

Abstract
Wild-type Chlamydomonas reinhardtii cells shifted
from high concentrations (5%) of CO2 to low, ambient
levels (0.03%) rapidly increase transcription of mRNAs from several
CO2-responsive genes. Simultaneously, they develop a
functional carbon concentrating mechanism that allows the cells to
greatly increase internal levels of CO2 and
HCO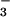 . The cia5 mutant is defective in
all of these phenotypes. A newly isolated gene, designated
Cia5, restores transformed cia5 cells to
the phenotype of wild-type cells. The 6,481-bp gene produces a 5.1-kb
mRNA that is present constitutively in light in high and low
CO2 both in wild-type cells and the cia5
mutant. It encodes a protein that has features of a putative
transcription factor and that, likewise, is present constitutively in
low and high CO2 conditions. Complementation of
cia5 can be achieved with a truncated
Cia5 gene that is missing the coding information for 54
C-terminal amino acids. Unlike wild-type cells or cia5
mutants transformed with an intact Cia5 gene,
cia5 mutants complemented with the truncated gene
exhibit constitutive synthesis of mRNAs from CO2-responsive
genes in light under both high and low CO2 conditions.
These discoveries suggest that posttranslational changes to the
C-terminal domain control the ability of CIA5 to act as an inducer and
directly or indirectly control transcription of
CO2-responsive genes. Thus, CIA5 appears to be a master
regulator of the carbon concentrating mechanism and is intimately
involved in the signal transduction mechanism that senses and allows
immediate responses to fluctuations in environmental CO2
and HCO
. The cia5 mutant is defective in
all of these phenotypes. A newly isolated gene, designated
Cia5, restores transformed cia5 cells to
the phenotype of wild-type cells. The 6,481-bp gene produces a 5.1-kb
mRNA that is present constitutively in light in high and low
CO2 both in wild-type cells and the cia5
mutant. It encodes a protein that has features of a putative
transcription factor and that, likewise, is present constitutively in
low and high CO2 conditions. Complementation of
cia5 can be achieved with a truncated
Cia5 gene that is missing the coding information for 54
C-terminal amino acids. Unlike wild-type cells or cia5
mutants transformed with an intact Cia5 gene,
cia5 mutants complemented with the truncated gene
exhibit constitutive synthesis of mRNAs from CO2-responsive
genes in light under both high and low CO2 conditions.
These discoveries suggest that posttranslational changes to the
C-terminal domain control the ability of CIA5 to act as an inducer and
directly or indirectly control transcription of
CO2-responsive genes. Thus, CIA5 appears to be a master
regulator of the carbon concentrating mechanism and is intimately
involved in the signal transduction mechanism that senses and allows
immediate responses to fluctuations in environmental CO2
and HCO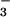 concentrations.
concentrations.
Prokaryotic and eukaryotic
algae are responsible for a significant portion of global
photosynthesis and carbon cycling (1). For most algae a carbon
concentrating mechanism (CCM) is essential for photosynthesis (2–5).
This mechanism enables algae to accumulate intracellular concentrations
of CO2 and HCO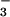 that are much
in excess of the CO2 and HCO
that are much
in excess of the CO2 and HCO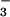 concentrations in their often inorganic carbon
(Ci)-deprived aqueous environments. The
cia5 mutant of Chlamydomonas reinhardtii lacks
the ability to acquire CO2 and
HCO
concentrations in their often inorganic carbon
(Ci)-deprived aqueous environments. The
cia5 mutant of Chlamydomonas reinhardtii lacks
the ability to acquire CO2 and
HCO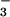 and must be maintained under high
CO2 conditions (i.e., 5%
CO2). It also lacks the ability to support the
formation of the CCM and the activation of several genes that are
induced (or derepressed) when wild-type C. reinhardtii cells
are transferred from an environment of high CO2
to ambient (i.e., 0.03%) levels of CO2 (reviewed
in refs. 4 and 5). Levels of mRNAs encoding most, if not all, of the
proteins that are “induced” under low CO2
show a rapid increase soon after transfer of cells to limiting
CO2 concentrations. Among the presently
identified proteins whose synthesis increases markedly during a shift
to low CO2 concentrations are: the major
periplasmic carbonic anhydrase (CA) (pCA1 encoded by the
Cah1 gene); two identical mitochondrial CAs (mtCA1 and mtCA2
encoded by the two highly homologous Mca1 and
Mca2 genes); Ccp1 and Ccp2 (also known as LIP36 and encoded
by the Ccp1 and Ccp2 genes); a chloroplast inner
envelope protein, Lci1 (a protein of unknown function encoded by the
Lci1 gene); a chloroplast stromal protein, GS2 (encoded by
the Gs2 gene); AlaAT, an alanine:α-ketoglutarate
aminotransferase (encoded by the Att1 gene); and two
photorespiratory enzymes, a glycolate dehydrogenase and a
phosphoglycolate phosphatase, whose genes have not been isolated
(reviewed in refs. 4 and 5).
and must be maintained under high
CO2 conditions (i.e., 5%
CO2). It also lacks the ability to support the
formation of the CCM and the activation of several genes that are
induced (or derepressed) when wild-type C. reinhardtii cells
are transferred from an environment of high CO2
to ambient (i.e., 0.03%) levels of CO2 (reviewed
in refs. 4 and 5). Levels of mRNAs encoding most, if not all, of the
proteins that are “induced” under low CO2
show a rapid increase soon after transfer of cells to limiting
CO2 concentrations. Among the presently
identified proteins whose synthesis increases markedly during a shift
to low CO2 concentrations are: the major
periplasmic carbonic anhydrase (CA) (pCA1 encoded by the
Cah1 gene); two identical mitochondrial CAs (mtCA1 and mtCA2
encoded by the two highly homologous Mca1 and
Mca2 genes); Ccp1 and Ccp2 (also known as LIP36 and encoded
by the Ccp1 and Ccp2 genes); a chloroplast inner
envelope protein, Lci1 (a protein of unknown function encoded by the
Lci1 gene); a chloroplast stromal protein, GS2 (encoded by
the Gs2 gene); AlaAT, an alanine:α-ketoglutarate
aminotransferase (encoded by the Att1 gene); and two
photorespiratory enzymes, a glycolate dehydrogenase and a
phosphoglycolate phosphatase, whose genes have not been isolated
(reviewed in refs. 4 and 5).
