

Abstract
Aquatic photosynthetic organisms, including the green alga Chlamydomonas reinhardtii, induce a set of genes for a carbon-concentrating mechanism (CCM) to acclimate to CO2-limiting conditions. This acclimation is modulated by some mechanisms in the cell to sense CO2 availability. Previously, a high-CO2-requiring mutant C16 defective in an induction of the CCM was isolated from C. reinhardtii by gene tagging. By using this pleiotropic mutant, we isolated a nuclear regulatory gene, Ccm1, encoding a 699-aa hydrophilic protein with a putative zinc-finger motif in its N-terminal region and a Gln repeat characteristic of transcriptional activators. Introduction of Ccm1 into this mutant restored an active carbon transport through the CCM, development of a pyrenoid structure in the chloroplast, and induction of a set of CCM-related genes. That a 5,128-base Ccm1 transcript and also the translation product of 76 kDa were detected in both high- and low-CO2 conditions suggests that CCM1 might be modified posttranslationally. These data indicate that Ccm1 is essential to control the induction of CCM by sensing CO2 availability in Chlamydomonas cells. In addition, complementation assay and identification of the mutation site of another pleiotropic mutant, cia5, revealed that His-54 within the putative zinc-finger motif of the CCM1 is crucial to its regulatory function.
Keywords: zinc-finger motif, carbon transport, signal transduction, photosynthesis, acclimation
Photosynthetic organisms sense environmental changes, e.g., light, temperature, and various nutrient availabilities, to modulate and optimize photosynthetic activities. A number of aquatic photosynthetic organisms are able to concentrate dissolved inorganic carbon (DIC) intracellularly, allowing rapid growth despite low-CO2 availability externally (1). This carbon-concentrating mechanism (CCM) shows acclimation to external DIC to optimize CO2 fixation efficiency (2). During acclimation these organisms induce the expression of a set of genes required for various aspects of the CCM. In Chlamydomonas reinhardtii, several genes have been shown to be regulated in response to changes in external CO2 concentration, including periplasmic carbonic anhydrase (CA; ref. 3), mitochondrial CA (4), and a chloroplast envelope protein, LIP-36 (5). A development of pyrenoid structure in the chloroplasts also is modulated by the supply of CO2 (6). This acclimation to CO2-limiting conditions suggests the existence of sensory mechanisms by which cells perceive the shortage of CO2 and pathways by which the signal is transduced into specific gene regulation. However, the regulation of gene expression during this acclimation in eukaryotic organisms is still poorly understood. One reason is the paucity of mutants impaired in induction processes. We have isolated previously one C. reinhardtii mutant (C16) by gene-tagging mutagenesis, which was high-CO2 requiring (7), and we showed that C16 exhibited a defect in CCM induction, with a low affinity for DIC and a low level of DIC accumulation under CO2-limiting conditions. It has been assumed that C16 had a mutation in a gene that serves as a regulatory switch to induce a set of genes for acclimation to low-CO2 conditions, and that this gene may play an important role for a CO2 signal-transduction pathway.
In the present study, we have cloned a nuclear regulatory gene, Ccm1, which complements the mutation in C16. In addition, we show that a similar induction mutant, cia5 (8), also is complemented by the same genomic DNA fragment encoding Ccm1. On the basis of the structure and expression of the Ccm1 gene, we discuss the function and importance of the CCM1 in the CO2 signal-transduction pathways in the eukaryotic photosynthetic organism C. reinhardtii.
Materials and Methods
Strains.
C. reinhardtii strain 5D (nit1–305, cw15), which is the wild-type (WT) cell for photosynthesis, was kindly provided by P. A. Lefebvre (Univ. of Minnesota, St. Paul). High-CO2-requiring mutant C16 was described (7) and another high-CO2-requiring mutant, cia5, was kindly provided by J. V. Moroney (Louisiana State Univ., Baton Rouge, LA) (8). Cells were cultured in tris-acetate-phosphate medium (9) for maintenance or in a modified high-salt (HS) medium supplemented with 20 mM Mops (pH 7.2) for photosynthetic growth under aeration with ordinary air containing 0.04% CO2 (low-CO2) or air enriched with 5% CO2 (high-CO2).
Transformation of Chlamydomonas Cells.
