

Abstract
The present study was designed to investigate the cardiac benefits of M3 muscarinic receptor (M3-mAChR) overexpression and whether these effects are related to the regulation of the inward rectifying K+ channel by microRNA-1 (miR-1) in a conditional overexpression mouse model. A cardiac-specific M3-mAChR transgenic mouse model was successfully established for the first time in this study using microinjection, and the overexpression was confirmed by both reverse transcriptase–polymerase chain reaction and Western blot techniques. We demonstrated that M3-mAChR overexpression dramatically reduced the incidence of arrhythmias and decreased the mortality in a mouse model of myocardial ischemia-reperfusion (I/R). By using whole-cell patch techniques, M3-mAChR overexpression significantly shortened the action potential duration and restored the membrane repolarization by increasing the inward rectifying K+ current. By using Western blot techniques, M3-mAChR overexpression also rescued the expression of the inward rectifying K+ channel subunit Kir2.1 after myocardial I/R injury. This result was accompanied by suppression of upregulation miR-1. We conclude that M3-mAChR overexpression reduced the incidence of arrhythmias and mortality after myocardial I/R by protecting the myocardium from ischemia in mice. This effect may be mediated by increasing the inward rectifying K+ current by downregulation of arrhythmogenic miR-1 expression, which might partially be a novel strategy for antiarrhythmias, leading to sudden cardiac death.
INTRODUCTION
Sudden cardiac death is a major contributor to mortality in industrialized nations, affecting around 500,000 individuals annually in the Western world. It can also cause more deaths than acquired immunodeficiency syndrome (AIDS), lung and breast cancer and stroke combined (1). In China, preliminary data suggest that there are over 544,000 adults with sudden cardiac death each year (2). About 75% of the sudden cardiac death cases are linked to a previous myocardial infarction, and 80% show a clinical record with the history of coronary artery disease (2). Intriguingly, life-threatening cardiac arrhythmias, one of the leading causes of sudden cardiac death, have been clinically demonstrated in the vast majority of severe heart diseases, especially myocardial infarction. Unfortunately, inefficacy of conventional antiarrhythmic drugs has also been recognized in healing the electrical remodeling process or reducing sudden cardiac death (3,4). Therefore, the primary purpose of the present study was designed to develop rational approaches to prevent lethal cardiac arrhythmias that may lead to a consequent sudden cardiac death.
Since the initial identification of the M3 subtype of the muscarinic acetylcholine receptors (M3-mAChR) in the heart, there have been increasing interest and advances in studies on the pathophysiological roles of M3-mAChR in the heart (5). Recent studies from several laboratories have provided compelling and solid evidence in support of the important roles of M3-mAChR in the regulation and maintenance of cardiac function and heart disease (6–8). M3-mAChR activates a delayed rectifying K+ current IKM3 to participate in cardiac repolarization, negative chronotropic actions and antidysrhythmic activity (suppresses ischemic dysrhythmias) (9). M3-mAChR interacts with gap-junctional channel connexin 43 to maintain cell–cell communication and excitation propagation (10). M3-mAChR regulates intracellular phosphoinositide hydrolysis to improve cardiac contraction and hemodynamic function (11). M3-mAChR also activates antiapoptotic signaling molecules, enhances endogenous antioxidant capacity and diminishes intracellular Ca2+ overload, all of which contribute to protecting the heart against ischemic injuries (12). However, it is generally difficult to predict the simultaneous involvement of two or more mAChR subtypes in a specific functional response by using antagonists of limited subtype selectivity. Development of transgenic mice overexpressing specific mAChR subtypes seems to allow a more definitive approach to defining the pathophysiological roles of mAChRs. To further explore the potential roles of M3-mAChR in myocardial ischemia and sudden cardiac death, we created a transgenic mouse model in which M3-mAChR was overexpressed conditionally.
There is a growing body of evidence showing that the dysfunction of the potassium channel is closely related to cardiac diseases (13). Among them, the Kir family, which expresses two transmembrane-spanning segments flanking a conserved pore region, is structurally and functionally different from other ion channels. Kir2.1 subunits, encoded by the potassium inwardly-rectifying channel, subfamily J, member 2 (KCNJ2) gene, is the main K+ channel subunit that mediates inward rectifier potassium channel current (IK1) in cardiomyocytes (14), which plays a key role in setting up the resting membrane potentials and in the repolarization of cardiac myocytes, especially affecting the action potential duration (APD) at 90% repolarization (APD90) (15). Many studies have demonstrated that IK1 is dys-functional in several cardiac diseases such as cardiac ischemia-reperfusion (I/R) injury, arrhythmia and heart failure (16–18).
Since the discovery of microRNA, a single-stranded noncoding RNA of approximately 22 nucleotides long and >700 miRNAs have been identified in humans (19), which regulates up to 20–30% of gene expression by binding to 3′ untranslated regions (UTRs) of mRNAs through inexact sequence matching, targeting numerous mRNAs for translational inhibition or degradation (20). For example, the overexpressed microRNA-1 (miR-1) is arrhythmogenic either in ischemic or normal hearts, which could slow the conduction and depolarized the cytoplasmic membrane by posttranscriptionally repressing KCNJ2 (21).
