

Abstract
Dysregulated release of neutrophil azurophilic granules causes increased tissue damage and amplified inflammation during autoimmune disease. Antineutrophil cytoplasmic antibodies (ANCAs) are implicated in the pathogenesis of small vessel vasculitis and promote adhesion and exocytosis in neutrophils. ANCAs activate specific signal transduction pathways in neutrophils that have the potential to be modulated therapeutically to prevent neutrophil activation by ANCAs. We have investigated a role for diacylglycerol kinase (DGK) and its downstream product phosphatidic acid (PA) in ANCA-induced neutrophil exocytosis. Neutrophils incubated with the DGK inhibitor R59022, before treatment with ANCAs, exhibited a reduced capacity to release their azurophilic granules, demonstrated by a component release assay and flow cytometry. PA restored azurophilic granule release in DGK-inhibited neutrophils. Confocal microscopy revealed that R59022 did not inhibit translocation of granules, indicating a role for DGK during the process of granule fusion at the plasma membrane. In investigating possible mechanisms by which PA promotes neutrophil exocytosis, we demonstrated that exocytosis can only be restored in R59022-treated cells through simultaneous modulation of membrane fusion and increasing cytosolic calcium. PA and its associated pathways may represent viable drug targets to reduce tissue injury associated with ANCA-associated vasculitic diseases and other neutrophilic inflammatory disorders.
INTRODUCTION
Human neutrophils contain granules that, when released into the extracellular environment, can promote cellular adhesion, facilitate transmigration into inflamed tissue and provide the cell with essential antimicrobial capability. Because of the high concentration of proteolytic enzymes contained within neutrophil granules, their release from the neutrophil is a tightly controlled and regulated process. For a granule to be successfully released from the cell, key events must occur. First, powered by actin polarization, granules must translocate from the cytosol to the plasma membrane, where they tether and dock at predetermined areas. After this, the granule must fuse and integrate with the plasma membrane to make the granular contents accessible to the outer environment. A number of intracellular signaling molecules are used by the cell to promote each step of exocytosis (1–3). Exacerbation of inflammation is associated with a number of autoimmune diseases where increased release of proteolytic enzymes enhances tissue damage (4,5). Delineating the mechanisms and pathways associated with this aberrant process of exocytosis may reveal a number of targets to control and reduce it.
Antineutrophil cytoplasmic antibodies (ANCAs) are implicated in the pathogenesis of small vessel vasculitides such as granulomatosis with polyangiitis (Wegener’s), microscopic polyangiitis and Churg-Strauss syndrome (6). Antibodies with specificities to either proteinase 3 (PR3) or myeloperoxidase (MPO) are believed to contribute to development of acute disease by activating neutrophils within the small vessels of the lung, kidney or other organs. Neutrophil exocytosis of the azurophilic granules is likely to play a role in promoting endothelial cell damage in the blood vessel, with serine proteases and MPO released from activated neutrophils able to induce damage to both endothelial cells and the basement membrane (7–9).
ANCA IgG promotes distinct signal transduction pathways compared with neutrophil activation by either immune complexes or chemoattractants, and the pathways are independent of phospholipase D involvement (10). During ANCA-induced superoxide production, ANCA IgG F(ab′)2 (fragment antigen-binding) binds to its antigen on the surface of primed cells, resulting in the activation of the heterotrimeric G protein Gi, thereby stimulating phosphatidylinositol 3-kinase (PI3K) type 1γ (PI3Kγ), which activates protein kinase B (10,11). Binding of the Fc fragment of ANCA IgG to either FcγRIIa or FcγRIIIb results in the autophosphorylation of the tyrosine kinases syk and src and the adaptor cbl (12). We have also previously demonstrated the importance of phosphatidic acid (PA) production in the promotion of ANCA-induced neutrophil adhesion in vitro (13). PA production was shown to depend on the activation of the enzyme diacylglycerol kinase (DGK), to phosphorylate the lipid diacylglycerol (DAG). ANCA stimulation results specifically in the phosphorylation of both saturated and monounsaturated forms of DAG (13). These forms of PA are believed to act as signaling secondary messengers compared with polyunsaturated forms of the same molecules that are considered nonsignaling, transient metabolites (14,15).
The azurophilic granules contain the highest concentrations of both MPO and serine proteases compared with other granules and are likely to be released in a highly inflammatory setting (16). We investigated the release of these granules after ANCA IgG activation. Here, we study the signal transduction pathways activated by ANCA IgG that promote the exocytosis of azurophilic granules in vitro, specifically focusing on the activation of DGK through the production of PA. We report that ANCA-induced release of MPO depends on the production of PA, through the activation of DGK. We also demonstrate a dual mechanism for the actions of PA through both the generation of intracellular calcium and the ability to modulate the plasma membrane topography. The work highlights an intriguing pathway that may be activated by ANCAs and that can be further studied and targeted by rational drug design.
MATERIALS AND METHODS
IgG Isolation
Total IgG was isolated from anonymized plasmapheresis samples obtained from ANCA-positive patients. All patients fulfilled Chapel Hill definitions and had active renal disease. IgG in serum was isolated on a protein G-Sepharose column (GE Healthcare, Buckinghamshire, UK). Both PR3- and MPO-specific ANCA IgGs were isolated and included in each set of experiments. We found no statistical difference in the ability of either type of ANCA to promote neutrophil activation in any of the assays performed in this study. The protocol was approved by the local ethics committee.
Generation of F(ab′)2 Fragments
Both ANCA and normal IgG preparations were pepsin digested to generate F(ab′)2 fragments. Preparations of anti-body (0.5 mg) were dialyzed overnight in acetate buffer (0.1 mol/L sodium acetate, 0.1 mol/L acetic acid, pH 4) and then incubated with pepsin (20 μg/mL) in acetate buffer for 24 h. The preparation was then dialyzed overnight in 2× phosphate-buffered saline. Fc fragments were removed through incubation with immobilized protein G beads (Thermo Scientific). Concentration of F(ab′)2 was determined by spectrophotometry and generation of F(ab′)2 fragments confirmed on an sodium dodecyl sulfate–polyacrylamide gel electrophoresis gel.
Neutrophil Isolation
Blood from healthy donors was obtained by venipuncture into acid citrate dextrose, and neutrophils were isolated by centrifugation over a Percoll discontinuous gradient (GE Healthcare) as described previously (17). Informed consent was received from all donors, and the local ethics committee approved the protocol.
Exocytosis Assay
Isolated neutrophils were diluted to 2.5 × 106/mL in Hanks buffered salt solution with 10 mmol/L 4-(2-hydroxyethyl)-1-piperazine ethanesulfonic acid (HEPES) (pH 7.4) and primed for 15 min with tumor necrosis factor (TNF)-α (2 ng/mL) (NIBSC, Potters Bar, UK) in the presence of cytochalasin B (10 μmol/L) (Sigma) at 37°C. Neutrophils (2.5 × 105) were added to wells of a round-bottom 96-well plate and stimulated with 200 μg/mL ANCA IgG, normal IgG or f-Met-Leu-Phe (fMLF) (1 μmol/L) (Sigma) as a positive control for 15 min at 37°C. After stimulation, cells were spun at 400g for 5 min and supernatants were removed. The activity of MPO within each sample was investigated using the o-phenylenediamine dihydrochloride (OPD) substrate (Sigma) (according to the manufacturer’s instructions). The reaction was stopped with 100% glacial acetic acid and read at 450 nm (Multiskan Thermo-Fisher, Waltham, MA, USA). Optical density readings from each sample were normalized as a percentage of the unstimulated control. Anti-MPO and anti-PR3 IgG had no effect on the enzymatic activity of MPO. Exocytosis was also measured after neutrophil adhesion. Neutrophils (2.5 × 106/mL) were primed with TNF-α (2 ng/mL) in the absence of cytochalasin B for 15 min and stimulated in 96-well flat-bottom plates previously coated with fibrinogen (25 μg/mL) as described previously (13). After 2 h of treatment, supernatants were removed and spun down to remove any unbound neutrophils and MPO release was measured as above. After the assay, neutrophils were found to be over 90% viable by try-pan blue (Sigma) staining.
