JY, Ahn K, Jang BG, et al. Nicotinamide reduces dopamine in postnatal hypothalamus and causes dopamine-deficient phenotype. Neuroscience Letters. 2009;461(2):163–166. doi: 10.1016/j.neulet.2009.06.005. [DOI] [PubMed] [Google Scholar]
- 264.Bisaglia M, Soriano ME, Arduini I, Mammi S, Bubacco L. Molecular characterization of dopamine-derived quinines reactivity toward NADH and glutathione: implications for mitochondrial dysfunction in Parkinson’s disease. Biochimica et Biophysica Acta. 2010;1802(9):699–706. doi: 10.1016/j.bbadis.2010.06.006. [DOI] [PubMed] [Google Scholar]
- 265.Kim B, Kim JE, Lee SM, et al. N-Nicotinoyl dopamine, a novel niacinamide derivative, retains high antioxidant activity and inhibits skin pigmentation. Experimental Dermatology. 2011;20(11):950–952. doi: 10.1111/j.1600-0625.2011.01345.x. [DOI] [PubMed] [Google Scholar]
- 266.Chan A, Tchantchou F, Graves V, Rozen R, Shea TB. Dietary and genetic compromise in folate availability reduces acetylcholine, cognitive performance and increases aggression: critical role of S-adenosyl methionine. Journal of Nutrition, Health and Aging. 2008;12(4):252–261. doi: 10.1007/BF02982630. [DOI] [PubMed] [Google Scholar]
- 267.Bosco DA, Lavoie MJ, Petsko GA, Ringe D. Proteostasis and movement disorders: parkinson’s disease amyotrophic lateral sclerosis. Cold Spring Harbor Perspectives in Biology. 2011;3(10) doi: 10.1101/cshperspect.a007500. Article ID a007500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 268.Carroll JS, Munchel SE, Weis K, et al. The DExD/H box ATPase Dhh1 functions in translational repsression, mRNA decay, and processing body dynamics. The Journal of Cell Biology. 2011;194:527–537. doi: 10.1083/jcb.201007151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 269.Batey RT. Recognition of S-adenosylmethionine by riboswitches. Wiley Interdisciplinary Reviews RNA. 2011;2(2):299–311. doi: 10.1002/wrna.63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 270.Szyf M. The implications of DNA methylation for toxicology: toward toxicomethylomics, the toxicology of DNA methylation. Toxicological Sciences. 2011;120(2):235–255. doi: 10.1093/toxsci/kfr024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 271.Waring RH, Steventon GB, Sturman SG, Heafield MT, Smith MC, Williams AC. S-methylation in motorneuron disease and Parkinson’s disease. The Lancet. 1989;2(8659):356–357. doi: 10.1016/s0140-6736(89)90538-2. [DOI] [PubMed] [Google Scholar]
- 272.Buratti E. TDP-43 and FUS in ALS/FTLD: will common pathways fit all? Neurology. 2011;77(17):1588–1589. doi: 10.1212/WNL.0b013e31823433ba. [DOI] [PubMed] [Google Scholar]
- 273.Hitchins MP. Inheritance of epigenetic aberrations (constitutional epimutations) in cancer susceptibility. Advances in Genetics. 2010;70:201–243. doi: 10.1016/B978-0-12-380866-0.60008-3. [DOI] [PubMed] [Google Scholar]
- 274.Krutovskikh V, Partensky C. Nouvelles perspectives en oncologie: epigénétique et cellules souches cancéreuses. Cancer Radiotherapie. 2011;15(8):716–722. doi: 10.1016/j.canrad.2011.05.012. [DOI] [PubMed] [Google Scholar]
- 275.Song S-H, Han S-W, Bang Y-J. Epigenetic-based therapies in cancer: progress to date. Drugs. 2011;71(18):2391–2403. doi: 10.2165/11596690-000000000-00000. [DOI] [PubMed] [Google Scholar]
- 276.Vel Szic KS, Ndlovu MN, Haegeman G, Vanden Berghe W. Nature or nurture: let food be your epigenetic medicine in chronic inflammatory disorders. Biochemical Pharmacology. 2010;80(12):1816–1832. doi: 10.1016/j.bcp.2010.07.029. [DOI] [PubMed] [Google Scholar]
- 277.Martin DI, Cropley JE, Suter CM. Epigenetics in disease: leader or follower? Epigenetics. 2011;6(7):843–848. doi: 10.4161/epi.6.7.16498. [DOI] [PubMed] [Google Scholar]
- 278.Ng SF, Lin RC, Laybutt DR, Barres R, Owens JA, Morris MJ. Chronic high-fat diet in fathers programs β 2-cell dysfunction in female rat offspring. Nature. 2010;467(7318):963–966. doi: 10.1038/nature09491. [DOI] [PubMed] [Google Scholar]
- 279.Margulis L, Dolan MF, Guerrero R. The chimeric eukaryote: origin of the nucleus from the karyomastigont in amitochondriate protists. Proceedings of the National Academy of Sciences of the United States of America. 2000;97(13):6954–6959. doi: 10.1073/pnas.97.13.6954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 280.Martin W, Müller M. The hydrogen hypothesis for the first eukaryote. Nature. 1998;392(6671):37–41. doi: 10.1038/32096. [DOI] [PubMed] [Google Scholar]