This study was designed to investigate the cardiac beneficial effects of M3-mAChR overexpression and whether they are related to the regulation of inward rectifying K+ channel by miR-1 and how the molecular processes within the cardiac cell are controlled after the overexpression of M3-mAChR. These can definitely help us better understanding the pathology of myocardial ischemia that are associated with the regulation of gene expression and transcriptional activity, which also might be a novel strategy for preventing sudden cardiac death.
MATERIALS AND METHODS
Vector Construction and Generation of M3-mAChR Transgenic Mouse
The cDNA (1,836 bp) of M3-mAChR from the University of Missouri–Roller (UMR) cDNA Resource Center (www.cdna.org) was cloned into α-myosin heavy chain (α-MHC) vector (a gift from Dr J. Robbins, Children’s Hospital Research Foundation, Cincinnati, OH, USA). M3-mAChR cDNA was subcloned between SalI and HindIII of the α-MHC expression cassette. The construct also contains a human growth hormone polyadenylylation (Hgh polyA) sequence. The M3-mAChR cDNA was amplified by the PCR, and the vector was extracted and cut using the SpeI restriction enzyme (Figure 1A). The DNA was then prepared for microinjection to produce transgenic mouse via cDNA microinjection of fertilized 562-L oocytes according to established techniques (22) (see the online supplementary materials).
Figure 1.

Characterization of expression levels of M3-mAChR in α-MHC promoter M3-mAChR transgenic gene (M3Tg) mice. (A) Schematic of the M3-mAChR gene constructed in the α-MHC promoter-M3Tg. The 13-kb transgene construct used to produce the transgenic mice was composed of cDNA (1,836 bp) of M3 from the UMR cDNA Resource Center, subcloned between SalI and HindIII of the α-MHC expression cassette. (B) The mRNA expression of M3-mAChR determined by real-time RT-PCR of total RNA in WT and M3Tg mice. (C) Top, examples of Western blot bands; bottom, expression levels of M3-mAChR. (D) Raw traces of IKM3 recorded from WT and M3Tg mice. (E) I-V relationships of IKM3 currents in WT and M3Tg mice. The average data presented were mean ± SEM; *P < 0.05, **P < 0.01 versus WT.
Mouse Model of Myocardial Ischemia Reperfusion
The 562-L male M3 transgenic (M3Tg) mice and their wild-type (WT) litter-mates, weighing approximately 25 g, were subjected to the sham or myocardial I/R model as previously described (23). The ECG and mortality were observed in each group to evaluate the severity of arrhythmia. Then, the hearts were removed for ventricular cardiomyocyte isolation, Western blot and real-time reverse transcriptase–polymerase chain reaction (RT-PCR) assay. This study was approved and supervised by the Institutional Animal Care and Use Committee of Harbin Medical University. Use of animals was in accordance with the regulations of the ethic committees of Harbin Medical University and was confirmed with the Guide for the Care and Use of Laboratory Animals, published by the U.S. National Institutes of Health (NIH publication no. 85-23, revised 1996) (see the online supplementary materials).
Arrhythmia Scores
The incidence of arrhythmias and the mortality rate were registered and evaluated for a continuous period of 2 h after myocardial I/R injury or sham surgery. An arrhythmia score was used to include the incidence and duration of arrhythmias by giving a grade to the animals (24) (see the online supplementary materials).
Arrhythmic Model Induced by Aconitine in Mice
Aconitine (5 mg/mL, 2 μg/10 g mice; Sigma, St. Louis, MO, USA) was pumped at a constant speed using Longerpump (LSP02-1B; Baoding Longer Precision Pump Company, Baoding, China) to induce the ventricular arrhythmias within 4 min. In each group, the arrhythmia was observed until the death of the mice. The survival time was estimated in WT and M3Tg mice.
Ventricular Cardiomyocyte Isolation and Patch-Clamp Recording
The dissociation procedures for ventricular cardiomyocytes were similar to that described previously (24). The cardiomyocytes were transferred to a chamber mounted on an inverted microscope (Nikon Diaphot; Nikon, Tokyo, Japan) for electrophysiological recording. Action potential (AP) and potassium currents (K+) were recorded under current and voltage-clamp configuration, respectively, using the whole-cell patch-clamp technique with an Ax-opatch 200B amplifier (Axon Instruments, Foster City, CA, USA) (see the online supplementary materials).
Western Blot Analysis
The procedures for membrane protein extraction from ventricular tissue and immunoblotting are described in detail elsewhere (12). The primary antibodies of anti-Kir2.1, anti-Kv4.2 (Alomone Labs, Jerusalme, Israel), and anti-Cav1.2, anti-M3 (Santa Cruz Technology, Santa Cruz, CA, USA), were used, with GAPDH (anti-GAPDH antibody from Kangcheng, Shanghai, China) as an internal control. Western blot bands were quantified using Odyssey v1.2 software by measuring the band intensity (area × optical density [OD]) for each group (see the online supplementary materials).
Determination of M3-mAChR mRNA, KCNJ2, KCND2, CACNA1C and miR-1 Levels
RT-PCR assay was carried out to quantify the levels of M3-mAChR mRNA, KCNJ2, KCND2 (potassium voltage-gated channel, Shal-related subfamily, member 2), CACNA1C (calcium channel, voltage-dependent, L type, alpha 1C subunit) transcripts and miR-1. The total RNA samples were isolated with a mirVana™ miRNA Isolation Kit (Ambion Inc., Austin, TX, USA) from mouse myocardial I/R. The miR-1 levels were measured using the mirVana™ qRT-PCR miRNA Detection Kit (Ambion), which is a kind of quantitative RT-PCR kit and was used in conjunction with real-time PCR with SYBR Green I for quantification of miR-1 transcripts (see the online supplementary materials).