Because the cia5 mutant lacks the ability to support a
functional CCM and rapid activation of several
CO2-responsive genes when shifted to low
CO2 concentrations, it has been suggested that
the cia5 mutant might be defective in the regulation of
genes and/or signal transduction pathways associated with the CCM
(4–6). Here we report the cloning of a gene, Cia5, that
complements the cia5 mutant and restores the ability of the
cell to develop a functional CCM. We provide evidence that the CIA5
protein is a master regulatory element that acts as an inducer in
controlling transcription of one or more
CO2-responsive genes that are essential to the
function of the CCM in C. reinhardtii. We report cloning and
characterization of a gene involved in the signal transduction pathways
that allow photosynthetic organisms to sense and respond to changes in
CO2 and HCO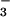 concentrations in
their environment.
concentrations in
their environment.
Materials and Methods
Strains and Growth Conditions.
Wild-type CC-125 originally was obtained from the Chlamydomonas Stock Center, Duke University, Durham, NC. The cell wall-deficient, high CO2-requiring cia5 double mutant was obtained from James Moroney (Louisiana State University, Baton Rouge) and was maintained in Tris acetate phosphate (TAP) medium (7). Liquid cultures were grown on a rotary shaker in white light (≈100 μmol/sec per m2) at 26°C. For experiments in which cells were shifted from high CO2 to low CO2 conditions, cells were cultured in TP medium (TAP lacking acetate) in 5% CO2 conditions to a density of ≈2 × 106 cells/ml and then shifted to ambient CO2 conditions for various times.
Genomic Complementation, Gene Isolation, and Characterization.
The mutant, cia5, was transformed with 120 DNA pools isolated from an indexed cosmid library of C. reinhardtii DNA (8). Recipient cells were resuspended at a cell density of 4 × 108 cells/ml in 250 μl TAP medium containing 60 mM sucrose and electroporated with cosmid DNA under conditions (25 μF, 1875 V/cm) described by Shimogawara et al. (10). Transformed cells were allowed to recover in 50 ml TAP medium containing 60 mM sucrose for 24 h in the light with shaking. Cells were collected by centrifugation, resuspended in TP medium (TAP lacking acetate), plated in the presence of 10% corn starch on TP medium, and maintained at ambient CO2 in light. Two cosmid pools produced complemented cia5 mutants, and two single cosmids from each pool were pinpointed by methods previously described (8, 9). Fragments of one of the cosmids, c118C2, were subcloned into pGEM-5Zf(+). A complementing 4.2-kb NdeI–NotI fragment was identified and sequenced. Subsequently, an additional 5 kb of DNA downstream of this fragment and containing portions of the Cia5 gene was cloned and sequenced.
DNA and RNA Hybridization.
DNA and RNA blot hybridizations were performed with the Nucleic Acid Hybridization and Detection kit (Roche, Indianapolis). For RNA analyses, total RNA was isolated from cells grown in TP medium in high CO2 and from cells shifted to low CO2 for 0, 45, 90, and 180 min. Ten micrograms of total RNA was resolved by electrophoresis on a formaldehyde-agarose gel, blotted onto nylon membrane, and hybridized with digoxinin-labeled DNA amplified as recommended by the kit manufacturer (Roche) from the Cia5 gene or CO2-response genes.
Cloning of the Cia5 cDNA.
The complete DNA sequence of the Cia5 gene was determined in the University of Nebraska-Lincoln DNA Core Facility and was analyzed via genemark computer software (http://genemark.biology.gatech.edu/GeneMark/hmmchoice.html) to predict exons and introns. Primers were designed to amplify different predicted exons via reverse transcriptase (RT)–PCR (11). Total RNA from cells grown in low CO2 conditions was reverse-transcribed in 20 μl of volume with a specific primer, 1-forward (5′-GAACAGCAGCTGCATGAGG-3′), at 50°C for 1 h using Superscript II RNase H− RT (GIBCO/BRL). For cloning the cDNA sequences encoded within the NdeI–NotI fragment, a pair of primers, 1-forward and p1757 (5′-GATGGGGACCCGCACAAG-3′), were used to generate a 1.2-kb product. To determine the transcriptional initiation site, 5′ rapid amplification of cDNA ends (RACE) was performed by using a 5′RACE kit (GIBCO/BRL) with a specific primer, p1316 (5′-CGGCTGCTGGTACCGCTG-3′) that produced a 771-bp product that overlapped with the 5′ portion of the 1.2-kb cDNA fragment. The transcriptional initiation site was further confirmed by using RNA ligase-mediated RACE with a RLM-RACE kit from Ambion. After exon regions in the 4.2-kb NdeI–NotI fragment of the Cia5 gene were confirmed, potential exons in 5 kb of DNA downstream of the predicted TAG termination codon were sought. RT-PCR was performed with primers p5143 (5′-CTAATCGCAGGACTGCAG-3′) and p3770 (5′-ACGGTGACGTAATGTACGACC-3′) to generate a 584-bp cDNA product that overlapped at its 5′ end with the 1.2-kb cDNA fragment mentioned above. Because Northern hybridization results showed the Cia5 mRNA was ≈5.1 kb in size, further cloning and sequencing of downstream cDNA sequences were conducted. Oligo(dT)30 was used as a 3′ primer for production of a RT product. This RT product, in turn, was used for PCR amplification of cDNA fragments using specific primers spaced at ≈400-bp intervals. The 3′ end of the Cia5 cDNA was cloned and identified by using a 3′ RACE kit (GIBCO/BRL).
