

Summary
Normal human lymphocytes resisted the hydrolytic action of secretory phospholipase A2 but became susceptible to the enzyme following treatment with a calcium ionophore, ionomycin. To test the hypothesis that this susceptibility requires exposure of the anionic lipid phosphatidylserine on the external face of the cell membrane, experiments were repeated with a human Burkitt’s lymphoma cell line (Raji cells). In contrast to normal lymphocytes or S49 mouse lymphoma cells, most of the Raji cells (83%) did not translocate phosphatidylserine to the cell surface upon treatment with ionomycin. Those few that did display exposed phosphatidylserine were hydrolyzed immediately upon addition of phospholipase A2. Interestingly, the remaining cells were also completely susceptible to the enzyme but were hydrolyzed at a slower rate and after a latency of about 100 s. In contradistinction to the defect in phosphatidylserine translocation, Raji cells did display other physical membrane changes upon ionomycin treatment that may be relevant to hydrolysis by phospholipase A2. These changes were detected by merocyanine 540 and trimethylammonium diphenylhexatriene fluorescence and were common among normal lymphocytes, S49 cells, and Raji cells. The levels of these latter effects corresponded well with the relative rates of hydrolysis among the three cell lines. These results suggested that while phosphatidylserine enhances the rate of cell membrane hydrolysis by secretory phospholipase A2, it is not an absolute requirement. Other physical properties such as membrane order contribute to the level of membrane susceptibility to the enzyme independent of phosphatidylserine.
Introduction
Secretory phospholipase A2 (sPLA2) hydrolyzes phospholipids at the sn-2 position producing free fatty acid and lysophospholipid. It represents a family of isozymes and is thought to participate in a broad range of functions including generation of pro-inflammatory mediators, protection against bacterial and fungal infections, digestion, participation in reproduction, nervous system development, and wound healing. Furthermore, sPLA2 has been implicated in several pathologies such as atherosclerosis, inflammatory diseases, septic shock, and cancer (reviewed in [1]).
A major characteristic of sPLA2 is its extreme sensitivity to the biophysical properties of the bilayers on which it acts. This sensitivity is important because it prevents damage to the membranes of normal healthy mammalian cells while allowing efficient catalysis of bacterial membranes and those of damaged or dying cells [2–6]. In particular, the membranes of several types of cultured leukocytes have been reported to convert from being resistant to becoming susceptible to the action of the enzyme during apoptosis and other forms of biochemical cell death [4,5,7–10].
The nature of the biophysical changes in the cell membrane that trigger catalysis by sPLA2 during cell death is not yet established. Nevertheless, important clues have been identified from biophysical studies of both artificial bilayers and living cells [6–8,10–18]. Two prominent candidates have emerged: 1) a reduction in the strength of interactions among adjacent phospholipids resulting in increased lipid mobility and greater interlipid spacing and 2) the presence of anionic lipids on the membrane surface [6,10,14,19]. The hypothesis is that reduced interactions among neighboring phospholipids allow substrates to migrate into the active site of adsorbed sPLA2, while negative charge facilitates initial adsorption of the enzyme to the membrane surface [11,18,20–22]. The primary source of these surface anions during cell death is phosphatidylserine (PS). Ordinarily, this lipid is confined to the inner leaflet of the cell membrane, and several enzymes are involved in maintaining that asymmetry [23]. However, during apoptosis and other forms of cell death, PS is translocated to the outer leaflet [23–25]. Consequently, the exposed PS functions as a marker for processes that eliminate the dying cell such as phagocytosis [24,26–28] and, perhaps, hydrolysis by sPLA2 [10,18,19]. Arguments for the importance of lipid-neighbor interactions and of surface PS are accumulating, although the existing evidence from cells depends on temporal correlations rather than direct experimental manipulations [6–8,10,14].
In order to distinguish more directly the potential roles of PS and membrane lipid interactions in capacitating hydrolysis during cell death, we have initiated a series of experiments with Raji Burkitt’s lymphoma cells. These cells are deficient in their ability to express the gene for scamblase, the enzyme primarily responsible for exposure of PS during apoptosis [29–31]. Here, we report investigations with a calcium ionophore, ionomycin, as the stimulator of cell death. Ionomycin was chosen for our first study because it stimulates complete immediate exposure of PS and full hydrolytic susceptibility to sPLA2 in other cultured lymphoma cells [7,9]. Since Raji cells originated from a human tumor, we used freshly-isolated normal human lymphocytes as a control for comparison. Data were also compared to that attained with a cell line used in previous studies, S49 mouse lymphoma cells, as a positive control.
Materials and methods
2.1 Reagents
The monomeric aspartate-49 phospholipase A2 from the venom of the water moccasin, Agkistrodon piscivorus piscivorus was isolated according to the procedure by Maraganore et al. [32]. Ionomycin, merocyanine 540 (MC540), propidium iodide, 1-(trimethylammoniumphenyl)- 6-phenyl-1,3,5 hexatriene p-toluenesulfonate (TMA-DPH), and Alexa Fluor 488 annexin V conjugate were acquired from Invitrogen (Carlsbad, CA). The acrylodan-labeled fatty acid-binding protein (ADIFAB) was purchased from FFA Sciences (San Diego, CA). Ionomycin and MC540 were dissolved as stock solutions in dimethylsulfoxide (DMSO), and TMA-DPH was suspended in dimethylformamide. Annexin V, ADIFAB, and propidium iodide were stored as aqueous solutions. Other reagents (buffer, salts, detergent) were obtained from standard sources.
2.2 Preparation of Cells
S49 mouse lymphoma cells were grown in DMEM at 37°C in humidified air containing 10% CO2. Raji human Burkitt’s lymphoma cells were grown at 37°C at 5% CO2 in RPMI containing 10% fetal bovine serum and L-glutamine. S49 and Raji cells were prepared as described [5]. Normal human lymphocytes were isolated from residual whole blood samples obtained during routine physical exams at the Brigham Young University Student Health Center by density centrifugation with Histopaque-1077 from Sigma-Aldrich (St. Louis, MO) according to the manufacturer protocol. Purity of preparations was determined by microscopic analysis of Wright-stained aliquots. On average, 91% of the leukocytes in the samples were lymphocytes. Control experiments revealed that standard procedures for eliminating contaminating erythrocytes in lymphocyte preparations (≤ 12%) were not usable in these experiments because they perturb cell membranes in ways that interfere with the results. We therefore tested the impact of added erythrocytes on the experimental observations and determined that they did not alter the results at concentrations below 50%. In flow cytometry experiments, erythrocytes and non-lymphocyte leukocytes were excluded from the analyses.
For experiments, cells were washed and suspended in MBSS (134 mM NaCl, 6.2 mM KCl, 1.6 mM CaCl2, 1.2 nM MgCl2, 18.0 mM Hepes, 13.6 mM glucose, pH 7.4 at 37 °C) at a density of 0.4–3.0 × 106 cells/ml. Aliquots were transferred to quartz fluorometer sample cells and allowed 5 min to equilibrate in the relevant spectrofluorometer (see below) for measurements of fluorescence emission intensity and anisotropy or to polycarbonate tubes in a shaking water bath for flow cytometry. Sample homogeneity and temperature were maintained in the spectrofluorometers by magnetic stirring and by circulating heated water through a jacket in the sample chamber. All experiments and incubations were performed at 37°C. In each procedure where ionomycin was added to cell samples (600 nM final), the experiment was repeated with an equivalent volume of DMSO (0.5% v/v final) to control for the effects of the solvent.