Data Analysis
Average data were expressed as mean ± standard error of the mean (SEM). Statistical comparisons (performed using analysis of variance [ANOVA]) were carried out using SPSS 13.0. A two-tailed P value <0.05 indicated a statistically significant difference. Nonlinear least-square curve fitting was performed with CLAMPFIT in pCLAMP 10.0 or GraphPad Prism.
All supplementary materials are available online at www.molmed.org.
RESULTS
Characterization of M3-mAChR in Tg Mice
Our data showed that the mRNA levels of M3-mAChR were higher in M3-overexpressing mice than in their WT counterparts (Figure 1B; P < 0.01). Meanwhile, the protein expression of M3-mAChR in M3-overexpressing mice (1.56 ± 0.17) was also higher than that in WT mice (Figure 1C; P < 0.05). To further confirm the successful establishment of M3-mAChR overexpression, whole-cell patch-clamp experiments were performed using ventricular cardiomyocytes in the presence of 2 mmol/L 4-aminopyridine (4-AP) in the perfusate to antecedently block the transient outward rapid K+ current (Ito). As shown in Figures 1D and E, 5 mmol/L choline (an agonist of M3 muscarinic receptor; Sigma) was applied to the bath. The voltage-clamp data showed that choline activated a delayed rectifier type of current (IKM3) when depolarizing voltage steps were applied.
The IKM3 density in ventricular cardiomyocytes of M3Tg mice at +50 mV (46.3 ± 5.8 pA/pF) was higher than that of WT mice (15.4 ± 2.5 pA/pF, P < 0.01 WT versus M3Tg mice).
Cardiac Beneficial Effects of M3-mAChR Overexpression against Mice Myocardial I/R and Aconitine Injury
To detect the protective role of over-expression of M3-mAChR in myocardial I/R injury, the WT and M3Tg mice were subjected to 30 min of ischemia and then 2 h of reperfusion. Representative electrocardiogram after myocardial I/R is shown in Figure 2A, and the abnormalities included ventricular fibrillation, ventricular tachyarrhythmia, premature ventricular contraction, bigeminy and atrioventricular block. The onset of atrial ventricular block after myocardial I/R injury in M3Tg mice (58.4 ± 15.3 min) were prolonged compared with that in WT I/R mice (17.4 ± 5.6 min, P < 0.01). The duration of atrial ventricular block in M3Tg I/R mice (7.27 ± 2.56 min) was just slightly shortened compared with that in WT I/R mice (11.8 ± 2.2 min), but there was no statistical significance. Overexpression of M3-mAChR lowered the arrhythmia score after myocardial I/R from 3.56 ± 0.39 (n = 32 for WT I/R mice) to 1.78 ± 0.31 (n = 33 for M3Tg I/R mice, P < 0.01). The mortality rate was then determined as the percentage of mice that initially recovered from surgery but subsequently died. M3Tg I/R mice tended to have a lower mortality rate: 9% (3 out of 33) for the M3Tg I/R mice versus 28% (9 out of 32) for the WT I/R mice (Figure 2E). None of the sham M3Tg mice or WT controls died during or after the operation.
Figure 2.

Cardiac beneficial effects of M3-mAChR overexpression against mice myocardial I/R and aconitine injury. (A) Representative electrocardiogram after mice myocardial I/R. The abnormalities include ventricular fibrillation, ventricular tachyarrhythmia, premature ventricular contraction, bigeminy and atrioventricular block. (B) Onset of atrial ventricular block (AVB). (C) Duration of AVB. (D) Arrhythmia score. (E) Mortality after experimental myocardial I/R. *P < 0.05, **P < 0.01 versus WT I/R. (F) Survival time after aconitine-induced arrhythmias in WT and M3Tg mice. **P < 0.01 versus WT + Aco. Aco, aconitine (5 mg/mL, 2 μg/10 g mice). The average data were presented as mean ± SEM.
Aconitine is a highly poisonous alkaloid derived from various aconite species and is used for creating models of cardiac arrhythmia (25). The arrhythmia was also induced by aconitine, and the survival time was obviously prolonged in M3Tg mice compared with WT mice (Figure 2G; P < 0.01).
M3-mAChR Overexpression Shortened APD90 of Ventricular Cardiomyocytes
Figure 3A showed representative AP recorded in ventricular cardiomyocytes derived from WT and M3Tg mice with or without myocardial I/R. The resting membrane potential (RMP) (−63.4 ± 1.5 mV) was significantly hyperpolarized in M3Tg mice compared with that in WT mice (−54.3 ± 0.8 mV; Figure 3B, top; P < 0.05 WT versus M3Tg). Obviously, the APD90 was shortened significantly in M3Tg mice compared with that in WT mice after myocardial I/R (P < 0.05), although there was no significant changes before myocardial I/R (Figure 3B, bottom right panel). Action potential duration at 50% (APD50) was not significantly changed between strains in all groups (Figure 3B, bottom left panel). We also measured the ratio of APD50/APD90 and found no significant difference in different experimental groups (data not shown). Comparison of AP characteristics of ventricular cardiomyocytes from both WT and M3Tg mice with or without myocardial I/R were summarized in Table 1.