Inhibition of DGK and Phospholipase D Activity
Neutrophils, being terminally differentiated cells, are not amenable to many molecular and genetic techniques that may be used in other cell types to investigate signal transduction pathways. Therefore, we used specific chemical inhibitors to study the role of secondary messengers in ANCA-induced activation. All treatments had no significant effect on neutrophil viability analyzed by trypan blue exclusion and forward/side scatter properties using flow cytometry. For the study of DGK, we used the specific inhibitor R59022, which directly targets Ca2+-sensitive isoforms of DGK selectively (3,18,19). For the study of phospholipase D (PLD), we used the inhibitor VU0155069, which at concentrations >1 μmol/L, inhibits the two mammalian isoforms of PLD (PLD1 and PLD2). Concentrations of inhibitors used were selected by known selectivity of isoforms in cell systems and had no effect on neutrophil viability in vitro assays and no effect on TNF-α priming. As a result, neutrophils were incubated with the DGK-selective inhibitor R59022 (18 μmol/L) (Calbiochem, La Jolla, CA, USA) or the PLD-selective inhibitor VU0155069 (1 μmol/L) for 10 min (Cayman Chemicals, Ann Arbor, MI, USA) before priming. All incubations were carried out before neutrophil priming at 37°C. DMSO (Sigma) used as a vehicle control, at the highest concentration, did not have a significant effect on neutrophil activation.
Flow Cytometry
Neutrophils were analyzed for the surface expression of CD63 using a monoclonal fluorescein isothiocyanate (FITC)-conjugated antibody at 2 μg/mL (AbD Serotec, Oxford, UK) and an IgG isotype control (BD Pharmingen, Oxford, UK). Cells were incubated with antibody for 15 min at room temperature before being washed and fixed with 2% paraformaldehyde. Samples were acquired on a FACScalibur flow cytometer using Cell Quest software (Becton Dickinson, Oxford, UK) with 1 × 104 cells acquired per tube. List mode data were analyzed using WinDMI (2.9) software.
Addition of Short-Chain Phosphatidate and Lysophosphatidylcholine
Short-chain lipid phosphatidate (Di8:0) (PA; Sigma), 1,2-dioctanoyl-sn-glycerol (Di8:0) (DAG, Cayman Chemicals, Tallinn, Estonia) and lysophosphatidylcholine (LPC) (Sigma) was reconstituted in chloroform:methanol (2:1), vacuumdried, resuspended in phosphate-buffered saline and sonicated. PA, DAG (final concentration 12.5 μmol/L) and LPC (250 nmol/L) were added to cells after priming and before neutrophil stimulation at 37°C. The working concentration of both PA and LPC was added at the lowest concentration that did not cause significant exocytosis in unstimulated cells during titration experiments. The concentration of PA is within the range of what is found physiologically in stimulated neutrophils (20). All treatments had no significant effect on neutrophil viability analyzed by trypan blue exclusion and forward/side scatter properties using flow cytometry.
Confocal Microscopy
Neutrophils were treated as above, and the cells were then fixed with 2% paraformaldehyde before the addition of wheat-germ agglutinin conjugated to alexa fluor 594 (5 μg/mL) to label plasma membranes (LIVE plasma membrane stain; Invitrogen, Paisley, UK). Neutrophils were then resuspended in permeabilization buffer (e-Bioscience, San Diego, CA, USA) and labeled with anti-CD63 monoclonal FITC-conjugated antibody L (AbD Serotec) or IgG isotype (BD Pharmingen) both at 2 μg/mL or incubated with phalloidin-FITC (10 μmol/L) (Sigma). Staining was carried out for 20 min at room temperature. The stained cells were spun onto glass slides, mounted in SlowFade® Gold antifade buffer (Invitrogen) and viewed using a Zeiss LSM 510 confocal scanning microscope equipped with a c-apochromat 100×/1.2 oil immersion objective (Carl Zeiss, Jena, Germany). All images were processed and analyzed using LSM 510 software (version 2.3; Carl Zeiss). Isotype- and concentration-matched control antibodies yielded consistently negative results.
[Ca2+] Measurement by Spectrofluorimetry
Neutrophils were resuspended at 2 × 106 cells/mL in 1.4 mmol/L Ca2+ Hanks balanced salt solution + 20 mmol/L HEPES buffer + 7.5% NaHCO3 and loaded with Indo-1 AM ester (Invitrogen) at 1 μmol/L (21). To confirm that the cells were resting and produced a stable baseline, the ratio of the fluorescent intensity at the two emission wavelengths was observed for 100 s, and ANCAs (200 μg/mL) were added through a light occlusive stopper in the top of the fluorimeter. After addition of the stimulus, the trace was observed until a stable plateau response was achieved. A calibration file was generated to allow calculation of [Ca2+] from the ratio of fluorescence intensity, as described previously (22). To assess the relative contributions of intracellular-derived Ca2+ and the influx of Ca2+ from the extracellular space, investigations were performed under Ca2+ free conditions. Neutrophils were transferred into Ca2+ free Hanks balanced salt solution containing HEPES buffer (HBH). Any remaining Ca2+ was removed by the addition of ethyleneglycoltetraacetic acid (EGTA) (0.4 mmol/L) immediately before stimulation, thus minimizing the time that the cells spent in Ca2+ free medium. ANCAs (200 μg/mL) were then added, and the trace was observed until a stable plateau response was observed. A source of Ca2+ (CaCl2) (1.4 mmol/L) was then added to the cell suspension to determine the influence of system operating channels and enable the rate and magnitude of Ca2+ influx to be assessed.
Statistical Analysis
Where required, data were analyzed by one-way repeated-measures analysis of variance (ANOVA) with a Bonferroni posttest or paired Student t test using Graphpad Prism software (Graphpad, La Jolla, CA, USA).
RESULTS
Neutrophils Release MPO to Intact ANCA IgG, Dependent on Actin Cytoskeleton Modulation
We first investigated the ability of neutrophils to release MPO in vitro (Figure 1). Because neutrophils will not readily exocytose their azurophilic granules in suspension, many studies facilitate exocytosis through modulation of the actin cytoskeleton by cytochalasin. Neutrophils are unlikely to encounter this fungal metabolite physiologically, yet in an inflammatory environment, they may also release the azurophilic granules through modulation of the actin cytoskeleton by increased adhesion to the activated endothelium (23). We compared the ability of cells stimulated with ANCAs or fMLF in the presence of cytochalasin B in suspension (Figure 1A) with neutrophils allowed to adhere to fibrinogen (Figure 1B). In both assays, ANCA IgG and fMLF treatment induced similar significant release of MPO above that seen upon treatment with control IgG. The data suggest that neutrophils release azurophilic granules dependent on changes to the cellular cytoskeleton, which physiologically would be induced upon neutrophil adhesion, but can be mimicked in a similar fashion in vitro by cytochalasin B. This may be explained by the observations that neutrophil adhesion promoted by either cytochalasin or through ligation of β2 integrin directly modulate the actin-rich cortical cytoskeleton altering cell rigidity (24–27). By correlating the amount of MPO released after stimulation with total MPO measured in cell lysates (Figure 1C), we found that, on average, ANCA IgG induced the release of 44.6% (± 8.9%) of the total cellular MPO compared with 68.1% (± 11.46%) of MPO released when neutrophils were treated with fMLF. To investigate whether ligation of both the Fc and F(ab′)2 portions are required to promote neutrophil exocytosis, we created F(ab′)2 and Fc fragments of ANCA IgG by pepsin digestion and compared neutrophil responses to these fragments compared with responses to complete ANCA IgG (Figure 1D). Whole ANCA IgG-activated neutrophils exhibited significant MPO release above that seen in unstimulated cells, whereas treatment of cells with either F(ab′)2 or Fc fragments of ANCA antibody alone failed to cause any increase in the release of MPO. Treating neutrophils with both F(ab′)2 and Fc fragments concurrently also failed to induce a significant release of MPO from the cell, indicating that the ANCA IgG molecule must be intact to induce exocytosis in neutrophils. F(ab′)2 and Fc portions added back to whole ANCA preparations could also inhibit neutrophil exocytosis (data not shown), suggesting that blocking antigen availability or ligation of low-affinity FcγR is sufficient to inhibit exocytosis by whole ANCA IgG. The data demonstrate that colocalization of the signal transduction pathways promoted by both the F(ab′)2 and Fc portions of the ANCA IgG is required to promote neutrophil release of the azurophilic granules.