- 281.Orphan VJ, Hoehler TM. Microbiology: hydrogen for dinner. Nature. 2011;476:154–155. doi: 10.1038/476154a. [DOI] [PubMed] [Google Scholar]
- 282.Belenky P, Racette FG, Bogan KL, McClure JM, Smith JS, Brenner C. Nicotinamide riboside promotes sir2 silencing and extends lifespan via nrk and Urh1/Pnp1/Meu1 pathways to NAD+ . Cell. 2007;129(3):473–484. doi: 10.1016/j.cell.2007.03.024. [DOI] [PubMed] [Google Scholar]
- 283.Mentel M, Martin W. Energy metabolism among eukaryotic anaerobes in light of Proterozoic ocean chemistry. Philosophical Transactions of the Royal Society B. 2008;363(1504):2717–2729. doi: 10.1098/rstb.2008.0031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 284.Anderson RM, Bitterman KJ, Wood JG, Medvedik O, Sinclair DA. Nicatinamide and PNC1 govern lifespan extension by calorie restriction in Saccharomyces cerevisiae. Nature. 2003;423(6936):181–185. doi: 10.1038/nature01578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 285.Lu SP, Kato M, Lin SJ. Assimilation of endogenous nicotinamide riboside is essential for calorie restriction-mediated life span extension in Saccharomyces cerevisiae. Journal of Biological Chemistry. 2009;284(25):17110–17119. doi: 10.1074/jbc.M109.004010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 286.Kasting JF. Runaway and moist greenhouse atmospheres and the evolution of Earth and Venus. Icarus. 1988;74(3):472–494. doi: 10.1016/0019-1035(88)90116-9. [DOI] [PubMed] [Google Scholar]
- 287.Shinmura K, Tamaki K, Sano M, et al. Caloric restriction primes mitochondria for ischemic stress by deactelyating specific mitochondrial proteins of the electron transport chain. Circulation Research. 2011;109(4):396–406. doi: 10.1161/CIRCRESAHA.111.243097. [DOI] [PubMed] [Google Scholar]
- 288.Bitterman KJ, Anderson RM, Cohen HY, Latorre-Esteves M, Sinclair DA. Inhibition of silencing and accelerated aging by nicotinamide, a putative negative regulator of yeast Sir2 and human SIRT1. Journal of Biological Chemistry. 2002;277(47):45099–45107. doi: 10.1074/jbc.M205670200. [DOI] [PubMed] [Google Scholar]
- 289.Estep PW, III, Warner JB, Bulyk ML. Short-term calorie restriction in male mice feminizes gene expression and alters key regulators of conserved aging regulatory pathways. Plos ONE. 2009;4(4) doi: 10.1371/journal.pone.0005242. Article ID e5242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 290.Calabrese EJ, Mattson MP, Calabrese V. Dose response biology: the case of resveratrol. Human and Experimental Toxicology. 2010;29(12):1034–1037. doi: 10.1177/0960327110383641. [DOI] [PubMed] [Google Scholar]
- 291.De Petrocellis L, Di Marzo V. Cell signalling: why fasting worms age slowly. Nature. 2011;473(7346):161–163. doi: 10.1038/473161a. [DOI] [PubMed] [Google Scholar]
- 292.Rodgers JT, Lerin C, Haas W, Gygi SP, Spiegelman BM, Puigserver P. Nutrient control of glucose homeostasis through a complex of PGC-1α and SIRT1. Nature. 2005;434(7029):113–118. doi: 10.1038/nature03354. [DOI] [PubMed] [Google Scholar]
- 293.Chen WY, Wang DH, RayWhay CY, Luo J, Gu W, Baylin SB. Tumor suppressor HIC1 directly regulates SIRT1 to modulate p53-dependent DNA-damage responses. Cell. 2005;123(3):437–448. doi: 10.1016/j.cell.2005.08.011. [DOI] [PubMed] [Google Scholar]
- 294.Mineur YS, Abizaid A, Rao Y, et al. Nicotine decreases food intake through activation of POMC neurons. Science. 2011;332(6035):1330–1332. doi: 10.1126/science.1201889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 295.Balan V, Miller GS, Kaplun L, et al. Life span extension and neuronal cell protection by Drosophila nicotinamidase. Journal of Biological Chemistry. 2008;283(41):27810–27819. doi: 10.1074/jbc.M804681200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 296.Suchankova G, Nelson LE, Gerhart-Hines Z, et al. Concurrent regulation of AMP-activated protein kinase and SIRT1 in mammalian cells. Biochemical and Biophysical Research Communications. 2009;378(4):836–841. doi: 10.1016/j.bbrc.2008.11.130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 297.Salminen A, Kaarniranta K. SIRT1: regulation of longevity via autophagy. Cellular Signalling. 2009;21(9):1356–1360. doi: 10.1016/j.cellsig.2009.02.014. [DOI] [PubMed] [Google Scholar]
- 298.Kang HT, Hwang ES. Nicotinamide enhances mitochondria quality through autophagy activation in human cells. Aging Cell. 2009;8(4):426–438. doi: 10.1111/j.1474-9726.2009.00487.x. [DOI] [PubMed] [Google Scholar]