Figure 3.

APD characteristics in M3Tg mice using a whole-cell patch-clamp. (A) Representative traces of AP recorded from both WT and M3Tg mice with or without myocardial I/R. (B) RMP. (C) Pooled data for APD at 50% (APD50) repolarization. (D) Pooled data for APD at 90% (APD90) repolarization. The average data were presented as mean ± SEM. *P < 0.05, **P < 0.01 versus WT or WT I/R.
Table 1.
AP characteristics of cardiac myocytes in WT and M3-overexpressing (M3Tg) mice from both control and myocardial I/R injury.
| Group | na | RMP (mV) | Overshoot (mV) | Amplitude of action (mV) | APD (ms) |
|---|---|---|---|---|---|
| WT | 6 | −54.3 ± 0.8 | 40.9 ± 3.2 | 95.3 ± 3.7 | 182.7 ± 16.0 |
| M3Tg | 6 | −63.3 ± 1.5b | 40.7 ± 3.1 | 104.1 ± 3.4 | 130.2 ± 16.7c |
| WT I/R | 14 | −54.5 ± 0.9 | 32.1 ± 2.6 | 86.7 ± 3.5 | 161.1 ± 10.4 |
| M3Tg I/R | 14 | −56.1 ± 1.2 | 34.8 ± 2.6 | 90.9 ± 3.5 | 131.6 ± 7.9d |
Data are mean ± SEM.
The number of cardiac myocytes.
P < 0.01,
P < 0.05 versus WT group.
P < 0.05 versus WT I/R group.
M3-mAChR Overexpression Reduced IK1 but Not Ito and ICa,L in Ventricular Cardiomyocytes
IK1 plays an important role in setting up the RMP and in modulating the APD90 of the ventricular AP (15). To investigate whether the shortened APD90 and hyperpolarized RMP was accompanied by IK1 change, we compared IK1 in ventricular cardiomyocytes between WT and M3Tg mice both in control and I/R hearts. Figure 4A showed typical examples of IK1, which was elicited by a series of 300-ms depolarizing test potentials (TP) from −120 to +50 mV, with 10-mV increments from a holding potential (HP) of −40 mV. IK1 was substantially increased in M3Tg compared with that in WT mice, as illustrated in Figure 4B (P < 0.05, n = 10 cells from six mice). While IK1 from both M3Tg I/R and WT I/R mice was substantially decreased, the current density of IK1 in M3Tg I/R mice was higher than that in WT I/R mice (see Figure 4B; P < 0.05; n = 10 cells from six mice). Ito was defined as the difference between the peak outward current (Ipeak) and the sustained component remaining (Ipeak) at the end of the depolarizing pulse (Ito = Ipeak − Isus). The mean current-voltage relationship for Ito was illustrated in Figure 4D. Ito current density was not significantly changed between strains in all groups (Figures 4C, D; n = 8 cells from six mice). Our result indicated that L-type Ca2+ current (ICa,L) densities were maximal at 0 mV in four groups. Although peak densities of ICa,L were increased in M3Tg mice compared with that in WT mice with or without myocardial I/R, there was no statistical significance (P > 0.05; n = 8 cells from six mice; Figures 4E, F).
Figure 4.

Comparison of the inward rectifier K+ current (IK1) and transient outward K+ currents (Ito) from both WT and M3Tg mice with or without myocardial I/R. (A) Raw traces of IK1 recorded from WT and M3Tg mice. (B) The averaged current density–voltage (I–V) relationships of IK1. (C) Raw traces of Ito recorded from WT and M3Tg mice. (D) In the average I–V relationships of Ito, there were no differences among the four groups. I–V curves, mean from at least eight cells for each condition. (E) Raw traces of ICa,L recorded from WT and M3Tg mice. (F) In the average I–V relationships of ICa,L, there were no significant differences between the four groups. I–V curves, mean from at least eight cells for each condition. The average data were presented as mean ± SEM.
M3-mAChR Overexpression Altered Protein and mRNA Levels of Potassium and Calcium Channel Subunits in Myocardium
The Kir2.1 subunit is the main K+ channel subunit that mediates IK1 in cardiomyocytes (14). To investigate whether the changed IK1 is the result of Kir2.1 activation, we examined the expression level of Kir2.1 by Western blot experiments. The expression level of Kir2.1 was 2.69 times higher in the myocardium of M3Tg mice than in WT mice (P < 0.01; Figure 5A). Although the decreases in Kir2.1 was confirmed in ventricular cardiomyocytes from both M3Tg I/R and WT I/R mice, the expression level of Kir2.1 was much higher in M3Tg I/R mice than in WT I/R mice (P < 0.05). However, mRNA levels of KCNJ2 (encodes Kir2.1) were similar between strains in all groups (P > 0.05; Figure 5D). As expected, there were no significant differences observed in the mRNA and protein expressions of Kv4.2 (mediates Ito in cardiomyocytes) under the current experimental condition (P > 0.05; Figures 5B, E; n = 8). The mRNA levels of CACNA1C (encodes Cav1.2) was much higher in M3Tg I/R mice than in WT I/R mice (P < 0.01; Figure 5F). However, Cav1.2 (195 kDa), the major component of the L-type calcium channel, remained unchanged among four groups (P > 0.05; Figure 5C).
