

Abstract
In vivo assessment of gene expression is desirable to obtain information on the extent and duration of transduction of tissue after gene delivery. We have developed an in vivo, potentially noninvasive, method for detecting virally mediated gene transfer to the liver. The method employs an adenoviral vector carrying the gene for the brain isozyme of murine creatine kinase (CK-B), an ATP-buffering enzyme expressed mainly in muscle and brain but absent from liver, kidney, and pancreas. Gene expression was monitored by 31P magnetic resonance spectroscopy (MRS) using the product of the CK enzymatic reaction, phosphocreatine, as an indicator of transfection. The vector was administered into nude mice by tail vein injection, and exogenous creatine was administered in the drinking water and by i.p. injection of 2% creatine solution before 31P MRS examination, which was performed on surgically exposed livers. A phosphocreatine resonance was detected in livers of mice injected with the vector and was absent from livers of control animals. CK expression was confirmed in the injected animals by Western blot analysis, enzymatic assays, and immunofluorescence measurements. We conclude that the syngeneic enzyme CK can be used as a marker gene for in vivo monitoring of gene expression after virally mediated gene transfer to the liver.
Rapid development of human gene therapy has produced methods of efficiently transducing several tissues and organs in a variety of animal models of disease as well as in humans (1). The liver is a key target for gene therapy applications because, besides being the site of inherited and acquired diseases for which no alternative treatment is currently available (2), it may also serve as an efficient site for production of secreted therapeutic proteins (3–5). Despite the impressive advances that have characterized the field of virally mediated gene transfer in the last decade, for most vectors the site and persistence of expression after administration is variable and remains to be defined. Several in vitro markers have been used for this purpose, consisting of either cellular proteins whose presence can be histologically detected (β-galactosidase or enhanced green fluorescent protein) (6, 7) or proteins secreted in the bloodstream, whose concentration can be measured in the serum. In the first case a tissue biopsy is required, and the presence of the reporter gene product will not provide precise insight into the extent of the region that has been transduced. In the second case, detectable circulating levels of a secreted reporter protein suggest transduction of target cells but do not delineate their location.
In vivo imaging techniques can noninvasively provide quantitative information on both the presence and anatomical location of a signal. The methods that have been proposed are all based on coexpression of a reporter or marker gene that can be detected by an appropriate noninvasive imaging modality. The marker gene is coexpressed with the therapeutic gene under the control of the same promoter within a bicistronic unit that includes a type II internal ribosomal entry site (8–10). Luciferase has been used as a marker gene detectable by bioluminescence (11–14), and the green fluorescent protein has been used as an optically detected reporter gene (15). Imaging of adenoviral gene expression has been accomplished by positron emission tomography and single photon emission computed tomography using herpes simplex virus 1 thymidine kinase (8, 16–19) and dopamine type 2 receptors (19). Weissleder and co-workers (20, 21) have developed a transferrin receptor marker gene that lacks the iron regulatory region and expresses an approximately 5-fold excess of transferrin receptors on the cell surface. Gradient echo MRI is used to detect T2 decreases induced by delivery of transferrin that is covalently linked to magnetite–dextran complexes. Meade's laboratory (22) has developed a unique Gd (gadolinium) chelate that is screened from contact with water by a galactopyranose residue that can be removed by β-galactosidase. This reaction produces an analog of Gd-DTPA (diethylenetriaminepentaacetic acid), a well-known MRI contrast agent that is detected with T1-weighted imaging pulse sequences. Walter et al. (23) have introduced Drosophila melanogaster arginine kinase as a marker gene for magnetic resonance spectroscopy (MRS) detection of gene therapy of muscle diseases. This enzyme phosphorylates arginine, leading to production of arginine phosphate, a unique metabolite that is not otherwise found in mammalian tissues and is readily detectable by 31P MRS.