C. reinhardtii was transformed with 20 μg/ml plasmid DNA by electroporation (10) with the slight modification that cells were embedded in 0.5% agar containing HS medium instead of cornstarch. Photosynthetic colonies were selected on an HS agar plate in ordinary air.
cDNA Isolation and Northern Blotting.
RNA isolation from Chlamydomonas cells and Northern blot hybridization were performed as described (11). Poly(A)+ RNA was isolated with a polyATract kit (Promega). First-strand cDNA was synthesized with oligo(dT) nucleotides by using Superscript II (Life Technologies) in the presence of 0.6 M trehalose at 60°C (12). PCR was performed with GC buffer by using LA Taq polymerase (Takara Biomedical, Kyoto). Gene-specific oligonucleotide probes were generated according to the published sequences, e.g., Cah1 mRNAs (11), Mca (4), Lci1 (13), Att1 (14), Cah3 (15), and Cyp1 (16). A probe for Ccp2 mRNAs was generated by PCR, as described (5).
In Vivo Labeling and Immunoprecipitation.
Cells grown in HS medium lacking sulfur in high-CO2 conditions were transferred into two culture bottles and bubbled with 5% CO2 or ordinary air, respectively, then 2 μCi/ml (1 Ci = 37 GBq) of 35S-labeled methionine and cysteine mix (Amersham Pharmacia) was added to the cultures. After 12-hr incubation with 150 μE⋅m−2⋅s−1 at 28°C (E = einstein, 1 mol of photons), cells were lysed by sonication in a 50 mM Hepes buffer (pH 7.2) containing 150 mM NaCl, 5 mM EDTA, 0.5% Nonidet P-40, 0.5% Triton X-100, and 1 mM phenylmethylsulfonyl fluoride. The lysate was used for immunoprecipitation with rat anti-CCM1 antibody by using protein G-Sepharose 4FF (Amersham Pharmacia). To obtain anti-CCM1 antibody, a part of the CCM1 coding region was subcloned into an expression vector, pET15b (Novagen). A His-tagged CCM1 protein covering from Met-1 to Gly-499 was expressed in Escherichia coli BL21(DE3)pLysS and purified with Ni2+-NTA-agarose (Qiagen, Chatsworth, CA) followed by SDS/PAGE. This partial CCM1 protein was used for immunization of a rat.
Physiological Measurements.
The rates of photosynthesis were measured in a Clark-type O2 electrode (Chloroview 1, Hansatech Instruments, Pentney King's Lynn, UK) and the CO2-compensation concentration was determined by using gas chromatography (17). The intracellular DIC concentration and carbon fixation were measured by the silicon oil centrifugation method (7). Chlamydomonas cells were stained with 0.05% bromophenol blue in 0.1% HgCl2 (6) and were observed with an Axiophot microscope equipped with Nomarski differential interference optics (Zeiss).
Results
Isolation of a DNA Fragment That Complements the C16 Phenotype.
It was shown previously that the high-CO2-requiring mutant, C16, is defective in the ability to induce various aspects of the CCM, including induction of the periplasmic CA gene, Cah1. The mutant was also shown to contain the Nia1 gene (formerly named NIT1) as a single-copy tag in its genome (7). After crossing this C16 mutant with a nit− strain, the high-CO2-requiring phenotype cosegregated with nit+ (data not shown), suggesting that the pleiotropic mutation, named as ccm1 in C16, was tagged by the inserted Nia1 coding sequence. To isolate the Ccm1 gene, we determined the nucleotide sequences of the flanking regions from the Nia1 coding region. Four genomic clones were isolated from a genomic library of WT strain C9 by using specific oligonucleotide probes, C16–5 and C16–3, generated from the flanking sequences (Fig. 1). Transformation of one of the genomic clones, pKI4 (harboring a 38-kb insert of DNA), successfully restored the growth of the Chlamydomonas cells under low-CO2 (Fig. 2A). One of the transformants, named C16∷pKI4, recovered inducibility of the Cah1 under low-CO2 conditions (Fig. 2B). Southern blot analysis of DNA samples isolated from WT, C16, and the transformant C16∷pKI4 indicated that C16∷pKI4 contains both the 4.8-kb BamHI fragment detected in WT genomic DNA and the 9.4-kb BamHI fragment, which was generated by an insertion of Nia1 (Fig. 2C). Subsequent subcloning and transformation into C16 delimited the complementation region to a 5.1-kb XhoI–ApaI DNA fragment in pKI4XA (Fig. 2A). Therefore, the transformant, C16∷pKI4XA harboring this 5.1-kb XhoI–ApaI fragment, was characterized physiologically.