Figure 5.

Alterations of protein and mRNA levels of IK1 and Ito revealed by Western blot analysis and real-time RT-PCR. (A) Top, examples of Western blot bands of Kir2.1; bottom, quantification of Kir2.1 (B) Top, examples of Western blot bands of Kv4.2; bottom, quantification of Kv4.2. (C) Top, examples of Western blot bands of Cav1.2; bottom, quantification of Cav1.2. (D) Effects of transgenic M3-mAChR on expression of KCNJ2 determined by real-time RT-PCR of total RNA from control and myocardial I/R mice. (E) Effects of transgenic M3-mAChR on expression of KCND2 determined by real-time RT-PCR of total RNA from healthy and myocardial I/R mice. (F) Effects of transgenic M3-mAChR on expression of CACNA1C determined by real-time RT-PCR of total RNA from control and myocardial I/R mice. The average data were presented as mean ± SEM. **P < 0.01 versus WT or WT I/R mice.
M3-mAChR Overexpression Suppressed Upregulated miR-1 Induced by Myocardial I/R Injury
Our previous study has established KCNJ2 as a target of miR-1, and miR-1 is considered as a muscle-specific miRNA that plays an important role in ischemic arrhythmogenesis (21). Here, we detected the expression level of miR-1 by using qRT-PCR. Interestingly, the miR-1 expression was decreased in the myocardium of M3Tg compared with WT hearts (n = 5; P < 0.05; Figure 6B). Whereas the miR-1 levels were significantly increased after myocardial I/R injury in both WT and M3Tg mice, levels in M3Tg mice after myocardial I/R injury were obviously lower than those in the WT group (P < 0.01; Figure 6A).
Figure 6.
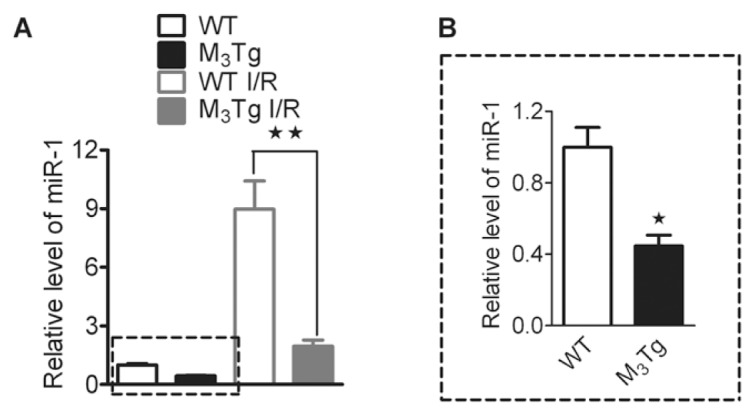
M3-mAChR overexpression on expression of miR-1 level in ventricular myocardium from control and myocardial I/R mice determined by real-time PCR. Data are expressed as mean ± SEM normalized to control. B is the magnification of part of A indicated by a dotted line. *P < 0.05. **P < 0.01 versus WT (n = 5) or WT I/R (n = 6).
DISCUSSION
Here we demonstrate that cardiac overexpression of M3-mAChR plays a critical role in the reduction of the incidence of arrhythmias and mortality in a mouse model by protecting the myocardium from ischemia, increasing the survival time after aconitine treatment and decreasing the arrhythmic score after myocardial I/R. Our patch-clamp data also demonstrate that M3-mAChR over-expression significantly shortened the APD by increasing the IK1 and consequently enhancing the membrane stability of ventricular cardiomyocytes without affecting Ito. More importantly, our molecular study clearly showed that the Kir2.1 was upregulated and miR-1 expression was downregulated in the myocardium from M3-overexpressing mice. This molecular evidence further confirmed our electrophysiological study. These findings not only help us understand the mechanisms underlying the cardioprotective effects of M3-mAChR from ischemia, but also conceptually advance our view of miRNAs that may serve as potential therapeutic and drug targets.
There are important fundamental implications of our findings. First, a large body of evidence indicates that muscarinic acetylcholine receptors, especially M3-mAChR, play critical roles in regulating the activity of many important functions of cardiovascular systems (26). Previous studies have demonstrated that M3-mAChR may possess a potential role in protecting the heart against ischemia through multiple mechanisms, including the interaction with gap junction channel connexin 43 to maintain cell–cell communication (10), activation of antiapoptotic signal molecule Bcl-2 and p38 mitogen-activated protein kinase and a decrease in Ca2+ overload induced by myocardial ischemia in ventricular cardiomyocytes (12), which has been known to improve cardiac contraction and hemodynamics by activating intracellular phosphoinositide hydrolysis via a Gq pathway (27).
Second, our study is the first to our knowledge to demonstrate that an M3Tg, specifically expressed in cardiac tissues, was successfully established and it is a useful animal model to study the M3-mAChR functions on cardiac diseases. The successful establishment of M3Tg was confirmed by its positive genotype of M3-mAChR by PCR data confirming much higher mRNA and protein expressions, and more importantly, the current density of IKM3 mediated by M3-mAChR was increased by an M3-mAChR agonist in the ventricular cardiomyocytes (Figure 1). Most importantly, the overexpression of M3-mAChR significantly reduced the incidence of arrhythmias induced by myocardial I/R, sequentially decreased the mortality of myocardial I/R and improved the survival time (Figure 2). These data significantly enhanced knowledge of the potential roles of M3-mAChR in myocardium against myocardial I/R injury, and this knowledge has been demonstrated by many researchers (28–31).