Figure 1.

Ability of whole ANCA and ANCA F(ab′)2 to induce azurophilic granule release in primed neutrophils. Neutrophils (2.5 × 105) were primed with TNF-α (2 ng/mL) in the presence of cytochalasin B (10 μmol/L) for 15 min. Neutrophils were then treated with ANCA IgG, normal control IgG (200 μg/mL) or the positive control fMLF (1 μmol/L) for 15 min (A). Neutrophils were also treated in the absence of cytochalasin B (B) and treated as above but allowed to adhere onto a fibrinogen-coated (50 μg/mL) plate for 2 h. After treatment, supernatants were collected and analyzed for MPO using a specific substrate. Data have been normalized as a percentage of the unstimulated (unstim) control cells. For neutrophils primed with TNF-α in the presence of cytochalasin B, the data are also expressed as a percentage of total MPO from lysates (C). Primed neutrophils (2.5 × 105), in the presence of cytochalasin B, were also treated separately with ANCA IgG (200 μg/mL), F(ab′)2 fragments, Fc fragments or preparations of both (300 μg/mL) from the same antibody preparation for 15 min (D). Data were normalized as a percentage of the unstimulated control cells. Data are a mean of five different neutrophil donor responses using a panel of two normal IgG preparations and three ANCA preparations (one MPO-ANCA, two PR3-ANCA). ***P < 0.001, **P < 0.01, *P < 0.05 by one-way ANOVA with posttest. Tested data samples were compared with normal IgG-treated (A, B and C) or ANCA IgG-treated (D) samples.
ANCA-Induced Azurophilic Granule Exocytosis Is Driven by DGK Activation and Phosphatidic Acid Production
To investigate the role of DGK activation in ANCA-induced azurophilic granule release, TNF-α primed neutrophils were stimulated in the presence or absence of R59022, a potent DGK inhibitor, and supernatants were analyzed for the activity of MPO (Figure 2A). Having previously demonstrated that DGK activation is pivotal to ANCA-induced neutrophil adhesion (13), we facilitated exocytosis in these assays with cytochalasin B. In the presence of R59022, neutrophils released significantly lower levels of MPO than control noninhibited cells. To rule out the possible impact of PLD-driven pathways, neutrophils were also treated in the presence of VU0155069, a PLD-selective inhibitor. VU0155069 had no effect on the release of MPO after ANCA treatment, but did significantly reduce levels of MPO released as a result of fMLF treatment. To confirm the importance of DGK activation on azurophilic granule exocytosis from neutrophils, we investigated neutrophil surface expression of the lysosomal marker CD63 (Figure 2B). After stimulation of neutrophils with ANCA IgG, we observed a significant increase in the surface expression of CD63 above that seen in normal IgG-treated cells. In the presence of the R59022 inhibitor, ANCA IgG stimulation caused no increase in CD63. To determine whether the action of DGK was independent of the action of cytochalasin B, we aimed to measure release of the azurophilic granules in the absence of cytochalasin B and adhesion. To achieve this aim, neutrophils were primed with TNF-α along with fMLF (1 μmol/L) before treatment with ANCAs in the presence or absence of the DGK inhibitor R59022 (Figure 2C). Neutrophils treated with ANCA IgG released significantly higher levels of MPO than neutrophils treated with normal IgG. Neutrophils treated in the presence of R59022 exhibited a significantly reduced ability to release MPO than noninhibited cells. These results suggest that R59022 inhibits ANCA-induced exocytosis, independently of the actions of cytochalasin B. We next investigated the role of PA as a downstream secondary messenger of the activation of DGK in ANCA IgG-stimulated neutrophils through the addition of a short-chain lipid analog of PA (Di8:0) to neutrophils in the presence or absence of the DGK inhibitor R59022 (Figure 2D). Although PA (DiC8:0) is a low abundance lipid in mammalian cells, lipids with acyl-chains of no more than eight carbons are soluble in aqueous solution, cell permeable and mimic the effects of their endogenously synthesized counterparts when added to cells (28). The addition of 12.5 μmol/L PA at the time of ANCA IgG stimulation restored the release of MPO significantly in DGK-inhibited cells. Phosphatidic acid alone had no significant effect on neutrophil exocytosis in unstimulated, normal IgG or uninhibited ANCA IgG-treated cells. Addition of the metabolite DAG also failed to restore exocytosis in ANCA-stimulated cells, indicating that PA is acting directly as a secondary messenger. The data confirm that DGK, through the production of PA, promotes exocytosis of the azurophilic granules when neutrophils are stimulated with ANCA IgG.
Figure 2.

The effect of the DGK inhibitor R59022 on exocytosis from ANCA IgG-stimulated primed neutrophils. TNF-α primed neutrophils in the presence of cytochalasin B (2.5 × 105) were treated with ANCA IgG, normal control IgG (200 μg/mL) or the positive control fMLF (1 μmol/L) for 15 min. Paired samples of cells were also treated with R59022 (18 μmol/L) or VU0155069 (1 μmol/L) for 10 min before priming. Supernatants were collected and analyzed for MPO using a specific substrate (A). Primed neutrophils in the presence of cytochalasin B (2.5 × 105) were treated with ANCA IgG as above in the presence or absence of R59022; CD63 surface expression was then analyzed by flow cytometry (B). Neutrophils were also primed with TNF-α for 15 min in the presence of fMLF (1 μmol/L) as a substitute for cytochalasin B, treated in suspension as above and measured for MPO release (C). Primed neutrophils in the presence of cytochalasin B (2.5 × 105) were treated with ANCA IgG as above in the presence or absence of PA or DAG (both at 12.5 μmol/L). Paired samples of cells were also treated with R59022 (18 μmol/L) for 10 min before priming. Supernatants were collected and analyzed for the activity of MPO (D). Data are the mean of six different neutrophil donor responses using a panel of two normal IgG preparations and three ANCA preparations (one MPO-ANCA, two PR3-ANCA). All data represent values that have been normalized as a percentage of the unstimulated (unstim) uninhibited control cells with optical density (OD) (A, C and D) or geometric mean (B) as the primary measurement. ***P < 0.001, **P < 0.01, *P < 0.05 by one-way ANOVA with posttest.