Each of these methods has advantages and limitations. The optical methods are relatively cheap and easy to implement. Bioluminescence is, however, limited to relatively small animals, and the green fluorescent protein is suitable only for superficial detection except in transparent animals. Optical detection of internal tissues is best accomplished with near infrared (NIR) markers, which remain to be perfected. Positron emission tomography (PET) and single photon emission computed tomography (SPECT) offer the advantage of high sensitivity, and these methods (as well as MRI, MRS, and NIR imaging) are suitable for translation into the clinic. However, the spatial resolution of PET is currently limited to 2 mm in mice and about 4–6 mm in humans; SPECT has even poorer resolution. In addition, viral thymidine kinase is toxic to the liver (18), causes sterility in male mice (17), and elicits an immunogenic response after chronic use (17), and PET ligands that cross the blood–brain barrier are not yet available. The dopamine method avoids many of these difficulties but requires distinction from endogenous receptors. The MRI methods provide exquisite spatial resolution, but superparamagnetic and ferromagnetic agents that enhance T2 or T*2, such as magnetite, provide limited image contrast in tissues such as the liver. Additional limitations of the transferrin receptor methods are interference from endogenous transferrin receptors and the inability of transferrin to cross the blood–brain barrier. The caged Gd complex, while potentially providing higher tissue contrast, has lower sensitivity than superparamagnetic or ferromagnetic contrast agents and currently lacks a suitable marker enzyme for extracellular activation or a suitable transport vehicle for intracellular activation. In addition, it does not cross the blood–brain barrier. Arginine kinase provides a unique signal in mammalian muscle, but it is a nonmammalian protein that is expected to induce an immune response, and 31P has poor spatial resolution (several millimeters in mice at high magnetic field and 3 cm in humans at 1.5 T). This method also requires saturation-transfer experiments for quantitation. Therefore, none of these methods is perfect, and there remains a need to develop marker genes that may overcome at least some of these limitations.
Localized MRS or spectroscopic imaging allows noninvasive mapping of the distribution of metabolites and quantitative measurement of enzymatic activity by magnetization-transfer experiments (24, 29). Ideally, an MRS reporter system should provide a unique resonance in the spectrum of the target tissue to eliminate the need for discrimination from background signals emanating from endogenous genes; it should employ a syngeneic marker protein to evade immunological responses; the reporter gene should be small enough to fit in the same expression vector as the therapeutic gene; and its enzymatic activity should not interfere with the function of the target tissue. Creatine kinase (CK), which was suggested as a marker gene by Koretsky et al. (25), satisfies all these criteria. It is expressed primarily in muscle (MM isoform) and brain (BB isoform), but is absent from the liver, kidney, and pancreas (26). It catalyzes the reaction
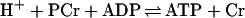 |
1 |
that provides energy buffering in the tissues where the enzyme is expressed (PCr, phosphocreatine; Cr, creatine). Ectopic expression of CK in the liver has been accomplished in transgenic mice by Koretsky's laboratory (25, 27), who demonstrated that CK expression has no harmful side effects on liver and may even protect this organ against ischemic insults. While the potential utility of CK as a marker gene has been suggested, until the present study the feasibility of implementing this strategy in vivo by viral delivery of the CK gene had not been demonstrated.
Adenoviral vectors administered to the general circulation mainly target the liver (28). We injected an adenoviral vector carrying the mouse gene for the brain CK isozyme (CK-B) into the tail veins of mice and were able to detect in vivo a PCr peak in the surgically exposed livers by 31P MRS. Our study demonstrates that CK can be used as a marker for in vivo, potentially noninvasive, monitoring of gene expression in liver. It may also be suitable as a marker gene even in tissues that normally express this enzyme, because the increment in CK activity can be determined by magnetization-transfer experiments, as recently demonstrated by Roman et al. (29) in mouse skeletal muscle. In summary, the key advantages of CK as a marker gene are absence of immune reaction, compatibility with magnetic resonance equipment that is readily available in the laboratory and the clinic, absence of radioactivity, and apparent absence of harmful side effects. The chief limitation is limited sensitivity and spatial resolution, but in organs such as liver, muscle, and brain the resolution and sensitivity are adequate and are improving as higher-field instruments become available.
Methods
Adenoviral Vector Construction.
The coding sequence of the mouse CK-B gene was amplified from mouse brain cDNA (CLONTECH) by using the following primers: forward, ATGCCCTTCTCCAACAGCCATAA; reverse, TAGCTCTTCGACCGTCATCTTC.