Figure 1.

Gene organization of Ccm1. The solid rectangles indicate the position of the protein-coding regions. The insertion site of the Nia1 gene in the C16 mutant is depicted by a large open triangle with an arrow. Positions of the oligonucleotide probes (C16–5 and C16–3) are shown by small triangles. Positions of a probe for the 1.5-kb EcoT22I DNA fragment (P-1) corresponding to the 5′-upstream region of Nia1, and Ccm1-specific probes (P-3 and P-5) are indicated by thick bars. A 4.8-kb BamHI fragment is between two BamHI sites. A, ApaI; B, BamHI; S, SacI; X, XhoI; (A)n, poly(A) tail.
Figure 2.

Complementation of C16 by transformation. (A) Relative positions of a genomic clone, pKI4, and its derivatives. Transformants of C16 cells, which photoautotrophically grew in ordinary air, are indicated by a +. (B) Northern blot analyses of Cah1 expression. Ten micrograms of total RNA from cells grown under high-CO2 (HC) or low-CO2 (LC) conditions were hybridized by a 32P-labeled Cah1-specific oligonucleotide probe. (C) Southern blot analysis of BamHI-digested total genomic DNA from WT, C16, and C16∷pKI4 cells. The blot was hybridized with a 32P-labeled Ccm1-specific probe, P-5.
A 5.1-kb Genomic DNA Fragment Complements the C16 Characteristics.
The transgenic strain C16∷pKI4XA grown under
low-CO2 conditions showed a high affinity against
DIC for photosynthesis with
Km(HCO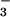 ) value of
122 μM, which is comparable to 145 μM of the WT (Fig.
3A). In addition, although the
C16 mutant was defective in DIC accumulation and carbon fixation,
C16∷pKI4XA accumulated DIC and fixed carbon similarly to WT cells
(Fig. 3 B and C). Moreover, C16∷pKI4XA
developed its pyrenoid structure in the chloroplast under
low-CO2 conditions, as did the WT, whereas C16
failed to do so (Fig. 4). These results
demonstrate that a single nuclear mutation at the ccm1 locus
resulted in multiple deficiencies in components of the CCM that are
needed to acclimate to CO2-limiting conditions.
) value of
122 μM, which is comparable to 145 μM of the WT (Fig.
3A). In addition, although the
C16 mutant was defective in DIC accumulation and carbon fixation,
C16∷pKI4XA accumulated DIC and fixed carbon similarly to WT cells
(Fig. 3 B and C). Moreover, C16∷pKI4XA
developed its pyrenoid structure in the chloroplast under
low-CO2 conditions, as did the WT, whereas C16
failed to do so (Fig. 4). These results
demonstrate that a single nuclear mutation at the ccm1 locus
resulted in multiple deficiencies in components of the CCM that are
needed to acclimate to CO2-limiting conditions.
Figure 3.

Photosynthetic characteristics of a transformant C16∷pKI4XA. (A) Relative rate of photosynthesis at various carbon concentrations of low-CO2-grown cells (pH 7.8) at 25°C. The maximum rates of O2 evolution were 90, 92, and 106 μmol⋅mg−1 of Chl⋅h−1 for WT, C16, and C16∷pKI4XA, respectively. (B) Intracellularly accumulated DIC concentration measured by silicone-oil centrifugation method (7). (C) Carbon fixation. The results are the average of three determinations, with standard deviations represented by bars. SIS, sorbitol-impermeable space.
Figure 4.

Pyrenoid structures in WT, C16, and C16∷pKI4XA. To visualize pyrenoids and nuclei, high-CO2 (HC)- and low-CO2 (LC)-grown cells were stained with bromophenol blue. Red arrowheads indicate positions of pyrenoids. (Bars = 10 μm.)
