

Abstract
The mediators of tissue damage in systemic lupus erythematosus (SLE) such as antibodies, cytokines and activated immune cells have direct access to most organs in the body but must penetrate the blood-brain barrier (BBB) to gain access to brain tissue. We hypothesized that compromise of the BBB occurs episodically such that the brain will acquire tissue damage slowly and not at the same rate as other organs. On the basis of these assumptions, we wished to determine if duration of disease correlated with brain injury, as measured with functional magnetic resonance imaging (fMRI), and if this was independent of degree of tissue damage in other organs. We investigated differences in brain activation patterns using fMRI in 13 SLE patients stratified by disease duration of ≤2 years (short-term [ST]) or ≥10 years (long-term [LT]). Two fMRI paradigms were selected to measure working memory and emotional response (fearful faces task). Performance in the working memory task was significantly better in the ST group for one and two shape recall; however, both groups did poorly with three shape recall. Imaging studies demonstrated significantly increased cortical activation in the ST group in regions associated with cognition during the two shape retention phase of the working memory task (P < 0.001) and increased amygdala (P < 0.05) and superior parietal (P < 0.01) activation in response to the fearful faces paradigm. In conclusion, analysis of activation patterns stratified by performance accuracy, differences in co-morbid disease, corticosteroid doses or disease activity suggests that these observed differences are attributable to SLE effects on the central nervous system exclusive of vascular disease or other confounding influences. Our hypothesis is further supported by the lack of correlation between regional brain abnormalities on fMRI and the Systemic Lupus International Collaborating Clinics (SLICC) damage index.
INTRODUCTION
The pathogenic mechanisms contributing to central nervous system (CNS) injury in neuropsychiatric systemic lupus erythematosus (NPSLE) are poorly understood. Because of the presence of the blood-brain barrier (BBB), mechanisms for brain injury in systemic lupus erythematosus (SLE) are likely to be different than other organ systems that are more vulnerable to circulating cytokines, autoantibodies and activated lymphocytes. Vascular compromise resulting in brain ischemia is frequently related to anti phospholipid antibodies with associated microvascular disease and thrombotic events, premature atherosclerosis and, rarely, vasculitis. However, these mechanisms do not account for the majority of symptom complexes associated with NPSLE—in particular, the insidious cognitive decline manifested by up to 80% of lupus patients and the common mood disorders. Recent studies have demonstrated direct neurotoxic effects of lupus autoantibodies; antibodies to n-methyl-d-aspartate receptor (NMDAR), ribosomal P (anti-P), α-tubulin and phospholipid have been shown to bind neurons ex vivo with harmful effects (1–5) and all have been identified in the cerebrospinal fluid of SLE patients with CNS symptoms attributable to SLE (6–9). In murine studies, circulating antibrain antibodies can cause neuronal dysfunction and apoptosis after breach of the BBB (5,10–12), resulting in functional impairment on cognitive and behavioral tasks, as can antibodies directly injected intraventricularly. Moreover, studies of cerebrospinal fluid in patients with CNS manifestations of SLE have shown that a number of cytokines that can trigger brain tissue inflammation are also present, even when the cerebrospinal fluid is acellular (13–16). Finally, some of the medications taken by lupus patients also have the potential to damage neurons if they penetrate the BBB.
Nicotine, stress, hypertensive episodes and inflammatory mediators secondary to infection and other insults have been demonstrated to increase permeability of the BBB (17–19), for which the former status as an impermeable barricade has evolved to that of a responsive gatekeeper with transport, secretory and signaling functions. Repeated episodes of increased BBB permeability over time may play a permissive role in allowing harmful molecules from the circulation access to the CNS, where they subsequently initiate neurotoxic events (20). We hypothesized that SLE patients with long-term (LT) disease duration would be more likely to have sustained repeated episodes of BBB disruption and would therefore display more neuronal dysfunction than patients with short-term (ST) disease duration. Additionally, because of the protection afforded by the BBB, we predicted that the Systemic Lupus International Collaborating Clinics (SLICC) damage index (DI) would not correlate with CNS damage. The SLICC DI is a validated measure of irreversible organ damage that has accrued since diagnosis of SLE (21), and damage scores have been associated with disease outcome (22,23).
To that end, we sought to explore differences in brain activation patterns using functional magnetic resonance imaging (fMRI) studies. fMRI is a noninvasive imaging technology that contributes information on the neurophysiology of perception and cognition with high- resolution images of brain activity produced by local distribution of oxygenated hemoglobin (blood oxygen level–dependent [BOLD] signal) in response to specific tasks or stimuli. Therefore, fMRI provides an objective measure of functional neural networks solicited by specific probes. Three previous fMRI studies in SLE patients compared with healthy controls have all demonstrated abnormally increased regional brain activation patterns in areas targeted by specific tasks (24–26). We asked whether these abnormalities also correlate with disease duration. The goal of this pilot project was to investigate the ability of fMRI to ascertain differences in brain activation patterns in SLE patients stratified by disease duration. We expected that the effects of potentially neurotoxic molecules that have gained access to the CNS would accumulate as a function of time and that patients with LT disease would exhibit greater neural dysfunction.
MATERIALS AND METHODS
Patients
All subjects were 18 years of age or older, fulfilled American College of Rheumatology revised criteria for SLE and had stable disease activity and medication doses for 4 wks before screening. Exclusion criteria were designed to limit potentially confounding factors and included a history of neuropsychiatric impairment from any cause; current use of antidepressant, antipsychotic or anxiolytic drugs; a history of illicit drug use; claustrophobia; or metal implants. No patients with evidence of cerebrovascular disease on structural MRI scan were studied. Subjects were enrolled at Columbia University Medical Center (CUMC). The study was approved by the institutional review boards at CUMC and the Feinstein Institute for Medical Research and informed consent was obtained before study procedures for each subject.