Myocardial ischemia results in a dramatic disorder of automaticity, in an increased heterogeneity of conduction and structural remodeling and in alterations of ion channel protein expression and distribution (32). These results contribute to the development of an electrical disturbance that leads to life-threatening cardiac arrhythmias. To explore the underlying ion channel mechanisms of M3-mAChR in its cardiac protection against myocardial I/R injury, the APD and some currents responsible for the characteristic differences in AP wave shapes were also examined. Our results indicate that M3-mAChR overexpression shortened the APD and increased the RMP. Effects of 4-diphenylacetoxy-N-methylpiperidine-methiodide (4-DAMP), a M3 receptor blocker, on the RMP between WT and M3Tg mice were also detected in our recent experiments. However, a significant difference of the RMP between WT and M3Tg mice was not detected in the presence of either 4.0 or 8.0 nmol/L 4-DAMP (data not shown), suggesting that 4-DAMP does not affect/reverse an increased inward rectifying K+ current (especially the upregulation of Kir2.1) nor the hyperpolarized RPM mediated by M3-mAChR overexpression. Although the activation of M3-mAChR can be blocked by 4-DAMP, but the channel protein of Kir2.1, a downstream product of M3-mAChR overexpression, was upregulated already. Blocking the receptor itself will not affect the Kir2.1 functional expression, and that is the reason why the RMP was not changed accordingly.
Consistent with our previous study (21), although individuals surviving from myocardial I/R presented a RMP depolarization, IK1 density reduction and Kir2.1 protein downregulation, interestingly, IK1 density and Kir2.1 expression were significantly higher in myocardial I/R hearts of M3Tg mice compared with WT mice (Figures 4 and 5). These results suggest that M3-mAChR overexpression plays, at least in part, an important role in cardioprotection. Under normal conditions, cardiac IK1 dominates the K+ conductance near the RMP. In accordance with this theory, our current-clamp data demonstrate that the greater the outward currents of IK1, the shorter the APD. It is also well known that IK1 stabilizes the RMP and determines the cell input resistance, which is the resistance of the membrane in the subthreshold voltage range (33). Other researches have reported that IK1 is downregulated in several other cardiac disorders such as heart failure and long QT syndrome (34,35). The downregulation of IK1 is also observed in the models of cardiac hypertrophy and in the patients with cardiomyopathies (36). This downregulation of IK1 has greater impacts on life-threatening arrhythmias due to membrane depolarization, APD prolongation, and both early and delayed after depolarizations (17).
It has been known that KCNJ2, which encodes the cardiac IK1 channels, is responsible for setting and maintaining the cardiac RMP. Although the protein level for Kir2.1 was obviously different, there were no detectable changes of KCNJ2 among the four experimental groups (Figure 5). This discrepancy between mRNA and protein levels may come from the function of microRNA, such as miR-1, which displayed opposite trends compared with the Kir2.1 protein level. As many studies have reported (21,37), microRNA can control the translation of mRNA by targeting its 3′UTR but without affecting the mRNA expression. The increased Kir2.1 protein function and expression may have resulted from the low expression of miR-1. Our data show that the mRNA level of CACNA1C was much higher in M3Tg I/R mice than in WT I/R mice. However, the protein levels of Cav1.2 were not significantly different. The reasons for this may be as follows: first, the translational level was affected by many factors posttranscription, which may be related to the regulator control system in different organs; and, second, the intensity of protein function correlates closely with the protein expression level but not its mRNA level, which was certificated with no differences of ICa,L among these groups.
Our previous study indicated that Kir2.1 was downregulated by miR-1, which was overexpressed in patients with coronary artery disease. Furthermore, repression of these genes by miR-1 contributes significantly to the arrhythmogenic potential of miR-1 during myocardial injury (21). Interestingly, our data demonstrated that miR-1 was downregulated in M3Tg mice to further lead to the increase of IK1 functional expression (Figure 6). Here, we conclude that the over-expression of M3-mAChR significantly suppressed upregulated miR-1 after myocardial I/R injury and restored the impaired KCNJ2 gene expression and function. These changes may be a novel molecular and cellular mechanism underlying the ischemic cardioprotective effects of M3-mAChR.
Supplemental Data
ACKNOWLEDGMENTS
This study was supported in part by the National Basic Research Program (973 Program) of China (2007CB512000/ 2007CB512006) and the National Natural Science Foundation of China (81072639, 30973531). We thank J Robbins for the gift of the α-MHC vector.
Footnotes
Online address: http://www.molmed.org
DISCLOSURE
The authors declare that they have no competing interests as defined by Molecular Medicine, or other interests that might be perceived to influence the results and discussion reported in this paper.