Induction of the CCM under low-CO2 stress is accompanied by an activation of the expression of a set of CCM-related genes, e.g., Cah1 (3), Mca encoding a mitochondrial CA (4), Ccp2 encoding the chloroplast envelope protein LIP-36 (5), Lci1 (13), and Att1 encoding the alanine α-ketoglutarate aminotransferase (14). Northern blot analyses using gene-specific probes indicated that activation of these five genes in low-CO2 conditions was recovered in C16∷pKI4XA (Fig. 5). Because RNA transcripts for these five genes were not detected in high-light (1,000 μE⋅m−2⋅s−1) and high-CO2 conditions, it is obvious that these genes are regulated preferentially by changes of CO2 availability. In addition, the mRNA transcripts of Cah3 encoding a chloroplastic CA (15) and Cyp1 encoding the cyclophilin (16), which are reported to be up-regulated under low-CO2 conditions in WT cells, were not detected in C16 but were detected in the C16∷pKI4XA under low-CO2 conditions. These results strongly suggest that the 5.1-kb genomic DNA fragment in pKI4XA encodes a key regulatory gene, which controls the expression of at least seven genes and modulates physiological properties and pyrenoid-development in response to CO2 availability.
Figure 5.

RNA blot analyses of total RNA. Ten micrograms of total RNA isolated from WT, C16, or C16∷pKI4XA cells grown under high-CO2 (HC) or low-CO2 (LC) was loaded. Cells were illuminated at 1,000 μE⋅m−2⋅s−1 (HL) or 150 μE⋅m−2⋅s−1 (ML). RNA blots were hybridized with probes for Cah1 coding for a periplasmic CA, Mca for a mitochondrial CA, Ccp2 for the chloroplastic envelope protein LIP-36, Lci1 for a low-CO2 inducible protein, Att1 for the alanine α-ketoglutarate aminotransferase, Cah3 for a chloroplastic CA, and Cyp1 for the cyclophilin. A 32P-labeled cDNA probe for Cblp encoding the G-protein β-subunit (29), which is expressed constitutively, was used as a loading control.
Structure and Expression of Ccm1.
Based on the nucleotide sequence of the 5,148-bp XhoI–ApaI genomic DNA fragment, which complements the ccm1 mutant phenotype, oligonucleotide primers were generated and used for reverse transcription–PCR to isolate corresponding cDNA. A 5,128-bp cDNA was isolated by using poly(A)+ RNA from 2-hr low-CO2-grown cells of the 5D strain. This cDNA consists of a 2,097-bp ORF encoding a 699-aa hydrophilic protein, a 176-bp 5′ untranslated region, and a 2,855-bp 3′ untranslated region (Fig. 1). Although the genomic clone complements the mutant phenotype, it lacks the 1,845-bp 3′ terminus of the cDNA. Because the sequence of the 502-bp 5′ upstream region did not contain any large ORF, the gene encoding this cDNA was assigned to be responsible to the complementation and was named Ccm1. To elucidate the entire structure of Ccm1, we determined the nucleotide sequence of the 8,474-bp region in the 12-kb XhoI genomic DNA cloned in pKI4Xh, which complements the C16 mutation (Fig. 2A). The Ccm1 mRNA is encoded by a 6,491-bp region in the 12-kb XhoI genomic region (Fig. 1). Sequence comparison of the flanking region around the Nia1 insertional site in C16 with the corresponding region in the WT revealed that the Nia1 was inserted into the coding region of the fourth exon of Ccm1 (Fig. 1). That Ccm1-specific probe P-5 hybridized with a 4.8-kb BamHI DNA fragment (Fig. 2C), a 12-kb XhoI fragment, and a 7.2-kb SacI fragment (data not shown) in the genomic Southern blot analyses, revealed that Ccm1 was a single-copy gene in C. reinhardtii.
In Northern blots using Ccm1-specific probes, P-5 and P-3 (Fig. 1), 5.1-kb mRNA was detected in high- and low-CO2 conditions in WT (lanes 1 and 2 in Fig. 6A). On the other hand, the mutant C16 did not accumulate the mRNA (lane 3 in Fig. 6A). These results indicate that Ccm1 is expressed constitutively in WT but the mature Ccm1 mRNA is not accumulated in C16 because of the Nia1 insertion.
Figure 6.

Expression of Ccm1 gene. (A) Ten micrograms of poly(A)+ RNA from WT in high-CO2 (lanes 1), WT in low-CO2 (lanes 2) and C16 in low-CO2 (lanes 3) were electrophoresed and hybridized with 32P-labeled Ccm1-specific probes P-5 (A a) and P-3 (A c). The same blot was reprobed by a 32P-labeled DNA fragment designated P-1 in Fig. 1 (A b) and by a Cblp cDNA (A d). (B) High-CO2-grown WT cells (lane 1), low-CO2-grown WT cells (lane 2), and low-CO2-grown C16 cells (lane 3) were labeled with [35S]-methionine and [35S]-cysteine, and immunoprecipitated with anti-CCM1 antibody. As a control, low-CO2-grown WT cells were labeled and immunoprecipitated with rat normal serum (lane 4).