2.3 Membrane Hydrolysis
As a generic model for sPLA2, we used enzyme purified from snake venom as described above. This source of enzyme was selected because of its previous use in biophysical studies of cell death by calcium ionophore and because of its strong similarity to the human group X sPLA2, the isoform that responds best to ionophore-stimulated cells [10,16,17]. We verified these assertions for human lymphocytes using samples of human groups IIa, V, and X sPLA2 generously provided by Michael Gelb (University of Washington) and Wonhwa Cho (University of Illinois, Chicago). In these calibration experiments, the group IIa enzyme displayed 0.4% of the activity observed with snake venom sPLA2 toward human lymphocytes treated with ionomycin whereas the groups V and X isozymes exhibited 25.7% and 82.8% respectively. These relative responses of the isozymes to ionomycin-treated lymphocytes were identical to those reported previously for S49 lymphoma cells and therefore justified the use of the snake venom enzyme as an assay for cell susceptibility to sPLA2 [10,16].
Fatty acid release from cell membranes was assayed in real time by fluorescence spectroscopy (Fluoromax 3, Horiba Jobin-Yvon, Edison, NJ ) using ADIFAB (excitation = 390 nm, emission = 432 and 505 nm, bandpass = 4 nm) [33]. After data acquisition was initiated, ADIFAB (65 nM final) was added and allowed adequate time to equilibrate. The sample was then incubated with ionomycin for 10 min after which sPLA2 (70 nM final) was added. The generalized polarization (GP) was calculated from the fluorescence emission intensities at 505 and 432 nm (I505 and I432) in order to quantify the amount of fatty acid produced [34].
| (1) |
Propidium iodide fluorescence (37 μM final, excitation = 537 nm, emission = 617 nm, bandpass = 4 nm) was used to assess the proportion of cells in the sample that were susceptible to hydrolysis by sPLA2 [8,9]. Addition and incubation times of reagents were as stated for ADIFAB assays with the exception that triton X-100 (0.25% final v/v) was added after the propidium iodide fluorescence intensity became stable (often > 2000 s in human lymphocytes and Raji cells) to permeabilize all cells and obtain a maximum signal. The proportions of cells in relevant categories (PIA, “already dead”; PIB, “killed by ionomycin”; PIC, “alive, but susceptible to sPLA2”) were quantified as follows (illustrated in Fig. 1):
| (2A) |
| (2B) |
| (2C) |
where I0 is the average intensity during the first 20 s after addition of propidium iodide to the sample, Iss is the average intensity during the 50 s prior to addition of ionomycin, Iiono is the average intensity during the 50 s prior to addition of sPLA2, IsPLA2 is the average intensity during the 50 s prior to addition of triton X-100, and Imax is the maximum intensity after adding triton X-100.
Fig. 1.

Idealized diagram of propidium iodide fluorescence time course during treatment with ionomycin and sPLA2. See Eq. 2A-2C and associated text for explanation. The dotted lines represent the times of addition of ionomycin (“+ iono”), sPLA2 (“+ sPLA2”), and triton X-100 (“+ triton”).
2.4 Membrane Biophysical Properties
Data using MC540 (170 nM final) was acquired at 585 nm (excitation = 540 nm, bandpass = 4 nm) before the addition of the probe (for background scattered and stray light), after a 5-min equilibration, 10 min after addition of ionomycin or DMSO, and during hydrolysis by sPLA2. Data were analyzed by subtracting the background intensity and calculating the change in emission intensities normalized to the initial intensity (before ionomycin).
Steady-state fluorescence anisotropy measurements (TMA-DPH) were obtained with a PC-1 spectrofluorometer in the “L” configuration (ISS, Champaign, IL) using Glan-Thompson polarizers. Fluorescence intensity was measured with excitation (350 nm, 16 nm bandpass) and emission (452 nm, 16 nm bandpass) polarizers oriented at 0° from vertical (parallel) and repeated with the excitation polarizer at 0° and the emission polarizer at 90° (perpendicular). The data were adjusted for transmission efficiency of the emission monochromator by calculating the ratio of emission intensity at 0° (v) and 90° (h) with the excitation polarizer oriented at 90° (G):
| (3) |
The TMA-DPH anisotropy was calculated using the traditional equation:
| (4) |
where Ivv and Ivh are the intensities with parallel and perpendicular polarizer configurations, respectively.
2.5 Flow Cytometry
The amount of PS exposed on the outer leaflet of the cell membrane was determined by annexin V binding to the cells. Flow cytometry data were collected using a FACSCanto flow cytometer (BD Biosciences, San Jose, CA) with an argon laser (488 nm) and an emission bandpass filter that transmits 515–545 nm. Cells were treated with DMSO or ionomycin (10–15 min) and concurrently labeled in MBSS with conjugated annexin V at the dilution indicated by the manufacturer (Invitrogen).
Results
3.1 Exposure of PS
Figure 2A displays the effect of calcium ionophore (ionomycin) treatment to stimulate exposure of PS on the outer surface of the plasma membrane of normal human lymphocytes. As has been reported for multiple cell lines, introduction of calcium into normal lymphocytes caused immediate exposure of the anionic lipid (dotted lines in Fig. 2A) [7]. In contrast, Raji Burkitt’s lymphoma cells showed minimal enhancement of annexin binding following ionophore treatment; 7% of the cells stained positive for annexin V under control conditions (similar to the other cell lines shown in Fig. 2B), and this number increased to only 17% upon addition of ionomycin. This observation of minimal annexin staining has been reported previously for Raji cells under a variety of experimental conditions [29–31]. As shown in Fig. 2B, the difference between normal (black) and Raji (dark gray) lymphocytes was reproducible and statistically significant. Addition of a higher concentration of ionomycin overcame the limitation in Raji cells, as shown by the white bar in Fig. 2B. Data obtained with S49 mouse lymphoma cells (light gray) are included as a positive reference.
Fig. 2.
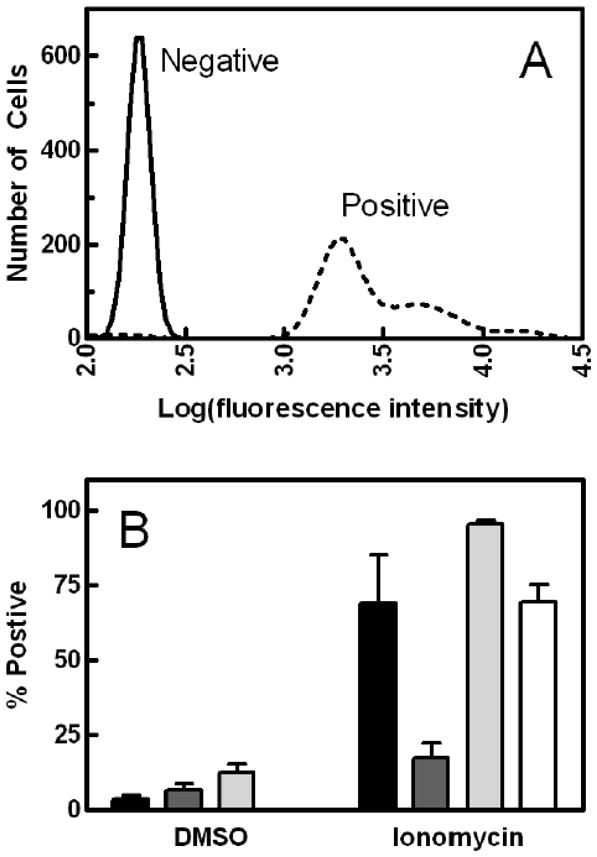
Effect of ionomycin treatment on PS exposure. Panels A and B: Data represent the level of staining by annexin V among cell subpopulations assayed by flow cytometry. Normal lymphocytes (A) were treated with DMSO (solid line) or ionomycin (dotted line) and incubated with annexin V for 15 min and immediately transferred to the flow cytometer. Panel B: The percentages of normal lymphocytes (black), Raji cells (dark gray), and S49 cells (light gray) staining positive for annexin V after addition of DMSO or ionomycin were identified from experiments such as those of panel A for 3–5 independent samples. The white bar for “ionomycin” represents Raji cells treated with 1.2 μM ionomycin instead of the typical 600 nM. The effects of cell type (22.2% of the variation) and ionomycin (52.7% of the variation) were both significant (p < 0.0001) by two-way analysis of variance. Interaction between cell type and drug treatment was also significant, indicating that the size of the effect of ionomycin depended on cell type (17.9% of the variation, p < 0.0001). One-way analysis of variance with a Bonferroni post-test on the four groups treated with ionomycin revealed that the Raji cells with normal ionomycin was the only distinguishable group (p < 0.01).