Patients were additionally selected on the basis of disease duration >10 years (LT) or <2 years (ST), and 19 subjects were recruited. Because of technical issues with the paradigms and subject compliance, 13 subjects (6 ST, 7 LT) had analyzable data for the working memory task and 11 subjects (6 ST, 5 LT) had analyzable data for the amygdala task. At enrollment, all subjects had a clinical evaluation including assessments of disease activity (Systemic Lupus Erythematosus Disease Activity Index [SLEDAI] [27]), damage (SLICC DI [21]) and comorbid disease. Subjects’ records were also reviewed for elevated titers of IgG or IgM anticardiolipin antibodies. fMRI scans were scheduled within 2 wks of the clinical assessment.
Data Acquisition
MR imaging was done with a 3T GE Signa MRI Scanner, using a gradient-echo, echo planar imaging with BOLD contrast pulse sequence. A total of 28 axial slices were acquired along the anterior commissure–posterior commissure (AC-PC) plane, with a 64 × 64 image acquisition matrix and 19-cm field of view (voxel size 3.0 × 3.0 × 4.5 mm). A total of 280 volumes were acquired during the face paradigm, and 312 volumes were acquired in the shapes paradigm of the functional scans. Repetition time, (TR) was 2 s, and echo delay time (TE) was 40 ms. Structural MRI data were acquired using a three-dimensional longitudinal relaxation time (T1)-weighted spoiled gradient recalled pulse sequence with isomorphic voxels (1 × 1 × 1 mm) with a 24-cm field of view (256 × 256 matrix, 128 slices, TR 34 ms, TE 3 ms). All subjects had a noncontrast structural MRI scan before fMRI assessment, and only individuals with no evidence of cerebrovascular disease were included in this study.
fMRI Paradigms
Selection of fMRI paradigms designed to probe areas that contribute to working memory (hippocampus) and fear/anxiety functions (amygdala) was guided by the presence of high-density NMDAR in the hippocampus and amygdala (28), the prevalence of cognitive and emotional impairment in SLE patients and previous work demonstrating that these areas are targeted by the anti-NMDAR antibody in the murine model.
Modified Sternberg/working memory paradigm
This delayed response task is based on the original Sternberg letter recognition task (29); it uses unfamiliar shapes as stimuli and uses set size as a proxy for working memory load. Subjects are shown a blank screen for 3 s, followed by one, two or three test figures for 3 s (encoding phase). The blank screen returns for 7 s as subjects are instructed to remember the figures (retention phase). A probe image is then displayed, and the subject presses a button to indicate whether this figure matches one of the previously shown test figures. Three experimental blocks of 620 s were presented with 60 s between blocks. Each block contained 30 trials in total; 10 for each of the three set sizes (one, two or three figures) with five correct (matching) and five incorrect (non-matching) probe figures per set size. Three training blocks were administered before fMRI testing, with feedback about accuracy and response time to reduce task-related skill learning during the experiment.
Fearful faces paradigm
Conscious (nonmasked) and unconscious (masked) perception of fearful faces displayed on a screen has been used previously to investigate amygdala function (30). Fearful and neutral faces were chosen from a standardized series developed by Ekman and Friesen (31), cropped into an elliptical shape and color coded. After a brief fixation cue (200 ms) followed by a blank screen (400 ms), either a nonmasked (200 ms for fearful or neutral face) or masked (33 ms for fearful or neutral face, followed by 167 ms of a neutral face) face was presented. The patient had 1,200 ms to indicate the color of the face. There were eight trials for each of the stimulus types, and each stimulus type was performed three times (30).
Image Processing and Statistical Analysis
The SPM5 image analysis package (http://www.fil.ion.ucl.ac.uk/spm/software/spm5) was used to process fMRI data. After the first four volumes were excluded, the remaining images were slice time-corrected, coregistered to the patient’s structural image and realigned to the 100th image volume to correct for subject movement. Volumes were then normalized to the standardized template at a voxel size of 3 × 3 × 3 mm and smoothed using a Gaussian kernel of 6 mm full width at half-maximum (32).
Statistical analysis of the functional data for individual subjects used a standard general linear model using realignment parameters as a covariate. The contrast images produced from the first-level subject analysis were used in a group level analysis. The amygdala mask was created using an automated atlas-based masking technique, where the standard mask was dilated by a factor of 5 to ensure that it encompassed the entire amygdala (33). A group level simple regression model using contrast images of one, two and three shapes retention during the modified Sternberg test was used with amygdala data: age and DI as covariates of interest. Mann-Whiney and Fisher exact tests were used to determine significant differences in accuracy performance for each set size between groups during the working memory paradigm as well as differences in baseline clinical characteristics between groups.
RESULTS
Subjects and Task Performance
Subject characteristics are shown in Table 1. Other than the expected difference in disease duration, the LT group had increased mean age and DI compared with the ST group. Task performance was assessed by looking at whole brain composites from both groups for each task (Figure 1).
Table 1.