- 299.Sasaki T, Kitamura T. Roles of FoxO1 and Sirt1 in the central regulation of food intake. Endocrine Journal. 2010;57(11):939–946. doi: 10.1507/endocrj.k10e-320. [DOI] [PubMed] [Google Scholar]
- 300.Satoh A, Brace CS, Ben-Josef G, et al. SIRT1 promotes the central adaptive response to diet restriction through activation of the dorsomedial and lateral nuclei of the hypothalamus. Journal of Neuroscience. 2010;30(30):10220–10232. doi: 10.1523/JNEUROSCI.1385-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 301.Malavasi F, Deaglio S, Zaccarello G, et al. The hidden life of NAD+-consuming ectoenzymes in the endocrine system. Journal of Molecular Endocrinology. 2010;45(4):183–191. doi: 10.1677/JME-10-0082. [DOI] [PubMed] [Google Scholar]
- 302.Noriega LG, Feige JN, Canto C, et al. CREB and ChREBP oppositely regulate SIRT1 expression in response to energy availability. EMBO Reports. 2011;12(10):1069–1076. doi: 10.1038/embor.2011.151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 303.Braidy N, Guillemin GJ, Mansour H, Chan-Ling T, Poljak A, Grant R. Age related changes in NAD+ metabolism oxidative stress and sirt1 activity in wistar rats. Plos ONE. 2011;6(4) doi: 10.1371/journal.pone.0019194. Article ID e19194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 304.Imai SI. Dissecting systemic control of metabolism and aging in the NAD world: the importance of SIRT1 and NAMPT-mediated NAD biosynthesis. FEBS Letters. 2011;585(11):1657–1662. doi: 10.1016/j.febslet.2011.04.060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 305.Sauro MD, Greenberg RP. Endogenous opiates and the placebo effect: a meta-analytic review. Journal of Psychosomatic Research. 2005;58(2):115–120. doi: 10.1016/j.jpsychores.2004.07.001. [DOI] [PubMed] [Google Scholar]
- 306.Cheng HL, Mostoslavsky R, Saito S, et al. Developmental defects and p53 hyperacetylation in Sir2 homolog (SIRT1)-deficient mice. Proceedings of the National Academy of Sciences of the United States of America. 2003;100(19):10794–10799. doi: 10.1073/pnas.1934713100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 307.Liu D, Pitta M, Mattson MP. Preventing NAD+ depletion protects neurons against excitotoxicity: bioenergetic effects of mild mitochondrial uncoupling and caloric restriction. Annals of the New York Academy of Sciences. 2008;1147:275–282. doi: 10.1196/annals.1427.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 308.Anderson DW, Bradbury KA, Schneider JS. Broad neuroprotective profile of nicotinamide in different mouse models of MPTP-induced parkinsonism. European Journal of Neuroscience. 2008;28(3):610–617. doi: 10.1111/j.1460-9568.2008.06356.x. [DOI] [PubMed] [Google Scholar]
- 309.Yang J, He L, Wang J, Adams JD., Jr. Early administration of nicotinamide prevents learning and memory impairment in mice induced by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine. Pharmacology Biochemistry and Behavior. 2004;78(1):179–183. doi: 10.1016/j.pbb.2004.03.007. [DOI] [PubMed] [Google Scholar]
- 310.Outeiro TF, Grammatopoulos TN, Altmann S, et al. Pharmacological inhibition of PARP-1 reduces alpha-synucleinand MPP+-induced cytotoxicity in Parkinson's disease in vitro models. Biochemical and Biophysical Research Communications. 2007;357(3):596–602. doi: 10.1016/j.bbrc.2007.03.163. [DOI] [PubMed] [Google Scholar]
- 311.Outeiro TF, Kontopoulos E, Altmann SM, et al. Sirtuin 2 inhibitors rescue α-synuclein-mediated toxicity in models of Parkinson’s disease. Science. 2007;317(5837):516–519. doi: 10.1126/science.1143780. [DOI] [PubMed] [Google Scholar]
- 312.Barber-Singh J, Seo BB, Nakamaru-Ogiso E, Lau YS, Matsuno-Yagi A, Yagi T. Neuroprotective effect of long-term NDI1 gene expression in a chronic mouse model of parkinson disorder. Rejuvenation Research. 2009;12(4):259–267. doi: 10.1089/rej.2009.0854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 313.Kim SH, Lu HF, Alano CC. Neuronal sirt3 protects against excitotoxic injury in mouse cortical neuron culture. Plos ONE. 2011;6(3):13 pages. doi: 10.1371/journal.pone.0014731. Article ID e14731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 314.Choi WS, Palmiter RD, Xia Z. Loss of mitochondrial complex I activity potentiates dopamine neuron death induced by microtubule dysfunction in a Parkinson's disease model. Journal of Cell Biology. 2011;192(5):873–882. doi: 10.1083/jcb.201009132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 315.Park G, Jeong JW, Kim JE. SIRT1 deficiency attenuates MPP+-induced apoptosis in dopaminergic cells. FEBS Letters. 2011;585(1):219–224. doi: 10.1016/j.febslet.2010.11.048. [DOI] [PubMed] [Google Scholar]