Photosynthetic Response Curves.
Oxygen evolution in response to increasing levels of KHCO3 by air-adapted, midlog cells was measured with a Clarke oxygen electrode (Hansatech, Norfolk, U.K.) at pH 7.0 according to Van and Spalding (12).
Construction of CIA5 with a 3× Hemagglutinin (HA) Epitope.
A 120-bp 3× HA sequence was amplified by PCR from pDW23 kindly provided by Arthur Grossman, Carnegie Institution of Washington. The amplified sequence was inserted at the 3′ terminus of the Cia5 gene to produce the fused CIA5∷3×HA gene. DNA sequencing was used to confirm that the genes were fused in the correct reading frame.
Western Blot Analysis of CIA5 levels.
Cells (≈5 × 106 wild-type cells, cia5 mutants, or cia5 mutants complemented with the CIA5∷3×HA gene) in exponential growth phase under high or low CO2 conditions were harvested by centrifugation and resuspended in 30 μl of SDS lysis buffer (11). After electrophoresis through a 10% SDS polyacrylamide gel, proteins were blotted onto a nitrocellulose membrane by using standard methods (11) and probed with rabbit polyclonal antibodies to HA (Roche) as the primary antibody and anti-rabbit Ig linked to horseradish peroxidase as the secondary antibody (Amersham Pharmacia).
Nuclear Localization of the CIA5 Protein.
A 1,592-bp fragment of the coding region of the Cia5 cDNA from the initiator ATG to a downstream NcoI was inserted into the NcoI site at the initiator codon downstream of the cauliflower mosaic virus (CaMV) 35S promoter in the plant cell expression vector pPTN134 (kindly provided by Tom Clemente, University of Nebraska). The resultant plasmid, designated as pCia5-glucuronidase (GUS), was sequenced to confirm junction sites and the ORF. Targeting of CIA5 protein to cell nuclei was conducted by introducing the chimeric Cia5-gus gene into onion epidermal cells by using methods described by Scott et al. (13). The control plasmid, pPTN134, and plasmids containing the fused gene construct were coated on tungsten particles and delivered to freshly prepared onion epidermal cells by particle bombardment. The bombarded cells then were cultured on hormone-free Murashige and Skoog (MS) medium without vitamins for 20 h under moderate light at 25°C. The cells then were stained in 5-bromo-4-chloro-3-indoly-β-d-glucuronide solution at 37°C for 7 h. The cells were affixed to slides, stained with propidium iodide, and observed with a fluorescent microscope.
Results
Genomic Complementation of the cia5 Mutant.
To isolate the gene involved in the defective phenotype of the high
CO2-requiring mutant
cia5, mutant cells were transformed with
pools of DNA from an indexed cosmid library of C.
reinhardtii DNA (8, 9) using a newly developed electroporation
procedure for Chlamydomonas transformation (10). Pools of
cosmid DNAs obtained from bacteria in two of the 120 microtiter dishes
comprising the library proved capable of complementing the
cia5 mutation when transformed cells were plated on minimal
medium in light under ambient (0.03%) CO2
conditions. Cosmid DNAs from bacteria in each row and each file of one
microtiter dish (no. 118) were used to transform the cia5
mutant and pinpoint the well (118C2) containing the DNA clone
responsible for complementation. The cosmid, c118C2, from this well
transformed the cia5 mutant at high rates (1.8 ×
10−4 transformants/cell) and produced
complemented cells with photosynthetic growth rates comparable to those
of wild-type cells (data not shown). The complemented cells also
regained the ability to concentrate Ci. An active
CCM was demonstrated in duplicate experiments using oxygen electrode
techniques (12) to measure the photosynthetic response
(O2 evolution) to increasing concentrations of
Ci (supplied as HCO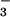 ) in
air-adapted cells at pH 7.0. Measurements of
K1/2 (Ci) for
cells grown in ambient concentrations of CO2 were
17.3 ± 7.3 μM for wild-type (CC125) cells; 98.2 ± 21.7
μM for cia5 ; 10.3 ± 3.0 μM for cia5
complemented with an intact Cia5 gene; and 12.9 ± 1.2
μM for cia5 complemented with a truncated Cia5
gene. The control K1/2
(Ci) value for wild-type cells grown in high
CO2 was 164 ± 49 μM in this experiment.
) in
air-adapted cells at pH 7.0. Measurements of
K1/2 (Ci) for
cells grown in ambient concentrations of CO2 were
17.3 ± 7.3 μM for wild-type (CC125) cells; 98.2 ± 21.7
μM for cia5 ; 10.3 ± 3.0 μM for cia5
complemented with an intact Cia5 gene; and 12.9 ± 1.2
μM for cia5 complemented with a truncated Cia5
gene. The control K1/2
(Ci) value for wild-type cells grown in high
CO2 was 164 ± 49 μM in this experiment.
Cia5 Gene and mRNA.