The 5,128-bp Ccm1 cDNA encodes a 699-aa polypeptide with no apparent transmembrane domains. To identify the CCM1 protein in the cell, total protein in WT and C16 cells was labeled with [35S]-methionine and [35S]-cysteine in vivo and was immunoprecipitated with anti-CCM1 antibody. After SDS/polyacrylamide gel separation, a single band with a molecular mass of 76 kDa was detected in WT cells by autoradiogram in high- and low-CO2 conditions (lanes 1 and 2 in Fig. 6B), indicating that the CCM1 protein abundance was not affected by a change of CO2 level, as was the case with the mRNA transcripts. In the C16 mutant, a single band with a molecular mass of 80 kDa was detected (lane 3 in Fig. 6B). Considering that a 2.2-kb mRNA transcript was detected by a 32P-labeled probe, P-5 (lane a3 in Fig. 6A), as well as probe P-1 corresponding to the 5′ part of the inserted-tag DNA containing the Nia1 gene (lane b in Fig. 6A and see also Fig. 1), and that no hybridization signal was detected by probe P-3 (lane c3 in Fig. 6A) in the C16 mutant, this 80-kDa protein in C16 could be generated from a 2.2-kb fusion-mRNA transcript made from a 5′ part of Ccm1 and the 5′ part of the inserted-tag DNA.
CCM1 Shares Significant Sequence Similarity to Zinc-Finger Domains.
When the deduced amino acid sequence of CCM1 was used for a similarity search against the nonredundant protein databases, no sequences showing high levels of similarity overall were detected. However, three characteristic sequence stretches were identified (Fig. 7A). First, a 27-aa sequence stretch, FxCxxxxCxRxFxxxxxLxxHxxxxxD (from Phe-34 to Asp-60), showing significant similarities with those of C2H2-type zinc-finger motifs in other organisms, which function in protein–DNA or protein–protein interactions (ref. 18; Fig. 7B); e.g., ZNF76 from Homo sapiens, TFIIIa-2 from Xenopus laevis, ZFH2 and Krüppel from Drosophila melanogaster, EPF1–1 from Petunia hybrida, and WZF1 from Triticum aestivum. Second, a Gln-repeat in the middle of the protein spanning from Gln-317 to Gln-330. Third, a Gly-rich domain at the C-terminal region spanning from Gly-555 to Gly-628.
Figure 7.

Amino acid sequence of CCM1 protein. (A) Predicted amino acid sequence of CCM1 is shown as one-letter symbols. A putative zinc-finger motif is highlighted and a Gln-repeat region is italicized. The His residue at position 54, which is changed into Tyr in the cia5 mutant, is depicted as His-54. Asp-183 and Arg-184 and a Gly-rich region are underlined. (B) Sequence comparison of the putative zinc-finger motif of CCM1 with zinc-finger motifs in other organisms: Homo sapiens (Hom) ZNF76 (NM_003427), Xenopus laevis (Xen) TFIIIa2 (K02938), Drosophila melanogaster (Dro) ZFH2 (P28167), Dro Krüppel (X03414), Petunia hybrida (Pet) EPF1–1 (X60700), and Triticum aestivum (Tri) WZF1 (D16415). Filled circles denote zinc-liganding amino acids. Conserved amino acids are highlighted.
Identification of a Mutation Site of Ccm1 in the High-CO2-Requiring Mutant, cia5.