- 316.Kitaoka Y, Hayashi Y, Kumai T, et al. Axonal and cell body protection by nicotinamide adenine dinucleotide in tumor necrosis factor-induced optic neuropathy. Journal of Neuropathology and Experimental Neurology. 2009;68(8):915–927. doi: 10.1097/NEN.0b013e3181afecfa. [DOI] [PubMed] [Google Scholar]
- 317.Ohsawa I, Ishikawa M, Takahashi K, et al. Hydrogen acts as a therapeutic antioxidant by selectively reducing cytotoxic oxygen radicals. Nature Medicine. 2007;13(6):688–694. doi: 10.1038/nm1577. [DOI] [PubMed] [Google Scholar]
- 318.Nakashima I, Kawamoto Y, Takeda K, Kato M. Control of genetically prescribed protein tyrosine kinase activities by environment-linked redox reactions. Enzyme Research. 2011;2011:10 pages. doi: 10.4061/2011/896567. Article ID 896567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 319.Kaidi A, Weinert B, Choudhary C, Jackson SP. Human SIRT6 promotes DNA end resection through CtIP deacetylation. Science. 2010;329(5997):1348–1353. doi: 10.1126/science.1192049. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 320.Stover PJ. One-carbon metabolism-genome interactions in folate-associated pathologies. Journal of Nutrition. 2009;139(12):2402–2405. doi: 10.3945/jn.109.113670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 321.Van Os J, Kenis G, Rutten BP. The environment and schizophrenia. Nature. 2010;468(7321):203–212. doi: 10.1038/nature09563. [DOI] [PubMed] [Google Scholar]
- 322.Roseboom TJ, Painter RC, van Ableen AF, Veenendaal MV, de Rooij SR. Hungary in the womb: what are the consequences? Lessions from the Dutch famine. Maturitas. 2011;70(2):141–145. doi: 10.1016/j.maturitas.2011.06.017. [DOI] [PubMed] [Google Scholar]
- 323.Poulsen P, Esteller M, Vaag A, Fraga MF. The epigenetic basis of twin discordance in age-related diseases. Pediatric Research. 2007;61(5, part 2):38R–42R. doi: 10.1203/pdr.0b013e31803c7b98. [DOI] [PubMed] [Google Scholar]
- 324.Konner M, Eaton SB. Paleolithic nutrition: twenty-five years later. Nutrition in Clinical Practice. 2010;25(6):594–602. doi: 10.1177/0884533610385702. [DOI] [PubMed] [Google Scholar]
- 325.Chen JH, Cottrell EC, Ozanne SE. Early growth and ageing. Nestle Nutrition Workshop Series. 2010;65:41–54. doi: 10.1159/000281144. [DOI] [PubMed] [Google Scholar]
- 326.Libert S, Cohen D, Guarente L. Neurogenesis directed by Sirt1. Nature Cell Biology. 2008;10(4):373–374. doi: 10.1038/ncb0408-373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 327.Noble M, Mayer-Pröschel M, Pröschel C. Redox regulation of precursor cell function: insights and paradoxes. Antioxidants and Redox Signaling. 2005;7(11-12):1456–1467. doi: 10.1089/ars.2005.7.1456. [DOI] [PubMed] [Google Scholar]
- 328.Prozorovski T, Schulze-Topphoff U, Glumm R, et al. Sirt1 contributes critically to the redox-dependent fate of neural progenitors. Nature Cell Biology. 2008;10(4):385–394. doi: 10.1038/ncb1700. [DOI] [PubMed] [Google Scholar]
- 329.Barker DJ. The foetal and infant origins of inequalities in health in Britain. Journal of Public Health Medicine. 1991;13(2):64–68. [PubMed] [Google Scholar]
- 330.Soto AM, Sonnenschein C. The tissue organization field theory of cancer: a testable replacement for the somatic mutation theory. BioEssays. 2011;33(5):332–340. doi: 10.1002/bies.201100025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 331.Ganesan A, Nolan L, Crabb SJ, Packham G. Epigenetic therapy: histone acetylation, DNA methylation and anti-cancer drug discovery. Current Cancer Drug Targets. 2009;9(8):963–981. doi: 10.2174/156800909790192428. [DOI] [PubMed] [Google Scholar]