Comparison of DNA sequences of cDNAs produced from a Cia5 mRNA with the DNA sequences of the Cia5 gene revealed that the gene is ≈6,481 bp in length and contains five introns and six exons, including an exceptionally long 2,904-bp 3′ exon (Fig. 1A). Northern blot analyses of mRNAs from complemented cia5 mutants and wild-type cells revealed that the Cia5 gene produces a mRNA of ≈5.1 kb in size (Fig. 1B, lanes 1–16). Transcription of the mutant Cia5 gene (Fig. 1B, lanes 2, 6, 10 and 14) produced a mRNA of the same size and abundance as the Cia5 mRNA produced by wild-type cells (Fig. 1B, lanes 1, 5, 9 and 13). There was no significant change in levels of Cia5 mRNA after a shift from high to low CO2 concentrations (Fig. 1B, lanes 1–4 vs. lanes 5–16). Constant levels of Cia5 mRNA under both high and low CO2 conditions suggested that expression of the CIA5 protein, likewise, might be constitutive.
Figure 1.

Cia5 gene, mRNA, and CIA5 protein. (A) Structure of the Cia5 gene and mRNA. The schematic diagram at the top shows the intron/exon configuration of the Cia5 gene along with a portion of the promoter region (from the SalI restriction site at −163 bp to the transcription initiation site at +1). The lower diagram illustrates the 2,094-nt coding region of the Cia5 mRNA along with its exceptionally long, 2,904-nt 3′ UTR. (B) Northern blot analysis of Cia5 mRNAs from wild-type cells (lanes 1, 5, 9, and 13), cia5 mutant cells (lanes 2, 6, 10, and 14), cia5 mutants complemented with an intact Cia5 gene (lanes 3, 7, 11, and 15), and cia5 mutants complemented with a slightly truncated Cia5 gene (lanes 4, 8, 12, and 16). Total RNA was extracted from cells maintained in high concentrations of CO2 (lanes 1–4) or from cells after they were switched from high CO2 levels to low levels of CO2 for 45 min (lanes 5–8), 90 min (lanes 9–12), or 180 min (lanes 13–16). The last lane of the Northern blot (M) contains labeled RNA markers of 1.5, 2.6, and 4.7 kb in size. (C) Deduced amino acid sequence of the CIA5 protein. The CIA5 protein contains a number of notable features including an abundance of glutamine residues (red) and repeats (residues 316–329), alanine residues (blue) and repeats (residues 352–363), and a region interspersed with glycines and alanines (italicized residues 550–627). The cysteine and histine residues contained in two potential, but noncanonical, zinc-finger motifs are underlined (residues C35 and C41, H54 and H69; C77 and C79, H89 or H90 and C93). The cia5 mutation is the result of a single nucleotide base pair change that converts the H54 histidine residue (noted above as a component of a potential zinc-finger motif) to a tyrosine residue. The underlined amino acid sequences at the C-terminal end of the protein represents a potential protein kinase C phosphorylation site. The italicized glutamine residue at position 642 represents the last complete codon encoded by the Cia5 gene truncated at its single NotI restriction enzyme cut site.
CIA5 Levels in High and Low CO2.
To determine whether the level of CIA5 was constant or changed with shifts in CO2 concentrations, the cia5 mutant was transformed with a Cia5 gene encoding an epitope-tagged CIA5 (i.e., a CIA5/HA fusion protein). In duplicate experiments using three separate transformants, it was observed that the level of CIA5 protein of the expected size was identical or nearly identical under low CO2 conditions (Fig. 2, lanes 4, 6, and 8) and high CO2 conditions (Fig. 2, lanes 5, 7, and 9). Specificity of the antibodies used was demonstrated by the lack of immunoreactive proteins in nontransformed cia5 mutants (Fig. 2, lane 1) or wild-type cells (Fig. 2, lanes 2 and 3).
Figure 2.

Cellular levels of CIA5 under low and high CO2 conditions. Western blot analysis was performed with total cell lysates of three separate cia5 mutants (lanes 4 and 5; lanes 6 and 7; lanes 8 and 9, respectively) complemented with an intact, HA-tagged Cia5 gene and maintained under low CO2 conditions (lanes 4, 6, and 8) or high CO2 conditions (lanes 5, 7, and 9). Epitope-tagged CIA5 was detected with antibodies to HA. Specificity of the antibody for epitope-tagged CIA5 was demonstrated by the lack of detection of proteins of the expected size in extracts from cia5 mutants (lane 1) or from wild-type cells maintained in high (lane 2) or low (lane 3) CO2 conditions.
Features of the CIA5 Protein.
The CIA5 protein (Fig. 1C) contains a number of striking features. First, CIA5 is a protein of unusual amino acid composition and configuration. It is unusually rich in alanines (16.2%), glycines (16.7%), prolines (10%), and glutamines (7.3%). The amino acid sequence of CIA5 reveals that the protein contains a long segment of repeated glutamine residues, a similar stretch of alanine residues, and a long stretch of glycine residues interspersed with alanine residues. Moreover, it contains an N-terminal region with an abundance (27.5%) of charged amino acids (23 lysine and arginine residues and 16 aspartate and glutamate residues in the first 142 aa) and a C-terminal region that is predominantly acidic. Second, the region from amino acid position 46 to 93 contains two potential, but noncanonical, zinc finger domains (CxxCxxxxxxxxxHxxxxxxxxH and CxxxCxxxxxxxxxHHxxC) (Fig. 1C). Third, there are numerous sites for potential posttranslational modifications of CIA5. These include a site for phosphorylation of the C terminus by protein kinase C, a modification whose potential relevance is discussed below. Finally, comparison of DNA sequences from the mutant Cia5 gene and those from the wild-type gene reveals a single nucleotide mutation (C > T) at nucleotide position 431. This mutation results in the substitution of a tyrosine residue for one of the two histidine residues that comprise the first zinc finger motif noted above (Fig. 1C).