3.2 Hydrolysis
The action of sPLA2 toward normal human lymphocytes was qualitatively similar to that reported previously for cultured cells (Fig. 3A and Ref. [7]). In the absence of ionomycin, a transient burst of hydrolysis was observed followed by slow restoration of fatty acid levels eventually to the baseline value. This reaction profile has been shown to represent hydrolysis of a small subpopulation of cells made vulnerable to the enzyme by trauma during sample preparation followed by clearance of released fatty acid through reacylation by the other healthy cells in the sample [5]. Addition of ionomycin enhanced the total amount of hydrolysis observed. Both the initial rate (23%) and total amount (87%) of hydrolysis in the normal lymphocytes (Fig. 3A) were lower than those observed with S49 cells (Fig. 3C).
Fig. 3.

Membrane hydrolysis by sPLA2 assayed by ADIFAB fluorescence. Cells were treated with ionomycin (black) or DMSO (gray) for 10 min and then with sPLA2 (time 0 on the graphs). Hydrolysis of normal lymphocytes (A), Raji cells (B), and S49 cells (C) was assayed using ADIFAB, and the experimental data were averaged to create a composite representation of the time profile for each cell type. Dotted lines indicate the range of ± one SEM for 3–9 independent samples. Panel D: The maximum rate of hydrolysis during the first 12 s after sPLA2 addition was calculated from the data of ionomycin-treated samples in panels A–C. One-way analysis of variance followed by a Bonferroni post-test revealed that these rates were statistically greater for S49 cells compared to the other cell types (p < 0.001), which were indistinguishable from each other.
The data for control samples of Raji cells were similar to control observations for the other cell types (Fig. 3B). However, the ionophore-treated Raji samples displayed time courses with a more complex shape, beginning with an initial rate comparable to that of the normal lymphocytes (Fig. 3D) followed by a latent secondary phase of hydrolysis that eventually matched the total of the S49 cells.
One of the challenges of comparing hydrolysis time courses beyond the initial phase is that enzyme may eventually be able to enter the cell and hydrolyze internal membranes thus clouding interpretation of observed totals. It is therefore useful to also estimate the percentage of cells for which an intact plasma membrane has been attacked by sPLA2. As explained in earlier studies, this estimate can be obtained using the vital stain propidium iodide [9]. Cells that resist hydrolysis exclude the dye (low fluorescence intensity) regardless of the presence of sPLA2. Alternatively, cells that have become vulnerable to the enzyme fluoresce brightly upon introduction of sPLA2 as hydrolysis damages the membrane allowing propidium iodide to enter and bind to nucleic acids. Figure 4 displays this phenomenon for normal (Fig. 4A), Raji (Fig. 4B), and S49 (Fig. 4C) cells. In all three cases, treatment with ionomycin caused all of the cells in the sample to become susceptible to sPLA2. Moreover, the shape of the time profile of propidium iodide uptake mirrored the hydrolysis data in Fig. 3, although the proportion of total uptake was always about one-third the proportion of total hydrolysis at the early time points. This 3:1 ratio of percent total hydrolysis to percent of cells immediately permeabilized was consistent among experimental conditions and cell types (3.2 ± 0.5, n = 19, p = 0.0003 by one-sample t-test, H0 = 1.0; condition not significant by one-way ANOVA, p = 0.52).
Fig. 4.

Percentage of cells susceptible to hydrolysis by sPLA2 assayed by propidium iodide fluorescence. The fluorescence intensity of propidium iodide for normal lymphocytes (A), Raji cells (B), and S49 cells (C) was measured during the ionomycin experiments of Fig. 3, and the data were averaged as explained in the legend to that figure (average: solid, SEM: dotted). Panel D: The percentages of cells already dead (white, PIA in Fig. 1 and Eq. 2A), killed by DMSO or Ionomycin (black, PIB in Fig. 1 and Eq. 2B), or alive, but susceptible to sPLA2 (gray PIC in Fig. 1 and Eq. 2C) were calculated as explained in Materials and Methods. Dotted lines and error bars represent the SEM for 3–10 independent samples. Panel E: Contour plot from flow cytometry analysis of Raji cells treated with 1.2 μM ionomycin showing numbers of cells staining negative for both annexin V binding and propidium iodide uptake (lower left, 19%); positive for annexin V, but negative for propidium iodide (upper left, 73%); positive for both probes (upper right, 7%); and negative for annexin V, but positive for propidium iodide (lower right, 1%). Panel F: Percentage of Raji cells originating from the upper left quadrant that stain positive for propidium iodide after incubation with sPLA2 for the indicated times. Cells were treated with normal (600 nM, black) or high (1.2 μM, red) ionomycin prior to addition of sPLA2.
The propidium iodide assay allowed us to use flow cytometry to determine whether the Raji cells hydrolyzed during the early phase of the time course were those that also had phosphatidylserine exposed. Figure 4E shows a sample plot for cells labeled with both propidium iodide and annexin. As expected from the data of Panels A–D, addition of sPLA2 resulted in conversion of cells from staining negative for propidium iodide to staining positive. Apparently, all of the cells that became permeable to propidium iodide originated from the population that stained positive for annexin binding (none migrated from annexin-negative: 5.1 ± 6.4%, n = 5, p = 0.47 by one-sample t-test, H0 = 0). Figure 4F shows the percentage of cells that have migrated from the propidium iodide-negative/annexin-positive field (upper left quadrant in Fig. 4E) to propidium iodide-positive during early hydrolysis by sPLA2 for both low and high ionomycin treatments.
3.3 Changes in membrane physical properties
The data of Figs. 3 and 4 demonstrated clearly that ionophore treatment still rendered Raji cells vulnerable to attack by sPLA2 (albeit at a lower rate compared to S49 cells) even though little increase in the amount of PS on the membrane surface was observed. We tested whether other relevant membrane changes induced by ionomycin (spacing and order of membrane lipids) were present in the Raji cells. We used two fluorescent membrane probes, MC540 and TMA-DPH, to assess these changes (Figs. 5 and 6).
Fig. 5.