- 332.Kenny PA, Lee GY, Bissell MJ. Targeting the tumor microenvironment. Frontiers in Bioscience. 2007;12(9):3468–3474. doi: 10.2741/2327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 333.Czarnecka AM, Bartnik E. The role of the mitochondrial genome in ageing and carcinogenesis. Journal of Aging Research. 2011;2011:10 pages. doi: 10.4061/2011/136435. Article ID 136435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 334.Hansen KD, Timp W, Bravo HC, et al. Increased methylation variation in epigenetic domains across cancer types. Nature Genetics. 2011;43(8):768–775. doi: 10.1038/ng.865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 335.Fassati A, Mitchison NA. Testing the theory of immune selection in cancers that break the rules of transplantation. Cancer Immunology, Immunotherapy. 2010;59(5):643–651. doi: 10.1007/s00262-009-0809-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 336.Gatenby RA, Gillies RJ, Brown JS. The evolutionary dynamics of cancer prevention. Nature Reviews Cancer. 2010;10(8):526–527. doi: 10.1038/nrc2892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 337.Merlo LMF, Pepper JW, Reid BJ, Maley CC. Cancer as an evolutionary and ecological process. Nature Reviews Cancer. 2006;6(12):924–935. doi: 10.1038/nrc2013. [DOI] [PubMed] [Google Scholar]
- 338.Peto R, Roe FJC, Lee PN. Cancer and ageing in mice and men. British Journal of Cancer. 1975;32(4):411–426. doi: 10.1038/bjc.1975.242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 339.Vaisitti T, Audrito V, Serra S, et al. NAD+-metabolizing ecto-enzymes shape tumor-host interactions: the chronic lymphocytic leukemia model. FEBS Letters. 2011;585(11):1514–1520. doi: 10.1016/j.febslet.2011.04.036. [DOI] [PubMed] [Google Scholar]
- 340.D’Andrea FP, Safwat A, Kassem M, Gautier L, Overgaard J, Horsman MR. Cancer stem cell overexpression of nicotinamide N-methuyltransferase enhances cellular radiation resistance. Radiotherapy & Oncology. 2011;99(3):373–378. doi: 10.1016/j.radonc.2011.05.086. [DOI] [PubMed] [Google Scholar]
- 341.Albracht SP, Meijer AJ, Rydstrom J. Mammalian NADH: ubiquinone oxidoreductase (Complex I) and nicotinamide nucleotide transhydrogenase (Nnt) together regulate the mitochondrial production of H2O2-implications for their role in disease, especially cancer. Journal of Bioenergetics and Biomembranes. 2011;43(5):541–564. doi: 10.1007/s10863-011-9381-4. [DOI] [PubMed] [Google Scholar]
- 342.Chandra N, Bhagavat R, Sharma E, Sreekanthreddy P, Somasundaram K. Virtual screening, identification and experimental testing of novel inhibitors of PBEF1/Visfatin/NMPRTase for glioma therapy. Journal of Clinical Bioinformatics. 2011;1(5) doi: 10.1186/2043-9113-1-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 343.Bass J, Takahashi JS. Circadian rhythms: redox redux. Nature. 2011;469(7331):476–478. doi: 10.1038/469476a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 344.Reed JC. Cancer: priming cancer cells for death. Science. 2011;334(6059):1075–1076. doi: 10.1126/science.1215568. [DOI] [PubMed] [Google Scholar]
- 345.Kirkland JB. Niacin status and genomic instability in bone marrow cells; mechanisms favouring the progression of leukemogenesis. Subcellular Biochemistry. 2012;56:21–36. doi: 10.1007/978-94-007-2199-9_2. [DOI] [PubMed] [Google Scholar]
- 346.Prendergast GC. Cancer: why tumours eat tryptophan. Nature. 2011;478(7368):192–194. doi: 10.1038/478192a. [DOI] [PubMed] [Google Scholar]
- 347.Griguer CE, Oliva CR. Bioenergetics pathways and therapeutic resistance in gliomas: emerging role of mitochondria. Current Pharmaceutical Design. 2011;17(23):2421–2427. doi: 10.2174/138161211797249251. [DOI] [PubMed] [Google Scholar]
- 348.Prensner JR, Chinnaiyan AM. Metabolism unhinged: IDH mutations in cancer. Nature Medicine. 2011;17(3):291–293. doi: 10.1038/nm0311-291. [DOI] [PubMed] [Google Scholar]
- 349.Yan H, Bigner DD, Velculescu V, Parsons DW. Mutant metabolic enzymes are at the origin of gliomas. Cancer Research. 2009;69(24):9157–9159. doi: 10.1158/0008-5472.CAN-09-2650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 350.Metallo CM, Gameiro PA, Bell EL. Reductive metabolism by IDH1 mediates lipogenesis under hypoxia. doi: 10.1038/nature10602. Nature. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 351.Anastasiou D, Poulogiannis G, Asra JM, et al. Inhibition of pyruvate kinase m2 by reactive oxygen species contributes to cellular antioxidant reponses. Science. 2011;334(6060):78–1283. doi: 10.1126/science.1211485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 352.Vander Heiden MG. Targeting cancer metabolism: a therapeutic window opens. Nature Reviews Drug Discovery. 2011;10(9):671–684. doi: 10.1038/nrd3504. [DOI] [PubMed] [Google Scholar]