The PCR product was cloned in the PCR 2.1 vector (Invitrogen, Carlsbad, CA) and sequenced. The mouse CK-B sequence was then cloned in the pAd-CMV-link shuttle plasmid (Vector Core, Institute for Human Gene Therapy, Univ. of Pennsylvania; CMV indicates the cytomegalovirus promoter). The pAd-CMV-transgene plasmid was used for cotransfection of 293 cells (American Type Culture Collection) with ClaI-digested H5.010CMVEGFP viral backbone containing deletions in map units 1–9 of the E1 region and in the E3 region (sub360) to create adenoviruses by homologous recombination. Recombinant plaques were initially isolated through green/white selection (30) and confirmed by restriction enzyme analysis of DNAs extracted by the Hirt procedure (30). The positive plaques were purified and grown up in 293 cells for CsCl gradient purification (28).
In Vivo MRS.
Six- to 8-week-old NCR-nude female mice (Taconic Farms) were injected with 1 × 1011 particles of the Ad-CMV-CK-B vector via the tail vein. Noninjected mice were used as controls. Animals were fed with 2% (wt/vol) creatine (Sigma) in water for 5 days and then given 100 μl of the same solution intraperitoneally (i.p.) 1 hr before performing both ex vivo and in vivo tests.
Seven days after injection, mice were anesthetized by i.p. injection of 0.18 ml of ketamine/xylazine (10/2 mg/ml) per mouse. In vivo MRS studies were performed on a Varian INOVA spectrometer interfaced to an Oxford 9.4-T/8.9-cm vertical bore magnet. Magnetic field homogeneity was improved by shimming on the 23Na signal to a linewidth at half-maximum of ≈60 Hz. A homemade single-turn solenoidal surface coil (8-mm diameter) (31) tuned to 31P (162 MHz) and to 23Na (105 MHz) was placed on the exposed liver. The tissues surrounding the liver (abdominal muscle, skin, and intestines) were isolated with a copper Faraday shield to eliminate magnetic resonance (MR) signals originating from these tissues. The animal was placed on a water-circulating blanket to maintain its core temperature at 37°C. In vivo 31P spectra were acquired by using the following parameters: 256 scans with an interpulse delay of 4 s, a radio frequency (rf) pulse of 22 μs corresponding approximately to a 90° flip angle; 15-kHz sweep width; 2,048 data points; and a line-broadening of 25 Hz to increase the signal-to-noise ratio.
CK-B Agarose Gel, Western Blot, Enzymatic Assay, Immunofluorescence, and MRS of Liver Extracts.
For ex vivo detection of CK-B in transduced livers, animals were injected with 1 × 1011 particles of Ad-CMV-CK-B via tail vein, injected i.p. with 2% creatine 4 days later, and killed after 1 hr. Mouse livers were flash-frozen and used independently for the various assays of CK expression. Necropsied livers and a control brain from the animals were homogenized in lysis buffer as described elsewhere (32). The protein concentration in the liver lysates was assessed by the Bradford assay (Bio-Rad). Ten micrograms of each lysate was loaded on a 1% agarose gel, electrophoretically separated, and stained with the CK isoenzyme kit (Sigma Diagnostics). Ten micrograms of the same lysates was added to 4× Laemmli reducing sample buffer, heated for 5 min at 95°C, and loaded on an SDS/glycine/4–12% polyacrylamide gradient gel (Novex, San Diego). Electrophoresis was at room temperature for 90 min at 90 V, proteins were transferred to a nitrocellulose filter, and the filter was blotted with a 1:200 dilution of a rabbit anti-human CK-B polyclonal antibody (Chromaprobe, Mountain View, CA).
Quantitative enzymatic assay (25) (Sigma Diagnostics) of liver extracts was performed according to the supplier's instructions.
For measurement of immunofluorescence, livers were harvested 4 days after tail vein administration of 1 × 1011 particles of Ad-CMV-CK-B or PBS, embedded, and frozen in liquid nitrogen. Ten-micrometer-thick cryosections were fixed in 4:1 methanol/acetone for 20 min at −20°C, blocked with PBS/2% nonfat dry milk at 37°C for 30 min, incubated at 37°C for 1 hr with biotin-conjugated anti-CK-B antibody (Chromaprobe) diluted 1:200 in PBS/2% milk, washed three times with PBS/2% milk, incubated at 37°C for 20 min with streptavidin coupled to the fluorescent dye CY3 (Sigma) diluted 1:200 in PBS/2% milk, washed three times with PBS/2% milk and once in PBS, and finally mounted with Vectashield mounting medium containing 4′,6-diamidino-2-phenylindole (DAPI; Vector Laboratories). Images were captured by using a fluorescence microscope with a triple filter for detection of fluorescein, rhodamine, and DAPI.