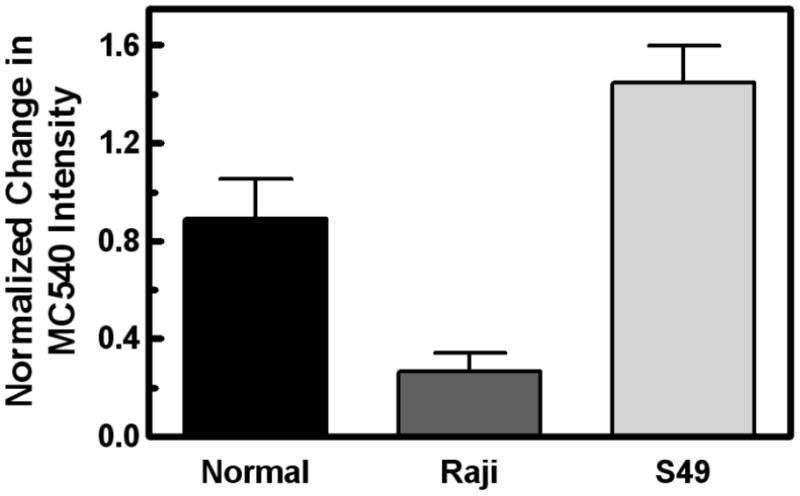
Effect of ionomycin treatment on MC540 fluorescence. The normalized change in intensity of MC540 fluorescence for normal lymphocytes (black), Raji cells (dark gray), and S49 cells (light gray) was calculated as the ratio of the intensity 10 min after ionomycin addition to the intensity before treatment minus 1 for 5–13 independent samples (error bars are SEM). The effect of cell type was significant (p < 0.0001) by one-way analysis of variance. A Bonferroni post-test revealed that the S49 samples were different from the other cell types (p < 0.05), which were indistinguishable from each other.
Fig. 6.
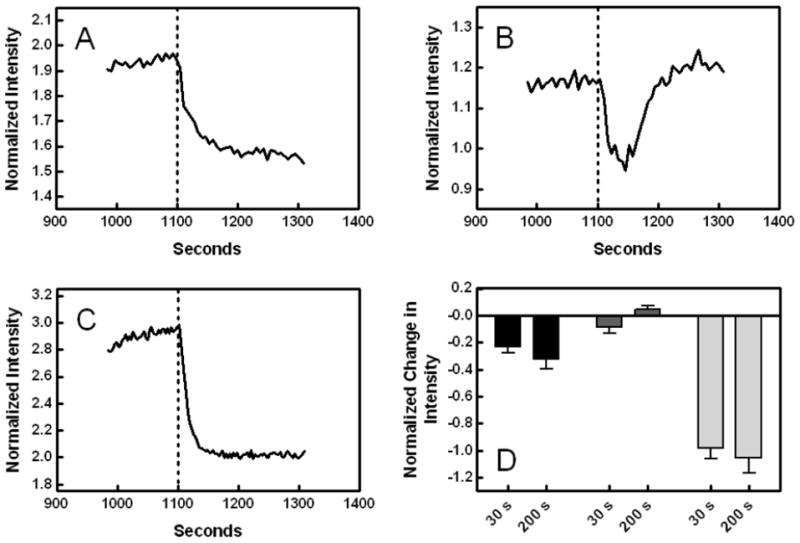
Effect of initial membrane hydrolysis on MC540 fluorescence intensity in ionomycin-treated samples. The fluorescence intensity of MC540 for normal lymphocytes (A), Raji cells (B), and S49 cells (C) was measured over time following addition of ionomycin. The data were normalized to the intensity prior to addition of ionomycin, and the traces shown in the figures begin approximately 600 s after that addition. Secretory PLA2 was added at the dotted lines. Panel D: The change in intensity at 30 s and again 200 s after addition of sPLA2 was calculated for each cell type (normal lymphocytes, black; Raji, dark gray; S49, light gray) as explained in the legend to Fig. 5. Only the effects of cell type (90.9% of the variation) were significant (p ≤ 0.0001) by two-way analysis of variance (n = 2–6). A Bonferroni post-test revealed that every pair-wise comparison between cell types at the 200 s time point was significant (p < 0.001), and that the 30 s point was indistinguishable between Raji and normal lymphocytes but different for S49 cells (p < 0.001).
Addition of ionomycin to S49 cells caused a 150% increase in the intensity of MC540 fluorescence (Fig. 5 and Ref. [7]). This intensity change has been shown previously to represent enhanced binding of the probe to the cell membrane [35]. This enhancement of binding occurs when the packing of membrane lipids has become looser resulting in greater interlipid distances [36]. The rise in MC540 emission intensity after ionomycin treatment for the normal lymphocytes was half the size of that observed with S49 cells. For Raji cells, the effect was even smaller, about one-fifth the size of the increment in S49 cells suggesting a link between exposure of PS and the complete effect of ionomycin on membrane properties detected by MC540. Nevertheless, the rise in MC540 fluorescence was statistically significant in each case (p < 0.03 for each by one-sample t-tests with H0 = 0). During hydrolysis by sPLA2, the rise in MC540 fluorescence observed in normal lymphocytes and S49 cells partially reversed (Figs. 6A and C). In contrast, the reversal was transient in Raji cells followed by a secondary rise that exceeded the MC540 intensity prior to addition of sPLA2 (Fig. 6B). These observations were reproducible as shown in Fig. 6D. Control experiments demonstrated that the drug solvent (DMSO) had no effect on MC540 intensity (not shown).
The apparent order of membrane lipids was assessed by TMA-DPH anisotropy; a decrease in anisotropy is generally interpreted as a reduction in order. As shown in Fig. 7, the initial TMA-DPH anisotropy prior to addition of ionomycin was highly reproducible for each of the three cell types. Although, this initial anisotropy appeared to vary among the cells, the only difference that was statistically significant was between the S49 cells (light gray) and the normal lymphocytes (black). For all three cell types, the effect of ionomycin to lower the anisotropy was significant. Furthermore, the decrement was greater for S49 cells than for either of the other two. Again, the effect was smallest with Raji cells. It has been suggested previously that ionophore treatment can cause cells to internalize TMA-DPH, an effect manifest by an increase in probe emission intensity [37]. In fact, with both S49 cells and normal lymphocytes, a reproducible rise in TMA-DPH intensity was observed upon addition of ionomycin (S49: 69.6 ± 2.2% enhancement, p < 0.0001, n = 5; lymphocytes: 37.9 ± 8.7%, p = 0.007, n = 6). This result raises the question of whether the observed change in anisotropy simply reflects uptake of the probe into a more fluid internal environment. Observations with the Raji cells, clarified this concern since enhanced intensity did not occur after stimulation with ionomycin suggesting that they did not internalize the probe (change in intensity = −1.1 ± 2.6%, p = 0.68, n = 7). Since the Raji cells still displayed a reproducible reduction in TMA-DPH anisotropy with ionophore treatment, we concluded that at least some of the result observed with all three cell types reflected disordering of the cell membrane. This conclusion was corroborated by previously-reported two-photon images with another probe of membrane order, laurdan [7,8]. This interesting result also suggests that the mechanism responsible for TMA-DPH uptake with ionophore treatment could involve the enzyme scramblase. Control experiments demonstrated that the drug solvent (DMSO) had no effect on TMA-DPH anisotropy (not shown).
Fig. 7.