Subject characteristics.
| ST (n = 6) (disease duration <2 years) | LT (n = 7) (disease duration >10 years) | P | |
|---|---|---|---|
| Disease duration (years) | 0.79 ± 0.49 | 15.29 ± 5.53 | 0.003a |
| Age | |||
| Mean | 25.7 ± 2.66 | 43.1 ± 9.16 | 0.008a |
| Median | 25.5 | 45 | |
| Range | 23–29 | 27–55 | |
| Sex | |||
| Female (%) | 83 | 100 | |
| Male (%) | 17 | 0 | |
| Ethnicity | |||
| African American (%) | 17 | 57 | 0.242 |
| Hispanic (%) | 83 | 29 | 0.103 |
| Caucasian (%) | 0 | 14 | 0.462 |
| Education (years) | 12.83 ± 2.14 | 12.85 ± 3.02 | 0.942 |
| SLEDAI | 1.67 ± 1.5 | 2.0 ± 2.30 | 0.940 |
| DI | 0.17 ± 0.41 | 1.29 ± 1.25 | 0.048a |
| History of NPSLE | 0 | 0 | |
| Anti-dsDNA titer (%)b | 33 | 43 | 0.59 |
| Anticardiolipin (%)c | 33 | 29 | 0.72 |
| Prednisoned | 14.17 ± 14.71 | 4.11 ± 5.62 | 0.127 |
| Cellcept(current) | 67% | 14% | 0.063 |
| Azathioprine(current) | 0 | 14% | 0.355 |
| Cyclophosphamide(current) | 17% | 0 | 0.280 |
| Current DMARDe | 83% | 29% | 0.058 |
| DMARD ever | 83% | 71% | 0.626 |
| Hypertensionf | 50% | 0 | 0.181 |
| Diabetes mellitusg | 0 | 14% | 0.355 |
P < 0.05.
Current anti-dsDNA titer; percent positive.
Anticardiolipin antibody titer; percent positive IgG or IgM by history.
Current prednisone dose.
DMARD, disease-modifying antirheumatic drugs include cyclophosphamide, methotrexate, azathioprine or cellcept for >2 months.
Current hypertension requiring medication.
Current diabetes mellitus requiring medication.
Figure 1.

Composite activation areas. (A) A composite of activation areas with a minimum size of 10 voxels from all subjects during the two shapes retention phase of the working memory paradigm. Areas with increased activation include BA40 (supramarginal gyrus, parietal lobe), somatomotor cortex, cingulated gyrus, BA9 (granular/prefrontal area), BA22 (superior temporal; auditory) and BA17 (occipital; visual). (B) A composite of activation areas from all subjects during unmasked (conscious) stimulus with fearful and neutral faces.
Working Memory Paradigm
LT subjects demonstrate decreased accuracy on behavioral tests
In the modified Sternberg task, accuracy inversely correlated with test set size, and significant performance differences between groups were noted for one and two figure tasks. For LT subjects, average accuracy for one, two and three test figures was 79.51%, 68.57% and 67.71%, respectively, compared with 92.8%, 85.57% and 62.33% for the ST group. The group differences were statistically significant for one (P = 0.038) and two (P = 0.032) test figures; both groups performed equally poorly on the three-figure tests. BOLD contrast levels during the two shape retention phase were selected for analysis on the basis of the performance data demonstrating the largest discrepancy in accuracy for two shape recall.
ST subjects demonstrate increased whole-brain activation patterns
There was slightly increased hippocampus activation in the ST group during the two shape retention phase (P < 0.05 uncorrected, data not shown). Additionally, analysis of whole brain activation patterns outside the hippocampus during the two shapes retention phase revealed an increased BOLD signal in the cingulate gyrus, prefrontal cortex, somatomotor cortex and Brodmann’s area 40 (BA40) in ST subjects compared with LT subjects (P < 0.001) (Figure 2 and Table 2). Significant inverse correlations were found between disease duration and activation in the cingulate gyrus (R = −0.9, P < 0.001), prefrontal cortex (R = −0.641, P < 0.05), somatomotor cortex (R = −0.739, P = 0.009), BA40 (R = −0.732, P = 0.01) and dorsolateral pre-frontal cortex (DLPFC) (R = −0.718, P < 0.05). There were no significant group differences in whole brain activation patterns during one and three shape retention or encoding phases. Additionally, there were no significant correlations between SLEDAI scores and brain activation patterns.
Figure 2.

Group differences during the working memory task. Significant group differences in regional brain activation outside the hippocampus during two figure retention in the working memory paradigm shows increased activation in ST subjects compared to LT subjects in the somatomotor cortex, BA40, cingulated gyrus and prefrontal cortex (P < 0.001). The coordinates and voxel sizes of these clusters are listed in Table 2.
Table 2.
Group differences in activation during the working memory two shapes retention.
| Brain region | Coordinates | Cluster size (number voxels) | Z value | P value | ||
|---|---|---|---|---|---|---|
|
| ||||||
| X | Y | Z | ||||
| Somatomotor cortex | 39 | −39 | 33 | 34 | 4.18 | <0.001 |
| BA40 | 21 | −24 | 39 | 37 | 4.01 | <0.001 |
| Cingulate gyrus | 0 | 6 | 24 | 45 | 3.76 | <0.001 |
| Prefrontal cortex | 53 | 3 | 24 | 39 | 3.67 | <0.001 |
The coordinates and cluster sizes of the regions with significant differences in BOLD signal intensity between the ST and LT subjects during performance of the modified Sternberg task (two shapes retention phase) are listed. These results have been corrected for cluster extent (P ≤ 0.001) and for multiple comparisons (P ≤ 0.05).
Activation patterns stratified by performance accuracy
Activation patterns of selected subgroups performing with similar accuracy on the Sternberg test were compared using an equiperformance analysis to investigate the potentially confounding influence of age on observed group differences in whole brain activation patterns during the two-shape retention phase (34,35). Three LT subjects and two ST subjects with accuracy scores at the extremes of high and low were not evaluated, leaving four subjects with significant age differences in each group who displayed similar performance accuracy on each of the Stern-berg tests. Comparison of BOLD signal intensity during the two-shape retention phase between the selected subgroups showed no group differences at the P < 0.001 level. These data indicate that age is not a significant contributor to accuracy and that the observed differences in activation patterns during the two shape retention phase are reflective of disease-related injury and not age.