Because the pleiotropic phenotype of C16 was similar to that of another high-CO2-requiring mutant, cia5 (7), it was necessary to know whether Ccm1 could complement the cia5 mutation. When the pKI4XA plasmid was transformed into the cia5 mutant cells, many colonies appeared on HS agar plates in low-CO2 condition. One of the transformants, named cia5∷pKI4XA, restored the inducibility of Cah1 expression under low-CO2 conditions (Fig. 2B). In addition, induction of other CCM-related genes and pyrenoid development under low-CO2 conditions also were recovered, as was the case for C16 (data not shown). To identify the cia5 mutation site, we isolated a genomic clone containing a 12-kb XhoI fragment from cia5 cells by using the Ccm1-specific probe P-3 (Fig. 1) and determined the nucleotide sequence of the 8,474-bp cia5 genomic region corresponding to that of Ccm1. This determination revealed that the cia5 mutant had a single point mutation from T to C in the second exon of Ccm1. As a consequence of this base substitution, His-54 in the putative zinc-finger motif of the CCM1 protein was replaced by Tyr in the cia5 mutant (Fig. 7). These findings of functional complementation of the cia5 mutant and the mutation site in cia5 strongly suggest that this His-54 in the CCM1 plays a critical role in the regulation of CCM, including the induction of a set of CCM-related genes and pyrenoid development.
Discussion
In this article we have identified a Ccm1 gene encoding a hydrophilic protein, which is shown to be essential to regulate CCM induction in the eukaryote C. reinhardtii by sensing CO2 availability. The gene product, CCM1, appears to control the expression of at least seven CCM-related genes—Cah1, Mca, Ccp2, Lci1, Att1, Cah3, and Cyp1—as well as the induction of CCM functions, including active DIC transport and the development of pyrenoid structure in the chloroplast during acclimation processes in CO2-limiting stress conditions. It is shown that the mutant cia5 forms the pyrenoid starch sheath and suggested that the starch sheath is not involved in the operation of CCM (19). Therefore, it is most likely that Ccm1 does not regulate genes controlling the formation of the pyrenoid starch sheath but those for the development of the pyrenoid structure. Because Ccm1 seems to be at a higher position in gene hierarchy and 2,665 low-CO2-specific expressed sequence tag groups have been described in this organism (20), it is possible that some of other unidentified low-CO2-inducible genes also are regulated by Ccm1. In those genes controlled by Ccm1, there should be genes that encode CCM components such as CO2 or bicarbonate transporters as well as enzymes for the development of the pyrenoid structure. Because a pleiotropic mutant, which does accumulate DIC by constitutively active CCM even in a high-CO2 condition, has been isolated from Chlorella elipsoidia (17), Ccm1 orthologues might also operate in other aquatic photosynthetic organisms.
Candidates for the nature of the signal that induces response to ambient CO2 level are proposed to be the total inorganic carbon level, the ratio [CO2]/[O2], or [CO2(aq)] (reviewed in ref. 1). It is possible that this CO2 signal is transduced through kinase cascades, because the CCM1 protein with 14 putative phosphorylation sites is essential to the low-CO2 signal transduction. Because it is reported that CCM and the Cah1 expression are affected by light regime and also the phase of the cell cycle (21), Ccm1 transcription and modification of the CCM1 protein may depend on light regime or the phase of the cell cycle.
Based on sequence similarities, CCM1 is possibly one of the C2H2-type zinc-finger transcription factors, although the Asp-60 in CCM1, corresponding to the second His in the C2H2 motif, is not conserved (Fig. 7B). Because Asp serves as a zinc ligand in other cellular components such as adenylate kinase from Bacillus subtilis (22) and ferredoxin from Sulfolobus (23), it is possible that Asp ligands to zinc to form a finger-like secondary structure as predicted in other transcriptional factors with C2H2-type zinc-finger motifs (24). Moreover, other amino acid residues such as Phe-34, Arg-43, Phe-45, and Leu-51, and β-sheet structures in this domain of CCM1 are highly conserved with other zinc-containing transcription factors (24). It is also possible that His or Cys around the motif could serve as a zinc ligand. Identification of a mutation site in cia5 revealed that His-54 in the putative zinc-finger motif in CCM1 is essential to transduce a low-CO2 signal into regulatory systems for CCM-related genes (Fig. 7). Taken with the fact that CCM1 also contains a Gln-repeat stretch, which is reported to be necessary for the regulatory functions of several eukaryotic transcription factors (25), CCM1 may interact directly with DNA or regulatory proteins through putative zinc-finger domain(s) and activate the transcription of CCM-related genes under low-CO2 conditions. Although the coding region for this putative zinc-finger domain of CCM1 is expressed in the mutant C16, the CO2-responsive regulation of CCM is impaired. Therefore, it is obvious that the C-terminal region of CCM1 is essential to transduce low-CO2 signal in addition to the putative zinc-finger domain.