Nuclear Localization of CIA5.
Computer-assisted analysis of the CIA5 aa sequence (protcomp, version 4, http://www.softberry.com) predicted a nuclear localization of the protein. However, because of the apparently low concentrations of the CIA5 in the C. reinhardtii cell, immunodetection of the protein in situ was not successful. As an alternative approach, a chimeric construct containing the Cia5 gene fused in-frame with the β-GUS gene and driven by the CaMV 35S promoter was used to transform onion bulb epidermal cells (13) by particle bombardment. Fig. 3A illustrates the apparently exclusive nuclear localization of the GUS activity of the CIA5/GUS fusion protein. Of 63 onion cells transformed with the chimeric CIA5/GUS gene and displaying GUS staining, all exhibited staining only of the nucleus. Transformation of onion epidermal cells with the analogous CaMV 35S/GUS gene construct lacking the Cia5 coding sequence showed no evidence for nuclear localization of GUS activity (Fig. 3C). That is, in 287 epidermal cells showing GUS staining after transformation with the CaMV 35S/GUS construct, the staining was uniformly distributed throughout the cytoplasm of each cell.
Figure 3.

Nuclear localization of the CIA5 protein. Onion epidermal cells were transformed by particle bombardment with a chimeric gene construct containing a truncated Cia5 gene fused in-frame with a β-GUS gene and driven by the promoter from the CaMV 35S promoter. Cells were fixed and prepared for detection of the GUS enzyme activity by reaction with 5-bromo-4-chloro-3-indoly-β-d-glucuronide. (A) Epidermal cells bombarded with the Cia5/GUS chimeric gene. (B) Cells in A stained with propidium iodide for DNA detection. (C) Epidermal cells bombarded with a CaMV 35S/GUS gene construct. (D) Cells in C stained with propidium iodide for DNA detection. (Magnification: ×100).
Regulation of CO2-Responsive Genes.
The kinetics of appearance of mRNAs encoded by different CO2-responsive genes after transfer of cells from high CO2 to low CO2 were determined (Fig. 4). The results revealed that cia5 mutants complemented with an intact copy of the Cia5 gene produced essentially identical increases in mRNAs to those observed in wild-type cells (Fig. 4, lanes 3, 7, 11 and 15 vs. lanes 1, 5, 9 and 13, respectively). As expected from the earlier results of others (4, 5, 14), no transcripts from any of the CO2-responsive genes were detectable in RNA extracted from the cia5 mutant grown under either high or low CO2 conditions (Fig. 4, lanes 2, 6, 10, and 14).
Figure 4.

Northern blot analysis of the induction or derepression of synthesis of mRNAs encoded by four different CO2-responsive genes on shift of cells from high concentrations of CO2 to low, ambient concentrations of CO2. Total RNA was extracted from wild-type cells (lanes 1, 5, 9, and 13), cia5 mutant cells (lanes 2, 6, 10, and 14), cia5 mutants complemented with an intact Cia5 gene construct (lanes 3, 7, 11, and 15), and cia5 mutants complemented with a truncated Cia5 gene (lanes 4, 8, 12, and 16). Total RNA was extracted from cells maintained in high concentrations of CO2 (lanes 1–4) or from cells after they were switched from high CO2 levels to low levels of CO2 for 45 min (lanes 5–8), 90 min (lanes 9–12), or 180 min (lanes 13–16). (A) mRNAs detected by hybridization with probes produced from the cDNA encoding Ccp2 (LIP36), a chloroplast inner envelope protein. (B) mRNAs detected with hybridization probes to the CAH1 cDNA encoding the major C. reinhardtii periplasmic CA. (C) mRNAs detected with probes to Att1 cDNA sequences encoding an alanine:α-ketoglutarate aminotransferase. (D) mRNAs detected with probes produced from the mtCA1 cDNA encoding a mitochondrial CA. (E) Ethidium bromide-stained gel illustrating uniform loading of total RNAs in each lane. The same RNA preparations used for the Northern blot analyses depicted in Fig. 1B were used for the blots shown.
Altered Regulation of CO2-Responsive Genes in Cells Transformed with a Truncated Cia5 Gene.
In subcloning segments of the c118C2 cosmid to recover a minimally sized Cia5 gene, we discovered that complementation of the cia5 mutant could be achieved at high rates (>1.9 × 10−4 transformants/cell) with a 4.2-kb NdeI/NotI fragment. This result was surprising in light of the size of the Cia5 mRNA (i.e., 5.1 kb) detected on Northern blots (Fig. 1B). Subsequent sequencing of the Cia5 gene and cDNAs revealed that the 4.2-kbp fragment was lacking 161 nt of the 3′ coding region of the gene. The appearance of a 2.1-kb mRNA hybridizing to a Cia5 gene probe only in cia5 mutants complemented with the truncated, 4.2-kbp Cia5 gene (Fig. 1B, lanes 4, 8, 12, and 16) strongly suggested that the 2.1-kb mRNA was transcribed from the truncated Cia5 gene. Importantly, when RNAs were extracted from cia5 mutants complemented with the 4.2-kbp fragment, it was discovered that all four CO2-responsive genes examined were constitutively expressed in complemented mutants grown in light under both high and low CO2 conditions (Fig. 4, lanes 4, 8, 12, and 16). This finding is in marked contrast to the situation with mRNAs isolated from wild-type cells or cia5 mutants complemented with an intact Cia5 gene cultured in high CO2. In this case, transcripts from same CO2-responsive genes are not detectable (Fig. 4, lanes 1 and 3, respectively).