Fearful Faces Paradigm
ST subjects demonstrate increased amygdala response
There was increased amygdala response to stimulation with unmasked neutral and fearful faces in the ST group compared with the LT group (P < 0.05 uncorrected) (Figure 3 and Table 3). In contrast to the working memory task, cortical representation of the fearful faces paradigm demonstrated only one area, BA7 (superior parietal area), with significant regional brain activation outside of the amygdala in the ST group compared with the LT group (P < 0.01). Similar to the results of the modified Sternberg test, a significant inverse correlation between amygdala activation and disease duration was demonstrated (R = −0.846, P = 0.001) and no correlations were demonstrated between SLEDAI scores and amygdala activation patterns. Few discrepancies in BOLD signal intensity were noted between subjects in the ST group (coefficient of variance 0.387) compared with a large variance demonstrated between subjects in the LT group (coefficient of variance 15.98).
Figure 3.
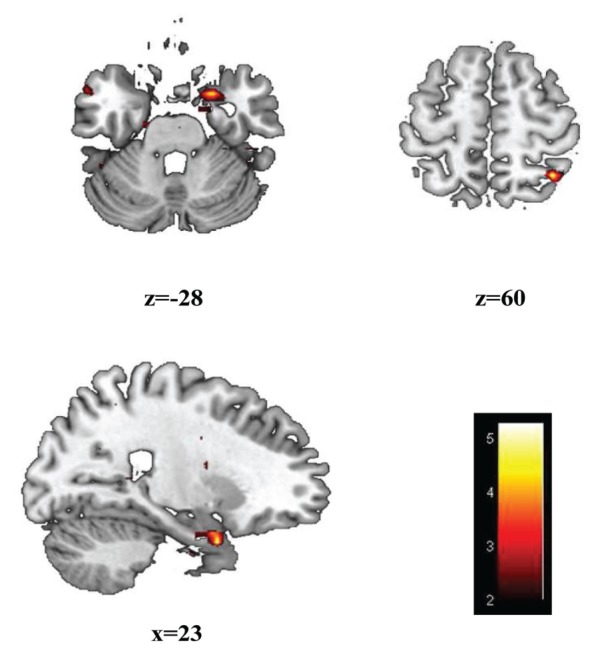
Group differences during the fearful faces paradigm. Significant group differences (P < 0.05) during conscious stimulation with fearful and neutral faces during the fearful faces paradigm shows increased activation in ST subjects compared to LT subjects in the amygdala and BA7 (superior parietal lobe). The coordinates and voxel sizes of these clusters are listed in Table 3.
Table 3.
Group differences in activation during the fearful faces paradigm.
| Brain region | Coordinates | Cluster size (number voxels) | Z value | P value | ||
|---|---|---|---|---|---|---|
|
| ||||||
| X | Y | Z | ||||
| Amygdala-R | −18 | −15 | −33 | 27 | 3.48 | <0.05 |
| Amygdala-L | 21 | 6 | −30 | 159 | 3.41 | <0.05 |
| BA7 | 36 | −54 | 60 | 74 | 3.39 | <0.05 |
The coordinates and cluster sizes of the regions with significant differences in BOLD signal intensity between the ST and LT subjects during performance of the fearful faces task are listed. These results have been corrected for cluster extent (P ≤ 0.05).
Interparadigm Correlations
Amygdala activation stimulated by the fearful faces paradigm was used as a covariate in a group level regression analysis of the working memory paradigm, yielding significant positive correlations in BA9 (granular frontal area), BA42 (posterior transverse temporal area), BA40 (supramarginal area) and three distinct regions in the left and right DLPFC (P < 0.001 for all). A negative correlation was demonstrated between amygdala activation and activation in the precuneus (P < 0.001).
Associations with Damage Index
No associations were noted between the DI and brain activation patterns
We wanted to determine whether abnormality on fMRI correlated with the SLICC DI (21) or whether, because the brain is protected by the BBB from potential tissue insults, there would be no correlation. Importantly, none of the subjects scored damage in the neurologic domains on the DI. The simple group regression analysis yielded no significant correlations between DI and age or regional brain activation during two shape retention in the modified Sternberg test. Additionally, analysis of contrast images from one, two and three shape retention phases of the modified Sternberg task in subjects stratified by DI (DI = 0, n = 7, and DI >0, n = 6) revealed no significant between-group differences. Comparison of contrast images during the fearful faces task from subjects grouped by DI (DI = 0, n = 6, and DI >0, n = 5) also showed no significant group differences.
DISCUSSION
This study demonstrates significant differences in brain activation patterns elicited by specific probes for cognitive and emotional response using fMRI imaging in SLE patients stratified by disease duration. Analysis of activation patterns stratified by performance accuracy, the absence of prior CNS events or structural changes on MRI in any subject and absence of significant differences in comorbid disease, current steroid doses or active systemic disease between the two groups suggest that the differences in brain activation patterns are attributable to SLE effects on the CNS, exclusive of vascular disease or other confounding influences, and these effects are a function of time, as predicted by our hypothesis.
Two fMRI studies previously reported increased regional activation patterns in patients with NPSLE in response to memory and simple motor tasks compared with healthy controls (25,26). Another fMRI study of 10 pediatric SLE patients, not selected for NPSLE, also reported increased cortical activation patterns compared with healthy controls in response to memory, attention and language processing tasks (24). Similarly, ST patients in this study demonstrated significantly increased neural activation in cortical areas associated with working memory (cingulate gyrus, prefrontal cortex, somatomotor cortex and BA40) compared with the LT group. Our findings of increased cortical activation patterns in ST patients suggest that ST patients do not display normal cognitive processes assessed by fMRI and that LT patients have evidence of greater neuronal damage reflected in decreased BOLD signal intensity. The abnormal findings in our ST population are consistent with previously reported findings of abnormal MRI, manifested as atrophy and/or focal lesions, in 25% of newly diagnosed patients (36).