- 353.Locasale JW, Grassian AR, Melman T, et al. Phosphoglycerate dehydrogenase diverts glycolytic flux and contributes to oncogenesis. Nature Genetics. 2011;43(9):869–874. doi: 10.1038/ng.890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 354.Metallo CM, Vander Heiden MG. Metabolism strikes back: metabolic flux regulates cell signaling. Genes and Development. 2010;24(24):2717–2722. doi: 10.1101/gad.2010510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 355.Dang L, White DW, Gross S, et al. Cancer-associated IDH1 mutations produce 2-hydroxyglutarate. Nature. 2010;465(7300):p. 966. doi: 10.1038/nature09132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 356.Hamanaka RB, Chandel NS. Warburg effect and redox balance. Science. 2011;334(6060):1219–1220. doi: 10.1126/science.1215637. [DOI] [PubMed] [Google Scholar]
- 357.Brundin P, Li JY, Holton JL, Lindvall O, Revesz T. Research in motion: the enigma of Parkinson’s disease pathology spread. Nature Reviews Neuroscience. 2008;9(10):741–745. doi: 10.1038/nrn2477. [DOI] [PubMed] [Google Scholar]
- 358.Segovia G, Arco AD, Mora F. Environmental enrichment, prefrontal cortex, stress, and aging of the brain. Journal of Neural Transmission. 2009;116(8):1007–1016. doi: 10.1007/s00702-009-0214-0. [DOI] [PubMed] [Google Scholar]
- 359.Okuda H, Tatsumi K, Makinodan M, Yamauchi T, Kishimoto T, Wanaka A. Environmental enrichment stimulates progenitor cell proliferation in the amygdala. Journal of Neuroscience Research. 2009;87(16):3546–3553. doi: 10.1002/jnr.22160. [DOI] [PubMed] [Google Scholar]
- 360.Stadtfeld M, Hochedlinger K. Induced pluripotency: history, mechanisms, and applications. Genes and Development. 2010;24(20):2239–2263. doi: 10.1101/gad.1963910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 361.Poss KD. Advances in understanding tissue regenerative capacity and mechanisms in animals. Nature Reviews Genetics. 2010;11(10):710–722. doi: 10.1038/nrg2879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 362.Lamour G, Eftekhari-Bafrooei A, Borguet E, Souès S, Hamraoui A. Neuronal adhesion and differentiation driven by nanoscale surface free-energy gradients. Biomaterials. 2010;31(14):3762–3771. doi: 10.1016/j.biomaterials.2010.01.099. [DOI] [PubMed] [Google Scholar]
- 363.Winner B, Kohl Z, Gage FH. Neurodegenerative disease and adult neurogenesis. European Journal of Neuroscience. 2011;33(6):1139–1151. doi: 10.1111/j.1460-9568.2011.07613.x. [DOI] [PubMed] [Google Scholar]
- 364.Fruscione F, Scarfì S, Ferraris C, et al. Regulation of human mesenchymal stem cell functions by an autocrine loop involving NAD+ release and P2Y11-mediated signaling. Stem Cells and Development. 2011;20(7):1183–1198. doi: 10.1089/scd.2010.0295. [DOI] [PubMed] [Google Scholar]
- 365.Conboy IM, Conboy MJ, Wagers AJ, Girma ER, Weismann IL, Rando TA. Rejuvenation of aged progenitor cells by exposure to a young systemic environment. Nature. 2005;433(7027):760–764. doi: 10.1038/nature03260. [DOI] [PubMed] [Google Scholar]
- 366.Ding L, Ellis MJ, Shunqiang L, et al. Genome remodelling in a basal-like breast cancer metastasis and xenograft. Nature. 2010;464(7291):999–1005. doi: 10.1038/nature08989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 367.Capp J-P. Stochastic gene expression is the driving force of cancer. BioEssays. 2011;33(10):781–782. doi: 10.1002/bies.201100092. [DOI] [PubMed] [Google Scholar]
- 368.Masip M, Veiga A, Belmonte JC, Simón C. Reprogramming with defined factors: from induced pluripotency to induced transdifferentiation. Molecular Human Reproduction. 2010;16(11):856–868. doi: 10.1093/molehr/gaq059. [DOI] [PubMed] [Google Scholar]
- 369.Hochedlinger K, Blelloch R, Brennan C, et al. Reprogramming of a melanoma genome by nuclear transplantation. Genes and Development. 2004;18(15):1875–1885. doi: 10.1101/gad.1213504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 370.Bely AE, Nyberg KG. Evolution of animal regeneration: re-emergence of a field. Trends in Ecology and Evolution. 2010;25(3):161–170. doi: 10.1016/j.tree.2009.08.005. [DOI] [PubMed] [Google Scholar]
- 371.Anderson ML. Neural reuse: a fundamental organizational principle of the brain. Behavioral and Brain Sciences. 2010;33(4):245–266. doi: 10.1017/S0140525X10000853. [DOI] [PubMed] [Google Scholar]