MRS was also conducted on perchloric acid extracts of livers. The exposed liver was flash-frozen by clamping between two aluminum plates that were precooled in liquid nitrogen. Frozen liver was weighed and pulverized 1:4 (wt/vol) in perchloric acid (12%, vol/vol) at 4°C and centrifuged at 15,555 × g for 20 min. The supernatant was neutralized with KOH to pH = 7.4 ± 0.1 and centrifuged at 1,200 × g for 10 min. The supernatant was freeze-dried and stored at 4°C. The sample was subjected to 1H followed by 31P MRS on a 9.4-T/8.9-cm Bruker Avance DMX 400 spectrometer. For analysis, the lyophilized powder was reconstituted in 0.6 ml of deuterated water (D2O), pH adjusted to 7.4 ± 0.1 (uncorrected for isotope effects), and transferred to a 5-mm MR tube. Proton MR spectra were acquired using 65,536 data points, 48 scans, 6.6-kHz sweep width, interscan delay of 9 s, and rf pulse of 4 μs corresponding to a 45° flip angle. The same sample was used for 31P MRS experiments after 0.2 M EDTA had been added (final concentration = 50 mM). 31P spectra were acquired with the following parameters: overnight scans with a delay of 4 s; rf pulse of 8 μs corresponding to a 45° flip angle; 65,536 data points; 13-kHz sweep width, and a line-broadening of −1 Hz. A capillary insert containing the sodium salt of 3-(trimethylsilyl)propionic-2,2,3,3,-d4 acid (TSP) or methylene diphosphonic acid (MDP) was used as reference for 1H and 31P MRS, respectively; the chemical shift of TSP was set to 0 ppm in 1H spectra, and MDP was set to 16.8 ppm in 31P spectra. Each capillary was calibrated for spectroscopic quantitation of metabolite concentrations in the extracted sample.
Results
In Vivo Detection of CK Expression by 31P MRS in Transduced Livers.
Seven days after tail vein administration of the Ad-CMV-CK-B vector, mice were subjected to in vivo 31P MRS. Fig. 1A shows a representative in vivo 31P spectrum from control livers (n = 6) and Fig. 1B, a spectrum from livers transduced with the Ad-CMV-CK-B vector (n = 6). A unique PCr resonance was observed in all of the livers of injected mice, but was absent from livers of control mice. The intensity of the PCr peak in transduced livers varies among individual animals. The chemical shift of PCr (0.3 ppm) relative to the α-phosphorus of ATP (−7.3 ppm) did not differ between experimental groups and was the same as previously reported in CK-expressing livers of transgenic mice (25).
Figure 1.

In vivo 31P MRS detection of murine livers transduced with the CK-B gene. (A) Representative spectrum from control livers (n = 6). (B) Spectrum from transduced livers (n = 6). PME, phosphomonoesters; Pi, inorganic phosphate; PDE, phosphodiesters; γ-, α-, and β-ATP, γ, α and β resonance of adenosine triphosphate.
Ex Vivo Detection of CK Expression in Transduced Livers.
To test ex vivo for the expression of biologically active CK in transduced livers, 5 μg of liver homogenate was loaded on a 1% agarose gel and separated by electrophoresis. The gel was then stained for CK activity. Fig. 2A displays the result of gel electrophoresis of liver homogenates. A stained band corresponding to CK-B was observed from a sample of purified rabbit brain CK (lane 1), from the homogenate of the brain of a control animal (lane 2), as well from an equal amount of liver homogenate from an injected animal (lane 3). No band was detected at the same location in the extract from the liver of a control mouse (lane 4).
Figure 2.

Ex vivo analysis of adenovirus-mediated CK-B expression in murine livers. (A) CK-B agarose assay. (B) Western blot using anti-human CK-B antibody. Lane 1 contains commercial CK-B from rabbit brain, lanes 2 and 4 originate from brain and liver extracts, respectively, from a noninjected mouse, and lane 3 is a liver extract from an injected animal. (C) CK quantitative enzymatic assay. The striped bar represents the levels of enzymatic activity present in the liver homogenates of three injected animals (26,030 ± 5,600 Sigma units per mg of total protein); the corresponding levels for three control mice (on the right) are undetectable.