The molecular mass of the CCM1 was estimated to be 76 kDa by an in vivo labeling experiment (Fig. 6B), which is slightly larger than that of 70 kDa deduced from the cDNA nucleotide sequence. In addition, CCM1 has 14 putative phosphorylation sites, three putative glycosylation sites, and putative myristylation sites. The level of CCM1 protein accumulation does not change during acclimation to low-CO2. These findings suggest that modifications of CCM1 protein under specific conditions, such as high- or low-CO2 stress conditions, might occur posttranslationally in response to CO2 availability. Further biochemical characterization of CCM1 protein may reveal the structure–function relationships of the CCM1 protein, concerning its regulatory roles in CCM induction processes.
During the course of the isolation of cDNA for CCM1, namely CCM1-A, a 3-bp shorter cDNA of 5,125 bp encoding another CCM1 protein, named CCM1-B, also was isolated from Chlamydomonas cells. Sequence comparison between two kinds of cDNAs and Ccm1 revealed that the shorter cDNA was generated by alternative splicing between the third and fourth exons, causing a 3-nt deletion. This shorter cDNA encodes a 698-aa CCM1-B, which has a substitution of Glu-183 instead of Asp-183 in CCM1-A and a deletion of Arg-184 present in CCM1-A (see Fig. 7A). It is possible that heterologous CCM1 proteins are translated simultaneously from the alternatively spliced mRNA transcripts in the cells. The influence of this heterogeneity in the CCM1 proteins on their regulatory functions remains to be elucidated. Recently, Cia5 was deposited in the GenBank database (accession no. AF317732) by Y. Xiang, J. Zang, and D. P. Weeks. Its gene product has an amino acid sequence identical to that of CCM1-B.
In Chlamydomonas, several regulatory genes have been identified, e.g., SacI (26), Psr1 (27), and Crd1 (28), which control the acclimation to limitation of sulfur, phosphorus, and copper ion, respectively. Although nuclear localization of the Psr1 gene product has been reported, no nuclear localization signal has been elucidated so far in Chlamydomonas. Therefore, there may be Chlamydomonas-specific nuclear localization signals in these regulatory proteins as well as in CCM1. Biochemical characterization of Chlamydomonas CCM1 protein would lead to a better understanding of CO2 sensing and regulatory control of the CCM in eukaryotic photosynthetic organisms. This discovery also may open the door to a greater understanding of crosstalk between signal transduction cascades that are involved in acclimation of eukaryotic photosynthetic organisms to environmental stresses such as nutrient shortage.
Acknowledgments
We thank Shoji Matsueda, Yuri Kobayashi, and Kiwako Hayashi for their technical assistance. This work was supported by Grants-in-Aid from the Japanese Ministry of Education, Science, and Culture (nos. 12025217 and 12660300) and the Japan Society for the Promotion of Science Grant JSPS-RFTF97R16001 (to H.F.).
Abbreviations
- CA
carbonic anhydrase
- CCM
carbon-concentrating mechanism
- DIC
dissolved inorganic carbon
- high-CO2
air enriched with 5% CO2
- low-CO2
air containing 0.04% CO2
- WT
wild type
- HS
high salt
Footnotes
References
- 1.Kaplan A, Reinhold L. Annu Rev Plant Physiol Plant Mol Biol. 1999;50:539–570. doi: 10.1146/annurev.arplant.50.1.539. [DOI] [PubMed] [Google Scholar]
- 2.Badger M R, Kaplan A, Berry J A. Plant Physiol. 1980;66:407–413. doi: 10.1104/pp.66.3.407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Fukuzawa H, Fujiwara S, Yamamoto Y, Deonisio-Sese M L, Miyachi S. Proc Natl Acad Sci USA. 1990;87:4383–4387. doi: 10.1073/pnas.87.11.4383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Eriksson M, Karlsson J, Ramazanov Z, Gardeström P, Samuelsson G. Proc Natl Acad Sci USA. 1996;93:12031–12034. doi: 10.1073/pnas.93.21.12031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Chen Z-Y, Lavigne L L, Mason C B, Moroney J V. Plant Physiol. 1997;114:265–273. doi: 10.1104/pp.114.1.265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kuchitsu K, Tsuzuki M, Miyachi S. Can J Bot. 1991;69:1062–1069. [Google Scholar]