REFERENCES
- 1.Adabag AS, Luepker RV, Roger VL, Gersh BJ. Sudden cardiac death: epidemiology and risk factors. Nat Rev Cardiol. 2010;7:216–25. doi: 10.1038/nrcardio.2010.3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Zhang S. Sudden cardiac death in China. Pacing Clin Electrophysiol. 2009;32:1159–62. doi: 10.1111/j.1540-8159.2009.02458.x. [DOI] [PubMed] [Google Scholar]
- 3.Clements-Jewery H, Hearse DJ, Curtis MJ. Phase 2 ventricular arrhythmias in acute myocardial infarction: a neglected target for therapeutic antiarrhythmic drug development and for safety pharmacology evaluation. Br J Pharmacol. 2005;145:551–64. doi: 10.1038/sj.bjp.0706231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Walker MJ. Antiarrhythmic drug research. Br. J. Pharmacol. 2006;147(Suppl 1):S222–31. doi: 10.1038/sj.bjp.0706500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Shi H, Wang H, Wang Z. Identification and characterization of multiple subtypes of muscarinic acetylcholine receptors and their physiological functions in canine hearts. Mol Pharmacol. 1999;55:497–507. [PubMed] [Google Scholar]
- 6.Lamping KG, Wess J, Cui Y, Nuno DW, Faraci FM. Muscarinic (M) receptors in coronary circulation: gene-targeted mice define the role of M2 and M3 receptors in response to acetylcholine. Arterioscler Thromb Vasc Biol. 2004;24:1253–8. doi: 10.1161/01.ATV.0000130661.82773.ca. [DOI] [PubMed] [Google Scholar]
- 7.Shi H, Wang H, Li D, Nattel S, Wang Z. Differential alterations of receptor densities of three muscarinic acetylcholine receptor subtypes and current densities of the corresponding K+ channels in canine atria with atrial fibrillation induced by experimental congestive heart failure. Cell Physiol Biochem. 2004;14:31–40. doi: 10.1159/000076924. [DOI] [PubMed] [Google Scholar]
- 8.Pönicke K, Heinroth-Hoffmann I, Brodde OE. Demonstration of functional M3-muscarinic receptors in ventricular cardiomyocytes of adult rats. Br J Pharmacol. 2003;138:156–60. doi: 10.1038/sj.bjp.0704997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Shi H, Wang H, Lu Y, Yang B, Wang Z. Choline modulates cardiac membrane repolarization by activating an M3 muscarinic receptor and its coupled K+ channel. J Membr Biol. 1999;169:55–64. doi: 10.1007/pl00005901. [DOI] [PubMed] [Google Scholar]
- 10.Yue P, et al. Ischemia impairs the association between connexin 43 and M3 subtype of acetylcholine muscarinic receptor (M3-mAChR) in ventricular myocytes. Cell Physiol Biochem. 2006;17:129–36. doi: 10.1159/000092074. [DOI] [PubMed] [Google Scholar]
- 11.Wang H, Lu Y, Wang Z. Function of cardiac M3 receptors. Auton Autacoid Pharmacol. 2007;27:1–11. doi: 10.1111/j.1474-8673.2006.00381.x. [DOI] [PubMed] [Google Scholar]
- 12.Yang B, et al. Choline produces cytoprotective effects against ischemic myocardial injuries: evidence for the role of cardiac m3 subtype muscarinic acetylcholine receptors. Cell Physiol Biochem. 2005;16:163–74. doi: 10.1159/000089842. [DOI] [PubMed] [Google Scholar]
- 13.Li GR, Lau CP, Leung TK, Nattel S. Ionic current abnormalities associated with prolonged action potentials in cardiomyocytes from diseased human right ventricles. Heart Rhythm. 2004;1:460–8. doi: 10.1016/j.hrthm.2004.06.003. [DOI] [PubMed] [Google Scholar]
- 14.Diaz RJ, et al. Selective inhibition of in- ward rectifier K+ channels (Kir2.1 or Kir2.2) abolishes protection by ischemic preconditioning in rabbit ventricular cardiomyocytes. Circ Res. 2004;95:325–32. doi: 10.1161/01.RES.0000137727.34938.35. [DOI] [PubMed] [Google Scholar]
- 15.Xu Y, Zhang Q, Chiamvimonvat N. IK1 and cardiac hypoxia: after the long and short QT syndromes, what else can go wrong with the inward rectifier K+ currents? J Mol Cell Cardiol. 2007;43:15–7. doi: 10.1016/j.yjmcc.2007.04.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Dhamoon AS, Jalife J. The inward rectifier current (IK1) controls cardiac excitability and is involved in arrhythmogenesis. Heart Rhythm. 2005;2:316–24. doi: 10.1016/j.hrthm.2004.11.012. [DOI] [PubMed] [Google Scholar]
- 17.Beuckelmann DJ, Näbauer M, Erdmann E. Alterations of K+ currents in isolated human ventricular myocytes from patients with terminal heart failure. Circ Res. 1993;73:379–85. doi: 10.1161/01.res.73.2.379. [DOI] [PubMed] [Google Scholar]
- 18.Ravens U, Cerbai E. Role of potassium currents in cardiac arrhythmias. Europace. 2008;10:1133–7. doi: 10.1093/europace/eun193. [DOI] [PubMed] [Google Scholar]