To confirm that the enzymatic activity detected in our samples corresponded to CK-B, the same amount of homogenates as was used for agarose staining was separated by SDS/PAGE, transferred to a nitrocellulose filter, and blotted with an anti-CK-B antibody (Fig. 2B). A band of the expected size was detected in the lanes corresponding to the positive control (lane 1), to mouse brain (lane 2), and to the transduced liver (lane 3), but was absent from the liver of the noninjected mouse (lane 4).
CK enzymatic activity present in the transduced livers was then independently quantified by using an enzymatic assay in which ADP and PCr were converted to ATP and creatine, depending on the amount of recombinant CK present in the sample tested. The ATP was then used in a coupled reaction to generate NADPH, whose absorbance was measured. The liver homogenates from three animals injected with the Ad-CMV-CK-B showed high levels of CK activity, which was undetectable in three control homogenates (Fig. 2C).
To analyze the pattern of CK expression in the liver after administration of 1 × 1011 particles of adenoviral vector via the tail vein, we used an anti-CK antibody to perform immunofluorescence assays on liver cryosections. Whereas no fluorescent signal was detectable in the control liver (Fig. 3A), 80–100% of hepatocytes from the transduced liver exhibited a homogeneous cytoplasmic signal corresponding to CK (Fig. 3B).
Figure 3.

Ex vivo analysis of adenovirus-mediated CK-B expression in murine livers: immunofluorescence on control (A) and transduced (B) livers. CK signal is red fluorescent, whereas the blue signal corresponds to 4′,6-diamidino-2-phenylindole (DAPI)-stained nuclei.
Quantitation of creatine, PCr, and other metabolites in liver was achieved by 1H and 31P MRS of extracts (3–5 animals in each group). As shown in Table 1, the PCr level in transduced livers was 0.8 ± 0.3 μmol per gram of wet tissue, whereas it was 0.0 in control livers. Both transduced and control livers had detectable levels of free creatine with large variations from animal to animal (1.7–6.6 μmol/g wet weight in transduced livers and 4.5–11.9 μmol/g wet weight in controls). The variation in creatine level may primarily reflect the different uptake of creatine by individual animals.
Table 1.
Concentration of selected metabolites from liver extracts estimated by 31P and 1H MR spectroscopy
| Livers | PCr, μmol/gww (n = 5) | ATP,* μmol/gww (n = 5) | Pi, μmol/gww (n = 5) | Creatine, μmol/gww (n = 3) |
|---|---|---|---|---|
| Transfected | 0.8 ± 0.3 | 2.0 ± 0.2 | 4.5 ± 0.5 | 4.0 ± 1.4 |
| Control | 0.0 ± 0.0 | 1.7 ± 0.5 | 4.7 ± 0.7 | 8.2 ± 1.8 |
Data are expressed as mean ± SEM; gww, gram of wet weight of tissues.
Measured from resonance of the β phosphorus of ATP.
Discussion
As viral vectors able to evade the immune response, such as adeno-associated virus (AAV), are being validated in clinical trials (33), interest is growing in monitoring gene expression in vivo. In fact, in humans vectors are used to transduce large volumes of tissue, e.g., muscle, or are injected into the general circulation. Characterization of the degree of gene transfer and the duration of expression is important and currently requires invasive analysis of biopsy specimens. Several noninvasive imaging techniques have been recently used to monitor gene expression in vivo. Each of these methods has distinct limitations (see above) that make it essential to develop additional methods.
By employing an adenoviral vector, we have successfully introduced the BB isoenzyme of CK into livers of nude mice. The expression of CK activity was detected in vivo by the unique resonance of PCr in the 31P MR spectra of transduced livers. Livers were surgically exposed to facilitate exclusion of PCr signals emanating from muscle, which exhibits high levels of this metabolite. In the future, localized MRS, such as three-dimensional 31P spectroscopic imaging, will be used to detect PCr in the transduced liver without surgical exposure of the liver. Such procedures, which are routinely performed on humans in the clinic (34), are considerably more demanding to implement on mice because of problems with probe design, spatial resolution, and signal-to-noise ratio. Furthermore, MRS imaging using the PCr signal may be facilitated by increasing PCr signal intensity substantially either by increasing the creatine concentration (35) or by administering cyclocreatine, which has an equilibrium constant for the ATP to phosphocyclocreatine reaction (i.e., the reverse of reaction 1) 30-fold higher than for cyclocreatine (35, 36).