Discussion
Genomic complementation of the cia5 mutant with an indexed cosmid library of C. reinhardtii DNA (8, 9) was used to isolate a cosmid, c118C2, containing a gene capable of restoring a wild-type phenotype to transformed cia5 mutants. Complemented mutants were able to grow photoautotrophically and concentrate Ci at ambient levels of CO2 at rates similar to wild-type cells. Complemented cells also regained the ability to activate transcription from CO2-responsive genes when shifted from high to low CO2 concentrations (Fig. 4).
The Cia5 Gene and mRNA.
Sequencing of the Cia5 cDNA showed one primary ORF encoding a protein of 698 aa. Based on comparison of the Cia5 cDNA and gene DNA sequences, the gene appears to contain six exons and five introns (Fig. 1A). Perhaps the most striking feature of the Cia5 mRNA is the ≈2,900-nt-long 3′ untranslated region (UTR). This is, by far, the longest 3′ UTR reported for C. reinhardtii mRNAs (GenBank). Such long 3′ UTRs have been reported in a few eukaryotic systems (16), but the functional significance of most long UTRs has not been established. Interestingly, the 3′ UTR of the Cia5 gene lacks introns. This 2.9-kb intron-less region is exceptionally long for C. reinhardtii genes, which average ≈3.9 introns per 1,000 bp (15).
Cia5 mRNA was found to be present constitutively in light in both wild-type cells and the cia5 mutant under both high and low CO2 conditions (Fig. 1B). This finding suggested that the CIA5 protein, likewise, might be constitutively expressed. Western blot analysis of epitope-tagged CIA5 confirmed the constitutive presence of CIA5 in high and low CO2 in light (Fig. 2). As discussed below, the constitutive presence of CIA5 suggested that some type of posttranslational modification of CIA5 could be required to allow (or repress) its role in controlling the synthesis and/or function of components of the CCM in C. reinhardtii.
Complementation with a Subgenic Fragment of the Cia5 Gene.
During the course of subcloning the Cia5 gene from the parent cosmid, c118C2, it was discovered that a small 4.2-kb NdeI/NotI fragment of the cosmid could complement the cia5 mutant with high efficiency. Complementation was achieved using either a gel-purified 4.2-kb fragment or the fragment cloned into a plasmid vector. Two observations initially suggested that this fragment might contain only a portion of the Cia5 gene. First, sequencing of the Cia5 cDNA showed that the presumed termination codon in the coding region of the gene was located ≈161 bp downstream of the NotI cut site (Fig. 1A). Second, Northern blot analysis of Cia5 mRNA from wild-type cells and the cia5 mutant revealed a single major mRNA with a size of ≈5.1 kb (Fig. 1B). Finally, it was shown in Northern blot analysis of mRNAs from cia5 mutants complemented with the 4.2-kb fragment of the Cia5 gene that both a 5.1-kb and a 2.1-kb Cia5 mRNA were present in these cells (Fig. 1B, lanes 4, 8, 12, and 16). It was surmised that the upper 5.1-kb band corresponded to an mRNA transcribed from the endogenous mutant Cia5 gene and that the second smaller mRNA was transcribed from the truncated 4.2-kb Cia5 gene fragment integrated into the host genome.
Regulation of CO2-Responsive Genes by the Cia5 Gene Product.
A shift of wild-type C. reinhardtii cells (and cia5 mutants complemented with an intact Cia5 gene) from high CO2 levels to low CO2 levels results in a rapid activation of transcription of mRNAs from several CO2-responsive genes (Fig. 4) and the assembly of a fully functional CCM (4, 5). However, in cia5 mutants transformed with the truncated Cia5 gene contained in a 4.2-kb NdeI/NotI DNA fragment, CO2-responsive genes are expressed under both high and low CO2 conditions (Fig. 4, lanes 4, 8, 12, and 16). This finding strongly suggests that the 54 aa missing from the C terminus of the CIA5 protein in these cells are essential to the normal function of CIA5 in controlling transcription of CO2-responsive genes.
A controversy has existed from soon after the time that the cia5 mutant was first isolated and characterized (6) as to whether this potential regulator of CCM formation and function served to control induction of CO2-responsive genes or if it was responsible for strong repression of these genes under conditions of high CO2. Although more complex mechanisms can be envisioned, the simplest hypothesis that is consistent with present data is one in which CIA5 is an inducer of gene transcription in cells grown in low CO2 environments, but is inactivated by posttranslational modification in cells switched to high levels of CO2. The logic for this hypothesis is as follows. Our results demonstrate that the presence of a functional CIA5 is essential for induction of transcription of a specific set of genes. Also, we have observed that CIA5 is present in light at essentially constant levels regardless of changes in CO2 concentrations (Fig. 2). This observation strongly suggests that neither controls at the transcriptional level nor at the translational level lead to increases or decreases in CIA5 levels and, therefore, do not serve to regulate CIA5 activity. Other experiments have demonstrated that CIA5 truncated at its C terminus retains its ability to complement the cia5 mutant and to act as an inducer of transcription at low CO2 levels. However, unlike wild-type CIA5, the truncated CIA5 molecule allows continued transcription of CO2-responsive genes in complemented cia5 mutants grown in high CO2. These collective observations and arguments provide a strong base for the hypothesis that wild-type CIA5 is an inducer of gene transcription at low CO2 levels and that its activity is negated at high CO2 levels by modification of one or more sites in the C-terminal domain, a region that is missing in the truncated CIA5 molecule.
If the preceding hypothesis is true, CIA5 could be converted reversibly from active to inactive states via cycles of phosphorylation/dephosphorylation, acetylation/deacetylation, or any other of the myriad of posttranslational modifications known to control the activities of transcription factors (17). In this regard, the presence of a potential protein kinase C phosphorylation site at the extreme C terminus of CIA5 (Fig. 1C) is intriguing.