It is possible that the increased cortical activation pattern observed in the ST group reflects increased recruitment of neural networks to solve relatively simple tasks, implying that these patients have already sustained neurologic injury to areas related to working memory. This result is analogous to the increased cortical activation patterns reported with diffuse and focal brain damage in other neurodegenerative diseases and prior SLE studies. Functional neuroimaging studies of diffuse neurodegenerative disease such as human immunodeficiency virus (HIV), multiple sclerosis and Alzheimer’s disease demonstrate consistently increased neural activation in pathways related to cognitive processes, suggesting compensatory recruitment of neural circuitry for damaged brain (37–39). Additionally, cross-sectional and longitudinal imaging of focal CNS damage (such as stroke) demonstrates similar evidence of increased neural recruitment after localized brain injury, which contributes to the reorganization of neural networks and functional recovery (40,41). In this study, the LT group no longer displays the increased activation pattern demonstrated by the ST group, suggesting that this group has sustained more extensive damage. These data are supported by the performance scores. Although there was an expected performance decrement with increasing working memory load (set size) in both groups, the LT group displayed significantly greater impairment for one and two shapes. The precipitous drop in accuracy in the ST group with three shapes to a level lower than the LT group (62.33% versus 67.71%) suggests an abnormal threshold effect in the working memory response of this group. One important caveat is the potentially confounding influence of volitional parameters such as attention, concentration and motivation on performance in functional task-related studies, but there is no obvious reason to believe these should vary between groups.
The amygdala is critical to the formation of emotional memory, and we report significant group differences in amygdala activation during conscious perception of fearful faces. The amygdala response in the ST group is similar to that reported for healthy subjects (30), whereas the decreased amygdala activation in the LT group is again indicative of regional neuronal damage. Selective damage to the amygdala associated with anti-NMDAR antibodies has been previously reported in humans (42) and is supported by a murine model (10). BA7 overlaps with the precuneus in the parietal lobe and is considered to be involved in somatosensory association as well as episodic memory retrieval (43). Decreased activation of BA7 in the LT group may reflect damage to a neural network involving the amygdala and BA7, as seen in the cortical response to the memory task in this group.
The interdependency of the two paradigms was explored by using amygdala activation as a covariate in the working memory paradigm, and positive correlations were seen in areas involved in cognitive functions including working memory, sustained attention, executive function, visuospatial attention and emotional responses (the DLPFC, BA9, BA42 and BA40). The results of this analysis confirm the known connectivity of the brain regions activated by our paradigms and reinforce the hypothesis that disease-related brain damage increases with disease duration.
Because age-dependent changes in regional neural activity related to cognitive functions in healthy individuals were previously demonstrated (44,45), our findings are potentially confounded by the mean age difference between groups. The study was not designed to investigate differences between SLE patients and healthy controls, but to discern differences between SLE patients with LT or ST disease duration. Not surprisingly, these groups differed in age. However, an equiperformance analysis (34,35), using the data from the modified Stern-berg test, failed to demonstrate differences in BOLD signal intensity between groups when matched for performance, despite significant age differences. We therefore conclude, with the caveat that our numbers are small, that the observed decrease in brain activation patterns in the LT group in response to the two paradigms is independent of age and may be reflective of progressive disease- related neuronal dysfunction.
The SLICC DI is a validated measure of irreversible organ damage, from any etiology that has accrued since disease diagnosis, and higher scores are associated with poor outcome (21–23). We predicted that, because of the protection afforded by the BBB and therefore a different proposed mechanism of injury, the DI may not be a sensitive indicator of CNS damage. None of the subjects enrolled scored damage in the neurologic domains. As expected, the DI in the LT group was greater than the ST group; however, the absence of associations found between DI and age, DI and BOLD signal intensity or BOLD signal stratified by DI suggests that the DI is not a useful predictor of brain dysfunction and that susceptibility of the brain to damage is different from that of other organs. This result reinforces our hypothesis that antibody-mediated neuronal damage in SLE requires interruption of the BBB; as a validated measure of organ damage in SLE, most of which is mediated by circulating inflammatory cytokines or autoantibodies, the DI would not be expected to reflect damage in an organ protected by the BBB. This result is supported by previous studies that have reported correlations between NPSLE and antibrain antibodies or cytokines in cerebrospinal fluid, as opposed to peripheral blood (6–9) and studies showing that the progression of cognitive impairment is independent of disease activity. Most importantly, the current study suggests that DI is not a surrogate marker for neurological damage in SLE, and other measures such as fMRI must be explored.
The potential significance of medication effects is difficult to ascertain. More ST subjects were undergoing treatment with immunosuppressive medications at the time of evaluation, but exposure was equivalent between groups. Deleterious effects of cyclophosphamide on cognition have been reported in oncology; however, oncologic doses exceed those commonly used in SLE patients. Variable effects of corticosteroids on cognition have been reported and depend on the duration, total dose and timing of treatment; acute, high doses have reversible negative effects on memory performance (46,47), whereas chronic use results in impaired memory performance as well as hippocampus atrophy and gray matter reduction (48–53). Conversely, short-term, low-dose corticosteroid may enhance memory and learning and augment fear and anxiety responses through effects on the hippocampus and amygdala (46,54,55). Importantly, several studies have not found significant correlations between corticosteroids and cognitive dysfunction or abnormalities in brain volume in SLE (56–61). Our cross-sectional analysis of these two groups is not adequate to address this issue.