However, it must be noted that detection of PCr or phosphocyclocreatine in the liver serves as only a qualitative indication of CK expression. Because CK is an enzyme, its concentration modifies only the rate at which equilibrium is achieved and not the relative concentrations of reactants and products. The immunofluorescence measurements indicate that 80–100% of the liver has been transduced by a dose of 1011 particles. Provided that one employs a viral dose in excess of the threshold level required to transduce all of the cells in the liver (as this dose appears to be), changing the dose should not substantially modify the fraction of cells transduced, but should modify the level of expression in the cells. Consequently, the PCr level detected should not reflect the extent of transduction, but may reflect differences in levels of creatine or hypoxia (35). Therefore, quantitation of transduction requires measurement of CK activity. This can be accomplished noninvasively by a magnetization-transfer experiment that measures the rate of reaction 1 in the PCr to ATP direction (24, 29). This flux should be proportional to Vmax, which in turn should be proportional to the concentration of CK. It is also possible to detect CK expression even in brain and muscle by using the magnetization-transfer MR technique to measure incremental changes in CK activity (29). Consequently, CK may also prove to be a suitable marker of gene therapy of these tissues.
It should also be noted that the spectra in Fig. 1 show normal ATP/Pi ratios; hence, the absence of PCr in spectra of control livers is not due to ischemia.
We confirmed expression of CK in the transduced livers by using two different enzymatic assays, one more qualitative that was based on the migration of the enzyme in an agarose gel, and the other that was suitable for quantitating the activity of the enzyme in the liver. In addition, we demonstrated the correct size of the recombinant protein by Western blotting as well as the homogeneous distribution of CK expression by immunofluorescence in liver after systemic administration of vector at high dose.
Expression of CK should not result in toxicity because the end product is PCr, which naturally occurs in muscle and brain of the host. The sensitivity limits of this method can be delineated with animals administered various viral doses. In addition, this method may provide target cells with a rapidly accessible source of high-energy phosphates for maintenance of ATP homeostasis under conditions of high energy demand and during ischemic episodes.
The in vivo detection of CK gene expression here involves 31P MRS or spectroscopic imaging that has limited spatial resolution (several millimeters in mice and 3 cm in humans). This limitation may be overcome by developing a marker suitable for 1H MRI to achieve higher spatial resolution (22).
Gene transfer to liver holds considerable promise for curing a wide range of diseases. Adenoviral vectors that efficiently target hepatocytes but elicit a robust cytotoxic T lymphocyte response are already being tested in clinical trials for several tumors, including some in the liver (37). In addition, several groups have focused their interest on identification of a vector that permits sustained and prolonged expression of transgenes in this organ (4, 38, 39). The availability of a small nontoxic reporter system such as CK should permit repeated in vivo monitoring of gene transfer in this context.
Acknowledgments
The technical assistance of the Animal Model group of the Institute for Human Gene Therapy is gratefully acknowledged. The authors are indebted to Dr. Alan Koretsky for helpful suggestions. Dr. Susan Wehrli assisted in MRS study of liver extracts, and Dr. Navin Bansal provided helpful guidance in the surgical exposure of the liver. This work was sponsored by the National Institutes of Health (Grants P30 DK47757 to J.M.W., R24 CA83105 to J.D.G., and P20 CA86255 to J.D.G.), the Cystic Fibrosis Foundation, the Juvenile Diabetes Foundation, and Genovo, Inc. (now Targeted Genetics), a company J.M.W. founded and holds equity in. A.A. is the recipient of a fellowship from Telethon-Italia (371/B).
Abbreviations
- MRS
magnetic resonance spectroscopy
- MR
magnetic resonance
- CK
creatine kinase
- CK-B
brain isozyme of CK
- PCr
phosphocreatine
- Ad
adenovirus
- CMV
cytomegalovirus promoter
Footnotes
This paper was submitted directly (Track II) to the PNAS office.