- 19.Divakaran V, Mann DL. The emerging role of microRNAs in cardiac remodeling and heart failure. Circ Res. 2008;103:1072–83. doi: 10.1161/CIRCRESAHA.108.183087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Xie X, et al. Systematic discovery of regulatory motifs in human promoters and 3′UTRs by comparison of several mammals. Nature. 2005;434:338–45. doi: 10.1038/nature03441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Yang B, et al. The muscle-specific microRNA miR-1 regulates cardiac arrhythmogenic potential by targeting GJA1 and KCNJ2. Nat Med. 2007;13:486–91. doi: 10.1038/nm1569. [DOI] [PubMed] [Google Scholar]
- 22.Arimoto T, et al. Cardiac-specific overexpression of diacylglycerol kinase zeta prevents Gq protein-coupled receptor agonist-induced cardiac hypertrophy in transgenic mice. Circulation. 2006;113:60–6. doi: 10.1161/CIRCULATIONAHA.105.560771. [DOI] [PubMed] [Google Scholar]
- 23.Petzelbauer P, et al. The fibrin-derived peptide Bbeta15–42 protects the myocardium against ischemia-reperfusion injury. Nat Med. 2005;11:298–304. doi: 10.1038/nm1198. [DOI] [PubMed] [Google Scholar]
- 24.Zhang Y, et al. Resveratrol, a natural ingredient of grape skin: antiarrhythmic efficacy and ionic mechanisms. Biochem Biophys Res Commun. 2006;340:1192–9. doi: 10.1016/j.bbrc.2005.12.124. [DOI] [PubMed] [Google Scholar]
- 25.Amran MS, Hashimoto K, Homma N. Effects of sodium-calcium exchange inhibitors, KB-R7943 and SEA0400, on aconitine-induced arrhythmias in guinea pigs in vivo, in vitro, and in computer simulation studies. J Pharmacol Exp Ther. 2004;310:83–9. doi: 10.1124/jpet.104.066951. [DOI] [PubMed] [Google Scholar]
- 26.Willmy-Matthes P, Leineweber K, Wangemann T, Silber RE, Brodde OE. Existence of functional M3-muscarinic receptors in the human heart. Naunyn Schmiedebergs Arch Pharmacol. 2003;368:316–9. doi: 10.1007/s00210-003-0796-2. [DOI] [PubMed] [Google Scholar]
- 27.Shi H, Wang H, Yang B, Xu D, Wang Z. The M3 receptor-mediated K(+) current (IKM3), a G(q) protein-coupled K(+) channel. J Biol Chem. 2004;279:1774–2178. doi: 10.1074/jbc.C400100200. [DOI] [PubMed] [Google Scholar]
- 28.Wang YP, et al. M3 muscarinic acetylcholine receptor is associated with beta-catenin in ventricular myocytes during myocardial infarction in the rat. Clin Exp Pharmacol Physiol. 2009;36:995–1001. doi: 10.1111/j.1440-1681.2009.05176.x. [DOI] [PubMed] [Google Scholar]
- 29.Liu Y, et al. Role of M3 receptor in aconitine/barium-chloride-induced preconditioning against arrhythmias in rats. Naunyn Schmiedebergs Arch Pharmacol. 2009;379:511–5. doi: 10.1007/s00210-008-0376-6. [DOI] [PubMed] [Google Scholar]
- 30.Liu Y, et al. Choline produces antiarrhythmic actions in animal models by cardiac M3 receptors: improvement of intracellular Ca2+ handling as a common mechanism. Can J Physiol Pharmacol. 2008;86:860–5. doi: 10.1139/Y08-094. [DOI] [PubMed] [Google Scholar]
- 31.Zhao WM, et al. The antiarrhythmic effect and possible ionic mechanisms of pilocarpine on animal models. J Cardiovasc Pharmacol Ther. 2009;14:242–7. doi: 10.1177/1074248409339308. [DOI] [PubMed] [Google Scholar]
- 32.de Bakker JM, van Rijen HV. Electrocardiographic manifestation of anatomical substrates underlying post-myocardial infarction tachycardias. J Electrocardiol. 2007;40:21–5. doi: 10.1016/j.jelectrocard.2007.05.018. [DOI] [PubMed] [Google Scholar]
- 33.Panama BK, McLerie M, Lopatin AN. Heterogeneity of IK1 in the mouse heart. Am J Physiol Heart Circ Physiol. 2007;293:3558–67. doi: 10.1152/ajpheart.00419.2007. [DOI] [PubMed] [Google Scholar]
- 34.Fauconnier J, Lacampagne A, Rauzier JM, Vassort G, Richard S. Ca2+-dependent reduction of IK1 in rat ventricular cells: a novel paradigm for arrhythmia in heart failure? Cardiovasc Res. 2005;68:204–12. doi: 10.1016/j.cardiores.2005.05.024. [DOI] [PubMed] [Google Scholar]
- 35.Domenighetti AA, Boixel C, Cefai D, Abriel H, Pedrazzini T. Chronic angiotensin II stimulation in the heart produces an acquired long QT syndrome associated with IK1 potassium current downregulation. J Mol Cell Cardiol. 2007;42:63–70. doi: 10.1016/j.yjmcc.2006.09.019. [DOI] [PubMed] [Google Scholar]
- 36.Tomaselli GF, Marbán E. Electrophysiological remodeling in hypertrophy and heart failure. Cardiovasc Res. 1999;42:270–83. doi: 10.1016/s0008-6363(99)00017-6. [DOI] [PubMed] [Google Scholar]
- 37.Chen JF, et al. The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation. Nat Genet. 2006;38:228–33. doi: 10.1038/ng1725. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.