DGK Does Not Affect Azurophil Granule Movement
We next investigated whether DGK promotes the movement of azurophilic granules (Figure 3). To detect granule movement, neutrophils were labeled with the plasma membrane stain wheat-germ agglutinin and, after permeabilization, were labeled with anti-CD63 or the F-actin specific toxin phalloidin. Control unstimulated cells exhibited a diffuse cytoplasmic pattern of anti-CD63 staining and displayed a prominent cortical F-actin ring near the plasma membrane. Control cells in the presence of the inhibitor R59022 maintained this resting phenotype. After treatment with ANCA IgG, neutrophils exhibited a flattening and protrusion of the plasma membrane, usually to one side of the cell, with a distinct colocalization of both CD63 and F-actin to these areas. Cells treated in the presence of the DGK inhibitor R59022 also displayed this activated phenotype, indicating that R59022 had no effect, either on the movement of granules or the formation of F-actin that allows this movement to occur, and further suggesting that DGK activity may be focused downstream of the movement of granules from the cytosol to areas of actin polymerization.
Figure 3.

The localization of intracellular CD63 and F-actin in stimulated neutrophils treated in the presence of R59022. Primed neutrophils (2.5 × 105) were treated with ANCA IgG (200 μg/mL) for 15 min. Paired samples of cells were also treated with R59022 (18 μmol/L) for 10 min before priming. Cells were fixed and labeled with the plasma membrane stain wheat-germ agglutinin (alexa-fluor 594 conjugated) and then permeabilized and labeled with anti-CD63 (FITC conjugated) or phalloidin (FITC conjugated), spun onto microscope slides and visualized by confocal microscopy at 100× magnification, with an additional 3.5× digital zoom. One representative cell is shown for each treatment (with over 75% of cells exhibiting this phenotype) in blocks of three, comprising FITC labeling in green, plasma membrane labeling in red and merged images with areas of colocalization in yellow on the image. All merged images were analyzed for distribution of fluorescence intensity of anti-CD63-FITC or phalloidin-FITC staining with the plasma membrane wheat-germ agglutinin alexa-fluor 594 conjugate stain within the same cell. An arrow was placed (cross-sectional) over the cell and histograms generated to distinguish fluorescent intensities of both FITC and alexa-fluor staining along the line of the arrow. Red histograms represent alexa-fluor membrane staining, whereas the green histograms represent FITC staining. The y axis represents fluorescence intensity, whereas the x axis represents distance across the cell (μm). All neutrophils in the image are from the same donor and the same individual experiment. The figure is representative of three individual experiments.
Membrane Fusion Modulation Cannot Restore Exocytosis in DGK-Inhibited Cells
Phosphatidic acid, a cone-shaped lipid, has been reported to modulate curvature of the plasma membrane, thereby reducing the energy requirements for fusion between the granule and plasma membrane (29). To determine whether PA derived from DGK activation was providing a structural role in the plasma membrane, we investigated the ability of LPC, an inverted cone-shape lipid, to restore exocytosis in R59022 inhibited cells. LPC concentrated in the outer plasma membrane provides a positive curvature of the membrane from the outside, thereby recreating the physical modulation of the membrane provided by PA (30). LPC, at a concentration that did not induce exocytosis in control resting neutrophils (250 nmol/L), was unable to restore exocytosis in R59022 inhibited cells (Figure 4), indicating that DGK-driven PA production does not solely promote exocytosis through the acids’ ability to modulate membrane fusion.
Figure 4.
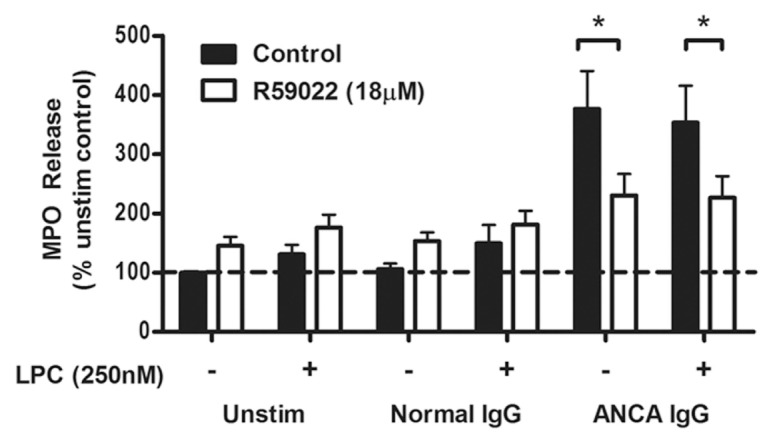
Assessment of LPC to restore ANCA induced exocytosis in DGK-inhibited neutrophils. Primed neutrophils (2.5 × 105) in the presence of cytochalasin B were treated with ANCA IgG (200 μg/mL) or IgG (200 μg/mL) from healthy control donors for 15 min in the presence or absence of LPC (250 nmol/L). Paired samples of cells were treated with R59022 (18 μmol/L) for 10 min before priming. Supernatants were collected and analyzed for the activity of MPO using a specific substrate. Data have been normalized as a percentage of the unstimulated (unstim) uninhibited control cells. Data are the mean of six different neutrophil donor responses using a panel of two normal IgG preparations and three ANCA preparations (one MPO-ANCA, two PR3-ANCA). *P < 0.05 by one-way ANOVA with posttest.
DGK, through the Production of PA, Promotes the Release of Intracellular Calcium by ANCAs
Increases in intracellular calcium are fundamental to both the movement and docking of primary granules, along with their subsequent fusion to the plasma membrane (31). To investigate the role of DGK activation in the generation of intra-cellular calcium levels, neutrophils were labeled with the calcium indicator dye INDO-1 and treated with ANCA IgG in the absence or presence of the DGK inhibitor R59022 (Figures 5A, B). Neutrophils treated with ANCA exhibited an increase in intracellular calcium above baseline that was significantly diminished in the presence of R59022. Control cells treated with ANCA IgG in the presence of PA exhibited no significant difference in release of intracellular calcium compared with cells treated with ANCA alone, whereas R59022-inhibited cells exhibited a significantly increased ability to release calcium when treated in the presence of PA. To investigate the source of Ca2+, which R59022 inhibits, we examined the effect of ANCA IgG treatment and R59022 inhibition on the generation of intracellular Ca2+ and the influx of Ca2+ across the plasma membrane (Figures 5C, D). Treatment of neutrophils in Ca2+ free HBH allowed us to measure the release of Ca2+ from intracellular stores. In the presence of R59022, this intracellular release was significantly inhibited. Adding back Ca2+ into the cell suspension enabled us to determine the effect of R59022 store-operated calcium channels. Both neutrophils stimulated in the presence or absence of R59022 exhibited similar levels of Ca2+ influx. The data suggest R59022 directly inhibits the release of Ca2+ from intracellular stores. We next investigated the ability of Ca2+ to restore exocytosis in R59022-inhibited cells. Addition of ionomycin, at a concentration that did not induce exocytosis in unstimulated cells (5 nmol/L), failed to restore exocytosis, measured by the release of MPO, in R59022-treated cells (Figure 5E). This concentration of ionomycin was sufficient to induce the intracellular release of calcium in R59022-inhibited cells, similar to calcium released in control cells treated with ANCA IgG (Figure 5F). The results indicate that DGK, through the production of PA, drives calcium generation in ANCA IgG-stimulated cells, yet this action is not solely responsible for the exocytosis of the azurophilic granules.
Figure 5.