References
- 1.Mountain A. Trends Biotechnol. 2000;18:119–128. doi: 10.1016/s0167-7799(99)01416-x. [DOI] [PubMed] [Google Scholar]
- 2.Raper S E, Wilson J M, Yudkoff M, Robinson M B, Ye X, Batshaw M L. J Inherit Metab Dis. 1998;21:119–137. doi: 10.1023/a:1005369926784. [DOI] [PubMed] [Google Scholar]
- 3.Snyder R O, Miao C, Meuse L, Tubb J, Donahue B A, Lin H F, Stafford D W, Patel S, Thompson A R, Nichols T, et al. Nat Med. 1999;5:64–70. doi: 10.1038/4751. [DOI] [PubMed] [Google Scholar]
- 4.Snyder R O, Miao C H, Patijn G A, Spratt S K, Danos O, Nagy D, Gown A M, Winther B, Meuse L, Cohen L K, et al. Nat Genet. 1997;16:270–276. doi: 10.1038/ng0797-270. [DOI] [PubMed] [Google Scholar]
- 5.Koeberl D D, Alexander I E, Halbert C L, Russell D W, Miller A D. Proc Natl Acad Sci USA. 1997;94:1426–1431. doi: 10.1073/pnas.94.4.1426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Fisher K J, Jooss K, Alston J, Yang Y, Haecker S E, High K, Pathak R, Raper S E, Wilson J M. Nat Med. 1997;3:306–312. doi: 10.1038/nm0397-306. [DOI] [PubMed] [Google Scholar]
- 7.Bennett J, Maguire A M, Cideciyan A V, Schnell M, Glover E, Anand V, Aleman T S, Chirmule N, Gupta A R, Huang Y, et al. Proc Natl Acad Sci USA. 1999;96:9920–9925. doi: 10.1073/pnas.96.17.9920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Tjuvajev J G, Joshi A, Callegari J, Lindsley L, Joshi R, Balatoni J, Finn R, Larson S M, Sadelain M, Blasberg R G. Neoplasia. 1999;1:315–320. doi: 10.1038/sj.neo.7900053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Gambhir S S, Barrio J R, Herschman H R, Phelps M E. J Nucl Cardiol. 1999;6:219–233. doi: 10.1016/s1071-3581(99)90083-1. [DOI] [PubMed] [Google Scholar]
- 10.Gambhir S S, Barrio J R, Herschman H R, Phelps M E. Nucl Med Biol. 1999;26:481–490. doi: 10.1016/s0969-8051(99)00021-9. [DOI] [PubMed] [Google Scholar]
- 11.Contag C H, Jenkins D, Contag P R, Negrin R S. Neoplasia. 2000;2:41–52. doi: 10.1038/sj.neo.7900079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Contag C H, Spilman S D, Contag P R, Oshiro M, Eames B, Dennery P, Stevenson D K, Benaron D A. Photochem Photobiol. 1997;66:523–531. doi: 10.1111/j.1751-1097.1997.tb03184.x. [DOI] [PubMed] [Google Scholar]
- 13.Contag P R, Olomu I N, Stevenson D K, Contag C H. Nat Med. 1998;4:245–247. doi: 10.1038/nm0298-245. [DOI] [PubMed] [Google Scholar]
- 14.Jacobs W R, Jr, Barletta R G, Udani R, Chan J, Kalkut G, Sosne G, Kieser T, Sarkis G J, Hatfull G F, Bloom B R. Science. 1993;260:819–822. doi: 10.1126/science.8484123. [DOI] [PubMed] [Google Scholar]
- 15.Misteli T, Spector D L. Nat Biotechnol. 1997;15:961–964. doi: 10.1038/nbt1097-961. [DOI] [PubMed] [Google Scholar]
- 16.Gambhir S S, Barrio J R, Wu L, Iyer M, Namavari M, Satyamurthy N, Bauer E, Parrish C, MacLaren D C, Borghei A R, et al. J Nucl Med. 1998;39:2003–2011. [PubMed] [Google Scholar]
- 17.Gambhir S S, Bauer E, Black M E, Liang Q, Kokoris M S, Barrio J R, Iyer M, Namavari M, Phelps M E, Herschman H R. Proc Natl Acad Sci USA. 2000;97:2785–2790. doi: 10.1073/pnas.97.6.2785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Tjuvajev J G, Chen S H, Joshi A, Joshi R, Guo Z S, Balatoni J, Ballon D, Koutcher J, Finn R, Woo S L, Blasberg R G. Cancer Res. 1999;59:5186–5193. [PubMed] [Google Scholar]