In conclusion, our results in this study indicate that use of fMRI with selected working memory and fear conditioning paradigms may provide a reliable measure of neuronal damage in SLE patients. The study design allowed us to evaluate the effect of disease duration on task-stimulated brain activation patterns that is independent of age and performance accuracy. We hypothesized that CNS damage, that is unrelated to vascular injury, is a function of circulating autoantibodies, BBB permeability and time. Patients with longer disease duration would therefore be expected to exhibit evidence of increased CNS damage. The group differences observed in behavioral performance as well as on the imaging studies support this hypothesis. We could not control for the effects of exposure to long-term medications, and this remains a potential confounder, as does age, since our numbers for the equiperformance analysis were small. Additionally, the DI was not predictive of neuronal damage in our cohort, reinforcing the hypothesis that mechanisms of tissue damage in the CNS differs from tissue damage in the periphery. Thus, studies of lupus therapeutics must begin to address CNS damage as well as disease activity and current measures of damage accrual. Further longitudinal investigations are warranted to determine the potential use of fMRI as an outcome measure in clinical trials of therapeutic agents. Identification of a noninvasive imaging technique that has the sensitivity to detect incremental change and can be used for determination of efficacy of agents designed to block antibody access to the CNS is critical to the success of a clinical trial.
ACKNOWLEDGMENTS
This study was supported by funding from the National Institutes of Health (Grant P01AI073693) and a Clinical Research Award from the New York Chapter of the Arthritis Foundation. None of the authors have financial interests that could create a potential conflict of interest or the appearance of a conflict of interest with regard to the work.
Footnotes
Online address: http://www.molmed.org
DISCLOSURE
The authors declare that they have no competing interests as defined by Molecular Medicine, or other interests that might be perceived to influence the results and discussion reported in this paper.
REFERENCES
- 1.Caronti B, et al. Anti-beta 2-glycoprotein I antibodies bind to central nervous system. J Neurol Sci. 1998;156:211–9. doi: 10.1016/s0022-510x(98)00027-6. [DOI] [PubMed] [Google Scholar]
- 2.DeGiorgio LA, et al. A subset of lupus anti-DNA antibodies cross-reacts with the NR2 glutamate receptor in systemic lupus erythematosus. Nat Med. 2001;7:1189–93. doi: 10.1038/nm1101-1189. [DOI] [PubMed] [Google Scholar]
- 3.Kent MN, et al. Ultrastructural localization of monoclonal antiphospholipid antibody binding to rat brain. Exp Neurol. 2000;163:173–9. doi: 10.1006/exnr.2000.7358. [DOI] [PubMed] [Google Scholar]
- 4.Matus S, et al. Antiribosomal-P autoantibodies from psychiatric lupus target a novel neuronal surface protein causing calcium influx and apoptosis. J Exp Med. 2007;204:3221–34. doi: 10.1084/jem.20071285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ndhlovu M, et al. Identification of alphatubulin as an autoantigen recognized by sera from patients with neuropsychiatric systemic lupus erythematosus. Brain Behav Immun. 2011;25:279–85. doi: 10.1016/j.bbi.2010.09.019. [DOI] [PubMed] [Google Scholar]
- 6.Arinuma Y, Yanagida T, Hirohata S. Association of cerebrospinal fluid anti-NR2 glutamate receptor antibodies with diffuse neuropsychiatric systemic lupus erythematosus. Arthritis Rheum. 2008;58:1130–5. doi: 10.1002/art.23399. [DOI] [PubMed] [Google Scholar]
- 7.Fragoso-Loyo H, et al. Serum and cerebrospinal fluid autoantibodies in patients with neuropsychiatric lupus erythematosus: implications for diagnosis and pathogenesis. PLoS One. 2008;3:e3347. doi: 10.1371/journal.pone.0003347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hirohata S, et al. Association of cerebrospinal fluid anti-ribosomal p protein antibodies with diffuse psychiatric/neuropsychological syndromes in systemic lupus erythematosus. Arthritis Res Ther. 2007;9:R44. doi: 10.1186/ar2184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Yoshio T, et al. Association of IgG anti-NR2 glutamate receptor antibodies in cerebrospinal fluid with neuropsychiatric systemic lupus erythematosus. Arthritis Rheum. 2006;54:675–8. doi: 10.1002/art.21547. [DOI] [PubMed] [Google Scholar]
- 10.Huerta PT, et al. Immunity and behavior: antibodies alter emotion. Proc Natl Acad Sci U S A. 2006;103:678–83. doi: 10.1073/pnas.0510055103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kowal C, et al. Human lupus autoantibodies against NMDA receptors mediate cognitive impairment. Proc Natl Acad Sci U S A. 2006;103:19854–9. doi: 10.1073/pnas.0608397104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Faust TW, et al. Neurotoxic lupus auto -antibodies alter brain function through two distinct mechanisms. Proc Natl Acad Sci U S A. 2010;107:18569–74. doi: 10.1073/pnas.1006980107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.George-Chandy A, Trysberg E, Eriksson K. Raised intrathecal levels of APRIL and BAFF in patients with systemic lupus erythematosus: relationship to neuropsychiatric symptoms. Arthritis Res Ther. 2008;10:R97. doi: 10.1186/ar2484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hirohata S, Miyamoto T. Elevated levels of interleukin-6 in cerebrospinal fluid from patients with systemic lupus erythematosus and central nervous system involvement. Arthritis Rheum. 1990;33:644–9. doi: 10.1002/art.1780330506. [DOI] [PubMed] [Google Scholar]