The effect of DGK inhibition on ANCA-induced intracellular calcium release. Neutrophils (2 × 106) were preincubated with INDO-1 (1 μmol/L) dye for 30 min before priming. Paired samples of cells were also treated with R59022 (18 μmol/L) for 10 min before priming. The ratio of the fluorescent intensity was observed for 100 s before treating neutrophils with ANCA (200 μg/mL) and recording changes in fluorescent intensity for 500 s. Paired readings were recorded for neutrophils treated in the presence of PA (12.5 μmol/L). Mean data (A) are shown as the increase in calcium with the highest value of calcium subtracted from the baseline levels. A representative time course assay is shown with fluorescent intensity ratio converted to calcium concentrations (B). To assess the relative contributions of the intracellular Ca2+ store and the influx of extracellular Ca2+ in response to ANCA IgG stimulation, neutrophils were stimulated in Ca2+ free medium containing EGTA, and then after a response plateau, Ca2+ was added to back to the cell suspension. A paired neutrophil sample also received R59022 (18 μmol/L) before priming. Mean data for both the release of intracellular Ca2+ and influx of extracellular Ca2+ are shown (C). A representative time course assay is also shown (D). The ability of ionomycin to restore exocytosis in R59022-inhibited cells was also assessed (E). Primed neutrophils (2.5 × 105) were treated with ANCA IgG (200 μg/mL) or control normal IgG (200 μg/mL) for 15 min in the presence or absence of ionomycin (5 nmol/L). Paired samples of cells were treated with R59022 (18 μmol/L) for 10 min before priming. Supernatants were collected and analyzed for the activity of MPO using a specific substrate. A representative time course assay is shown for the ability of ionomycin (5 nmol/L) to restore calcium levels in R59022-inhibited cells (F). Data have been normalized as a percentage of the unstimulated (unstim), uninhibited control cells. All graphs show the mean of four different neutrophil donor responses using a panel of two normal IgG preparations and two ANCA preparations (one MPO-ANCA, one PR3-ANCA). *P < 0.05, **P < 0.01, ***P < 0.001 by a paired Student t test.
Actions of LPC and Ionomycin Combine to Restore Exocytosis in R59022-Inhibited Cells
To investigate the possibility that PA may have dual roles in the process of ANCA IgG-induced exocytosis, we tested whether adding back calcium and modulating the neutrophil’s membrane fusion together may have a cumulative effect to restore exocytosis, similar to the actions of PA (Figure 6). Both LPC and ionomycin, when added alone in the presence of ANCA IgG treatment, showed no ability to restore exocytosis in the R59022-treated cells, whereas treating R59022-inhibited cells with ANCA in the presence of both LPC and ionomycin resulted in a significant increase in the release of MPO compared with R59022-inhibited cells alone. LPC alone demonstrated no ability to induce the release of intracellular Ca2+ (data not shown). These results suggest that the ability to modulate membrane fusion and promote the release of intracellular calcium are key roles of PA in the promotion of ANCA-induced exocytosis and that these actions occur in unison to achieve the release of inflammatory enzymes from the activated neutrophil.
Figure 6.
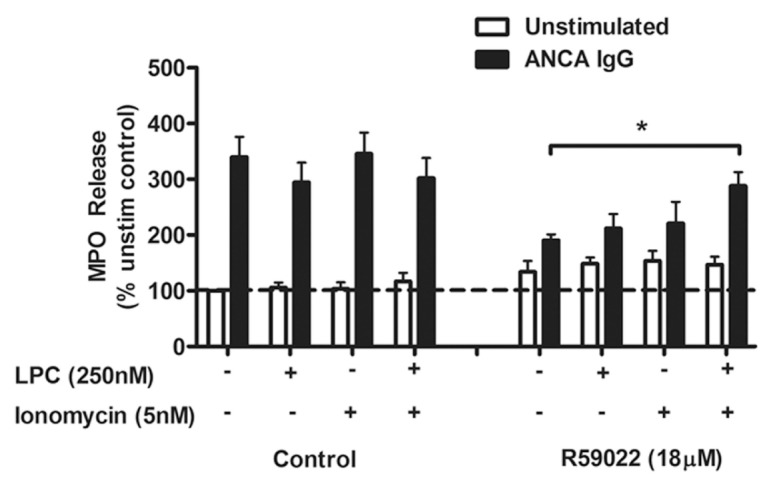
Restoration of ANCA-induced exocytosis with LPC and ionomycin. Primed neutrophils in the presence of cytochalasin B (2.5 × 105) were treated with ANCA IgG (200 μg/mL) or control IgG (200 μg/mL) for 15 min in the presence of LPC (250 nmol/L) and ionomycin (5 nmol/L) alone or together in the same treatment sample. Paired samples of cells were treated with R59022 (18 μmol/L) for 10 min before priming. Supernatants were collected and analyzed for the activity of MPO using a specific substrate. Data have been normalized as a percentage of the unstimulated (unstim), uninhibited control cells. Data are the mean of seven different neutrophil donor responses using a panel of two normal IgG preparations and two ANCA preparations (one MPO-ANCA, one PR3-ANCA). *P < 0.05 by one-way ANOVA with posttest.
DISCUSSION
Pathophysiological mediators of neutrophil activation, including the vasculitis-associated autoantibody ANCA, can use distinct signaling pathways to induce exocytosis of the cell’s azurophilic granules. We previously reported that ANCA IgG induces neutrophil adhesion through a pathway involving DGK-catalyzed monounsaturated or saturated PA formation, although this pathway is not used for ANCA-mediated superoxide release (13). In this study, we now report that ANCA-induced DGK-mediated formation of PA promotes exocytosis of the azurophilic granules by dual effects on calcium and membrane fusion.
Although the complete signaling pathways promoting ANCA-induced neutrophil exocytosis are yet to be fully defined, it is clear from this report that ligation of both the Fc and F(ab′)2 portions of ANCA IgG are required to promote the release of azurophilic granules. The work also confirms earlier studies examining superoxide release (32) where both fragments, through ligation, were shown to promote distinct signaling pathways. It is possible that ligation of receptors by the intact IgG molecule brings key receptors and signaling molecules together in the plasma membrane to promote adequate signal transduction. This result is feasible considering that the ANCA antigens have been reported to sit in distinct lipid rafts near both the β2 integrin complex and the low-affinity IgG receptor FcγRIIIb (33).
A degree of actin disruption induced by cytochalasin B has been shown, in a number of previous studies, to be required for chemoattractant-induced azurophilic granule release (3,25), bringing into question the ability of neutrophils to release their azurophilic granules physiologically. In this study, we were able to demonstrate that primed neutrophils stimulated with either autoantibody or fMLF, and left to adhere onto fibrinogen, released significant levels of MPO, suggesting that actin cytoskeleton modulation induced by adhesion and transmigration is sufficient to promote the release of the azurophilic granules. The mechanisms of neutrophil adhesion and exocytosis may well be entwined, since CD63 localizes with the β2 integrin CD11/CD18 in distinct lipid rafts of the plasma membrane and signal transduction downstream of CD63 promotes the distinct activation of the CD11/CD18 to promote increased neutrophil adhesion (34,35).
Exocytosis of granules is a complex and ordered process, beginning with the movement of granules from the cytosol to the plasma membrane, recognition and docking of the granule and complete fusion between vehicle and target membrane resulting in the release of granule constituents into the extracellular environment. Using confocal microscopy, we observed that DGK activation plays little or no role in the movement of granules from the cytosol to the membrane, pointing to more defined roles at the plasma membrane concerning granule fusion. Fusion of the target and vehicle membrane is achieved through the actions of SNARE (soluble N proteins ethylmalimide-sensitive factor accessory protein [SNAP] receptor) family proteins. SNARE family members are found on both the target membrane and the vehicle membrane and, according to the SNARE hypothesis, opposite SNARE complexes form a transient bridge promoting a trans-SNARE complex that promotes the fusion of two opposing biological membranes (36).