- 372.Lui JH, Hansen DV, Kriegstein AR. Development and evolution of the human neocortex. Cell. 2011;146(1):18–36. doi: 10.1016/j.cell.2011.06.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 373.Niven JE, Laughlin SB. Energy limitation as a selective pressure on the evolution of sensory systems. Journal of Experimental Biology. 2008;211(11):1792–1804. doi: 10.1242/jeb.017574. [DOI] [PubMed] [Google Scholar]
- 374.Adams DS, Masi A, Levin M. H+ pump-dependent changes in membrane voltage are an early mechanism necessary and sufficient to induce Xenopus tail regeneration. Development. 2007;134(7):1323–1335. doi: 10.1242/dev.02812. [DOI] [PubMed] [Google Scholar]
- 375.Pinker S. The cognitive niche: coevolution of intelligence, sociality, and language. Proceedings of the National Academy of Sciences of the United States of America. 2010;107(2):8993–8999. doi: 10.1073/pnas.0914630107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 376.Sinclair DA, Oberdoerffer P. The ageing epigenome: damaged beyond repair? Ageing Research Reviews. 2009;8(3):189–198. doi: 10.1016/j.arr.2009.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 377.Levin M. Bioelectric mechanisms in regeneration: unique aspects and future perspectives. Seminars in Cell and Developmental Biology. 2009;20(5):543–556. doi: 10.1016/j.semcdb.2009.04.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 378.Li Y, He X, Li Y, et al. Nicotinamide phosphoribosyltransferase (Nampt) affects the lineage fate determination of mesenchymal stem cells: a possible cause for reduced osteogenesis and increased adipogenesis in older individuals. Journal of Bone and Mineral Research. 2011;26(11):2656–2664. doi: 10.1002/jbmr.480. [DOI] [PubMed] [Google Scholar]
- 379.Lang Y, Arnepalli RR, Tiwari A, et al. A review of hydrogen production: methods, materials and nanotechnology. Journal of Nanoscience and Nanotechnology. 2011;11(5):3719–3739. doi: 10.1166/jnn.2011.4157. [DOI] [PubMed] [Google Scholar]
- 380.Rupprecht J, Hankamer B, Mussgnug JH, Ananyev G, Dismukes C, Kruse O. Perspectives and advances of biological H2 production in microorganisms. Applied Microbiology and Biotechnology. 2006;72(3):442–449. doi: 10.1007/s00253-006-0528-x. [DOI] [PubMed] [Google Scholar]
- 381.James W. The Principles of Psychology. New York, NY, USA: Dover press; 1890. [Google Scholar]
- 382.Savulescu J, Bostrom N. Human Enhancement. Oxford, UK: Oxford University Press; 2009. [Google Scholar]
- 383.Ball P. Physics of life: the dawn of quantum biology. Nature. 2011;474(7351):272–274. doi: 10.1038/474272a. [DOI] [PubMed] [Google Scholar]
- 384.Bähner F, Weiss EK, Birke G, et al. Cellular correlate of assembly formation in oscillating hippocampal networks in vitro. Proceedings of the National Academy of Sciences of the United States of America. 2011;108(35):E607–E616. doi: 10.1073/pnas.1103546108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 385.Kann O, Huchzermeyer C, Kovacs R, Wirtz S, Schuelke M. Gamma oscillations in the hippocampus require high complex I gene expression and stress functional performance of mitochondira. Brain. 2010;134, part 2:345–358. doi: 10.1093/brain/awq333. [DOI] [PubMed] [Google Scholar]
- 386.Druback DA, Rabinstein AA, Molano J. Free will, freedom of choice and frontotemporal lobar degerenation. Mens Sana Monographs. 2011;9(1):238–250. doi: 10.4103/0973-1229.77440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 387.Didic M, Barbeau EJ, Felician O, et al. Which memory system is impaired frist in Alzheimer’s disease? Alzheimer's Disease. 2011;27(1):11–22. doi: 10.3233/JAD-2011-110557. [DOI] [PubMed] [Google Scholar]
- 388.Fried I, Mukamel R, Kreiman G. Internally generated preactivation of single neurons in human medial frontal cortex predecits volition. Neuron. 2011;69:548–562. doi: 10.1016/j.neuron.2010.11.045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 389.Gailliot MT, Baumeister RF. The physiology of will power: linking blood glucose to self-control. Personality and Social Psychology Review. 2007;11(4):303–327. doi: 10.1177/1088868307303030. [DOI] [PubMed] [Google Scholar]
- 390.Bushey D, Tononi G, Cirelli C. Sleep and synaptic homeostasis: structural evidence in Drosophila . Science. 2011;332(6037):1576–1581. doi: 10.1126/science.1202839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 391.Sperling RA, LaViolette PS, O’Keefe K, et al. Amyloid deposition is associated with impaired default network function in older persons without dementia. Neuron. 2009;63(2):178–188. doi: 10.1016/j.neuron.2009.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 392.Klingberg T. The Overflowing Brain: Information Overload and the Limits of Working Memory. Oxford, UK: Oxford University Press; 2008. [Google Scholar]