- 7.Fukuzawa H, Ishizaki K, Miura K, Matsueda S, Ino-ue T, Kucho K, Ohyama K. Can J Bot. 1998;76:1092–1097. [Google Scholar]
- 8.Moroney J V, Husic H D, Tolbert N E, Kitayama M, Manuel L J, Togasaki R K. Plant Physiol. 1989;89:897–903. doi: 10.1104/pp.89.3.897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Harris E H. The Chlamydomonas Sourcebook. San Diego: Academic; 1989. [Google Scholar]
- 10.Shimogawara K, Fujiwara S, Grossman A R, Usuda H. Genetics. 1998;148:1821–1828. doi: 10.1093/genetics/148.4.1821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kucho K, Ohyama K, Fukuzawa H. Plant Physiol. 1999;121:1329–1337. doi: 10.1104/pp.121.4.1329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Carninci P, Nishiyama Y, Westover A, Itoh M, Nagaoka S, Sasaki N, Okazaki Y, Muramatsu M, Hayashizaki Y. Proc Natl Acad Sci USA. 1998;95:520–524. doi: 10.1073/pnas.95.2.520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Burow M D, Chen Z-Y, Mouton T M, Moroney J V. Plant Mol Biol. 1996;31:443–448. doi: 10.1007/BF00021807. [DOI] [PubMed] [Google Scholar]
- 14.Chen Z-Y, Burow M D, Mason C B, Moroney J V. Plant Physiol. 1996;112:677–684. doi: 10.1104/pp.112.2.677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Karlsson J, Clarke A K, Chen Z-Y, Hugghins S Y, Park Y-I, Husic H D, Moroney J V, Samuelsson G. EMBO J. 1998;17:1208–1216. doi: 10.1093/emboj/17.5.1208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Somanchi A, Moroney J V. Plant Mol Biol. 1999;40:1055–1062. doi: 10.1023/a:1006262123918. [DOI] [PubMed] [Google Scholar]
- 17.Matsuda Y, Coleman B. Plant Physiol. 1996;110:1283–1291. doi: 10.1104/pp.110.4.1283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Mackay J P, Crossley M. Trends Biochem Sci. 1998;23:1–4. doi: 10.1016/s0968-0004(97)01168-7. [DOI] [PubMed] [Google Scholar]
- 19.Villarejo A, Martínez F, Plumed M P, Ramazanov Z. Physiol Plant. 1996;98:798–802. [Google Scholar]
- 20.Asamizu E, Miura K, Kucho K, Inoue Y, Fukuzawa H, Ohyama K, Nakamura Y, Tabata S. DNA Res. 2000;7:305–307. doi: 10.1093/dnares/7.5.305. [DOI] [PubMed] [Google Scholar]
- 21.Marcus Y, Schuster G, Michaels A, Kaplan A. Plant Physiol. 1986;80:604–607. doi: 10.1104/pp.80.2.604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Perrier V, Surewicz W K, Glaser P, Martineau L, Craescu C T, Fabian H, Mantsch H H, Bârzu O, Gilles A-M. Biochemistry. 1994;33:9960–9967. doi: 10.1021/bi00199a019. [DOI] [PubMed] [Google Scholar]
- 23.Fujii T, Hata Y, Wakagi T, Tanaka N, Ohsima T. Nat Struct Biol. 1996;3:834–837. doi: 10.1038/nsb1096-834. [DOI] [PubMed] [Google Scholar]
- 24.Pabo C O, Sauer R T. Annu Rev Biochem. 1992;61:1053–1095. doi: 10.1146/annurev.bi.61.070192.005201. [DOI] [PubMed] [Google Scholar]
- 25.Courey A J, Tjian R. Cell. 1988;55:887–898. doi: 10.1016/0092-8674(88)90144-4. [DOI] [PubMed] [Google Scholar]
- 26.Davies J P, Yildiz F H, Grossman A R. EMBO J. 1996;15:2150–2159. [PMC free article] [PubMed] [Google Scholar]
- 27.Wykoff D D, Grossman A R, Weeks D P, Usuda H, Shimogawara K. Proc Natl Acad Sci USA. 1999;96:15336–15341. doi: 10.1073/pnas.96.26.15336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Moseley J, Quinn J, Eriksson M, Merchant S. EMBO J. 2000;19:2139–2151. doi: 10.1093/emboj/19.10.2139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Schloss J A. Mol Gen Genet. 1990;221:443–452. doi: 10.1007/BF00259410. [DOI] [PubMed] [Google Scholar]