- 15.Katsumata Y, et al. Diagnostic reliability of cerebral spinal fluid tests for acute confusional state (delirium) in patients with systemic lupus erythematosus: interleukin 6 (IL-6), IL-8, interferon-alpha, IgG index, and Q-albumin. J Rheumatol. 2007;34:2010–7. [PubMed] [Google Scholar]
- 16.Trysberg E, Carlsten H, Tarkowski A. Intrathecal cytokines in systemic lupus erythematosus with central nervous system involvement. Lupus. 2000;9:498–503. doi: 10.1177/096120330000900704. [DOI] [PubMed] [Google Scholar]
- 17.Banks WA, Erickson MA. The blood-brain barrier and immune function and dysfunction. Neurobiol Dis. 2010;37:26–32. doi: 10.1016/j.nbd.2009.07.031. [DOI] [PubMed] [Google Scholar]
- 18.Carvey PM, Hendey B, Monahan AJ. The blood-brain barrier in neurodegenerative disease: a rhetorical perspective. J Neurochem. 2009;111:291–314. doi: 10.1111/j.1471-4159.2009.06319.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hawkins BT, et al. Nicotine increases in vivo blood-brain barrier permeability and alters cerebral microvascular tight junction protein distribution. Brain Res. 2004;1027:48–58. doi: 10.1016/j.brainres.2004.08.043. [DOI] [PubMed] [Google Scholar]
- 20.Diamond B, et al. Losing your nerves? Maybe it’s the antibodies. Nat Rev Immunol. 2009;9:449–56. doi: 10.1038/nri2529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Gladman DD, et al. The reliability of the Systemic Lupus International Collaborating Clinics/American College of Rheumatology Damage Index in patients with systemic lupus erythematosus. Arthritis Rheum. 1997;1997;40:809–13. doi: 10.1002/art.1780400506. [DOI] [PubMed] [Google Scholar]
- 22.Nossent JC. SLICC/ J Rheumatol. Vol. 25. ACR; 1998. Damage Index in Afro-Caribbean patients with systemic lupus erythematosus: changes in and relationship to disease activity, corticosteroid therapy, and prognosis; pp. 654–9. [PubMed] [Google Scholar]
- 23.Gladman DD, Urowitz MB. The SLICC/ACR damage index: progress report and experience in the field. Lupus. 1999;8:632–7. doi: 10.1191/096120399680411335. [DOI] [PubMed] [Google Scholar]
- 24.DiFrancesco MW, et al. Functional magnetic resonance imaging assessment of cognitive function in childhood-onset systemic lupus erythematosus: a pilot study. Arthritis Rheum. 2007;56:4151–63. doi: 10.1002/art.23132. [DOI] [PubMed] [Google Scholar]
- 25.Fitzgibbon BM, et al. Functional MRI in NPSLE patients reveals increased parietal and frontal brain activation during a working memory task compared with controls. Rheumatology (Oxford) 2008;47:50–3. doi: 10.1093/rheumatology/kem287. [DOI] [PubMed] [Google Scholar]
- 26.Rocca MA, et al. An fMRI study of the motor system in patients with neuropsychiatric systemic lupus erythematosus. Neuroimage. 2006;30:478–84. doi: 10.1016/j.neuroimage.2005.09.047. [DOI] [PubMed] [Google Scholar]
- 27.Bombardier C, et al. Derivation of the SLEDAI: a disease activity index for lupus patients. The Committee on Prognosis Studies in SLE. Arthritis Rheum. 1992;35:630–40. doi: 10.1002/art.1780350606. [DOI] [PubMed] [Google Scholar]
- 28.Ozawa S, Kamiya H, Tsuzuki K. Glutamate receptors in the mammalian central nervous system. Prog Neurobiol. 1998;54:581–618. doi: 10.1016/s0301-0082(97)00085-3. [DOI] [PubMed] [Google Scholar]
- 29.Sternberg S. High-speed scanning in human memory. Science. 1966;153:652–4. doi: 10.1126/science.153.3736.652. [DOI] [PubMed] [Google Scholar]
- 30.Etkin A, et al. Individual differences in trait anxiety predict the response of the basolateral amygdala to unconsciously processed fearful faces. Neuron. 2004;44:1043–55. doi: 10.1016/j.neuron.2004.12.006. [DOI] [PubMed] [Google Scholar]
- 31.Ekman P, Friesen WV. Pictures of Facial Affect. Palo Alto, CA: Consulting Psychologists; 1976. [Google Scholar]
- 32.Friston KJ, et al. Statistical parametric maps in functional imaging: a general linear approach. Hum Brain Mapp. 1995;2:189–210. [Google Scholar]
- 33.Maldjian JA, et al. An automated method for neuroanatomic and cytoarchitectonic atlas-based interrogation of fMRI data sets. Neuroimage. 2003;19:1233–9. doi: 10.1016/s1053-8119(03)00169-1. [DOI] [PubMed] [Google Scholar]
- 34.Carbon M, et al. Increased cerebellar activation during sequence learning in DYT1 carriers: an equiperformance study. Brain. 2008;131:146–54. doi: 10.1093/brain/awm243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Mentis MJ, et al. Enhancement of brain activation during trial-and-error sequence learning in early PD. Neurology. 2003;60:612–9. doi: 10.1212/01.wnl.0000044154.92143.dc. [DOI] [PubMed] [Google Scholar]
- 36.Petri M, et al. Brain magnetic resonance imaging in newly diagnosed systemic lupus erythematosus. J Rheumatol. 2008;35:2348–54. doi: 10.3899/jrheum.071010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Bobholz JA, et al. fMRI study of episodic memory in relapsing-remitting MS: correlation with T2 lesion volume. Neurology. 2006;67:1640–5. doi: 10.1212/01.wnl.0000242885.71725.76. [DOI] [PubMed] [Google Scholar]