Effect of ionomycin treatment on TMA-DPH anisotropy. The baseline anisotropy of TMA-DPH was assayed for normal lymphocytes (black), Raji cells (dark gray), and S49 cells (light gray) (“before”, error bars are the SEM, n = 5–7). The anisotropy was then re-assessed 10 min after addition of ionomycin. The overall effects of cell type (51.9% of the variation) and ionomycin (29.6% of the variation) were both significant (p ≤ 0.0001) by two-way analysis of variance. Interaction between cell type and drug treatment was small but significant, indicating that the size of the effect of ionomycin depended slightly on cell type (3.4% of the variation, p = 0.0006). A Bonferroni post-test revealed that S49 samples were different from the normal lymphocytes prior to ionomycin (p < 0.01) and that they were different were different from both of the other cell types (p < 0.001) after ionomycin treatment. No other pairwise comparisons between cell types were significant.
Discussion
The basic question of whether exposure of PS is necessary for cells to become vulnerable to sPLA2 was addressed by the data in this paper. The short answer to that question appears to be “no” because only 17% of the Raji cells displayed normal exposure of PS in the presence of ionomycin, even though 100% of the cells became susceptible to the action of sPLA2 (Fig. 2 and 4). Nevertheless, certain nuances in the data suggest complexities that bear further discussion on this point.
As shown in Fig. 3, the time profile for hydrolysis by sPLA2 was more complicated with Raji cells compared to normal lymphocytes or S49 cells. In the latter two cases, the time profile could be described exactly (r2 = 0.999 for human lymphocytes and 1.000 for S49 cells) as the sum of two exponential (pseudo first-order) processes, presumably reflecting diversity of substrate pools in the sample. For human lymphocytes, the faster component represented 59% of the hydrolysis events with a first-order rate constant of 0.045 s−1. The slower component had a first-order rate constant of 0.002 s−1. Comparable results were identified with S49 cells, although at much higher rates (fast component: 57% at 0.38 s−1 and slow component at 0.052 s−1). This type of kinetic profile also indicated that the process that made the membranes vulnerable to hydrolysis was accomplished prior to addition of enzyme (i.e. within 10 min incubation with ionomycin) and that only substrate availability was limiting in the reaction thereafter. In contrast, the data obtained from Raji cells could not be rationalized as simple first-order processes. Instead, the time profile displayed a sharp initial hydrolysis representing about 45% of the total lipid hydrolyzed. This initial hydrolysis appeared to be completed by about 70 s with a second phase that began after a delay and displayed a modest acceleration. Such a shape implies two populations of cells: a small percentage fully susceptible to the enzyme (about 28%; summed from initial phase in Fig. 4B and black + white bar segments in Fig. 4D) and a second population for which hydrolytic capacity grows latently during the time course. Based on the proportion of hydrolysis represented by each phase, it appeared that the number of lipids hydrolyzed per cell was about three times greater for the initially-vulnerable subpopulation.
Statistical comparison of the data of Figs. 2 and 4 suggested that the subpopulation of Raji cells incorporating propidium iodide during the first hydrolysis phase could be synonymous with those that exposed PS in the presence of ionomycin (p = 0.40, n = 4–9 by unpaired t-test). Moreover, flow cytometry analysis verified that this subpopulation of cells vulnerable to immediate attack by sPLA2 originated entirely from cells staining positive for annexin and therefore exposing PS. Finally, when additional PS was exposed by treatment of Raji cells with a higher dose of ionomycin (Fig. 2B), the subpopulation attacked by sPLA2 during the initial phase also rose (Fig. 4F). The number of cells incorporating propidium iodide during hydrolysis observed by flow cytometry in Fig. 4F was lower than that implied by Fig. 4B due to competition between the annexin V probe and sPLA2 [38]. Nevertheless, those cells that did not bind annexin V would not be subject to competition, and none of them were attacked by sPLA2 during the initial hydrolysis phase. Hence, we concluded that the cells initially vulnerable to sPLA2 and hydrolyzed at the highest rate in Raji cells were those that displayed PS on the outer leaflet of the plasma membrane.
A similar match between the number of Raji cells showing exposed PS and vulnerable to sPLA2 was observed under control conditions (p = 0.48). Previous analysis demonstrated that hydrolysis by sPLA2 in control samples represents elimination of a cohort of cells already dead in culture or damaged during sample preparation [10]. Thus, the initial rates of hydrolysis in the control groups in Fig. 3 probably pertained entirely to those control cells that stained positive for annexin V in Fig. 2 and also incorporated propidium iodide in Fig. 4. Under such conditions, the initial hydrolysis rate in Raji cells was indistinguishable from those observed in human lymphocytes and S49 cells (p = 0.34 by analysis of variance). When cells were stimulated with ionomycin, however, nearly all the S49 cells and normal lymphocytes, but few of the Raji cells, stained positive for PS exposure. This event corresponded to a substantial rise in initial hydrolysis rate in S49 cells compared to the other two cell types. Hence, the observation that Raji cells were hydrolyzed at a rate similar to S49 cells only when cell surface PS was expressed argues that the presence of the anionic lipid is indeed important for efficient hydrolysis of cell membranes. In contrast, the result that hydrolysis was not stimulated in normal lymphocytes to the same level as in S49 cells even though PS was fully exposed in both cell types indicated that PS expression alone is insufficient.
The disproportionality between fatty acid release (ADIFAB) and the number of cells permeablized to propidium iodide during the initial phase of Raji cell hydrolysis was unexpected. The small cohort of cells in control samples already dead before introduction of sPLA2 showed the same ~3:1 disproportion of hydrolysis to permeabilization (3.93 ± 0.82, n = 10, p = 0.006 by one-sample t-test, H0 = 1.0). Hence, subpopulations of cells must differ in the amount of hydrolysis that occurs, perhaps reflecting varying degrees of accessibility of internal membranes to the enzyme. The assumption is that those cells that are already dead or have exposed PS in Raji samples are “super-susceptible” relative to the others such that hydrolysis of the plasma membrane produces sufficient damage for the enzyme to interact with internal membranes. The plasma membranes of the remaining cells in the Raji samples, would then be hydrolyzed to a lesser extent, sufficient to allow propidium iodide entry but still able to exclude proteins. As evidence of this idea, we have found from microscopy images after exposure of vulnerable samples to sPLA2 that most cells remain intact, even though they have become permeable to propidium iodide, consistent with the fact that they remain detectable as propidium iodide-positive entities of appropriate size by flow cytometry (as in Fig. 4).
The changes in interlipid spacing and membrane order assessed in Figs. 5 and 7 have been suggested to be essential for cells to become susceptible to sPLA2 [7,8,39,40]. The data shown here would argue the same since the cells hydrolyzed at the highest rate (S49) also displayed the largest alteration in MC540 and TMA-DPH fluorescence upon addition of ionomycin. In fact, a strong relationship between average ionomycin-stimulated hydrolysis rate and the increment of change in the emission of these probes was observed across all three cell types (p ≤ 0.0003 for both probes based on linear regression). Thus, it appeared that optimal hydrolysis depended on both the presence of PS and reduced interlipid interactions in the cell membrane. A similar conclusion was reached previously from less direct observations of the timing of PS exposure and vulnerability to hydrolysis during glucocorticoid-stimulated apoptosis [10].