- 19.Gambhir S S, Herschman H R, Cherry S R, Barrio J R, Satyamurthy N, Toyokuni T, Phelps M E, Larson S M, Balatoni J, Finn R, et al. Neoplasia. 2000;2:118–138. doi: 10.1038/sj.neo.7900083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wunderbaldinger P, Bogdanov A, Weissleder R. Eur J Radiol. 2000;34:156–165. doi: 10.1016/s0720-048x(00)00196-0. [DOI] [PubMed] [Google Scholar]
- 21.Weissleder R, Moore A, Mahmood U, Bhorade R, Benveniste H, Chiocca E A, Basilion J P. Nat Med. 2000;6:351–355. doi: 10.1038/73219. [DOI] [PubMed] [Google Scholar]
- 22.Louie A Y, Huber M M, Ahrens E T, Rothbacher U, Moats R, Jacobs R E, Fraser S E, Meade T J. Nat Biotechnol. 2000;18:321–325. doi: 10.1038/73780. [DOI] [PubMed] [Google Scholar]
- 23.Walter G, Barton E R, Sweeney H L. Proc Natl Acad Sci USA. 2000;97:5151–5155. doi: 10.1073/pnas.97.10.5151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Bittle J A, DeLayre J, Ingwall J S. Biochemistry. 1987;26:6083–6090. doi: 10.1021/bi00393a021. [DOI] [PubMed] [Google Scholar]
- 25.Koretsky A P, Brosnan M J, Chen L H, Chen J D, Van Dyke T. Proc Natl Acad Sci USA. 1990;87:3112–3116. doi: 10.1073/pnas.87.8.3112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Dawson D M, Fine I H. Arch Neurol. 1967;16:175–180. doi: 10.1001/archneur.1967.00470200063005. [DOI] [PubMed] [Google Scholar]
- 27.Koretsky A P. Am J Physiol. 1992;262:C261–C275. doi: 10.1152/ajpcell.1992.262.2.C261. [DOI] [PubMed] [Google Scholar]
- 28.Gao G P, Yang Y, Wilson J M. J Virol. 1996;70:8934–8943. doi: 10.1128/jvi.70.12.8934-8943.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Roman B B, Wieringa B, Koretsky A P. J Biol Chem. 1997;272:17790–17794. doi: 10.1074/jbc.272.28.17790. [DOI] [PubMed] [Google Scholar]
- 30.Davis A R, Meyers K, Wilson J M. Gene Ther. 1998;5:1148–1152. doi: 10.1038/sj.gt.3300705. [DOI] [PubMed] [Google Scholar]
- 31.Zhou R, Bansal N, Leeper D B, Glickson J D. Cancer Res. 2000;60:3532–3536. [PubMed] [Google Scholar]
- 32.Auricchio A, di Domenico M, Castona G, Bilancio A, Migliaccio A. FEBS Lett. 1994;353:16–20. doi: 10.1016/0014-5793(94)00987-2. [DOI] [PubMed] [Google Scholar]
- 33.Kay M A, Manno C S, Ragni M V, Larson P J, Couto L B, McClelland A, Glader B, Chew A J, Tai S J, Herzog R W, et al. Nat Genet. 2000;24:257–261. doi: 10.1038/73464. [DOI] [PubMed] [Google Scholar]
- 34.Negendank W G, Padovic-Shaller K A, Li C W, Murphy-Boesch J, Stoyanova R, Krigel R L, Schilder R J, Smith M R, Brown T R. Cancer Res. 1995;55:3286–3294. [PubMed] [Google Scholar]
- 35.Brosnan M J, Chen L, Van Dyke T A, Koretsky A P. J Biol Chem. 1990;265:20849–20855. [PubMed] [Google Scholar]
- 36.LoPresti P, Cohn M. Biochim Biophys Acta. 1989;998:317–320. doi: 10.1016/0167-4838(89)90291-4. [DOI] [PubMed] [Google Scholar]
- 37.Habib N A, Hodgson H J, Lemoine N, Pignatelli M. Hum Gene Ther. 1999;10:2019–2034. doi: 10.1089/10430349950017383. [DOI] [PubMed] [Google Scholar]
- 38.Wang L, Takabe K, Bidlingmaier S M, Ill C R, Verma I M. Proc Natl Acad Sci USA. 1999;96:3906–3910. doi: 10.1073/pnas.96.7.3906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kafri T, Blomer U, Peterson D A, Gage F H, Verma I M. Nat Genet. 1997;17:314–317. doi: 10.1038/ng1197-314. [DOI] [PubMed] [Google Scholar]