- 38.Rocca MA, et al. Adaptive functional changes in the cerebral cortex of patients with nondisabling multiple sclerosis correlate with the extent of brain structural damage. Ann Neurol. 2002;51:330–9. doi: 10.1002/ana.10120. [DOI] [PubMed] [Google Scholar]
- 39.Yetkin FZ, et al. FMRI of working memory in patients with mild cognitive impairment and probable Alzheimer’s disease. Eur Radiol. 2006;16:193–206. doi: 10.1007/s00330-005-2794-x. [DOI] [PubMed] [Google Scholar]
- 40.Karni A, et al. The acquisition of skilled motor performance: fast and slow experience-driven changes in primary motor cortex. Proc Natl Acad Sci U S A. 1998;95:861–8. doi: 10.1073/pnas.95.3.861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Calautti C, Baron JC. Functional neuroimaging studies of motor recovery after stroke in adults: a review. Stroke. 2003;34:1553–66. doi: 10.1161/01.STR.0000071761.36075.A6. [DOI] [PubMed] [Google Scholar]
- 42.Emmer BJ, et al. Selective involvement of the amygdala in systemic lupus erythematosus. PLoS Med. 2006;3:e499. doi: 10.1371/journal.pmed.0030499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Cavanna AE, Trimble MR. The precuneus: a review of its functional anatomy and behavioural correlates. Brain. 2006;129:564–83. doi: 10.1093/brain/awl004. [DOI] [PubMed] [Google Scholar]
- 44.Cabeza R, et al. Task-independent and task-specific age effects on brain activity during working memory, visual attention and episodic retrieval. Cereb Cortex. 2004;14:364–75. doi: 10.1093/cercor/bhg133. [DOI] [PubMed] [Google Scholar]
- 45.Rajah MN, D’Esposito M. Region-specific changes in prefrontal function with age: a review of PET and fMRI studies on working and episodic memory. Brain. 2005;128:1964–83. doi: 10.1093/brain/awh608. [DOI] [PubMed] [Google Scholar]
- 46.de Kloet ER, Oitzl MS, Joels M. Stress and cognition: are corticosteroids good or bad guys? Trends Neurosci. 1999;22:422–6. doi: 10.1016/s0166-2236(99)01438-1. [DOI] [PubMed] [Google Scholar]
- 47.de Quervain DJ, Roozendaal B, McGaugh JL. Stress and glucocorticoids impair retrieval of long-term spatial memory. Nature. 1998;394:787–90. doi: 10.1038/29542. [DOI] [PubMed] [Google Scholar]
- 48.Brown ES, et al. Hippocampal volume, spectroscopy, cognition, and mood in patients receiving corticosteroid therapy. Biol Psychiatry. 2004;55:538–45. doi: 10.1016/j.biopsych.2003.09.010. [DOI] [PubMed] [Google Scholar]
- 49.Keenan PA, et al. Commonly used therapeutic doses of glucocorticoids impair explicit memory. Ann N Y Acad Sci. 1995;761:400–2. doi: 10.1111/j.1749-6632.1995.tb31402.x. [DOI] [PubMed] [Google Scholar]
- 50.McEwen BS. Protective and damaging effects of stress mediators. N Engl J Med. 1998;338:171–9. doi: 10.1056/NEJM199801153380307. [DOI] [PubMed] [Google Scholar]
- 51.Sapolsky RM. Glucocorticoids and hippocampal atrophy in neuropsychiatric disorders. Arch Gen Psychiatry. 2000;57:925–35. doi: 10.1001/archpsyc.57.10.925. [DOI] [PubMed] [Google Scholar]
- 52.Young AH, et al. The effects of chronic administration of hydrocortisone on cognitive function in normal male volunteers. Psychopharmacology (Berl) 1999;145:260–6. doi: 10.1007/s002130051057. [DOI] [PubMed] [Google Scholar]
- 53.Appenzeller S, et al. Longitudinal analysis of gray and white matter loss in patients with systemic lupus erythematosus. Neuroimage. 2007;34:694–701. doi: 10.1016/j.neuroimage.2006.09.029. [DOI] [PubMed] [Google Scholar]
- 54.Holsboer F, Barden N. Antidepressants and hypothalamic-pituitary-adrenocortical regulation. Endocr Rev. 1996;17:187–205. doi: 10.1210/edrv-17-2-187. [DOI] [PubMed] [Google Scholar]
- 55.Lupien SJ, McEwen BS. The acute effects of corticosteroids on cognition: integration of animal and human model studies. Brain Res Brain Res Rev. 1997;24:1–27. doi: 10.1016/s0165-0173(97)00004-0. [DOI] [PubMed] [Google Scholar]
- 56.Ginsburg KS, et al. A controlled study of the prevalence of cognitive dysfunction in randomly selected patients with systemic lupus erythematosus. Arthritis Rheum. 1992;35:776–82. doi: 10.1002/art.1780350711. [DOI] [PubMed] [Google Scholar]
- 57.Hanly JG, et al. Cognitive impairment in patients with systemic lupus erythematosus. J Rheumatol. 1992;19:562–7. [PubMed] [Google Scholar]
- 58.Hay EM, et al. A prospective study of psychiatric disorder and cognitive function in systemic lupus erythematosus. Ann Rheum Dis. 1994;53:298–303. doi: 10.1136/ard.53.5.298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kozora E, et al. Analysis of cognitive and psychological deficits in systemic lupus erythematosus patients without overt central nervous system disease. Arthritis Rheum. 1996;39:2035–45. doi: 10.1002/art.1780391213. [DOI] [PubMed] [Google Scholar]
- 60.Papero PH, et al. Neuropsychologic deficits and antineuronal antibodies in pediatric systemic lupus erythematosus. Clin Exp Rheumatol. 1990;8:417–24. [PubMed] [Google Scholar]
- 61.Waterloo K, et al. Neuropsychological dysfunction in systemic lupus erythematosus is not associated with changes in cerebral blood flow. J Neurol. 2001;248:595–602. doi: 10.1007/s004150170138. [DOI] [PubMed] [Google Scholar]