With PA likely to exert its effects at the plasma membrane, we aimed to investigate the role of the phospholipid in its ability to modulate membrane fusion and promote the release of intracellular Ca2+. R59022-inhibited neutrophils exhibited diminished calcium generation upon ANCA treatment that was restored in the presence of exogenous PA. However, adding exogenous calcium into the cell with the ionophore ionomycin did little to restore exocytosis in R59022-inhibited cells, indicating that exocytosis did not solely depend on PA-induced calcium release. The ability of PA to modulate membrane fusion was assessed by adding LPC into R59022-inhibited cells; here, also, the addition of LPC had no restorative effect on exocytosis. The observation that exocytosis was only restored when R59022-inhibited cells were treated with ANCA in the presence of both ionomycin and LPC indicates dual or multiple roles for PA that are generated through neutrophil activation by ANCA.
The formation of the trans-SNARE complex does not happen readily, since the outer membrane leaflet needs to bend in a negative curvature to preserve membrane integrity, while the inner leaflet requires opposite positive curvature (37). PA is able to promote a negative curvature in the membrane, since it is principally a cone-shaped molecule with relatively small phospholipid head groups (38). It has been reported that inhibiting PA production in the plasma membrane, but not in the donor granule membrane, inhibits granule fusion and release (39). PA may also promote the formation of the SNARE fusion complex directly by interacting with syntaxin, acting as structural support and reducing the energetic barrier required to fuse two membranes (40). In many cell types, including neutrophils, PA was demonstrated to modulate levels of intracellular Ca2+ (41), which in turn tightly regulates SNARE fusion (42,43). The SNARE complex alone has no Ca2+ sensory capacity yet uses an additional factor (synaptotagmin-1) for this role (44). The complete role of synaptotagmin-1 is yet to be fully elucidated, but reports suggest that the molecule is able to promote positive curvature of the vesicle membrane as well as binding distinct anionic phospholipids (such as PA) and syntaxin in the plasma target membrane (45,46).
Phosphatidic acid also plays a role in promoting neutrophil activation in response to fMLF treatment (47). However, in our experiments, fMLF-induced MPO release did not depend on PA generated by the actions of DGK. A number of previous reports suggested that PA generated as a result of fMLF treatment of neutrophils is likely to be the product of a PLD-dominant and not a DGK-driven pathway (48,49). We can also confirm that fMLF promotes neutrophil activation through a PLD pathway, since VU0155069 was able to significantly inhibit fMLF-induced MPO release in vitro. Although both ANCA IgG and fMLF are able to induce PA generation, it is likely that these are indicative of two separate pools with distinct properties. For example, PA generated through the actions of PLD may have roles in chemotaxis and modulation of the actin cytoskeleton (50,51), whereas it is evident from work here that the actions of DGK activated by ANCA IgG ligation do not modulate the actin cytoskeleton. We have previously documented the importance of the PI3K pathway in promoting ANCA-induced neutrophil activation (11), and this is likely to be central in promoting changes to the actin cytoskeleton (52).
It remains unclear which upstream pathways lead to DGK activation and the production of PA in ANCA- stimulated neutrophils. However, it is evident from this study that ligation of both the F(ab′)2 and Fc portions are required for ANCA-induced MPO release, and signaling pathways downstream of this ligation may promote the activation of DGK and the production of PA. We previously demonstrated that PI3K inhibition abrogates ANCA-induced neutrophil adhesion in vitro, which can be subsequently reversed by adding PA (13). Indeed, downstream products of PI3K including phosphatidylinositol-3,4,5-trisphosphate (PIP3) are known to be activators of DGK (53). Downstream pathways of FcγR ligation may also promote DGK activation, for example, src was previously identified as an activating tyrosine kinase of DGKs as well as inducing Ca2+ generation in ANCA-stimulated neutrophils (12,54). We also cannot rule out the possibility that these distinct mechanisms of PA activity are provided by different pools of PA. Both the location and timing of PA generation may have different effects on ongoing signal transduction pathways (55). The generation of PA may directly modulate both membrane fusion and calcium generation; however, it is also feasible that PA promotes the activation of downstream signaling pathways, which in turn may promote both the membrane modulation, calcium generation and other processes involved in neutrophil exocytosis. It has been reported that PA may directly promote the activation of a number of tyrosine and serine-threonine kinases (41,56,57), which may have similar effects to what we have reported here. For example, PA is known to promote activation of sphingosine kinase (58), which also acts as a modulator of intracellular calcium release in neutrophils (59,60).
In conclusion, we have demonstrated that the release of azurophilic granules from ANCA-stimulated neutrophils depends on the production of PA through the activation of DGK. We propose that PA promotes azurophil granule membrane fusion through its dual ability to modulate membrane fusion and promote increases in intracellular calcium. This pathway may be key, not only to the neutrophils’ ability to release harmful proteins into their microenvironment, but also for the neutrophils’ ability to adhere. Involvement of these molecular mechanisms in ANCA-induced exocytosis suggests that they are worthy of consideration as novel targets for treating vasculitic glomerulonephritis.
ACKNOWLEDGMENTS
The work was funded by a Stuart Strange Vasculitis Trust postdoctoral fellowship.
Footnotes
Online address: http://www.molmed.org
DISCLOSURE
C Savage is currently employed by GlaxoSmithKline PLC. The authors declare that they have no other competing interests as defined by Molecular Medicine, or other interests that might be perceived to influence the results and discussion reported in this paper.
REFERENCES
- 1.Lacy P. The role of Rho GTPases and SNAREs in mediator release from granulocytes. Pharmacol Ther. 2005;107:358–76. doi: 10.1016/j.pharmthera.2005.03.008. [DOI] [PubMed] [Google Scholar]
- 2.Mollinedo F, et al. Combinatorial SNARE complexes modulate the secretion of cytoplasmic granules in human neutrophils. J Immunol. 2006;177:2831–41. doi: 10.4049/jimmunol.177.5.2831. [DOI] [PubMed] [Google Scholar]
- 3.Mitchell T, Lo A, Logan MR, Lacy P, Eitzen G. Primary granule exocytosis in human neutrophils is regulated by Rac-dependent actin remodeling. Am J Physiol Cell Physiol. 2008;295:C1354–65. doi: 10.1152/ajpcell.00239.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Pak V, Budikhina A, Pashenkov M, Pinegin B. Neutrophil activity in chronic granulomatous disease. Adv Exp Med Biol. 2007;601:69–74. doi: 10.1007/978-0-387-72005-0_7. [DOI] [PubMed] [Google Scholar]