- 393.Yizhar O, Fenno LE, Prigge M, et al. Neocortical excitation/inhibition balance in information processing and social dysfunction. Nature. 2011;477(7363):171–178. doi: 10.1038/nature10360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 394.Amodio DM, Frith CD. Meeting of minds: the medial frontal cortex and social cognition. Nature Reviews Neuroscience. 2006;7(4):268–277. doi: 10.1038/nrn1884. [DOI] [PubMed] [Google Scholar]
- 395.Piguet O, Petersén A, Yin Ka Lam B, et al. Eating and hypothalamus changes in behavioral-variant frontotemporal dementia. Annals of Neurology. 2011;69(2):312–319. doi: 10.1002/ana.22244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 396.Wang H-X, Xu W, Pei J-J. Leisure activities, cognition and dementia. Biochimica et Biophysica Acta. 2012;1822(3):482–491. doi: 10.1016/j.bbadis.2011.09.002. [DOI] [PubMed] [Google Scholar]
- 397.Kirkland JB. Niacin requirements for genomic stability. Mutation Research. doi: 10.1016/j.mrfmmm.2011.11.008. [DOI] [PubMed] [Google Scholar]
- 398.Harris LT, Fiske ST. Social groups that elicit disgust are differentially processed in mPFC. Social Cognitive and Affective Neuroscience. 2007;2(1):45–51. doi: 10.1093/scan/nsl037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 399.Penberthy WT, Tsunoda I. The importance of NAD in multiple sclerosis. Current Pharmaceutical Design. 2009;15(1):64–99. doi: 10.2174/138161209787185751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 400.Berer K, Mues M, Koutrolos M, et al. Commensal microbiota and myelin autoantigen cooperate to trigger autoimmune demyelination. Nature. 2011;479(7374):538–541. doi: 10.1038/nature10554. [DOI] [PubMed] [Google Scholar]
- 401.Finger BC, Dinan TG, Cryan JF. High-fat diet selectively protects against the effects of chronic social stress in the mouse. Neuroscience. 2011;192:351–360. doi: 10.1016/j.neuroscience.2011.06.072. [DOI] [PubMed] [Google Scholar]
- 402.Gesquiere LR, Learn NH, Simao MC, Onyango PO, Alberts SC, Altmann J. Life at the top: rank and stress in wild male baboons. Science. 2011;333(6040):357–360. doi: 10.1126/science.1207120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 403.Sachs JL, Essenberg CJ, Turcotte MM. New paradigms for the evolution of beneficial infections. Trends in Ecology and Evolution. 2011;26(4):202–209. doi: 10.1016/j.tree.2011.01.010. [DOI] [PubMed] [Google Scholar]
- 404.Wiessner P. Hunting, healing, and hxaro exchange: a long-term perspective on !Kung (Ju/’hoansi) large-game hunting. Evolution and Human Behavior. 2002;23(6):407–436. [Google Scholar]
- 405.Hamann K, Warneken F, Greenberg JR, Tomasello M. Collaboration encourages equal sharing in children but not in chimpanzees. Nature. 2011;476(7360):328–331. doi: 10.1038/nature10278. [DOI] [PubMed] [Google Scholar]
- 406.Hrdy SB. Mothers and Others: The Evolutionary Origins of Utual Understanding. Massachusetts, Mass, USA: Harvard University Press; 2009. [Google Scholar]
- 407.Murphy-Berman V, Berman JJ, Singh P, Pachauri A, Kuman P. Factors affecting allocation to needy and meritorious recipients: a cross-cultural comparison. Journal of Personality and Social Psychology. 1984;46(6):1267–1272. [Google Scholar]
- 408.Huang C, Andres AM, Ratliff EP, Hernandez G, Lee P, Gottlieb RA. Preconditioning involves selective mitophagy mediated by parkin and p62/SQSTM1. Plos ONE. 2011;6(6) doi: 10.1371/journal.pone.0020975. Article ID e20975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 409.Walter P, Ron D. The unfolded protein response: from stress pathway to homeostatic regulation. Science. 2011;334(6059):1081–1086. doi: 10.1126/science.1209038. [DOI] [PubMed] [Google Scholar]
- 410.Smith MH, Ploegh HL, Weissman JS. Road to ruin: targeting proteins for degradation in the endoplasmic reticulum. Science. 2011;334(6059):1086–1090. doi: 10.1126/science.1209235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 411.Turgut O, Tandogan I, Karapinar H, Aydin G. Preconditioning, postconditioning, stunning and hibernation: towards an integrated insight into the mechanisms of hypoperfusion/reperfusion. International Journal of Cardiology. 2011;146(3):442–443. doi: 10.1016/j.ijcard.2010.10.112. [DOI] [PubMed] [Google Scholar]