Whether the “activated” inducer form of the CIA5 protein acts first on a single gene whose product, in turn, activates downstream CO2-responsive genes or if it directly leads to the increased transcription of Cah1, Mca 1, Mca2, Ccp1, Ccp2, and other CO2-responsive genes is an open question. Because genes in C. reinhardtii, such as the tubulin genes, can be induced and expressed at elevated levels in less than 15 min (e.g., ref. 18), the 15 min (e.g., ref. 19) to 1 h needed for activation of many of the CO2-responsive genes potentially leaves time for a short gene activation cascade.
An Argument Against CIA5 as a Repressor of Transcription.
The fact that the cia5 mutant does not produce a functional CIA5 molecule—as evidenced by the ability of a gene encoding a wild-type CIA5 to complement this mutant—strongly argues against CIA5 being a repressor. That is, if CIA5 was a repressor, then mutant cia5 cells should exhibit constitutive expression of the CO2-responsive genes because they would not produce a functional repressor. The data of Fig. 4 showing the absence of mRNAs from CO2-responsive genes in cia5 mutants grown under either high or low CO2 conditions clearly demonstrate that this is not the case.
Is CIA5 a Transcription Factor?
The mechanism by which CIA5 controls expression of CO2-responsive genes and the development of the CCM in C. reinhardtii grown under low CO2 conditions is unknown. Nonetheless, several observations point to a potential role of CIA5 as a transcription factor. Clearly, the Cia5 gene is responsible for controlling the expression of CO2-responsive genes. The amino acid sequences of domains within the CIA5 protein show marked similarity to several known and putative transcription factors. The glutamine-rich region of CIA5 is similar to that found in a number of transcription factors (17) including the PSR1 protein, a putative transcription factor involved in the regulation of phosphate uptake and utilization in C. reinhardtii (20).
Two motifs within the CIA5 protein have clear similarities to zinc finger motifs found in numerous transcription factors (21, 22) (Fig. 1C). However, both potential zinc finger regions in CIA5 differ from the standard CX2–4CX9–12HX2–4H configuration. The first potential zinc finger conforms to the canonical zinc finger motif except for an extra long loop between the two histidine residues (i.e., 15 aa vs. the usual 2–4 aa). The second motif contains a cysteine in place of the terminal histidine residue. Such a substitution has been observed in a number of zinc fingers in various organisms (especially in the last zinc finger in a tandem array of zinc fingers) and has been demonstrated to allow formation of a functional zinc finger domain (23).
The lack of conformity to the canonical zinc finger motif might encourage dismissal of the potential role of these unusual zinc-finger motifs in CIA5 function. However, it is important to note that there is only one mutation in the Cia5 gene of the cia5 mutant that results in a change in the amino acid sequence of the CIA5 protein. This mutation causes a change of the first histidine residue (residue H54) in the upstream zinc-finger motif to a tyrosine. Whether the zinc finger domains of CIA5 are involved in binding specific DNA sequences in target genes or to associated transcription factors is an open question that will require further investigation. Finally, computer program predictions (e.g., protcomp version 4, http://www.softberry.com) for a nuclear localization of CIA5 and the clear-cut nuclear localization of CIA5 in onion epidermal cells (Fig. 3) provide additional weight to the argument that CIA5 may be a transcription factor. These observations, coupled with observations of constitutive expression of CO2-responsive genes in cia5 mutants transformed with the 4.2-kb truncated Cia5 gene, suggest that CIA5 may act directly at the gene level to control transcription of specific genes under varying CO2 conditions.
Perspectives.
The mechanism by which C. reinhardtii and other
photosynthetic organisms sense changes in Ci
concentrations is unknown. The availability of the Cia5 gene
should expedite progress in unraveling the signal transduction pathway
triggered by changes in CO2 concentrations. We
propose that such a pathway leads from an initial change in a
Ci sensor to the putative posttranslational
modification that activates or inactivates the CIA5 protein and finally
to the cascade of gene inductions that are necessary for the synthesis
and/or function of components of the CCM under low
CO2 conditions. Thus, from present observations,
it can be hypothesized that posttranscriptional modification of CIA5
permits an immediate response of cells to fluctuations in
CO2 and HCO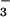 concentrations, a
common occurrence in the natural environment in which algae must
survive. The increasing pace with which genes and proteins associated
with the structure and function of the CCM can be isolated and studied
(4, 5, 9) provides promise of a much fuller understanding of the CCM in
the near future. From a longer term and more practical perspective, a
detailed knowledge of the CCM in C. reinhardtii and other
algae, ultimately, may allow for modifications to higher plants that
will permit more efficient uptake and utilization of the often
growth-limiting concentrations of CO2 in the
atmosphere (24–26).
concentrations, a
common occurrence in the natural environment in which algae must
survive. The increasing pace with which genes and proteins associated
with the structure and function of the CCM can be isolated and studied
(4, 5, 9) provides promise of a much fuller understanding of the CCM in
the near future. From a longer term and more practical perspective, a
detailed knowledge of the CCM in C. reinhardtii and other
algae, ultimately, may allow for modifications to higher plants that
will permit more efficient uptake and utilization of the often
growth-limiting concentrations of CO2 in the
atmosphere (24–26).
Acknowledgments
We thank Dr. James Moroney for providing the wall-less C. reinhardtii cia5 mutant used in the present studies, the laboratory of Dr. Heriberto Cerutti for assistance in nuclear localization studies, Dr. Ed van Hunnik for calculations dealing with oxygen electrode analyses, and the laboratory of Dr. Martin Spalding for providing clones of CO2-responsive genes and assisting in experiments using oxygen electrodes. We thank Dr. Spalding and Dr. Roel Funke for their careful review of the manuscript. This research is supported by a grant from the National Science Foundation (IBN-9723333) and funding from the University of Nebraska-Lincoln Center for Biotechnology. This manuscript is Journal Series no. 13,174 of the Agricultural Research Division, University of Nebraska.