- 5.Paulino EC, de Souza LJ, Molan NA, Machado MC, Jancar S. Neutrophils from acute pancreatitis patients cause more severe in vitro endothelial damage compared with neutrophils from healthy donors and are differently regulated by endothelins. Pancreas. 2007;35:37–41. doi: 10.1097/MPA.0b013e31805c177b. [DOI] [PubMed] [Google Scholar]
- 6.Pankhurst T, Savage CO. Pathogenic role of anti-neutrophil cytoplasmic antibodies in vasculitis. Curr Opin Pharmacol. 2006;6:190–6. doi: 10.1016/j.coph.2005.10.009. [DOI] [PubMed] [Google Scholar]
- 7.Lu X, Garfield A, Rainger GE, Savage CO, Nash GB. Mediation of endothelial cell damage by serine proteases, but not superoxide, released from antineutrophil cytoplasmic antibody-stimulated neutrophils. Arthritis Rheum. 2006;54:1619–28. doi: 10.1002/art.21773. [DOI] [PubMed] [Google Scholar]
- 8.Baldus S, et al. Endothelial transcytosis of myeloperoxidase confers specificity to vascular ECM proteins as targets of tyrosine nitration. J Clin Invest. 2001;108:1759–70. doi: 10.1172/JCI12617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Astern JM, et al. Myeloperoxidase interacts with endothelial cell-surface cytokeratin 1 and modulates bradykinin production by the plasma Kallikrein-Kinin system. Am J Pathol. 2007;171:349–60. doi: 10.2353/ajpath.2007.060831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ben Smith A, Dove SK, Martin A, Wakelam MJ, Savage CO. Antineutrophil cytoplasm autoantibodies from patients with systemic vasculitis activate neutrophils through distinct signaling cascades: comparison with conventional Fcgamma receptor ligation. Blood. 2001;98:1448–55. doi: 10.1182/blood.v98.5.1448. [DOI] [PubMed] [Google Scholar]
- 11.Williams JM, et al. Activation of the G(i) heterotrimeric G protein by ANCA IgG F(ab′)2 fragments is necessary but not sufficient to stimulate the recruitment of those downstream mediators used by intact ANCA IgG. J Am Soc Nephrol. 2003;14:661–9. doi: 10.1097/01.asn.0000050223.34749.f4. [DOI] [PubMed] [Google Scholar]
- 12.Hewins P, Williams JM, Wakelam MJ, Savage CO. Activation of Syk in neutrophils by antineutrophil cytoplasm antibodies occurs via Fcgamma receptors and CD18. J Am Soc Nephrol. 2004;15:796–808. doi: 10.1097/01.asn.0000113241.98702.77. [DOI] [PubMed] [Google Scholar]
- 13.Williams JM, et al. Antineutrophil cytoplasm antibody-stimulated neutrophil adhesion depends on diacylglycerol kinase-catalyzed phosphatidic acid formation. J Am Soc Nephrol. 2007;18:1112–20. doi: 10.1681/ASN.2006090973. [DOI] [PubMed] [Google Scholar]
- 14.Pettitt TR, et al. Diacylglycerol and phosphatidate generated by phospholipases C and D, respectively, have distinct fatty acid compositions and functions: phospholipase D-derived diacylglycerol does not activate protein kinase C in porcine aortic endothelial cells. J Biol Chem. 1997;272:17354–9. doi: 10.1074/jbc.272.28.17354. [DOI] [PubMed] [Google Scholar]
- 15.Wakelam MJ. Diacylglycerol: when is it an intracellular messenger? Biochim Biophys Acta. 1998;1436:117–26. doi: 10.1016/s0005-2760(98)00123-4. [DOI] [PubMed] [Google Scholar]
- 16.Lominadze G, et al. Proteomic analysis of human neutrophil granules. Mol Cell Proteomics. 2005;4:1503–21. doi: 10.1074/mcp.M500143-MCP200. [DOI] [PubMed] [Google Scholar]
- 17.Jepsen LV, Skottun T. A rapid one-step method for the isolation of human granulocytes from whole blood. Scand J Clin Lab Invest. 1982;42:235–8. [PubMed] [Google Scholar]
- 18.de Chaffoy de Courcelles DC, Roevens P, Van Belle H. R 59 022, a diacylglycerol kinase inhibitor: its effect on diacylglycerol and thrombin-induced C kinase activation in the intact platelet. J Biol Chem. 1985;260:15762–70. [PubMed] [Google Scholar]
- 19.Jiang Y, Sakane F, Kanoh H, Walsh JP. Selectivity of the diacylglycerol kinase inhibitor 3-[2-(4-[bis-(4-fluorophenyl)methylene]-1-piperidinyl)ethyl]-2, 3-dihydro-2-thioxo-4(1H)quinazolinone (R59949) among diacylglycerol kinase subtypes. Biochem Pharmacol. 2000;59:763–72. doi: 10.1016/s0006-2952(99)00395-0. [DOI] [PubMed] [Google Scholar]
- 20.Agwu DE, McPhail LC, Wykle RL, McCall CE. Mass determination of receptor-mediated accumulation of phosphatidate and diglycerides in human neutrophils measured by Coomassie blue staining and densitometry. Biochem Biophys Res Commun. 1989;159:79–86. doi: 10.1016/0006-291x(89)92407-8. [DOI] [PubMed] [Google Scholar]
- 21.Rabinovitch PS, June CH, Grossmann A, Ledbetter JA. Heterogeneity among T cells in intracellular free calcium responses after mitogen stimulation with PHA or anti-CD3: simultaneous use of indo-1 and immunofluorescence with flow cytometry. J Immunol. 1986;137:952–61. [PubMed] [Google Scholar]
- 22.Carruthers DM, Arrol HP, Bacon PA, Young SP. Dysregulated intracellular Ca2+ stores and Ca2+ signaling in synovial fluid T lymphocytes from patients with chronic inflammatory arthritis. Arthritis Rheum. 2000;43:1257–65. doi: 10.1002/1529-0131(200006)43:6<1257::AID-ANR8>3.0.CO;2-Q. [DOI] [PubMed] [Google Scholar]
- 23.Rainger GE, Rowley AF, Nash GB. Adhesion-dependent release of elastase from human neutrophils in a novel, flow-based model: specificity of different chemotactic agents. Blood. 1998;92:4819–27. [PubMed] [Google Scholar]
- 24.Ting-Beall HP, Lee AS, Hochmuth RM. Effect of cytochalasin D on the mechanical properties and morphology of passive human neutrophils. Ann Biomed Eng. 1995;23:666–71. doi: 10.1007/BF02584463. [DOI] [PubMed] [Google Scholar]
- 25.Jog NR, et al. The actin cytoskeleton regulates exocytosis of all neutrophil granule subsets. Am J Physiol Cell Physiol. 2007;292:C1690–700. doi: 10.1152/ajpcell.00384.2006. [DOI] [PubMed] [Google Scholar]
- 26.Sheikh S, Gratzer WB, Pinder JC, Nash GB. Actin polymerisation regulates integrin-mediated adhesion as well as rigidity of neutrophils. Biochem Biophys Res Commun. 1997;238:910–15. doi: 10.1006/bbrc.1997.7407. [DOI] [PubMed] [Google Scholar]
- 27.Anderson SI, Behrendt B, Machesky LM, Insall RH, Nash GB. Linked regulation of motility and integrin function in activated migrating neutrophils revealed by interference in remodelling of the cytoskeleton. Cell Motil Cytoskeleton. 2003;54:135–46. doi: 10.1002/cm.10091. [DOI] [PubMed] [Google Scholar]
- 28.Pagano RE, Longmuir KJ, Martin OC. Intracellular translocation and metabolism of a fluorescent phosphatidic acid analogue in cultured fibroblasts. J Biol Chem. 1983;258:2034–40. [PubMed] [Google Scholar]
- 29.Kooijman EE, et al. Spontaneous curvature of phosphatidic acid and lysophosphatidic acid. Biochemistry. 2005;44:2097–102. doi: 10.1021/bi0478502. [DOI] [PubMed] [Google Scholar]
- 30.Zeniou-Meyer M, et al. Phospholipase D1 production of phosphatidic acid at the plasma membrane promotes exocytosis of large dense-core granules at a late stage. J Biol Chem. 2007;282:21746–57. doi: 10.1074/jbc.M702968200. [DOI] [PubMed] [Google Scholar]
- 31.Logan MR, Odemuyiwa SO, Moqbel R. Understanding exocytosis in immune and inflammatory cells: the molecular basis of mediator secretion. J Allergy Clin Immunol. 2003;111:923–32. [PubMed] [Google Scholar]