The second phase of hydrolysis observed in the Raji cells was unexpected. In a simple sense, it revealed that the cells could be completely eliminated by sPLA2 even though initial ionomycin-stimulated PS exposure was minimal. The hydrolytic acceleration associated with this phase also demonstrated that some latent change must be occurring in the Raji membranes that further encourages hydrolysis after those cells with exposed PS have been exhausted. One possibility is that anionic lipid becomes available on the cell surface as damage from hydrolysis progresses. Assessing whether additional PS is exposed during this latent hydrolysis phase is complicated by the fact that sPLA2 and annexin V compete with each other for binding to the cell surface [38]. Nonetheless, as shown in Fig. 4F, hydrolysis in the presence of annexin V still proceeded, albeit at a lower rate, suggesting that any additional exposure of PS should also be detectable even though the exact amount would be underestimated. However, from the annexin staining observed during the experiments of Figs. 4E and F, there was no evidence for increased exposure of PS during incubation with sPLA2 (increase in annexin-positive cells after 10 min hydrolysis = −12 ± 18%, n = 4, p = 0.24; also p = 0.58 by linear regression of change in annexin-positive cells during 1–10 min hydrolysis time points, n = 10).
What we can say is that the membranes of Raji cells became increasingly perturbed during this secondary phase of hydrolysis as assessed by MC540 fluorescence. Ordinarily, as observed in both S49 cells and normal lymphocytes, the calcium-induced MC540 fluorescence change illustrated in Fig. 5 regresses during hydrolysis by sPLA2 probably because of membrane shrinkage as shown previously (Figs. 6A and C, Ref. [8]). Raji cells behaved similarly during the first 30 s after sPLA2 addition, but in contrast to the other cell types, the fluorescence trend thereafter reversed direction and instead increased, reaching a plateau after 200 s (Fig. 6B). The timing of this plateau corresponded to the moment of acceleration of hydrolysis illustrated in Fig. 3B. Afterwards, the MC540 fluorescence continued to rise in synchrony with propidium iodide uptake shown in Fig. 4B while it remained low in the other cell types (not shown). The biophysical reasons for this effect on MC540 fluorescence are unknown, but they could relate to accumulation of reaction products in the membrane, which is known to accelerate hydrolysis in artificial bilayers that are not already fully susceptible [13].
In addition to addressing the potential role of PS exposure in modulating the action of sPLA2, this report provides information regarding the relevance of studies from cultured lymphoma cells to normal human lymphocytes. Prior to this study, all data suggesting that nucleated cells become vulnerable to hydrolysis by sPLA2 during biochemically-programmed death involved cultured transformed cells. The data in this report make it clear that those studies with cultured cell lines were relevant to normal human lymphocytes. Nevertheless, as shown by the data of Figs. 3, 5, and 7, both the rate of hydrolysis and the extent of relevant physical changes to the cell membrane were substantially less in human lymphocytes compared to S49 cells. This observation raises the question of whether there is a difference between normal and transformed cells or whether these parameters simply vary depending on the source of the cells (i.e. mouse vs. human). The answer to that question requires inductive logic and therefore future comparisons among many examples. Nonetheless, preliminary findings suggest the intriguing possibility that dying tumor cells that are not deficient in their ability to expose PS are generally more vulnerable to hydrolysis by sPLA2 than are their normal counterparts. Data published from our lab several years ago contained results from another human transformed lymphocyte cell line that supported this notion (MOLT-4, acute lymphoblastic leukemia) [5]. The rate of hydrolysis of this cell line during ionomycin-induced cell death was comparable to that observed with S49 cells (0.058 GP units · s−1, 68% of the S49 cell rate) and therefore a factor of about three times greater than for normal lymphocytes.
There are two important implications of these findings. First, although the presence of PS on the surface of cell membranes is an important marker for stress responses such as blood clotting and clearance of dead cells by macrophages [24,26,27,41,42], it appears to be a secondary signal for hydrolysis of dying cells by sPLA2. As mentioned previously, this ability of some sPLA2 isoforms to recognize dying cells prior to or in the absence of PS exposure may be significant for the process of recruitment and activation of macrophages. Apparently, there are two steps to that process, an initial “find-me” signal for recruitment followed by an “eat-me” signal to specify which cells will be cleared [43,44]. While the eat-me signal almost surely represents the presence of PS on the external face of the cell membrane, the find-me signal may be accomplished by the release of one of several small molecules from the dying cell [43–47]. One of these molecules is the phospholipid hydrolysis product lysophospholipid, which would be released by the action of sPLA2 [44,46,47]. Since limited hydrolysis appears to be possible in the absence of PS exposure, sPLA2 would be able to help generate the find-me signal in dying cells before it becomes capable of destroying their membranes. Thus, the enzyme may perform a priming function to help initiate clearance of dying cells so that macrophages are present and ready when PS is exposed. Although sPLA2 would be now able to hydrolyze the membrane fully and release inflammatory mediators in large quantities, the cell removal by macrophages would mitigate that effect. Recent work indicates that this hypothetical priming function would most likely to be accomplished by the group X isoform of sPLA2 [10,17].
The second implication of this study is the possibility that death-induced vulnerability to sPLA2, while relevant to normal human lymphocytes, may be quantitatively more significant for tumor cells. This observation, if verified, has potential therapeutic implications, both positive and negative. On the positive side, it may suggest a novel treatment strategy that could take advantage of the enzyme’s lethal action. However, the attack of susceptible cells by sPLA2 during chemotherapy could produce an undesired inflammatory response.
Highlights.
Phosphatidylserine exposure is not an absolute requirement for membrane hydrolysis by secretory phospholipase A2
Dying lymphoma cells are more susceptible to hydrolysis by secretory phospholipase A2 than their normal human counterparts
Efficient hydrolysis of cell membranes correlates best with physical changes in the membrane that occur as cells proceed through ionomycin-induced cell death
Acknowledgments