Abbreviations
- CCM
carbon concentrating mechanism
- TAP
Tris acetate phosphate medium
- RACE
rapid amplification of cDNA ends
- RT
reverse transcriptase
- HA
hemagglutinin
- CaMV
cauliflower mosaic virus
- GUS
glucuronidase
- UTR
untranslated region
- CA
carbonic anhydrase
- Ci
inorganic carbon
Footnotes
References
- 1.Raven J A. Can J Bot. 1991;69:908–924. [Google Scholar]
- 2.Badger M R. In: The Biochemistry of Plants: A Comprehensive Treatise. Hatch M D, Boardman N K, editors. Vol. 10. New York: Academic; 1987. pp. 219–274. [Google Scholar]
- 3.Kaplan A, Reinhold L. Annu Rev Plant Physiol Plant Mol Biol. 1999;50:539–570. doi: 10.1146/annurev.arplant.50.1.539. [DOI] [PubMed] [Google Scholar]
- 4.Spalding M H. In: The Molecular Biology of Chloroplasts and Mitochondria of Chlamydomonas. Rochaix J D, Goldschmidt-Clermont M, Merchant S, editors. Dordrecht, The Netherlands: Kluwer; 1998. pp. 529–547. [Google Scholar]
- 5.Moroney J V, Somanchi A. Plant Physiol. 1999;119:9–16. doi: 10.1104/pp.119.1.9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Moroney J V, Husic H D, Tolbert N E, Kitayama M, Manuel L J, Togasaki R K. Plant Physiol. 1989;89:897–903. doi: 10.1104/pp.89.3.897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Gorman D S, Levine R P. Proc Natl Acad Sci USA. 1965;54:1665–1669. doi: 10.1073/pnas.54.6.1665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zhang H, Herman P L, Weeks D P. Plant Mol Biol. 1994;24:663–672. doi: 10.1007/BF00023562. [DOI] [PubMed] [Google Scholar]
- 9.Funke R P, Kovar J L, Weeks D P. Plant Physiol. 1997;114:237–244. doi: 10.1104/pp.114.1.237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Shimogawara K, Fujiwara S, Grossman A, Usuda H. Genetics. 1998;148:1821–1828. doi: 10.1093/genetics/148.4.1821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ausubel F M, Brent R, Kingston R E, Moore D D, Seidman J G, Smith J A, Struhl K. Current Protocols in Molecular Biology. New York: Wiley; 1995. [Google Scholar]
- 12.Van K, Spalding M. Plant Physiol. 1999;120:757–764. doi: 10.1104/pp.120.3.757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Scott A, Wyatt S, Tsou P-L, Robertson D, Stromgren-Allen N. BioTechniques. 1999;26:1125–1132. doi: 10.2144/99266st04. [DOI] [PubMed] [Google Scholar]
- 14.Somanchi A, Moroney J V. Plant Mol Biol. 1999;40:1055–1062. doi: 10.1023/a:1006262123918. [DOI] [PubMed] [Google Scholar]
- 15.Silflow C D. In: The Molecular Biology of Chloroplasts and Mitochondria of Chlamydomonas. Rochaix J D, Goldschmidt-Clermont M, Merchant S, editors. Dordrecht, The Netherlands: Kluwer; 1998. pp. 25–40. [Google Scholar]
- 16.Serikawa K A, Mandoli D. Plant Mol Biol. 1999;41:785–793. doi: 10.1023/a:1006387107071. [DOI] [PubMed] [Google Scholar]
- 17.Carey M, Smalle S T. Transcriptional Regulation in Eukaryotes. Plainview, NY: Cold Spring Harbor Lab. Press; 2000. [Google Scholar]
- 18.Brunke K, Anthony J, Kalish F, Sternberg E, Weeks D. In: Molecular Biology of the Cytoskeleton. Borisy G G, Cleveland D W, Murphy D B, editors. Plainview, NY: Cold Spring Harbor Lab. Press; 1984. pp. 367–380. [Google Scholar]
- 19.Eriksson M, Villand P, Gardestrom P, Samuelsson G. Plant Physiol. 1998;116:637–641. doi: 10.1104/pp.116.2.637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wykoff D D, Grossman A R, Weeks D P, Usada H, Shimogawara K. Proc Natl Acad Sci USA. 1999;96:15336–15341. doi: 10.1073/pnas.96.26.15336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Gogos J A, Hsu T, Bolton J, Kafatos F C. Science. 1992;257:1951–1955. doi: 10.1126/science.1290524. [DOI] [PubMed] [Google Scholar]
- 22.Bouhouche N, Syvanen M, Kado C I. Trends Microbiol. 2000;8:77–81. doi: 10.1016/s0966-842x(99)01679-0. [DOI] [PubMed] [Google Scholar]
- 23.Krizek B A, Amann B T, Kilfoil V J, Merkle D L, Berg J M. J Am Chem Soc. 1991;113:4518–4523. [Google Scholar]
- 24.Stitt M. Plant Cell Environ. 1991;14:741–762. [Google Scholar]
- 25.Bowes G. Annu Rev Plant Physiol Plant Mol Biol. 1993;44:309–332. [Google Scholar]
- 26.Culotta E. Science. 1995;268:654–656. doi: 10.1126/science.268.5211.654. [DOI] [PubMed] [Google Scholar]