- 32.Williams JM, et al. Activation of the G(i) heterotrimeric G protein by ANCA IgG F(ab′)2 fragments is necessary but not sufficient to stimulate the recruitment of those downstream mediators used by intact ANCA IgG. J Am Soc Nephrol. 2003;14:661–9. doi: 10.1097/01.asn.0000050223.34749.f4. [DOI] [PubMed] [Google Scholar]
- 33.David A, Fridlich R, Aviram I. The presence of membrane proteinase 3 in neutrophil lipid rafts and its colocalization with FcgammaRIIIb and cytochrome b558. Exp Cell Res. 2005;308:156–65. doi: 10.1016/j.yexcr.2005.03.034. [DOI] [PubMed] [Google Scholar]
- 34.Skubitz KM, Campbell KD, Skubitz AP. CD63 associates with CD11/CD18 in large detergent-resistant complexes after translocation to the cell surface in human neutrophils. FEBS Lett. 2000;469:52–6. doi: 10.1016/s0014-5793(00)01240-0. [DOI] [PubMed] [Google Scholar]
- 35.Skubitz KM, Campbell KD, Iida J, Skubitz AP. CD63 associates with tyrosine kinase activity and CD11/CD18, and transmits an activation signal in neutrophils. J Immunol. 1996;157:3617–26. [PubMed] [Google Scholar]
- 36.Sollner T, Bennett MK, Whiteheart SW, Scheller RH, Rothman JE. A protein assembly-disassembly pathway in vitro that may correspond to sequential steps of synaptic vesicle docking, activation, and fusion. Cell. 1993;75:409–18. doi: 10.1016/0092-8674(93)90376-2. [DOI] [PubMed] [Google Scholar]
- 37.Lee AG. How lipids affect the activities of integral membrane proteins. Biochim Biophys Acta. 2004;1666:62–87. doi: 10.1016/j.bbamem.2004.05.012. [DOI] [PubMed] [Google Scholar]
- 38.Dickey A, Faller R. Examining the contributions of lipid shape and headgroup charge on bilayer behavior. Biophys J. 2008;95:2636–46. doi: 10.1529/biophysj.107.128074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Vicogne J, et al. Asymmetric phospholipid distribution drives in vitro reconstituted SNARE-dependent membrane fusion. Proc Natl Acad Sci U S A. 2006;103:14761–6. doi: 10.1073/pnas.0606881103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Lam AD, Tryoen-Toth P, Tsai B, Vitale N, Stuenkel EL. SNARE-catalyzed fusion events are regulated by syntaxin1A-lipid interactions. Mol Biol Cell. 2008;19:485–97. doi: 10.1091/mbc.E07-02-0148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Siddiqui RA, English D. Phosphatidic acid binding to human neutrophils: effects on tyrosine kinase-regulated intracellular Ca2+ mobilization. Cell Signal. 1996;8:349–54. doi: 10.1016/0898-6568(96)00072-1. [DOI] [PubMed] [Google Scholar]
- 42.Wadel K, Neher E, Sakaba T. The coupling between synaptic vesicles and Ca2+ channels determines fast neurotransmitter release. Neuron. 2007;53:563–75. doi: 10.1016/j.neuron.2007.01.021. [DOI] [PubMed] [Google Scholar]
- 43.Korchak HM, et al. Activation of the human neutrophil by calcium-mobilizing ligands. II. Correlation of calcium, diacyl glycerol, and phosphatidic acid generation with superoxide anion generation. J Biol Chem. 1988;263:11098–105. [PubMed] [Google Scholar]
- 44.Fernandez-Chacon R, et al. Synaptotagmin I functions as a calcium regulator of release probability. Nature. 2001;410:41–9. doi: 10.1038/35065004. [DOI] [PubMed] [Google Scholar]
- 45.Stein A, Radhakrishnan A, Riedel D, Fasshauer D, Jahn R. Synaptotagmin activates membrane fusion through a Ca2+-dependent trans interaction with phospholipids. Nat Struct Mol Biol. 2007;14:904–11. doi: 10.1038/nsmb1305. [DOI] [PubMed] [Google Scholar]
- 46.Martens S, Kozlov MM, McMahon HT. How synaptotagmin promotes membrane fusion. Science. 2007;316:1205–8. doi: 10.1126/science.1142614. [DOI] [PubMed] [Google Scholar]
- 47.Agwu DE, McPhail LC, Sozzani S, Bass DA, Mc-Call CE. Phosphatidic acid as a second messenger in human polymorphonuclear leukocytes: effects on activation of NADPH oxidase. J Clin Invest. 1991;88:531–9. doi: 10.1172/JCI115336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Mahadevappa VG. [3H]phosphatidic acid formed in response to FMLF is not inhibited by R59 022, a diacylglycerol kinase inhibitor. Biochem Biophys Res Commun. 1988;153:1097–104. doi: 10.1016/s0006-291x(88)81341-x. [DOI] [PubMed] [Google Scholar]
- 49.Rossi F, Grzeskowiak M, Della Bianca V, Calzetti F, Gandini G. Phosphatidic acid and not diacylglycerol generated by phospholipase D is functionally linked to the activation of the NADPH oxidase by FMLF in human neutrophils. Biochem Biophys Res Commun. 1990;168:320–7. doi: 10.1016/0006-291x(90)91711-z. [DOI] [PubMed] [Google Scholar]
- 50.Siddiqui RA, English D. Phosphatidic acid elicits calcium mobilization and actin polymerization through a tyrosine kinase-dependent process in human neutrophils: a mechanism for induction of chemotaxis. Biochim Biophys Acta. 1997;1349:81–95. doi: 10.1016/s0005-2760(97)00085-4. [DOI] [PubMed] [Google Scholar]
- 51.Nishikimi A, et al. Sequential regulation of DOCK2 dynamics by two phospholipids during neutrophil chemotaxis. Science. 2009;324:384–7. doi: 10.1126/science.1170179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Srinivasan S, et al. Rac and Cdc42 play distinct roles in regulating PI(3,4,5)P3 and polarity during neutrophil chemotaxis. J Cell Biol. 2003;160:375–85. doi: 10.1083/jcb.200208179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Cipres A, et al. Regulation of diacylglycerol kinase alpha by phosphoinositide 3-kinase lipid products. J Biol Chem. 2003;278:35629–35. doi: 10.1074/jbc.M305635200. [DOI] [PubMed] [Google Scholar]
- 54.Cutrupi S, et al. Src-mediated activation of alpha-diacylglycerol kinase is required for hepatocyte growth factor-induced cell motility. EMBO J. 2000;19:4614–22. doi: 10.1093/emboj/19.17.4614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Hodgkin MN, et al. Diacylglycerols and phosphatidates: which molecular species are intracellular messengers? Trends Biochem Sci. 1998;23:200–4. doi: 10.1016/s0968-0004(98)01200-6. [DOI] [PubMed] [Google Scholar]
- 56.Siddiqui RA, English D. Phosphatidylinositol 3′-kinase-mediated calcium mobilization regulates chemotaxis in phosphatidic acid-stimulated human neutrophils. Biochim Biophys Acta. 2000;1483:161–73. doi: 10.1016/s1388-1981(99)00172-9. [DOI] [PubMed] [Google Scholar]
- 57.Winter JN, Fox TE, Kester M, Jefferson LS, Kimball SR. Phosphatidic acid mediates activation of mTORC1 through the ERK signaling pathway. Am J Physiol Cell Physiol. 2010;299:C335–44. doi: 10.1152/ajpcell.00039.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Delon C, et al. Sphingosine kinase 1 is an intracellular effector of phosphatidic acid. J Biol Chem. 2004;279:44763–74. doi: 10.1074/jbc.M405771200. [DOI] [PubMed] [Google Scholar]
- 59.Aas V, Algeroy S, Sand KL, Iversen JG. Fibronectin promotes calcium signaling by interferon-gamma in human neutrophils via G-protein and sphingosine kinase-dependent mechanisms. Cell Commun Adhes. 2001;8:125–38. doi: 10.3109/15419060109080712. [DOI] [PubMed] [Google Scholar]
- 60.Itagaki K, Hauser CJ. Sphingosine 1-phosphate, a diffusible calcium influx factor mediating store-operated calcium entry. J Biol Chem. 2003;278:27540–7. doi: 10.1074/jbc.M301763200. [DOI] [PMC free article] [PubMed] [Google Scholar]