The authors express appreciation to Mary Keller (BYU) for providing fresh samples of Raji cells and to Michael Gelb (University of Washington) and Wonhwa Cho (University of Illinois, Chicago) for generously providing samples of human sPLA2 isoforms. This work was supported by the National Institutes of Health (GM073997).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Murakami M, Taketomi Y, Miki Y, Sato H, Hirabayashi T, Yamamoto K. Recent progress in phospholipase A research: from cells to animals to humans. Prog Lipid Res. 2011;50:152–192. doi: 10.1016/j.plipres.2010.12.001. [DOI] [PubMed] [Google Scholar]
- 2.Beers SA, Buckland AG, Koduri RS, Cho W, Gelb MH, Wilton DC. The antibacterial properties of secreted phospholipases A2: a major physiological role for the group IIA enzyme that depends on the very high pI of the enzyme to allow penetration of the bacterial cell wall. J Biol Chem. 2002;277:1788–1793. doi: 10.1074/jbc.M109777200. [DOI] [PubMed] [Google Scholar]
- 3.Koduri RS, Gronroos JO, Laine VJ, Le Calvez C, Lambeau G, Nevalainen TJ, Gelb MH. Bactericidal properties of human and murine groups I, II, V, X, and XII secreted phospholipases A(2) J Biol Chem. 2002;277:5849–5857. doi: 10.1074/jbc.M109699200. [DOI] [PubMed] [Google Scholar]
- 4.Atsumi G, Murakami M, Tajima M, Shimbara S, Hara N, Kudo I. The perturbed membrane of cells undergoing apoptosis is susceptible to type II secretory phospholipase A2 to liberate arachidonic acid. Biochim Biophys Acta. 1997;1349:43–54. doi: 10.1016/s0005-2760(97)00082-9. [DOI] [PubMed] [Google Scholar]
- 5.Wilson HA, Waldrip JB, Nielson KH, Judd AM, Han SK, Cho W, Sims PJ, Bell JD. Mechanisms by which elevated intracellular calcium induces S49 cell membranes to become susceptible to the action of secretory phospholipase A2. J Biol Chem. 1999;274:11494–11504. doi: 10.1074/jbc.274.17.11494. [DOI] [PubMed] [Google Scholar]
- 6.Smith SK, Farnbach AR, Harris FM, Hawes AC, Jackson LR, Judd AM, Vest RS, Sanchez S, Bell JD. Mechanisms by which intracellular calcium induces susceptibility to secretory phospholipase A2 in human erythrocytes. J Biol Chem. 2001;276:22732–22741. doi: 10.1074/jbc.M010880200. [DOI] [PubMed] [Google Scholar]
- 7.Bailey RW, Olson ED, Vu MP, Brueseke TJ, Robertson L, Christensen RE, Parker KH, Judd AM, Bell JD. Relationship between Membrane Physical Properties and Secretory Phospholipase A2 Hydrolysis Kinetics in S49 Cells during Ionophore-Induced Apoptosis. Biophys J. 2007;93:2350–2362. doi: 10.1529/biophysj.107.104679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Bailey RW, Nguyen T, Robertson L, Gibbons E, Nelson J, Christensen RE, Bell JP, Judd AM, Bell JD. Sequence of physical changes to the cell membrane during glucocorticoid-induced apoptosis in S49 lymphoma cells. Biophys J. 2009;96:2709–2718. doi: 10.1016/j.bpj.2008.12.3925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Nielson KH, Olsen CA, Allred DV, O’Neill KL, Burton GF, Bell JD. Susceptibility of S49 lymphoma cell membranes to hydrolysis by secretory phospholipase A(2) during early phase of apoptosis. Biochim Biophys Acta. 2000;1484:163–174. doi: 10.1016/s1388-1981(00)00004-4. [DOI] [PubMed] [Google Scholar]
- 10.Olson ED, Nelson J, Griffith K, Nguyen T, Streeter M, Wilson-Ashworth HA, Gelb MH, Judd AM, Bell JD. Kinetic evaluation of cell membrane hydrolysis during apoptosis by human isoforms of secretory phospholipase A2. J Biol Chem. 2010;285:10993–11002. doi: 10.1074/jbc.M109.070797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Jain MK, Yu BZ, Kozubek A. Binding of phospholipase A2 to zwitterionic bilayers is promoted by lateral segregation of anionic amphiphiles. Biochim Biophys Acta. 1989;980:23–32. doi: 10.1016/0005-2736(89)90195-8. [DOI] [PubMed] [Google Scholar]
- 12.Singer AG, Ghomashchi F, Le Calvez C, Bollinger J, Bezzine S, Rouault M, Sadilek M, Lazdunski M, Lambeau G, Gelb MH. Interfacial kinetic and binding properties of the complete set of human and mouse groups I, II, V, X, and XII secreted phospholipases A2. J Biol Chem. 2002;277:48535–48549. doi: 10.1074/jbc.M205855200. [DOI] [PubMed] [Google Scholar]
- 13.Henshaw JB, Olsen CA, Farnbach AR, Nielson KH, Bell JD. Definition of the specific roles of lysolecithin and palmitic acid in altering the susceptibility of dipalmitoylphosphatidylcholine bilayers to phospholipase A2. Biochemistry. 1998;37:10709–10721. doi: 10.1021/bi9728809. [DOI] [PubMed] [Google Scholar]
- 14.Harris FM, Smith SK, Bell JD. Physical properties of erythrocyte ghosts that determine susceptibility to secretory phospholipase A2. J Biol Chem. 2001;276:22722–22731. doi: 10.1074/jbc.M010879200. [DOI] [PubMed] [Google Scholar]
- 15.Han SK, Kim KP, Koduri R, Bittova L, Munoz NM, Leff AR, Wilton DC, Gelb MH, Cho W. Roles of Trp31 in high membrane binding and proinflammatory activity of human group V phospholipase A2. J Biol Chem. 1999;274:11881–11888. doi: 10.1074/jbc.274.17.11881. [DOI] [PubMed] [Google Scholar]
- 16.Nelson J, Gibbons E, Pickett KR, Streeter M, Warcup AO, Yeung CH, Judd AM, Bell JD. Relationship between membrane permeability and specificity of human secretory phospholipase A(2) isoforms during cell death. Biochim Biophys Acta. 2011;1808:1913–1920. doi: 10.1016/j.bbamem.2011.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Bezzine S, Koduri RS, Valentin E, Murakami M, Kudo I, Ghomashchi F, Sadilek M, Lambeau G, Gelb MH. Exogenously added human group X secreted phospholipase A(2) but not the group IB, IIA, and V enzymes efficiently release arachidonic acid from adherent mammalian cells. J Biol Chem. 2000;275:3179–3191. doi: 10.1074/jbc.275.5.3179. [DOI] [PubMed] [Google Scholar]
- 18.Bezzine S, Bollinger JG, Singer AG, Veatch SL, Keller SL, Gelb MH. On the binding preference of human groups IIA and X phospholipases A2 for membranes with anionic phospholipids. J Biol Chem. 2002;277:48523–48534. doi: 10.1074/jbc.M203137200. [DOI] [PubMed] [Google Scholar]
- 19.Murakami M, Kambe T, Shimbara S, Higashino K, Hanasaki K, Arita H, Horiguchi M, Arita M, Arai H, Inoue K, Kudo I. Different functional aspects of the group II subfamily (Types IIA and V) and type X secretory phospholipase A(2)s in regulating arachidonic acid release and prostaglandin generation. Implications of cyclooxygenase-2 induction and phospholipid scramblase-mediated cellular membrane perturbation. J Biol Chem. 1999;274:31435–31444. doi: 10.1074/jbc.274.44.31435. [DOI] [PubMed] [Google Scholar]
- 20.Scott DL, White SP, Otwinowski Z, Yuan W, Gelb MH, Sigler PB. Interfacial catalysis: the mechanism of phospholipase A2. Science. 1990;250:1541–1546. doi: 10.1126/science.2274785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kinkaid AR, Wilton DC. The effect of anions on interfacial binding and activation of secretory phospholipase A2. Biochem Soc Trans. 1995;23:556S. doi: 10.1042/bst023556s. [DOI] [PubMed] [Google Scholar]
- 22.Diraviyam K, Murray D. Computational analysis of the membrane association of group IIA secreted phospholipases A2: a differential role for electrostatics. Biochemistry. 2006;45:2584–2598. doi: 10.1021/bi051901t. [DOI] [PubMed] [Google Scholar]
- 23.Bevers EM, Comfurius P, Zwaal RF. Regulatory mechanisms in maintenance and modulation of transmembrane lipid asymmetry: pathophysiological implications. Lupus. 1996;5:480–487. doi: 10.1177/096120339600500531. [DOI] [PubMed] [Google Scholar]
- 24.Fadeel B. Plasma membrane alterations during apoptosis: Role in corpse clearance. Antioxidants & Redox Signaling. 2004;6:269–275. doi: 10.1089/152308604322899332. [DOI] [PubMed] [Google Scholar]
- 25.Williamson P, Christie A, Kohlin T, Schlegel RA, Comfurius P, Harmsma M, Zwaal RF, Bevers EM. Phospholipid scramblase activation pathways in lymphocytes. Biochemistry. 2001;40:8065–8072. doi: 10.1021/bi001929z. [DOI] [PubMed] [Google Scholar]
- 26.Peter C, Wesselborg S, Lauber K. Molecular suicide notes: last call from apoptosing cells. J Mol Cell Biol. 2010;2:78–80. doi: 10.1093/jmcb/mjp045. [DOI] [PubMed] [Google Scholar]
- 27.Callahan MK, Williamson P, Schlegel RA. Surface expression of phosphatidylserine on macrophages is required for phagocytosis of apoptotic thymocytes. Cell Death Differ. 2000;7:645–653. doi: 10.1038/sj.cdd.4400690. [DOI] [PubMed] [Google Scholar]
- 28.Krahling S, Callahan MK, Williamson P, Schlegel RA. Exposure of phosphatidylserine is a general feature in the phagocytosis of apoptotic lymphocytes by macrophages. Cell Death Differ. 1999;6:183–189. doi: 10.1038/sj.cdd.4400473. [DOI] [PubMed] [Google Scholar]
- 29.Zhao J, Zhou Q, Wiedmer T, Sims PJ. Level of expression of phospholipid scramblase regulates induced movement of phosphatidylserine to the cell surface. J Biol Chem. 1998;273:6603–6606. doi: 10.1074/jbc.273.12.6603. [DOI] [PubMed] [Google Scholar]
- 30.Tepper AD, Ruurs P, Wiedmer T, Sims PJ, Borst J, Van Blitterswijk WJ. Sphingomyelin hydrolysis to ceramide during the execution phase of apoptosis results from phospholipid scrambling and alters cell-surface morphology. J Cell Biol. 2000;150:155–164. doi: 10.1083/jcb.150.1.155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kagan VE, Gleiss B, Tyurina YY, Tyurin VA, Elenstrom-Magnusson C, Liu SX, Serinkan FB, Arroyo A, Chandra J, Orrenius S, Fadeel B. A role for oxidative stress in apoptosis: oxidation and externalization of phosphatidylserine is required for macrophage clearance of cells undergoing Fas-mediated apoptosis. J Immunol. 2002;169:487–499. doi: 10.4049/jimmunol.169.1.487. [DOI] [PubMed] [Google Scholar]
- 32.Maraganore JM, Merutka G, Cho W, Welches W, Kezdy FJ, Heinrikson RL. A new class of phospholipases A2 with lysine in place of aspartate 49. Functional consequences for calcium and substrate binding. J Biol Chem. 1984;259:13839–13843. [PubMed] [Google Scholar]
- 33.Richieri GV, Kleinfeld AM. Continuous measurement of phospholipase A2 activity using the fluorescent probe ADIFAB. Anal Biochem. 1995;229:256–263. doi: 10.1006/abio.1995.1410. [DOI] [PubMed] [Google Scholar]
- 34.Wilson HA, Huang W, Waldrip JB, Judd AM, Vernon LP, Bell JD. Mechanisms by which thionin induces susceptibility of S49 cell membranes to extracellular phospholipase A2. Biochim Biophys Acta. 1997;1349:142–156. doi: 10.1016/s0005-2760(97)00089-1. [DOI] [PubMed] [Google Scholar]
- 35.Lagerberg JW, Kallen KJ, Haest CW, VanSteveninck J, Dubbelman TM. Factors affecting the amount and the mode of merocyanine 540 binding to the membrane of human erythrocytes. A comparison with the binding to leukemia cells. Biochim Biophys Acta. 1995;1235:428–436. doi: 10.1016/0005-2736(95)80032-b. [DOI] [PubMed] [Google Scholar]
- 36.Stillwell W, Wassall SR, Dumaual AC, Ehringer WD, Browning CW, Jenski LJ. Use of merocyanine (MC540) in quantifying lipid domains and packing in phospholipid vesicles and tumor cells. Biochim Biophys Acta. 1993;1146:136–144. doi: 10.1016/0005-2736(93)90348-4. [DOI] [PubMed] [Google Scholar]
- 37.Kuhry JG, Fonteneau P, Duportail G, Maechling C, Laustriat G. Tma-Dph - A Suitable Fluorescence Polarization Probe for Specific Plasma-Membrane Fluidity Studies in Intact Living Cells. Cell Biophysics. 1983;5:129–140. doi: 10.1007/BF02796139. [DOI] [PubMed] [Google Scholar]
- 38.Buckland AG, Wilton DC. Inhibition of secreted phospholipases A2 by annexin V. Competition for anionic phospholipid interfaces allows an assessment of the relative interfacial affinities of secreted phospholipases A2. Biochim Biophys Acta. 1998;1391:367–376. doi: 10.1016/s0005-2760(98)00026-5. [DOI] [PubMed] [Google Scholar]
- 39.Best KB, Ohran AJ, Hawes AC, Hazlett TL, Gratton E, Judd AM, Bell JD. Relationship between erythrocyte membrane phase properties and susceptibility to secretory phospholipase A2. Biochemistry. 2002;41:13982–13988. doi: 10.1021/bi026796r. [DOI] [PubMed] [Google Scholar]
- 40.Jensen LB, Burgess NK, Gonda DD, Spencer E, Wilson-Ashworth HA, Driscoll E, Vu MP, Fairbourn JL, Judd AM, Bell JD. Mechanisms governing the level of susceptibility of erythrocyte membranes to secretory phospholipase A2. Biophys J. 2005;88:2692–2705. doi: 10.1529/biophysj.104.056457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Peter C, Wesselborg S, Lauber K. Apoptosis: opening PANdora’s BoX. Curr Biol. 2010;20:R940–R942. doi: 10.1016/j.cub.2010.09.066. [DOI] [PubMed] [Google Scholar]
- 42.Peter C, Wesselborg S, Herrmann M, Lauber K. Dangerous attraction: phagocyte recruitment and danger signals of apoptotic and necrotic cells. Apoptosis. 2010;15:1007–1028. doi: 10.1007/s10495-010-0472-1. [DOI] [PubMed] [Google Scholar]
- 43.Chekeni FB, Elliott MR, Sandilos JK, Walk SF, Kinchen JM, Lazarowski ER, Armstrong AJ, Penuela S, Laird DW, Salvesen GS, Isakson BE, Bayliss DA, Ravichandran KS. Pannexin 1 channels mediate ‘find-me’ signal release and membrane permeability during apoptosis. Nature. 2010;467:863–867. doi: 10.1038/nature09413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Peter C, Waibel M, Radu CG, Yang LV, Witte ON, Schulze-Osthoff K, Wesselborg S, Lauber K. Migration to apoptotic “find-me” signals is mediated via the phagocyte receptor G2A. J Biol Chem. 2008;283:5296–5305. doi: 10.1074/jbc.M706586200. [DOI] [PubMed] [Google Scholar]
- 45.Chekeni FB, Ravichandran KS. The role of nucleotides in apoptotic cell clearance: implications for disease pathogenesis. J Mol Med. 2011;89:13–22. doi: 10.1007/s00109-010-0673-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Lauber K, Bohn E, Krober SM, Xiao YJ, Blumenthal SG, Lindemann RK, Marini P, Wiedig C, Zobywalski A, Baksh S, Xu Y, Autenrieth IB, Schulze-Osthoff K, Belka C, Stuhler G, Wesselborg S. Apoptotic cells induce migration of phagocytes via caspase-3-mediated release of a lipid attraction signal. Cell. 2003;113:717–730. doi: 10.1016/s0092-8674(03)00422-7. [DOI] [PubMed] [Google Scholar]
- 47.Mueller RB, Sheriff A, Gaipl US, Wesselborg S, Lauber K. Attraction of phagocytes by apoptotic cells is mediated by lysophosphatidylcholine. Autoimmunity. 2007;40:342–344. doi: 10.1080/08916930701356911. [DOI] [PubMed] [Google Scholar]